

Лесоведение, 2019, № 5, стр. 357-365
Влияние массового размножения сосновой пяденицы на радиальный прирост деревьев
В. Г. Суховольский a, b, *, П. А. Красноперова b, Е. Н. Пальникова c, И. В. Свидерская b, О. В. Тарасова b
a Институт леса им. В.Н. Сукачева СО РАН
660036 Красноярск, Академгородок, 50/28, Россия
b Сибирский федеральный университет
660041 Красноярск, пр. Свободный, 79, Россия
c Сибирский государственный университет науки и технологий имени, академика М.Ф. Решетнёва
660037 Красноярск, проспект имени газеты Красноярский рабочий, 31, Россия
* E-mail: soukhovolsky@yandex.ru
Поступила в редакцию 16.11.2017
После доработки 01.08.2018
Принята к публикации 05.06.2019
Аннотация
Изучены временные ряды радиального прироста сосны Pinus sylvestris L. с 1945 по 2015 г. в насаждениях, поврежденных в ходе вспышки сосновой пяденицы Bupalus piniarius L. в 1974–1978 г., а также в насаждениях, не поврежденных вредителем, на территории Краснотуранского соснового бора. Для оценки различий процессов радиального прироста деревьев в поврежденных и неповрежденных насаждениях до начала вспышки массового размножения предложено использовать регуляторные характеристики временны́х рядов, такие как величины n запаздывания (авторегрессии) и размаха s колебаний временны́х рядов. По этим показателями можно численно оценить инерционность процессов регуляции радиального прироста. Так как характеристики регуляции корректно вычислять только для стационарных временны́х рядов, для расчетов были использованы ряды первых разностей радиального прироста. Деревья в насаждениях, на которые напали насекомые, характеризовались либо отсутствием запаздывания во временны́х рядах, либо достаточно большими величинами порядка n авторегрессии. При этом колебания текущего радиального прироста относительно возрастного тренда у деревьев в поврежденных насаждениях были существенно больше, чем в неповрежденных. Дана оценка скорости восстановления прироста у поврежденных насекомыми деревьев в зависимости от регуляторных характеристик временны́х рядов радиального прироста до начала повреждений. Полученные характеристики можно использовать для оценки риска нападения филлофагов на насаждения и скорости восстановления насаждений, поврежденных насекомыми.
При изучении вспышек массового размножения насекомых важно понять, с чем связан выбор насекомыми насаждений, в которых возникают их очаги. Феномен вспышек массового размножения лесных насекомых объясняют, прежде всего, взаимодействием популяции с паразитами и хищниками и погодными условиями, привязанными к определенному моменту времени. Кроме того, для целого ряда видов лесных насекомых показано, что первичные очаги вспышек массового размножения характеризуются специфическими ландшафтными условиями (Исаев, Ряполов, 1979; Тарасова, 1982; Артемьева и др., 1995). Еще один возможный фактор, влияющий на пространственное размещение очага, – состояние кормовых растений и их привлекательность для вредителя (Thomas, 1989; Herms, Mattson, 1992; Тарасова и др., 2015; Isaev et al., 2017). Вспышка массового размножения будет начинаться в насаждении, привлекательном для насекомых, при определенных погодных условиях.
Для оценки устойчивости деревьев к нападению насекомых предложено большое число инструментальных показателей (Скрипальщикова и др., 2009; Суховольский и др., 2009; Мозолевская и др., 2011; Hard, 1985; Strauss, Agrawal, 1999; Haukioja et al., 2002; Larsson, 2002; Huberty, Denno, 2004; Haukioja, 2005; Mumm, Hilker, 2006; Stevens et al., 2007; Wise, Abrahamson, 2007; Barton, Koricheva, 2010; Delvas et al., 2011; Fuentealba, Bauce, 2012, Kumbas, Bauce, 2013; Castagneyrol et al., 2014; Ferrenberg et al., 2014; Buchel et al., 2016).
Однако при изучении вспышек массового размножения лесных насекомых-филлофагов исследователи обычно попадают в очаги, когда они уже сформировались, и сложно оценить физиологическое состояние деревьев в этих насаждениях до вспышки. Не вполне понятно, являются ли наблюдаемые отклонения в физиологическом состоянии деревьев причиной или же следствием воздействия насекомых.
Однако существует показатель, с помощью которого можно проводить ретроспективные исследования привлекательности деревьев для насекомых-филлофагов. Речь идет о радиальном приросте деревьев, характеризующем их ростовые процессы перед вспышкой, в ходе и после ее окончания.
Большинство работ по изучению связи радиального прироста и воздействия насекомых-филлофагов посвящено изменению радиального прироста после повреждения листового аппарата вредителями (Straw, 1996; Armour et al., 2003; Свидерская, Пальникова, 2003; Пальникова и др., 2016). Известно, что у деревьев с поврежденным фотосинтетическим аппаратом радиальный рост практически прекращается, но затем может восстановиться. Характер процесса восстановления зависит от уровня изъятия хвои (Пальникова и др., 2002).
В меньшей степени изучена связь между привлекательностью деревьев для насекомых и радиальным приростом в период перед вспышкой массового размножения.
В настоящей работе рассмотрены характеристики радиального прироста деревьев перед вспышкой массового размножения и изучена связь показателей ростовых процессов у деревьев до повреждения их насекомыми с ходом восстановления радиального роста после ее окончания.
В качестве рабочей гипотезы будет использовано представление о связи привлекательности деревьев для насекомых-филлофагов и процессов восстановления роста после повреждений с характеристиками регуляции ростовых процессов. Из теории автоматического регулирования известно, что при эффективной работе системы регуляции изучаемого объекта отклонение значений регулируемой переменной от нормы мало́, а после снятия внешнего воздействия такая система быстро возвращается к исходному состоянию (Поляк, Щербаков, 2002; Дорф, Бишоп, 2004; Ким, 2007; Веремей, 2013). При неустойчивости системы регуляции происходят сильные отклонения состояния системы от нормы в сочетании с достаточно большим временем запаздывания реакции на внешние воздействия. Регуляторные свойства, связанные с радиальным приростом деревьев, могут служить индикатором устойчивости деревьев к внешним воздействиям. Регуляторные характеристики системы можно оценивать, изучая свойства временны́х рядов регулируемой переменной методами корреляционного и спектрального анализа (Бокс, Дженкинс, 1974). При этом в качестве показателей регуляции можно использовать величины отклонения характеристик регулируемой системы от нормы и скорость реакции, которую можно найти по величине запаздывания ее характеристик (Веремей, 2013).
В рамках проведенного исследования изучены характеристики инерционности регуляторных процессов радиального прироста деревьев в очагах массового размножения сосновой пяденицы до начала вспышки массового размножения и в неповрежденных насаждениях.
ОБЪЕКТЫ И МЕТОДИКА
Сосновая пяденица (Bupalus piniarius L., Geometridae, Lepidoptera) имеет обширный ареал, совпадающий с распространением Pinus silvestris L. Вид имеет следующие особенности жизненного цикла: питание хвоей сосны на стадии гусеницы, отсутствие питания у имаго, окукливание и зимовка в подстилке в конце августа–сентябре, отсутствие личиночной диапаузы, кучная откладка яиц самками, ограниченная миграция гусениц, разнообразный комплекс энтомофагов, среди которых преобладают паразиты куколок. Основными факторами модификации численности сосновой пяденицы являются погодные условия в период питания гусениц. Вспышки массового размножения сосновой пяденицы – обычные явления в сосновых борах юга Сибири (Пальникова и др., 2002).
Для оценки зависимости между характеристиками регуляции радиального прироста стволов деревьев и привлекательностью деревьев для насекомых-филлофагов изучались временные ряды радиального прироста сосен Pinus sylvestris L. в очагах массового размножения сосновой пяденицы в 1974–1978 гг. на территории Краснотуранского соснового бора (54°16.315′ с.ш., 91°37.757′ в.д.) и деревьев на неповрежденных вредителем участках бора (Пальникова и др., 2002).
Для анализа на территории Краснотуранского бора для анализа было выбрано шесть пробных площадей (пр.пл.): пр. пл. 1 и 2 – очаги вспышки массового размножения 1974–1978 гг., деревья на которых были детально изучены, начиная с 1978 г. (Пальникова и др., 2002; Исаев и др., 2015), пр. пл. 3–6 расположены на территории, где деревья в 1974–1978 гг. не подвергались воздействию насекомых-вредителей.
Возраст деревьев к началу вспышки массового размножения как в очагах, так и вне их составлял 60–80 лет. Исследуемые деревья в очагах в процессе развития вспышки были дефолиированы на 30–50%, но впоследствии хвоя восстановилась. Для анализа радиального прироста на всех пробных площадях выбраны деревья с диаметром на высоте груди от 24 до 32 см. При этом на всех пробных площадях анализировались ряды радиального прироста деревьев до начала вспышки массового размножения сосновой пяденицы в период с 1945 по 1972 г., когда никаких повреждений хвои насекомыми еще не было, и ряды радиального прироста в период с 1972 по 2015 г.
С помощью приростного бурава были взяты керны из 44 деревьев в очагах массового размножения сосновой пяденицы и 44 образца в неповрежденных насаждениях. Ширина годичных колец измерялась на шлифованной поверхности кернов в поле зрения микроскопа на установке “Lintab 5 Tree-RingStation” (RINNTECH®) с точностью 0.1 мм. Датировка годичных колец выполнялась визуально по графикам в программе TSAP-WinTM (RINNTECH®), контроль датировки производился с помощью программы COFECHA (version 6.0P), содержащейся в библиотеке дендрохронологических программ DPL (https://www.ldeo.columbia.edu/tree-ring-laboratory/resources/software).
Методы спектрального и корреляционного анализа временны́х рядов корректно использовать, если изучаемые временные ряды стационарны (Бокс, Дженкинс, 1974). Однако для рядов радиального прироста деревьев $\{ x(j)\} $ характерно наличие возрастного тренда. В связи с этим прежде чем использовать методы корреляционного анализа необходимо убрать тренд изучаемых временны́х рядов. Распространенным методом детрендирования является переход к временны́м рядам первых разностей $z(j) = x(j + 1) - x(j)$ (Бокс, Дженкинс, 1974). В этом случае среднее значение временно́го ряда будет равно 0. Фактически переход к первым разностям эквивалентен выделению высокочастотной составляющей временно́го ряда радиального прироста.
Качество регуляции системы можно определить по двум показателям: порядку авторегрессии (зависимости значения переменной z(j) от величин z(j–n) предыдущих n лет) и колебаниям амплитуды временно́го ряда первых разностей.
Зависимость текущего значения переменной от n ее предыдущих значений характеризуется порядком авторегрессии (Бокс, Дженкинс, 1974):
где z0, a(r) – коэффициенты, n – порядок авторегресссии (число предыдущих значений z, от которых зависит z(j).Для определения порядка n авторегрессии рядов первых разностей прироста производился расчет парциальной авторегрессионной функции (ПАКФ) этих рядов (Бокс, Дженкинс, 1974). ПАКФ показывает корреляцию между значениями двух переменных временно́го ряда, разделенных временем r, за вычетом влияния всех внутренних значений автокорреляции. Наибольшее значение сдвига n, при котором ПАКФ статистически значимо еще отличается от нуля, дает величину порядка n авторегрессии ряда z(j) (Бокс, Дженкинс, 1974; Сток, Ватсон, 2015; Подкорытова, Соколов, 2016). Величина порядка n авторегрессии указывает на то, что значение временно́го ряда в момент времени t зависит от значений временно́го ряда, начиная с момента времени (t – 1) и заканчивая моментом времени (t – n). Можно охарактеризовать n как своеобразную “память” системы.
На рис. 1 приведены типичные ПАКФ у деревьев на разных пробных площадях. Как следует из рис. 1а, порядок ПАКФ n = 0, из рис. 1б – порядок ПАКФ n = 3.
Рис. 1.
Типичные парциальные автокорреляционные функции ряда первых разностей радиального прироста дерева. 1 – ПАКФ, 2 – границы доверительного интервала ПАКФ – стандартная ошибка ПАКФ; (а) – дерево на пробной площади 1, n = 0; (б) – дерево на пробной площади 3, n = 3.

Расчет ПАКФ рядов первых разностей деревьев в очагах массового размножения и неповрежденных насаждениях производился в пакете Statistica 6.0.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Средние значения Х1 радиального прироста в течение 1942–1972 гг. у деревьев в очагах массового размножения (Х1 = 0.85 мм при стандартном отклонении σ = 0.23 мм) значимо не отличались от средних значений Х2 радиального прироста в контрольных неповрежденных насаждениях (Х2 = 0.87 мм при стандартном отклонении σ = 0.16 мм). Таким образом, по абсолютным значениям радиального прироста до начала вспышки массового размножения деревья в очагах и на неповрежденных участках не различались. Поэтому для анализа связи между привлекательностью деревьев для насекомых и процессами их роста будем рассматривать характеристики рядов первых разностей радиального прироста.
Для оценки колебаний амплитуды рядов первых разностей радиального прироста деревьев использовалась величина стандартного отклонения s:
где m – выбранная длина временного ряда (30 лет) до начала вспышки массового размножения (1974 г.).У эффективно регулируемой системы должны быть достаточно малы величины стандартного отклонения s. Порядок n авторегрессии характеризует связь приростов соседних лет. Снижение качества регулирования будет отражаться как в увеличении значения s стандартного отклонения рядов первых разностей, так и в изменении порядка n авторегрессии.
Ряды первых разностей у деревьев в очаге массового размножения сосновой пяденицы и в неповрежденных насаждениях различаются по показателям запаздывания n и стандартного отклонения s, представленным в табл. 1. Каждая ее клетка характеризует долю деревьев в выборке с соответствующими значениями s и n. Как видно из таблицы, в очагах массового размножения преобладают (54.5%) деревья, у которых показатель запаздывания равен 0 (фактически это означает, что радиальный прирост дерева определяется текущими условиями внешней среды, тогда как в неповрежденных насекомыми насаждениях 60.5% деревьев характеризуются запаздываем n = 1 (т.е. текущее значение радиального прироста зависит от значения радиального прироста предыдущего года). У 81.4% деревьев вне очага массового размножения стандартное отклонение s рядов первых разностей радиального прироста не превышает 0.30, тогда как у деревьев в очаге массового размножения более чем у половины деревьев s > 0.30. У деревьев в очагах массового размножения либо отсутствует связь текущего прироста с приростом прошлых лет (их можно охарактеризовать как деревья с безинерционным типом регуляции роста), либо, напротив, порядок авторегрессии рядов первых разностей превосходит 1 (деревья с сильно инерционным типом регуляции радиального роста).
Таблица 1.
Сопряженность регуляторных характеристик s и n рядов первых разностей радиального прироста в период, предшествующий вспышке массового размножения*
| Показатель запаздывания n | Стандартное отклонение s | ||
|---|---|---|---|
| <0.30 | >0.30 | всего | |
| 0 | 0.186/0.284 | 0.070/0.261 | 0.256/0.545 |
| 1 | 0.512/0.080 | 0.093/0.170 | 0.605/0.250 |
| >1 | 0.116/0.102 | 0.023/0.102 | 0.140/0.205 |
| Всего | 0.814/0.466 | 0.186/0.534 | 1.000 |
Для оценки статистической значимости различия долей деревьев с различными регуляторными характеристиками в очагах массового размножения и неповрежденных насаждениях использовался t-критерий Стьюдента (Поллард, 1976). Расчеты показали, что доли деревьев с n = 0 в насаждениях, поврежденных насекомыми, значимо больше доли деревьев с n = 0 в неповрежденных насаждениях (t = 3.40 при табличном значении tтабл. = 2.6 для уровня значимости р = 0.99), а доля деревьев с n = 1 в поврежденных насаждениях значимо меньше доли таких деревьев в неповрежденных насаждениях (t = 4.04), Доля деревьев с размахом s, меньшем критического значения s = = 0.3 колебаний первых разностей радиальных приростов в неповрежденных насаждениях значимо (t = 4.36) больше доли таких деревьев в поврежденных насаждениях.
Таким образом, можно говорить о том, что деревья в очагах массового размножения сосновой пяденицы и в неповрежденных насаждениях различаются по инерционности процессов регуляции радиального прироста.
Возникает вопрос о связи между характеристиками регуляторных процессов у деревьев в очагах массового размножения и показателями восстановления радиального роста после окончания вспышки.
При повреждении деревьев насекомыми в течение одного-двух лет после повреждениях радиальный прирост отсутствует, а затем идет его восстановление (рис. 2).
Рис. 2.
Типичная динамика падения и восстановления радиального прироста деревьев после повреждения насекомыми (дерево № 55, пробная площадь 1).

Как видно из рис. 2, в течение 3 лет в ходе вспышки радиальный рост дерева прекратился. После этого началось восстановление радиального прироста, связанное с восстановлением хвои. В качестве показателей восстановления будем использовать продолжительность Т периода восстановления между датой t0 прекращения прироста и датой tm окончания восстановления, а также отношение $w = \frac{{{{x}_{m}}}}{{{{x}_{0}}}}$ радиальных приростов xm и x0 в моменты времени tm и t0. Если w > 1, можно говорить о перерегуляции в ходе процесса восстановления.
Для оценки даты прекращения процесса восстановления рассматривался временной ряд высокочастотной составляющей радиального прироста. Низкочастотная составляющая y(i) ряда радиального прироста x(i) с 1972 по 2015 г. находилась с помощью фильтра Ганна (Хемминг, 1980):
Высокочастотная составляющая $z(i) = x(i) - y(i).$На рис. 3 приведен исходный ряд радиального прироста x(i) и его высокочастотная соcтавляющая.
Рис. 3.
Исходный ряд (1) радиального прироста и высокочастотная соcтавляющая ширины годичного кольца (2) этого ряда.

Как видно из рис. 3, ряд высокочастотной составляющей радиального прироста гетероскедастичен и дисперсия значений его ряда на начальном этапе восстановления весьма мала, но затем сильно возрастает. Можно предположить, что возрастание колебаний ряда радиального прироста соответствует началу процесса регуляции прироста в норме. И в качестве даты окончания процесса восстановления выбран год, когда колебания ряда высокочастотной составляющей начинают возрастать.
При изучении связи стандартного отклонения s колебаний и порядка n рядов первых разностей радиального прироста до повреждения деревьев насекомыми и интенсивностью восстановления w рассматривались отдельно деревья с безинерционным типом регуляции (порядок авторегрессии n = 0) (рис. 4) и деревья c инерционным типом регуляции (порядок авторегресии n > 0) (рис. 5).
Рис. 4.
Связь между стандартным отклонением s рядов первых разностей радиального прироста деревьев с безинерционным типом регуляции (n = 0) до начала вспышки массового размножения и интенсивностью восстановления w радиального прироста после повреждения.

Рис. 5.
Связь между стандартным отклонением s рядов первых разностей радиального прироста деревьев с инерционным типом регуляции (n > 0) до начала вспышки массового размножения и интенсивностью восстановления w радиального прироста после повреждения.
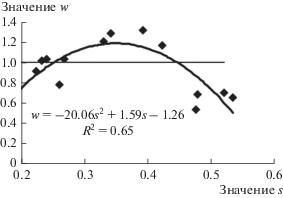
Как следует из рис. 4, для деревьев с безинерционным типом регуляции радиального прироста наблюдается следующая тенденция: чем больше значение s (т.е., чем неустойчивее регуляция дерева до его повреждения), тем менее выражен процесс восстановления радиального прироста и тем меньше значения w. При меньших значениях s имеет место перерегуляция процесса восстановления и w >1. При этом коэффициент детерминации R2 невелик (0.317) и жесткой связи между s и w нет. Для деревьев с инерционным типом регуляции (рис. 5) характер связи между стандартным отклонением s рядов первых разностей радиального прироста и интенсивностью восстановления w немонотонен. При значениях s от 0.27 до 0.40 имеет место перерегуляция процесса восстановления и w > 1. При увеличении значений s > 0.27 интенсивность восстановления w уменьшается с увеличением s. При малых же значениях s интенсивность восстановления растет с увеличением s.
Заключение. Для оценки состояния деревьев и возможности нападения на них насекомых-филлофагов предложено использовать характеристики регуляции радиального прироста деревьев. В качестве показателей регуляторного процесса радиального прироста использовано запаздывание (авторегрессия) и размах рядов первых разностей радиального прироста. Проведенный анализ позволяет говорить, что регуляторные процессы радиального прироста у большинства деревьев в неповрежденных насаждениях характеризовались запаздыванием n = 1 и малой амплитудой изменений прироста (s < 0.30) относительно его возрастного тренда. Большая часть деревьев, на которые напали насекомые, напротив, характеризовалась либо отсутствием запаздывания в рядах первых разностей радиального прироста, либо достаточно высокими значениями порядка n авторегрессии рядов первых разностей радиального прироста. При этом колебания значений текущего радиального прироста относительно возрастного тренда у деревьев в очагах массового размножения были существенно больше, чем у деревьев в неповрежденных насаждениях.
Было показано, что существует связь между характеристиками регуляторных процессов прироста деревьев до начала их повреждения филлофагами и скоростью восстановления радиального роста деревьев после повреждений.
Представляется, что анализ регуляторных процессов в рядах радиального прироста деревьев в насаждении может быть использован как для оценки риска возникновения вспышки массового размножения насекомых-вредителей в насаждении, так и для оценки продолжительности процессов восстановления деревьев после повреждений хвои вредителями.
Список литературы
Артемьева Н.В., Ряполов В.Я., Суховольский В.Г. Статистический анализ ландшафтной приуроченности природных явлений на примере очагов массового размножения лесных насекомых // География и природные ресурсы. 1995. № 2. С. 141–146.
Бокс Дж., Дженкинс Г. Анализ временных рядов. Прогноз и управление. М.: Мир. 1974. Вып 1. 406 с.
Веремей Е.И. Линейные системы с обратной связью. СПб.: Лань, 2013. 448 с.
Дорф Р.К., Бишоп Р.Х. Современные системы управления. М.: Лаборатория базовых знаний, 2004. 832 с.
Исаев А.С., Ряполов В.Я. Анализ ландшафтно-экологической приуроченности очагов сибирского шелкопряда с применением аэрокосмической съемки // Исследование таежных ландшафтов дистанционными методами. Новосибирск: Наука. 1979. С. 152–167.
Исаев А.С., Пальникова Е.Н., Суховольский В.Г., Тарасова О.В. Динамика численности лесных насекомых-филлофагов: модели и прогнозы. М.: Товарищество научных изданий КМК, 2015. 276 с.
Ким Д.П. Теория автоматического управления. Т. 1. Линейные системы. М.: ФИЗМАТЛИТ, 2007. 312 с.
Мозолевская Е.Г., Селиховкин А.В., Ижевский С.С., Захаров А.А., Голосова М.А., Никитский Н.Б. Лесная энтомология. М.: “Академия”, 2011. 416 с.
Пальникова Е.Н. Свидерская И.В., Тарасова О.В., Ягунов М.Н., Суховольский В.Г. Лесные насекомые как экологические инженеры: оценка воздействия на лесные экосистемы // Интенсификация лесного хозяйства России: проблемы и инновационные пути решения. Красноярск, 19 -23 сентября 2016 г. Красноярск: ИЛ СО РАН, 2016. С. 147–148.
Пальникова Е.Н., Свидерская И.В., Суховольский В.Г. Сосновая пяденица в лесах Сибири. Новосибирск: Наука, 2002. 232 с.
Подкорытова О.А., Соколов М.В. Анализ временных рядов. М.: Юрайт, 2016. 266 с.
Поллард Дж. Справочник по вычислительным методам статистики. М.: Финансы и статистика, 1982. 344 с.
Поляк Б.Т., Щербаков П.С. Робастная устойчивость и управление. М.: Наука, 2002. 303 с.
Свидерская И.В. Пальникова Е.Н. Радиальный рост сосны в связи с дефолиацией сосновой пяденицей // Лесоведение. 2003. № 5. С. 44–53.
Скрипальщикова Л.Н., Татаринцев А.И., Зубарева О.Н., Перевозникова В.Д., Стасова В.В., Грешилова Н.В. Экологическое состояние пригородных лесов Красноярска. Новосибирск: Гео, 2009. 179 с.
Сток Дж.Х., Ватсон М. Введение в эконометрику. М.: Издательский дом “Дело” РАНХиГС, 2015. 864 с.
Суховольский В.Г., Ковалев А.В., Сотниченко Д.В. Диэлектрическая спектроскопия прикамбиального комплекса тканей и экспресс-диагностика состояния древесных растений // Эколого-географические аспекты лесообразовательного процесса. Красноярск, 19–23 сентября 2014 г. Красноярск: ИЛ СО РАН, 2009. С. 335–336.
Тарасова О.В. Ландшафтно-экологическая специфика вредной лесной энтомофауны Минусинских ленточных боров // Насекомые лесостепных боров Сибири. Новосибирск: Наука, 1982. С. 18–34.
Тарасова О.В., Калашникова И.И., Кузнецова В.В. Энергетический баланс потребления корма насекомыми-филлофагами: оптимизационная модель // Сибирский лесной журнал. 2015. № 3. С. 83–92.
Хемминг Р.В. Цифровые фильтры. М.: Советское радио, 1980. 224 с.
Armour H., Straw N., Day K. Interactions between growth, herbivory and long-term foliar dynamics of Scots pine // Trees. 2003. V. 17 (1). P. 70–80.
Barton K.E., Koricheva J. The ontogeny of plant defense and herbivory: characterizing general patterns using meta-analysis// American Naturalist. 2010. V. 175 (4). P. 481–493.
Büchel K., Fenning T., Gershenzon J., Hilker M., Meiners T. Elm defence against herbivores and pathogens: morphological, chemical and molecular regulation aspects// Phytochemestry Reviews. 2016. V. 15 (5). P. 961–983.
Castagneyrol B., Régolini M., Jactel H. Tree species composition rather than diversity triggers associational resistance to the pine processionary moth // Basic and Applied Ecology. 2014, V. 15 (6). P. 516–523.
Delvas N., Bauce É., Labbé C., Ollevier T., Bélanger R. Phenolic compounds that confer resistance to spruce budworm// Entomologia Experimentalis et Applicata. 2011. V. 141 (1). P. 35–44.
Ferrenberg S., Kane J.M., Mitton·J.B. Resin duct characteristics associated with tree resistance to bark beetles across lodgepole and limber pines // Oecologia. 2014. V. 174 (4). P. 1283–1292.
Fuentealba A., Bauce É. Interspecific variation in resistance of two host tree species to spruce budworm // Acta Oecologica. 2016. V. 70. P. 10–20.
Fuentealba A., Bauce É. Site factors and management influence short-term host resistance to spruce budworm, Choristoneura fumiferana (Clem.), in a species-specific manner // Pest Management Science. 2012. V. 68 (2). P. 245–253.
Hard J.S. Spruce beetles attack slowly growing spruce// Forest Science. 1985. V. 31 (4). P. 839–850.
Haukioja E. Plant defenses and population fluctuations of forest defoliators: mechanism-based scenarios // Annales Zoologici Fennici. 2005. V. 42 (4). P. 313–325.
Haukioja E., Ossipov V., Lempa K. Interactive effects of leaf maturation and phenolics on consumption and growth of a geometrid moth // Entomologia Experimentalis et Applicata. 2002. V. 104 (1). P. 125–136.
Herms D.A., Mattson W.J. The dilemma of plants: to grow or defend // The Quarterly Review of Biology. 1992. V. 67 (3). P. 283–335.
Huberty A.F., Denno R.F. Plant water stress and its consequences for herbivorous insects: a new synthesis // Ecology. 2004. V. 85 (5). P. 1383–1398.
Isaev A.S., Soukhovolsky V.G., Tarasova O.V., Palnikova E.N. and Kovalev A.V. Forest insect population dynamics, outbreaks and global warming effects. New York: J. Wiley and Sons, 2017. 286 p.
Kumbas M., Bauce É. Spruce budworm biological and nutritional performance responses to varying levels of monoterpenes // iForest-Biogeosciences and Forestry. 2013. V. 6 (6). P. 310–314.
Larsson S. Resistance in trees to insects – an overview of mechanisms and interactions // Mechanisms and Deployment of Resistance in Trees to Insects (eds. Wagner M.R., Clancy K.M., Lieutier F., Paine T.D.). New York: Kluwer Academic Publishers, 2002. P. 1–29.
Mumm R., Hilker M. Direct and indirect chemical defence of pine against folivorous insects // Trends in Plant Science. 2006. V. 11 (7). P. 351–358.
Stevens M.T., Waller D.M., Lindroth R.L. Resistance and tolerance in Populus tremuloides: genetic variation, costs, and environmental dependency // Evolutionary Ecology. 2007. V. 21 (6). P. 829–847.
Strauss S.Y., Agrawal A.A. The ecology and evolution of plant tolerance to herbivory // Trends in Ecology & Evolution. 1999. V. 14 (5). P. 179–185.
Straw N.A. The impact of pine looper moth, Bupalus piniaria L. (Lepidoptera; Geometridae) on the growth of Scots pine in Tentsmuir Forest, Scotland. // Forest Ecology and Management. 1996. V. 87 (1–3). P. 209–232.
Thomas A.W. Food consumption and utilization by 6th-instar larvae of spruce budworm, Choristoneura fumiferana: a comparison on three Picea (spruce) species// Entomologia Experimentalis et Applicata. 1989. V. 52 (3). P. 205–214.
Wise M.J., Abrahamson W.G. Effects of resource availability on tolerance of herbivory: a review and assessment of three opposing models // American Naturalist. 2007. V. 169 (4). P. 443–454.
https://www.ldeo.columbia.edu/tree-ring-laboratory/resources/software Lamon-Doherty Earth Observatory, Columbia University Earth Institute (дата обращения 17.02. 2015)
Дополнительные материалы отсутствуют.