ПАРАЗИТОЛОГИЯ, 2023, том 57, № 2, с. 91-107.
УДК 576.895
О ПРОИСХОЖДЕНИИ ИКСОДОВЫХ КЛЕЩЕЙ
(PARASITIFORMES, IXODIDAE)
© 2023 г. С. А. Леонович*
Зоологический институт РАН,
Университетская наб., 1, Санкт-Петербург, 199034 Россия
*e-mail: Segei.Leonovich@zin.ru
Поступила в редакцию 27.02.2023 г.
После доработки 11.03.2023 г.
Принята к публикации 12.03.2023 г.
В статье рассматриваются различные гипотезы происхождения иксодовых клещей, пере-
носчиков множества опасных трансмиссивных инфекционных заболеваний человека и жи-
вотных, с привлечением литературных и частично собственных данных. Согласно последним
молекулярно-генетическим и палеонтологическим данным, проиксодиды появились не в середи-
не мезозоя, как считалось ранее, но скорее в середине девонского периода палеозоя. Наиболее
близкой к иксодидам сестринской группой паразитиформных клещей являются клещи-голотиры
(отряд Holothyrida Thon, 1905).
Ключевые слова: Ixodidae, происхождение, датировка, палеонтологические данные, органы
чувств
DOI: 10.31857/S0031184723020011; EDN: AZSEGK
Иксодовые клещи - временные эктопаразиты позвоночных животных, облигатные
гематофаги, характеризующиеся длительным питанием на хозяине. Интерес к иксодо-
вым клещам определяется в основном тем, что они служат переносчиками множества
опасных трансмиссивных заболеваний человека и животных (Балашов, 2009). Проис-
хождение этой группы остается совершенно неясным, так как морфологически эти
клещи сильно отличаются от прочих хелицеровых, обладая некоторыми чертами сход-
ства с гамазовыми, аргасовыми клещами, клещами-сенокосцами и голотирами и об-
разуя надотряд паразитиформных клещей. Одна из причин неясности происхождения
иксодид - весьма редкие и отрывочные палеонтологические данные и отсутствие (до
91
последнего времени) результатов молекулярно-генетических исследований как самих
иксодовых клещей, так и возможных сестринских групп. Молекулярно-генетические
методы широко используются при решении вопросов филогенетических отношении
внутри самой группы, практически не касаясь ее происхождения. Поэтому различные
гипотезы о происхождении иксодовых клещей основывались главным образом на ре-
зультатах сравнительно-морфологического анализа. Проведение такого анализа затруд-
нено тем, что разные авторы выбирают различные морфологические структуры, а это
приводит к противоречивым выводам о сестринских группах. Так, иксодовых клещей
сближают с клещами-голотирами (Lehtinen, 1991; Murrel et al., 2005), нуталлиелидами
(Mans et al., 2011) и даже некоторыми группами мезостигматических клещей, в част-
ности с клещами-уроподами (Karg, Schorlemmer, 2008). Временной отрезок, в течение
которого появились иксодовые клещи, сильно различается у разных авторов, причем
различия составляют сотни миллионов лет, от позднего мела-кайнозоя (Филиппова,
1977), до конца палеозоя (Hoogstraal, 1978). Не существует и общих представлений
относительно первичных групп хозяев иксодовых клещей. По мнению Н.А. Филип-
повой (1977), первичными хозяевами иксодид были мезозойские млекопитающие. По
мнению Хугстрала (Hoogstraal, 1978), хозяевами проиксодид были «большие, гладкоко-
жие рептилии». Некоторые авторы в качестве исходных прокормителей рассматривают
амфибий (Jeyaprakash, Hoy, 2009), По мнению Ю.С. Балашова (1989), главную роль
в эволюции иксодид играли не только коэволюционные отношения иксодовых клещей
и позвоночных животных, но в не меньшей мере и приспособления к определенным
типам биотопов.
В данном обзоре автор поставил перед собой цель разобраться с современным
состоянием вопроса о происхождении клещей и высказать свои представления об
этой проблеме с привлечением литературных и частично собственных данных по
строению органов чувств.
По современным представлением, иксодовые клещи (семейство Ixodidae Dugès,
1834) представляют собой семейство, относящееся к надотряду паразитиформных
клещей, включающему в себя четыре основных отряда (Krantz, Walter, 2009; Beaulieu
et al., 2011):
Надотряд Parasitiformes Reuter, 1909 (sensu Krantz & Walter 2009) (4 отряда)
Отряд Opilioacarida Zakhvatkin, 1952 (клещи-сенокосцы)
Отряд Holothyrida Thon, 1905 (клещи-голотиры)
Отряд Ixodida Leach, 1815
92
Надсемейство Ixodoidea Dugès, 1834 (3 семейства) (иксодоидные клещи)
Семейство Argasidae Koch, 1844 (4 рода, 188 видов) (аргасовые клещи)
Семейство Ixodidae Dugès, 1834 (14 родов, 682 вида) (иксодовые клещи)
Семейство Nuttalliellidae Schulze, 1935 (клещи-нуталлиелиды, 1 вид -
Nuttalliella namaqua Bedford, 1931.
Отряд Mesostigmata G. Canestrini, 1891 (мезостигматические или гамазовые кле-
щи) (109 семейств, 11424 вида) [численные данные взяты из работы Beaulieu
et al. (2011)].
По мнению многих исследователей, которое разделяет и автор, акариформные
(надотряд Acariformes) и паразитиформные (надотряд Parasitiformes) клещи пред-
ставляют собой две независимые группы хелицеровых членистоногих. Некоторые
черты сходства объясняются конвергенцией признаков, развившихся в ходе процесса
миниатюризации. Генетические исследования (молекулярная калибровка) показали,
что минимальный возраст общего предка акариформных клещей относится к кем-
брию-ордовику (примерно 455-552 mya), в то время как происхождение надотря-
да паразитиформных клещей относится к более позднему периоду (карбон-пермь)
(Arribas et al., 2020).
Палеонтологические данные
Палеонтологические данные, касающиеся иксодовых клещей, весьма отрывочны и
немногочисленны. Объясняется это тем, что клещи сохранились в основном в янтарях
(балтийском и бирманском).
Всего к настоящему времени в янтарях были обнаружены следующие виды ик-
содовых клещей:
Ixodes succineus Weidner, 1964 (Ixodidae), Балтийский янтарь (Третичный период,
35-50 mya) (Weidner, 1964);
Ixodes spp. Балтийский янтарь (Третичный период, 35-50 mya) (De la Fuente,
2003);
Ixodes antiquorum Chitimia-Dobler, Mans et Dunlop, 2022 (Сеноман-Альб, 94-113
mya) (Chitimia-Dobler et al., 2022);
Ixodes tertiarius Scudder, 1885 (Третичный период, олигоцен, 30 mya) (De la Fuente,
2003)
Ixodes spp. (голоцен, 11000 лет назад) Guerra et al., 2001; Guerra, 2002 (ссылки из
Rocha, da Serra-Freire, 2014)
93
Hyalomma spp. Балтийский янтарь (Третичный период, 35-50 mya) (De la Fuente,
2003);
Cornupalpatum burmanicum Poinar et Brown, 2003 (Сеноман-Альб, 94-113 mya)
(Chitimia-Dobler et al., 2022);
Deinocroton copia Chitimia-Dobler, Mans, Handsuch et Dunlop, 2022), (Сеноман-
Альб, 94-113 mya) (Chitimia-Dobler et al., 2022);
Khimaira fossus Chitimia-Dobler, Mans et Dunlop, 2022 ) (Сеноман-Альб, 94-113
mya) (Chitimia-Dobler et al., 2022);
Amblyomma testudinis Lane & Poinar, 1986, самец (Доминиканский янтарь, Третич-
ный период, 35-50 mya) (Lane, Poinar, 1986);
Amblyomma spp., личинка (Доминиканский янтарь, Третичный период, 15-40 mya)
(Poinar, 1992);
Dermacentor reticulatus Fabricius, 1794 (Четвертичной период, плиоцен, 2-5 mya)
(De la Fuente, 2003);
Amblyomma spp. (голоцен, 11000 лет назад) Guerra et al., 2001; Guerra, 2002 (ссылки
из Rocha, da Serra-Freire, 2014).
Мы можем заключить из этого весьма неполного списка (к сожалению, иксодовые
клещи сохраняются только в янтаре), что эволюция иксодовых клещей идет очень
медленно (виды современного рода Ixodes известны примерно со 100 mya до 50 mya,
а представители рода (хотя и другие виды) известны и в наше время.
На основе палеонтологических данных, имевшихся к началу 21 века, большинство
исследователей считали, что иксодовые клещи появились примерно в середине мезо-
зоя. Так, по мнению де ла Фуенте (De la Fuente, 2003), основанному на ископаемом
материале, имевшемся в тот период, происхождение иксодовых клещей относится
к меловому периоду (65-146 mya), причем основные этапы эволюции распростра-
нения этих клещей приходятся на третичный период (палеоген, неоген и часть ан-
тропогена).
Однако полученные в последнее время палеонтологические данные категориче-
ски не согласуются с такими представлениями. Так, недавно в бирманском янтаре
были обнаружены представители современных родов иксодовых клещей (Chitimia-
Dobler et al., 2022), в частности, Ixodes antiquorum Chitimia-Dobler, Mans et Dunlop,
2022, старейший из ископаемых видов рода Ixodes (Prostriata), а также представители
описанных вымерших семейств Deinocrotonidae (Cornupalpatum burmanicum Poinar
et Brown, 2003 Deinocroton copia Chitimia-Dobler, Mans, Handsuch et Dunlop, 2022) и
94
вновь описанного вымершего семейства Khimairidae (Khimaira fossus), датируемых
как Сеноманский-Альбский ярусы на границе верхнего и нижнего мела (94-113 mya).
Т.е. в этот период уже существовали представители как Prostriata, так и Metastriata
(Chitimia-Dobler et al., 2022), включая представителя рецентного рода Ixodes. Ины-
ми словами, предполагаемый период происхождения иксодовых клещей совпадает
с существованием рецентного рода Ixodes, чего, конечно же, быть никак не может.
Молекулярно-генетические данные также заставляют относить происхождение
иксодовых клещей не к середине мезозоя, а к более раннему периоду. Так, на осно-
вании результатов исследования митохондриальной ДНК у представителей 25 так-
сонов хелицерат, было показано, что расхождение эволюционных стволов пауков,
скорпионов, клещей (mites, т.е. кроме иксодоидных) и иксодоидных клещей (ticks)
произошло в конце палеозоя, т.е. значительно ранее, чем считалось согласно ран-
ним палеонтологическим находкам (Jeyaprakash, Hoy, 2009). Это хорошо согласуется
с последними палеонтологическими находками.
Согласно представлениям группы авторов-палеонтологов (Chitimia-Dobler
et al., 2022), основанных на находках представителей родов Ixodes, Amblyomma и
Haemaphysalis (находки датируются ими как примерно 100 mya) в Бирманском янтаре,
возникновение группы, которая впоследствии разделилась на иксодовых и аргасовых
клещей, произошло в период с середины пермского периода палеозоя до раннего
юрского периода мезозоя (273 и 192 mya).
Таким образом, согласно последним палеонтологическим и молекулярным дан-
ным, происхождение предков иксодовых клещей наиболее вероятно относится не
к средине мезозоя, как считалось ранее, а скорее к середине пермского периода (при-
мерно 250 mya).
Отсюда следует неожиданный вывод - первыми хозяевами проиксодид никак не
могли быть млекопитающие, которых в данный период просто не существовало.
С точки зрения автора, проблема первичных хозяев проиксодид не так важна, как
это представлялось многим авторам. На мой взгляд, наиболее обоснованной являет-
ся экологическая гипотеза Ю.С. Балашова (1989, 1998) о слабых коэволюционных
паразито-хозяинных связях на ранних этапах формирования иксодовых клещей и
о приспособлении проиксодид не к конкретным хозяевам, а скорее к конкретным
биотопам. В этих битопах круг хозяев определяется теми животными, которые здесь
обитают. Тесные паразито-хозяинные связи иксодовых клещей с конкретными груп-
пами наземных позвоночных, видимо, формировались позднее. По мнению некоторых
95
авторов, первичными хозяевами проиксодид (а также аргасовых клещей (Argasidae) и
клещей-нуталлиелид (Nittalielidae), образующих общую кладу среди Parasitiformes),
могли быть девонские лабиринтодонтные амфибии (Mans et al., 2012).
Родственные группы
Отряд Mesostigmata G. Canestrini, 1891
В ранних работах (Baker, Wharton, 1952) иксодовых и мезостигматических клещей
рассматривали в качестве родственных групп (Рис. 1Ba); в настоящее время (Klopmen,
2010) эти взгляды признаются устаревшими и не соответствующими современным
генетическим и морфологическим данным.
Отряд Mesostigmata, подотряд Uropodina
Некоторые авторы выделяют среди мезостигматических клещей группу клещей-
уропод, рассматривая их в качестве группы, наиболее близкой к иксодовым клещам
(Karg, Schorlemmer, 2008).
Уроподы - мелкие клещи (менее 2 мм). Для большинства представителей харак-
терны редукция хетотаксии ног и щупалец, вытянутые хелицеры с относительно ма-
ленькими терминальными пальцами. Форма тела округлая или овальная, выпуклая.
Дорзальный щит цельный, твёрдый. Внешне похожи на микроскопических черепах
или божьих коровок. Обитают в почве, навозе, муравейниках, лесной подстилке. Пи-
тание разнообразно (свободноживущие хищники, детритофаги, фитофаги, паразиты).
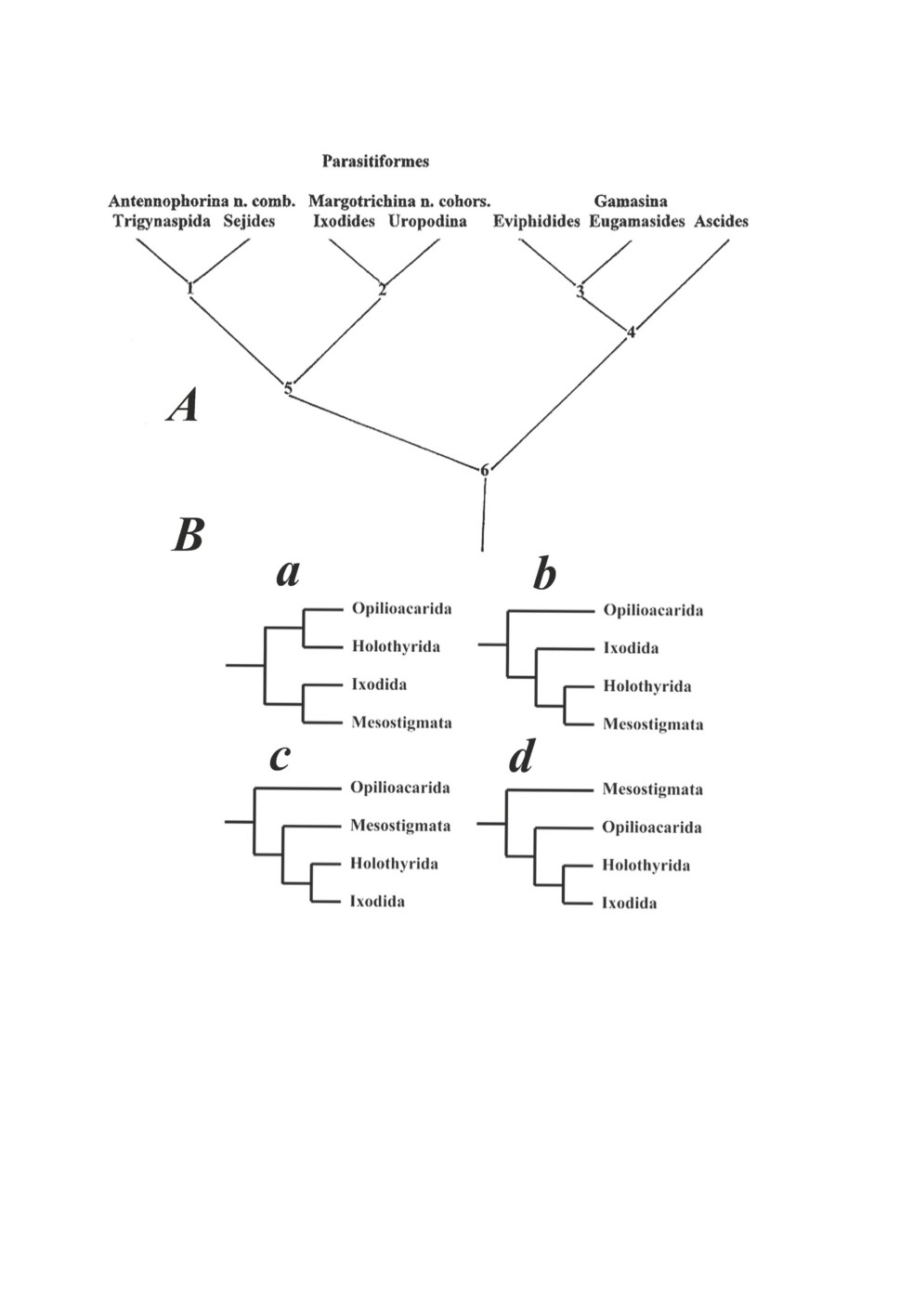
Карг и Шорлеммер (Karg, Schorlemmer, 2008) сближают иксодовых клещей
с клещами-уроподами (Uropodina) (Рис. 1A) на основании некоторых выделенных ими
синапоморфий, таких как расположение щетинок на пигидиальном щитке личинок
уропод и личинок иксодид (при этом пигидиальный щит у последних отсутствует,
а наличие этих щетинок трактуется как остaток щитка), наличие перигенитальных
бороздок, наличие анальных щетинок у всех фаз развития иксодид и у дейтонимф
уропод. На основании подобных морфологических признаков авторы разделяют
паразитиформных клещей на три основные группы: Antennophorina (включающей
Trigynopsida и Sejides), Margotrichina (включающей родственные группы Ixodida и
Uropodina) и отдельная группа остальных гамазовых клещей (Рис. 1А).
Однако родство уроподин и иксодовых клещей опровергается молекулярно-
генетическими исследованиями (Klompen et al., 2007). Исследования митогенома по-
казали, что мезостигматические (гамазовые) клещи и иксодовые клещи представля-
ют собой монофилетические группы, хотя и близкие, но не объединяемые в общую
96
группу (Xin-Chao Ban et al., 2022). Кроме того, выделяемые авторами морфологиче-
ские признаки не входят в перечень устойчивых структур, позволяющих сравнивать
ларвальную морфологию иксодовых клещей (Klompen et al., 1996). Мелкие размеры,
в значительной степени специализированный внешний облик и тип питания также
не позволяют считать уроподин сестринской группой иксодид.
Таким образом, мезостигматические (гамазовые) клещи, в том числе клещи-
уроподы, никак не могут быть группой, родственной проиксодидам.
Отряд Opilioacarida Zakhvatkin, 1952 (клещи-сенокосцы)
Небольшого размера (1-3 мм) клещи, головной и грудной отделы слиты в голо-
вогрудь, брюшко сохраняет внешнюю сегментацию. Имеются две или даже три пары
глаз. Ноги тонкие и длинные, придают клещам сходство с сенокосцами (Opiliones).
Питаются твердой пищей, хищники или сапрофаги. Старейшая палеонтологическая
находка Opilioacarida (Opilioacarus groehni Dunlop et Bernardi, 2014) датируется се-
номанским ярусом верхнего мела (Dunlop, Bernardi, 2014).
Клещи-голотиры (отряд Holothyridae, надотряд Parasitiformes)
Относительно крупные (до 7 мм) медлительные клещи, обитатели влажной лесной
подстилки. Исследования гена малой субъединицы рибосомальной РНК показали, что
Opilioacariformes являются сестринской группой по отношению к группе Ixodida-
Holothyrida, а не Ixodida-Mesostigmata-Holothyrida (Murrell et al., 2005).
Согласно работам Lehtinen (1991); d - Murrell et al. (2005), именно голотиры яв-
ляются наиболее близкой к иксодовым клещам группой паразитиформных клещей
(Риc. 1Bc, 1Cd)
Рассмотрим подробнее эти родственные группы с точки зрения их возможной
близости к предковой форме иксодид.
На взгляд автора, важную роль в анализе филогенетических связей может играть
строение сенсорных систем.
Нет никаких сомнений, что предки иксодовых клещей в течение длительного
времени обитали в почве, а еще ранее - на ее поверхности. Глаза иксодовых кле-
щей обладают уникальной ультраструктурной чертой, не встречающейся более ни
в одной из групп членистоногих (Eakin, 1979). Микроворсинки фоторецепторных кле-
ток, формирующие их рабдомеры, ориентированы не поперечно продольной оси, как
это типично для всех артропод, а параллельно этой оси, причем это характерно для
всех фаз развития изученных в этом отношении иксодовых клещей. Такое строение
выявлено и в фоторецепторных клетках, обнаруженных под гиподермой «безглазых»
видов клещей в тех местах, где находятся глаза у видов «глазастых» (Леонович, 2005).
97
Рисунок 1. Филогенетические отношения между различными группами клещей согласно:
А - Karg, Schorlemmer, 2008; В - другим авторам (a - Baker, Wharton, 1952,
b - Norton et al., 1993, c - Lehtinen, 1991, d - Murrell et al., 2005).
Схема приведена по: Klompen, 2010.
Figure 1. Phylogenetic relations between different groups of mites according to:
A - Karg, Schorlemmer, 2008; В - to other authors (a - Baker, Wharton, 1952;
b - Norton et al., 1993; c - Lehtinen, 1991; d - Murrell et al., 2005).
Scheme from Klompen, 2010.
98
Если бы глаза иксодид развились у них изначально, этот феномен невозможно
было бы объяснить. Единственное приемлемое объяснение заключается в том, что
у предков проиксодид глаза имелись, но потоми исчезли за десятки миллионов лет
обитания в почве и подстилке. Когда же проиксодиды вышли на поверхность, глаза
появились de novo. Таким образом, глаза иксодовых клещей не были унаследова-
ны от гипотетических предков, а возникли de novo, обладая идентичной функцией,
но принципиально отличаясь морфологически, в соответствии с принципом Долло
(Dollo, 1893). Современные же «безглазые» виды (роды Ixodes, Haemaphysalis) утрати-
ли глаза уже вторично. Точнее, не утратили, а сохранили их остатки в рудиментарном
виде (Леонович, 2005). Таким образом, как проиксодид, так и группы, родственные
проиксодидам, следует искать среди безглазых обитателей почвы или среди клещей,
структура глаз которых идентична таковой иксодовых клещей. Это сразу же исключает
Opilioacarina, обладающих двумя, а то и тремя парами глаз обычного для членисто-
ногих строения (Kaiser, Alberti, 1991). У мезостигматических (гамазовых) клещей и
клещей-голотир глаз нет.
Иксодовые клещи, точнее все иксодоидные клещи (Ixodoidea), обладают уникаль-
ным рецепторным образованием - органом Галлера, который не встречается больше
ни в одной группе клещей. Этот орган типичен как для иксодид, так и для аргазид,
причем, несмотря на все различия, и у тех и у других обладает общими чертами
строения. Орган Галлера - сложный полифункциональный орган чувств, расположен-
ный на тарзальных члениках передник конечностей. Главная часть органа Галлера -
капсула, более или менее глубокая ямка, в которой обнаруживаются пористые кутику-
лярные волоски обонятельных сенсилл (Леонович, 1977, 1978, 1980). Капсула может
быть открытой (у части представителей рода Ixodes (Leonovich, 2020) или закры-
той (все остальные роды Prostriata и все вида аргасовых клещей) (Leonovich, 2021).
Кроме ольфакторных сенсилл, на дне капсулы располагаются несенсорные выросты
сложной формы, так называемые плеоморфы (Dermacentor, Rhipicephalus, Hyalomma)
(Леонович, 2005). У представителей подсемейства Ixodinae (род Ixodes) плеоморфы
отсутствуют, у аргасовых клещей могут формировать крышку капсулы (Леонович,
1980). Перед капсулой (иногда также в ямке) располагается передняя группа сенсилл,
содержащая термо- и гигрорецепторые сенсиллы (Леонович, 2005).
Основной морфологической чертой строения органа Галлера является группа обо-
нятельных сенсилл, погруженная в ямку на дорсальной стороне переднего тарзуса.
Такая ямка, открытая или закрытая крышкой капсулы, имеется у всех без исключения
иксодоидных клещей, включая иксодовых (Ixodidae).
99
Рассматривая строение рецепторов на тарзальных члениках передних ног
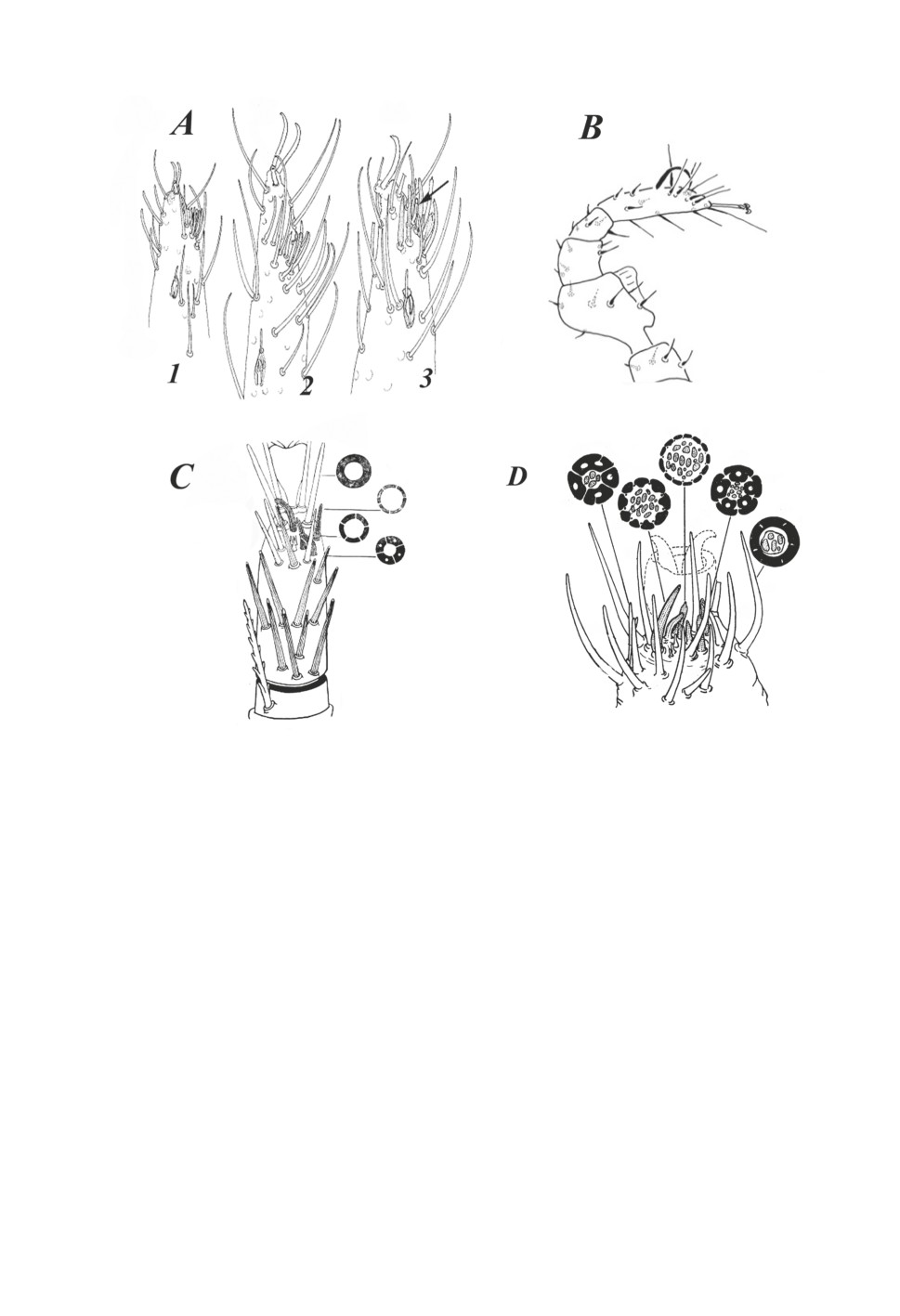
у Ixodidae, Gamasina, Uropodina, Opilioacarina и Holothyrida, можно заметить, что
у представителей Gamasina, Uropodina и Opilioacarina (Рис. 2) тарзальный рецеп-
торный комплекс представляет собой скопление сенсилл на дорсальной поверхности
переднего тарзуса, причем обращает на себя внимание явное сходство этого образова-
ния у опилиоакаран (Рис. 2А), уроподин (Рис. 2B) и гамазовых клещей (Mesostigmata)
(Рис. 2С, 2D). К сожалению, если рецепторные комплексы гамазовых клещей ис-
следованы методами электронной микроскопии, позволяющими четко выявлять мо-
дальность (Леонович, 1984, 1989), то опилиоакарины и уроподы в этом отношении
не исследованы. Однако даже по данным световой микроскопии видно, что, как
у гамазид, так и у уропод и клещей-сенокосцев, обонятельные сенсиллы (на Рис. 2С
и 2D волоски таких сенсилл покрыты точками, обозначающими поры - характер-
нейшую черту обонятельных сенсилл) не собраны в общую компактную группу и
не погружены в ямку.
В то же время, именно это явление мы наблюдаем в комплексах тарзальных
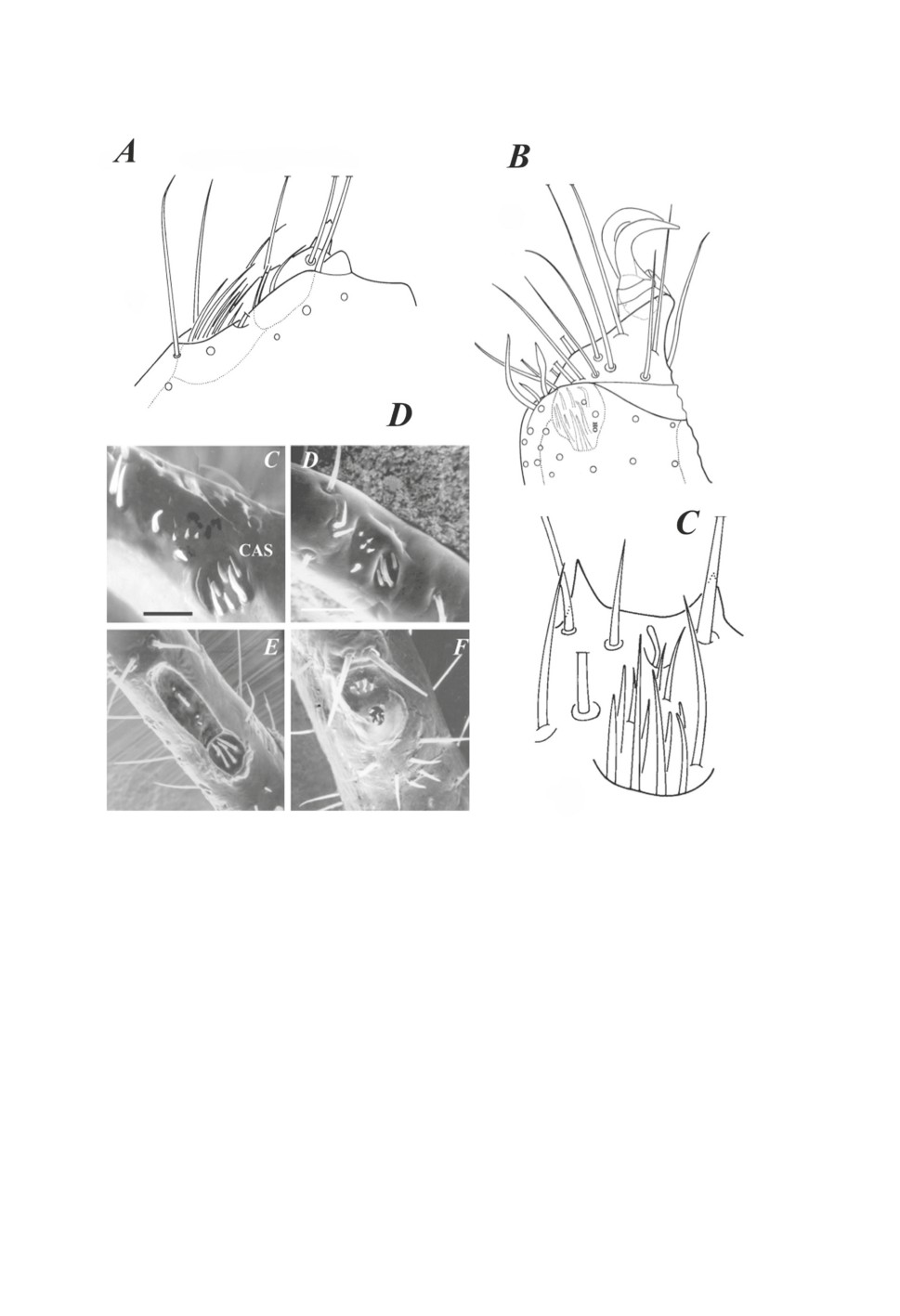
сенсилл клещей-голотир (Рис. 3A-3C). Обратим внимание на очевидное сходство
«Галлеро-подобных» органов клещей-голотир разных видов (Hammenius (Leiothyrus)
holthuisi Hammen, 1983 (Рис. 3A), Australothyrus ocellatus Hammen, 1983 (Рис. 3B) и
Holothyrus coccinella Gervais, 1842 (Рис. 3С) с органами Галлера некоторых видов
иксодовых клещей, обладающих открытой капсулой (Рис. 3D). Отметим, что никакого
функционального значения такое «погружение» сенсилл в ямку у клещей-голотир
явно не имеет. У иксодовых клещей форма ямки, форма крышки (если имеется) и
ряд других структур никак не связаны ни с местообитанием (нидиколия или обитание
на пастбище), ни с увлажненностью биотопа, ни с другими эколого-этологическими
факторами, но отражает исключительно филогенетические отношения (Leonovich,
2020, 2021).
Таким образом, и строение сенсорной системы указывает на клещей-голотир как
на наиболее близкую к иксодовым клещам группу.
Вызывает интерес и тип питания проиксодид. Очевидно, что питание твердой
пищей и питание жидкостями приводит к развитию совершенно разных морфоло-
гических приспособлений ротовых органов. Перестройка клешнеобразных хелицер,
характерных для клещей, питающихся твердой пищей, в сосущий ротовой аппарат
представляется крайне маловероятной. Поэтому наиболее вероятно проиксодиды
должны были быть преадаптированы к кровососанию, т.е. к высасыванию жидкого
100
Рисунок 2. Тарзальный рецепторный орган некоторых видов паразитиформных клещей:
A - Opilioacaridae, B - Uropodida, С-D - Gamasina.
A - тарзальный членик передней лапки Caribeacarus armasi Vázquez & Klompen, 2009
(1 - протонимфа, 2 - самка) и самки Neocarus siankaanensis Vazquez & Klompen, 2002 (3)
(по: Vázquez, Klompen, 2009).
B - тарзальный членик Urobovella sugiyamai Hiramatsu, 1979 (по: Hiromatsu, 1983).
C - тарзальный рецепторный комплекс клеща Haemogamasus ambulans Thorell, 1872
(Mesostigmata) (по: Леонович, 1984) .
D - тарзальный рецепторный комплекс клеща Hirstionyssus criceti Sulzer, 1774
(Mesostigmata) по данным электронной микроскопии, показывающим тонкое внутреннее
строениe всех типов сенсилл (по: Леонович, 1985).
Figure 2. Tarsal receptor complex in some species of mites:
A - Opilioacaridae, B - Uropodida, С-D - Gamasina.
A - foreleg tarsal segment of Caribeacarus armasi Vázquez & Klompen, 2009:
(1 - protonymph, 2 - female); female of Neocarus siankaanensis Vazquez & Klompen, 2002 (3)
(from: Vázquez, Klompen, 2009).
B - tarsal segment of Urobovella sugiyamai Hiramatsu, 1979 (from: Hiromatsu, 1983).
C - tarsal receptor complex of Haemogamasus ambulans Thorell, 1872 (Mesostigmata)
(from: Leonovich, 1984).
D - tarsal receptor complex of Hirstionyssus criceti Sulzer, 1774 (Mesostigmata)
(from: Leonovich, 1989).
101
Рисунок 3. Тарзальные рецепторы («орган Галлера») клещей-голотир (Holothyrida) (А-С)
и настоящий орган Галлера иксодовых клещей (Ixodidae, Ixodinae) (D).
A - Hammenius (Leiothyrus) holthuisi Hammen, 1983 (правая нога самца) (по: van der Hammen,
1983). В - Australothyrus ocellatus Hammen, 1983 (по: van der Hammen, 1983).
С - Holothyrus coccinella Gervais, 1842 (по: van der Hammen, 1983). D - орган Галлера
некоторых видов рода Ixodes (по: Leonovich, 2020); С - I. uriae, самка; D - I. uriae, личинка;
E - I. vespertilionis, самка; F - I. ovatus, самка. Масштабная линейка, мкм: A, B - 30;
C, D - 30; E, F - 50.
Figure 3. Tarsal receptors («Haller’r organ») in holothyrid mites (Holothyrida) (А-С) and the real
Haller’s organ of some ixodid ticks (Ixodidae, Ixodinae) (D).
A - Hammenius (Leiothyrus) holthuisi Hammen, 1983 (male right leg) (from van der Hammen,
1983). В - Australothyrus ocellatus Hammen, 1983 (from van der Hammen, 1983). С -
Holothyrus coccinella Gervais, 1842 (from van der Hammen, 1983). D - Haller’s organ in some
species of the genus Ixodes (from Leonovich, 2020) ; С - I. uriae, female; D - I. uriae, larva;
E - I. vespertilionis, female; F - I. ovatus, female. Scale (µm): A, B - 30; C, D - 30; E, F - 50.
102
содержимого и при этом не должны были испытывать сопротивления жестких по-
кровов. Те, кто к этому приспособился, не испытывают необходимости в изменении
своих адаптаций.
Исследование пищевого поведения в лабораторных условиях у 3 видов клещей-
голотир рода Allothyrus (Parasitiformes: Holothyrida: Allothyridae) (Walter, Proctor, 1998)
показало, что клещи-голотиры - это падальщики, способные поглощать (высасывать)
только жидкую пищу. Они пренебрегали живыми членистоногими, улитками (моллю-
сками), нематодами и аннелидами, но мертвые членистоногие служили прекрасной пи-
щей и прокармливали нимф и взрослых клещей в течение нескольких месяцев. Эти же
авторы изучали пищевое поведение ряда древних Mesostigmata (Sejina, представители
родов Sejus и Uropodella; Uropodina, Polyaspis sp. и Cercomegistina). Оказалось, что все
эти клещи были агрессивными хищниками, высасывавшими мелких беспозвоночных и
игнорировавшими мертвых животных. В кишечнике ряда видов Asternolaelaps (Sejina)
были обнаружены твердые частицы грибов и мелких членистоногих. Очень похожая
ситуация наблюдалась у клещей-сенокосцев (неопределенные до вида Opilioacarida из
Австралии). Все эти клещи поглощали исключительно твердую пищу. Таким образом,
согласно представлением авторов, питание исключительно жидкой пищей являет-
ся общей (объединяющей) особенностью для трех групп паразитиформных клещей
(Holothyrida, Ixodida и части Mesostigmata) (Walter, Proctor, 1998).
Таким образом, и тип питания указывает на клещей-голотир как на наиболее близ-
кую к проиксодидам группу паразитиформных клещей.
Как же могли выглядеть клещи-проиксодиды? Можно выдвинуть гипотезу, что
это были относительно крупные (5-7 мм) почвенные клещи-сапрофаги, которые вы-
сасывали гемолимфу мертвых членистоногих и несколько напоминали рецентных
Holothyrida. Хищничество требует высокой подвижности, в то время как малопод-
вижные клещи-сапрофаги имели больше шансов перейти к прокалыванию покровов
крупных и также малоподвижных палеозойских амфибий. Становление паразитизма,
скорее всего, произошло еще на стадии общих предков, позднее разделившихся на три
родственных ствола (Ixodidae, Argasidae, Nuttalielidae). На ранних этапах становления
группы основную роль играло приспособление к обитанию в определенной среде
(влажные жаркие биотопы, верхний слой почвы и подстилка), а не приспособление
к определенным хозяевам, что, видимо, способствовало становлению полифагии про-
иксодид. Коэволюционные паразито-хозяинные связи развились значительно позднее,
в середине мезозоя и в кайнозое.
103
БЛАГОДАРНОСТИ
Работа выполнена по Государственному заданию «Разнообразие паразитар-
ных систем, адаптаций и путей эволюции паразитов» (номер темы: АААА-А19-
119020690109-2).
СПИСОК ЛИТЕРАТУРЫ
Балашов Ю.С. 1989. Коэволюция иксодовых клещей и наземных позвоночных. Паразитология 23 (6):
457-468. [Balashov Yu.S. 1989. Coevolution of ixodid ticks and terrestrial vertebrates. Parazitologiya 23
(6): 457-468. (in Russian)].
Балашов Ю.С. 1998. Иксодовые клещи - паразиты и переносчики инфекций. СПб., Наука, 287 с. [Balashov
Yu.S. 1998. Ixodid ticks - parasites and vectors of infections. St. Petersburg, Nauka, 287 pp. (in Russian)].
Балашов Ю.С. 2009 . Паразитизм клещей и насекомых на наземных позвоночных. СПб., Наука, 358 с.
[Balashov Yu.S. 2007. Parasitism of acarines and insects on terrestrial vertebrates. SPb., Nauka, 358 pp.
(in Russian)].
Леонович C.А. 1977. Электронно-микроскопическое исследование органа Галлера клеща Ixodes persulcatus
(Ixodidae). Паразитология 11 (4): 340-347. [Leonovich S.A. 1977. Electron microscopy studies of Haller's
organ of the tick Ixodes persulcatus (Ixodidae). Parazitologiya 11 (4): 340-347. (in Russian)].
Леонович С.А. 1978. Тонкое строение органа Галлера иксодового клеща Hyalomma asiaticum P. Sch. et
E. Sch. (Parasitiformes, Ixodidae, Amblyomminae). Энтомологическое обозрение 57 (1): 221-226.
[Leonovich S.A. 1978. The fine structure of the Haller’s organ in the ixodid tick Hyalomma asiaticum P.
Sch. et E. Sch. (Parasitiformes, Ixodidae, Amblyomminae). Entomologicheskoe obozrenie 57 (1): 221-226.
(in Russian)].
Леонович С.А. 1980. Ультраструктурные исследования органа Галлера аргасовых клещей Ornithodoros
moubata и Alveonasus lahorensis (Argasidae). Паразитология 14 (5): 376-385. [Leonovich S.A. 1980.
Fine structural investigation of Haller' s organ in argasid ticks Ornithodoros moubata and Alveonasus
lahorensis (Argasidae). Parazitologiya 14 (5): 376-385. (in Russian)].
Леонович С.А. 1984. Тарзальные рецепторные комплексы гамазовых клещей семейства Haemogamasidae.
Паразитология 18 (6): 451-458. [Leonovich S.A. 1984. Tarsal receptor complexes of gamasid mites of
the family Haemogamasidae. Parazitologiya 18 (6): 451-458. (in Russian)].
Леонович С.А. 1985. Ультраструктурное исследование тарзального рецепторного комплекса гамазового
клеща Hirstionyssus criceti (Hirstionyssidae). Паразитология 19 (6): 456-463. [Leonovich S.A. 1985. Fine
structural investigation of tarsal receptor complex in gamasid mite Hirstionyssus criceti (Hirstionyssidae).
Parazitologiya 19 (6): 456-463. (in Russian)].
Леонович С.А. 1989. Тарзальный рецепторный комплекс и систематика гамазовых клещей (Parasitiformes,
Mesostigmata, Gamasina). Паразитология 23 (6): 469-479. [Leonovich S.A. 1989. Tarsal receptory
complex and systematics of gamasid mites (Parasitiformes, Mesostigmata, Gamasina). Parazitologiya 23
(6): 469-479. (in Russian)].
Леонович С.А. 2005. Сенсорные системы паразитических клещей. СПб.: Наука, 236 с [Leonovich S.A.
Sensory systems of parasitic ticks and mites. SPb, Nauka, 236 p. (in Russian)].
104
Филиппова Н.А. 1977. Иксодовые клещи подсем. Ixodinae Фауна СССР. Новая серия, № 114. Паукообраз-
ные. Т. 4. Вып. 4. Ленинград, Наука. [Filippova N.A. 1977. Ixodovye kleshchi podesemeistva Ixodinae.
Fauna SSSR, Novaya seriya, 114, Paukoobraznye, Vol. 3, № 4, Leningrad, Nauka. (In Russian)].
Arribas P., Andújar C., Moraza M.L., Linard B., Emerson B.C., Vogler A.P. 2020. Mitochondrial metagenomics
reveals the ancient origin and phylodiversity of soil mites and provides a phylogeny of the Acari. Molecular
Baker E.W., Wharton G.W. 1952. An introduction to Acarology. New York, Macmillan Company, 465 pp.
Beaulieu F., Dowling A.P.G., Klompen H., de Moraes G.J., Evans D.
2011. Superorder Parasitiformes Reuter,
1909. In: Zhang Z.-Q. (Ed.). Animal biodiversity: An outline of higher-level classification and survey of
Chitimia-Dobler L., Mans B.J., Handschuh S., Dunlop J.A. 2022. A remarkable assemblage of ticks from mid-Cre-
De la Fuente J. 2003. The fossil record and the origin of ticks (Acari: Parasitiformes: Ixodida). Experimental &
Applied Acarology 30: 331-344.
Dunlop J.A., Bernardi L.F.O. An opilioacarid mite in Cretaceous Burmese amber. The Science of Nature 101 (9):
759-763. DOI: 10.1007/s00114-014-1212-0
Eakin R.M. 1979. Evolutionary significance of photoreceptors in retrospect. American Zoologist 19: 647-653.
Hiramatsu N. 1983. Mannchen von drei bekannten Arten und eine neue Discourella-Art von Uropodidae aus
Japan. Acta arachnologica 32: 15-26.
Hoogstraal H. 1978. Biology of ticks. In: Tick borne diseases and their vectors. Ed. J.K.H Wilde. Proc. Internat.
Conf., Edimburgh, September-October, University of Edinburgh, Centre for tropical veterinary medicine,
Edinburgh, 3-14.
Jeyaprakash A., Hoy M.J. 2009. First divergence time estimate of spiders, scorpions, mites and ticks (subphylum:
Chelicerata) inferred from mitochondrial phylogeny. Experimental and Applied Acarology 17: 1-18.
Kaiser T., Alberti G. 1991. The fine structure of the lateral eyes of Neocarus texanus Chamberlin and Mulaik,
1942 (Opilioacarida, Acari, Arachnida, Chelicerata). Protoplasma 163: 19-33.
Karg W., Schorlemmer A. 2008. Origin and classification of the Ixodides (ticks) within the Parasitiformes Reuter
1909 (Acarina). Acarologia 48 (3-4): 123-134.
Klompen H. 2010. Holothyrids and ticks: new insights from larval morphology and DNA sequencing, with the
description of a new species of Diplothyrus (Parasitiformes: Neothyridae). Acarologia 50 (2): 269-285.
DOI: 10.1051/acarologia/20101970
Klompen H., Lekveishvili M., Black IV W.C. 2007. Phylogeny of parasitiform mites (Acari) based on rRNA.
Klompen J.S.H., Keirans J.E., Filippova N.A., Oliver Jr. J.H. 1996. Idiosomal lyrifissures, setae, and small glands
as taxonomic characters and potential indicators of ancestral segmentation patterns in larval Ixodidae (Acari:
Krantz G.W., Walter D.E. (Eds) 2009. A Manual of Acarology. 3rd Edition. Texas Tech University Press, Lubbock,
807 pp.
105
Lane R.S., Poinar Jr. G. 1986. First fossil tick (Acari: Ixodidae) in New World amber. International Journal of
Acarology 12: 75-78.
Lehtinen P.T. 1991. Phylogeny and zoogeography of the Holothyrida. In: Dusbábek F., Bukva V. (Eds). Modern
Acarology, Vol. II. The Hague, SPB Academic Press, 101-113.
Leonovich S.A. 2020. Structure of Haller’s Organ and Taxonomy of Hard Ticks of the Subfamily Ixodinae (Family
Ixodidae). Entomological Review 100 (9): 1387-1401. DOI: 10.1134/S0013873820090122
Leonovich S.A. 2021. Structure of Haller’s Organ and Taxonomy of Hard Ticks of the Subfamily Amblyomminae
(Family Ixodidae). Entomological Review 101 (5): 709-724. DOI: 10.1134/S0013873821050110
Mans B., de Klerk D.D., Pienaar R., de Castro M.D., Latif A. 2012. The mitochondrial genomes of Nutalliella
namaqua (Ixodoidea: Nuttallielidae) and Argas africolumbae (Ixodoidea: Argasidae): estimation of
divergence dates for the major tick lineages and reconstruction of ancestral blood-feeding characters.
PLoS ONE DOI: 1 0.1371/journal.pone.0049461
Mans B.J., de Klerk D., Pienaar R., Latif A.A. 2011. Nuttalliella namaqua: A living fossil and closest relative
to the ancestral tick lineage: implications for the evolution of blood-feeding in ticks. PLoS ONE 6 (8):
Murrell A., Dobson S.J., Walter D.E., Campbell N.J.H., Shao R., Barker S.C. 2005. Relationships among the
three major lineages of the Acari (Arthropoda: Arachnida) inferred from small subunit rRNA: paraphyly
of the Parasitiformes with respect to the Opilioacariformes and relative rates of nucleotide substitution.
Invertebrate Systematics 19: 383-389.
Norton R.A., Kethley J.B., Johnston D.E., O,Connor B.M. 1993. Phylogenetic perspectives on genetic systems
and reproductive modes of mites. In: Wrensch D.L., Ebbert M.A. (Eds). Evolution and diversity of sex
ratio in insects and mites. New York, Chapman & Hall, 8-99.
Poinar Jr., G.O. 1992. Life in amber. Palo Alto: Stanford University Press.
Rocha G.C. da, da Serra-Freire N.M. 2014. Mites, Ticks, and Paleoparasitology. In Ferreira L.F., Reinhard K.J.,
Araújo A. (Editors). Foundations of Paleoparasitology [online]. Rio de Janeiro, Editora FIOCRUZ, 462
pp. doi: 10.7476/9788575415986
Van der Hammen L. 1983. New notes on Holothyrida (Anactinotrichid mites). Zoologische Verhandelingen 207
(1): 1- 48.
Vázquez M.M., Klompen H. 2009. New species of New World Opilioacaridae (Acari: Parasitiformes)
with the description of a new genus from the Caribbean region. Zootaxa 2061 (1): 3. DOI:
Walter D.E., Proctor H.C. 1998 Feeding behaviour and phylogeny: observations on early derivative. Acari.
Experimental & Applied Acarology 22: 39-50.
Weidner H. 1964. Eine Zecke, Ixodes succineus sp. n., im Baltischen Bernstein. Veroff Uberseemuseum, Bremen
3: 143-151.
Xin-Chao Ban, Zi-Kai Shao, Li-Jun Wu, Jing-Tao Sun, Xiao-Feng Xue 2022. Highly diversified mitochondrial
genomes provide new evidence for interordinal relationships in the Arachnida. Cladistics 38 (4): 452-464.
106
ON THE ORIGIN OF IXODID TICKS
(PARASITIFORMES, IXODIDAE)
S. A. Leonovich
Keywords: Ixodidae, origin, dating, paleontological data, sense organs
SUMMARY
Different hypotheses on the origin of ixodid ticks, vectors of numerous dangerous transmissive
diseases of humans and animals are analyzed basing on literary and partly own data. According
to latest molecular-genetic and paleontological data, pro-ixodid ticks appeared not in mid-Mesosoic,
as it was believed earlier, but most likely in the middle of Devonian. Among Parasitiformes, mites
of the order Holothyrida Thon, 1905 form the most closely related to Ixodidae sister group.
107