Автоматика и телемеханика, № 6, 2022
© 2022 г. И.С. ПРОСКУРКИН, канд. физ.-мат. наук
(megavolt007@mail.ru),
В.К. ВАНАГ, д-р. физ.-мат. наук (vvanag@kantiana.ru)
(Центр нелинейной химии, Балтийский федеральный университет
им. Иммануила Канта, Калининград)
СЛУЧАЙНОЕ ПРИНЯТИЕ РЕШЕНИЙ В СЕТЯХ
ИМПУЛЬСНО СВЯЗАННЫХ СПАЙКОВЫХ
ОСЦИЛЛЯТОРОВ1
Теоретически и экспериментально исследована иерархическая сеть им-
пульсно связанных спайковых микроосцилляторов (МО), способная слу-
чайным образом реагировать на внешний сигнал. Сеть включает в себя
блоки Антенна, Центральный Генератор Ритмов (ЦГР) и блок Приня-
тия Решений (ПР). Внешний сигнал вызывает в Антенне противофазные
или синфазные колебания входящих в Антенну микроячеек. ЦГР так-
же обладает этими двумя колебательными модами. Какую моду принять
блоку ЦГР в ответ на появление колебательной моды в Антенне, решает
блок ПР. Благодаря своей конфигурации блок ПР принимает это реше-
ние случайно. В качестве МО используются микросферы с колебатель-
ной реакцией Белоусова-Жаботинского. Импульсные связи между МО
осуществляются сфокусированными на МО лучами света.
Ключевые слова: спайковые микроосцилляторы, импульсные связи, сети,
принятие решений, случайный выбор, реакция Белоусова-Жаботинского.
DOI: 10.31857/S0005231022060101, EDN: ADGWTB
1. Введение
Изучение сетей импульсно связанных спайковых осцилляторов приобрело
в последние годы особую актуальность [1-11]. В качестве таких осциллято-
ров рассматриваются математические модели, электрические схемы и даже
автоколебательные химические реакции [12-14]. Для изучения сетей спай-
ковых2 химических микроосцилляторов мы используем известную автоката-
литическую реакцию Белоусова-Жаботинского (БЖ) [15, 16]. Реакция БЖ
представляет собой окисление, как правило, малоновой кислоты (МА) бро-
матом в кислой среде, причем этот процесс катализируется ионами металлов
1 Данное исследование было поддержано из средств программы стратегического
академического лидерства “Приоритет 2030” Балтийского федерального университета
им. И. Канта.
2 Спайком БЖ реакции называется резкий переход системы из состояния с преиму-
щественно восстановленным катализатором в состояние с преимущественно окисленным
катализатором и обратно — из окисленного в восстановленное состояние. Такой быстрый
переход обеспечивается автокатализом.
136
или такими металло-комплексами, как ферроин, Fe(phen)2+3, или Ru(bpy)2+3,
где phen - это 1,10-фенантролин, а bpy - это 2,2′-бипиридин. Динамика БЖ
реакции подобна динамике нейронов. Поэтому построение сетей химических
микроосцилляторов позволяет выявлять и исследовать свойства, присущие
нейросетям мозга.
К настоящему времени нами исследована относительно небольшая иерар-
хическая сеть импульсно связанных химических осцилляторов (от
30
до 50 микроосцилляторов), способная демонстрировать адаптивное поведе-
ние [17]. Сеть импульсно связанных БЖ-осцилляторов может обладать муль-
тиритмичностью [18]. Эффекту мультиритмичности (или мультистабильно-
сти) даже посвящен специальный выпуск журнала Chaos [19]. Этот эффект
мы используем для создания мультиритмичного Центрального Генератора
Ритмов (ЦГР), контролирующего различные состояния (или моды) системы.
Переключения между этими состояниями осуществляются специальными им-
пульсами, которые формируются специальным блоком Принятия Решений
(ПР) [20].
Помимо функциональных блоков ПР и ЦГР наша иерархическая сеть со-
держит два блока “Ридер” и блок Антенна. Антенна принимает внешние сиг-
налы, которые ассоциируются Антенной с какой-либо модой, способной в ней
возникнуть. Ридеры считывают текущие ритмы блоков ЦГР и Антенны и
посылают информацию на блок ПР. Он в свою очередь сравнивает инфор-
мационные сигналы от двух Ридеров и в случае несовпадения ритма ЦГР
с ритмом Антенны посылает специальные импульсы на ЦГР, которые пере-
ключают его текущий ритм в другой ритм, аналогичный ритму Антенны.
Используемый нами термин “иерархическая сеть” подразумевает, что одни
блоки сети (например, Антенна) управляют динамикой других блоков сети
(например, ЦГР). Полученные ранее результаты [17, 20] наметили перспек-
тивы создания сетей, способных выполнять более сложные задачи.
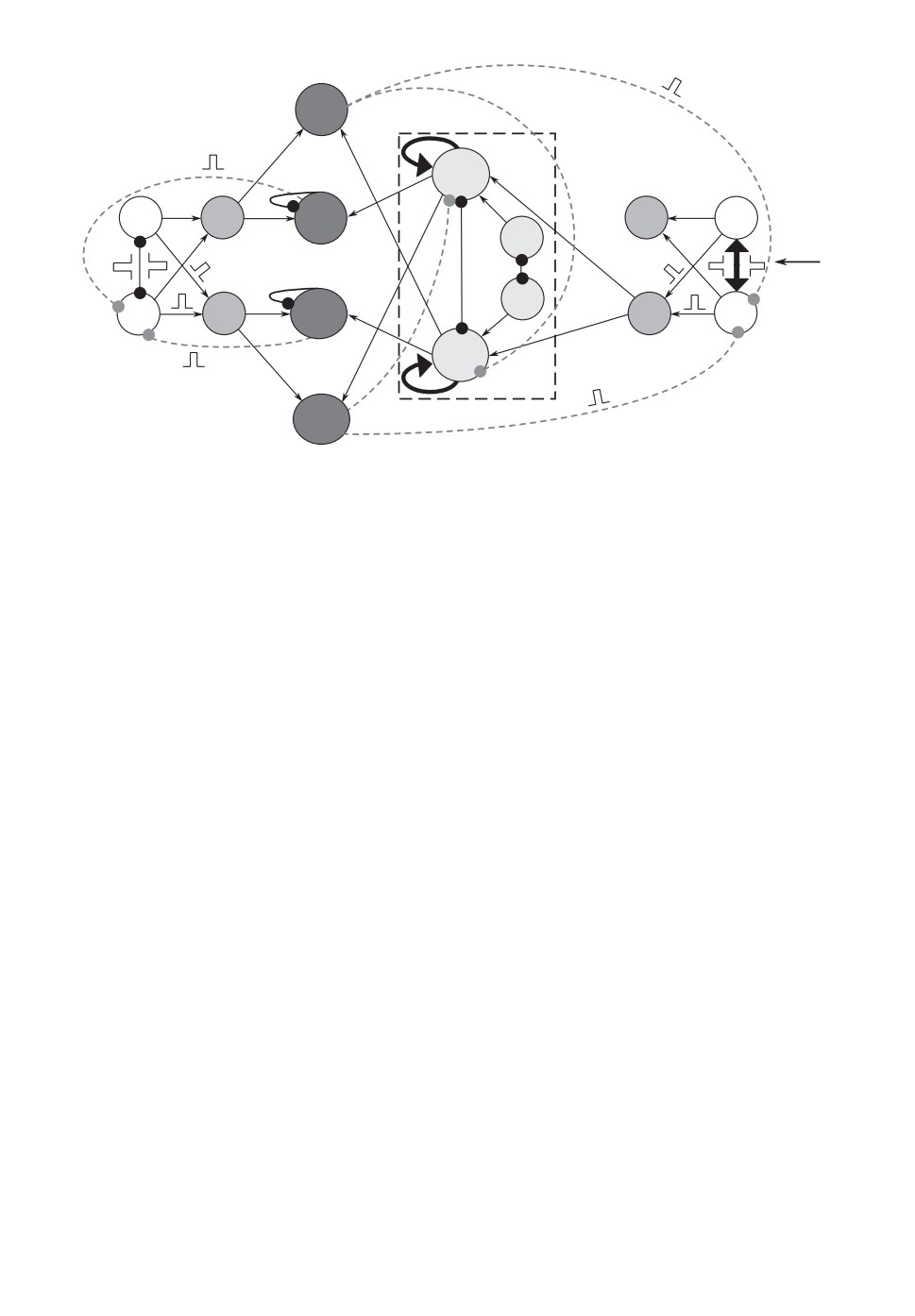
2. Принципиальное описание сети
В данной статье мы представляем новую схему иерархической нейроподоб-
ной сети БЖ осцилляторов и возбудимых ячеек (рис. 1), в которой старый
ПР блок разделен на две части. Одну часть мы назвали блоком “Исполни-
тель” (ячейки № 9-№ 12), а для другой части сохранили старое название ПР
(ячейки № 13, № 14, “Pro” и “Contra”). Но теперь новый блок ПР действи-
тельно принимает решения, а не выполняет команды, как это происходило
в старом блоке ПР. Функции же исполнения команд отданы блоку “Испол-
нитель”. Для простоты мы упростили блоки ЦГР (осцилляторы № 1 и № 2)
и Антенна (возбудимые ячейки № 5 и № 6). Теперь каждый из этих блоков
состоит всего из двух ячеек и может иметь только два ритма: синфазные и
противофазные колебания. Соответственно, и Ридеры этих блоков (“Р” и РA)
состоят только из двух возбудимых ячеек (Ридер “Р” состоит из возбудимых
ячеек № 3 и № 4, а ридер РA представлен возбудимыми ячейками № 7 и № 8).
Ячейки блока ЦГР (а также ячейки № 13 и № 14) находятся в колебательном
137
9
Pro
IP
1
3
10
7
5
IP
13
S
14
2
4
11
8
6
AP
AP
Contra
ЦГР
“P”
PA
Антенна
12
ПР
Исп
Рис. 1. Схема иерархической сети импульсно связанных спайковых осцилля-
торов и возбудимых ячеек, в которой реализуется случайный тип принятия
решения. Обозначения: ЦГР - центральный генератор ритмов; Исп - блок Ис-
полнитель; ПР - блок Принятия Решений. Черными жирными стрелочками
отмечены возбуждающие импульсы с амплитудой I, тонкими черными стре-
лочками - импульсы с амплитудой I/2. Связи с круглыми наконечниками -
это ингибиторные импульсы. Черными сплошными и серыми пунктирными
линиями обозначены ингибиторные связи внутри одного блока и между раз-
ными блоками соответственно. Подписи у ячеек Ридеров соответствуют опре-
деляемым ими ритмам, для IP (inphase) - синфазный, а для AP (antiphase) -
противофазный. Чтобы не перегружать рисунок, в схеме прорисованы лишь
те связи, которые необходимы для ее работы в случае возникновения в Ан-
тенне противофазного ритма.
режиме, а все остальные ячейки — в возбудимом состоянии с одинаковым
порогом возбуждения.
Как правило, в живых системах принятие решения является сложным
процессом, в который вовлечена обработка многих сигналов из разных ча-
стей мозга. Однако в некоторых случаях решение необходимо принять очень
быстро, не имея возможности на анализ всей информации. В принятии ре-
шения в таком случае, как правило, присутствует случайный выбор [21]. На
рис. 1 как раз и представлена сеть, предназначенная для случайного (в зна-
чении “равновероятного”) принятия решений.
Блок ЦГР может находиться в синфазном (IP) или противофазном ритме
(AP). При поступлении внешнего импульса S на Антенну в ней также возни-
кает IP или AP ритм, а Ридер РA определяет возникший ритм (= моду). Если
в Антенне возбудилась AP (IP) мода, то активируется ячейка № 8 (№ 7), ко-
торая с периодом противофазных или синфазных колебаний в Антенне (в за-
висимости от того, какая мода возбудилась в Антенне при приходе сигнала S)
138
посылает импульсы на блок ПР, точнее, на ячейки “Pro” и “Contra”. Для ак-
тивации ячеек “Pro” и “Contra”, а также ячеек № 8, № 7, № 3 и № 4 на них
должны одновременно прийти два импульса амплитудой I/2. Под словом
“одновременно” понимается такая последовательность импульсов, профили
которых в значительной степени пересекаются на временной шкале в момент
их прихода на ячейку. В этом случае суммарная амплитуда возбуждающего
сигнала достигает порогового значения I, при котором возбудимая ячейка
генерирует спайк.
Чтобы каждая ячейка Ридера РA активировалась в ответ на определенный
ритм, два импульса от ячеек Антенны приходят на ячейки Ридера с разными
фазами. На ячейку № 7 импульсы от ячеек № 5 и № 6 приходят без вре-
менных задержек. В этом случае ячейка № 7 возбуждается, если в Антенне
установилась IP мода. На ячейку № 8 импульсы от ячеек № 5 и № 6 приходят
с разностью фаз, равной половине периода противофазных колебаний TAP.
Поэтому ячейка № 8 возбуждается, если в Антенне установилась AP мода.
Аналогичным образом работает Ридер “Р” блока ЦГР.
Случайность принятия решения заключается в случайной активации яче-
ек “Pro” или “Contra”. Если случайно активируется ячейка “Pro”, то в блоке
ЦГР должен установиться такой же внутренний ритм, как и в Антенне. И на-
оборот, если случайно активируется ячейка “Contra”, то в блоке ЦГР должен
установиться ритм, противоположный ритму Антенны. Блок Исполнитель
выполняет всю “техническую” работу по установлению нужного ритма в ЦГР.
Ячейки блока Исполнитель активируются только при одновременном прихо-
де на них двух импульсов: одного от одной из ячеек Ридера “Р”, а другого от
одной из ячеек блока ПР. Если одна из ячеек № 10 или № 11 активировалась,
то Исполнитель отправляет переключающий сигнал на блок ЦГР, изменяя
его ритм на ритм, аналогичный или противоположный ритму Антенны, в
зависимости от того, активна ячейка “Pro” или “Contra”.
В качестве примера работы блока Исполнитель рассмотрим случай, когда
активны ячейки “Pro” и № 3. Одновременный приход двух импульсов от яче-
ек “Pro” и № 3 на ячейку № 10 возбуждает ее. В активированной ячейке № 10
рождается спайк и ячейка № 10 отправляет на ячейку № 2 блока ЦГР пере-
ключающий ингибиторный импульс. Этот импульс переключает синфазную
моду ЦГР в противофазную, а родившаяся в ЦГР AP мода регистрируется
ячейкой № 4 Ридера “Р”. Так как переключающий импульс, отправленный на
ячейку № 2, имеет временную задержку, превышающую периоды ритмов в
блоках ЦГР и Антенна, то за это время задержки ячейка № 10 может сгене-
рировать второй спайк, что нарушает правильную работу сети. Поэтому для
ячейки № 10 (это же относится и к ячейке № 11) дополнительно введено им-
пульсное самоингибирование, что позволяет ячейке № 10 оставаться какое-то
время после первого спайка нечувствительной к возбуждающим импульсам.
Случайный выбор принятия решения реализуется через совпадение двух
независимых событий, а именно: прихода на ячейки “Pro” и “Contra” двух им-
пульсов возбуждения от других, не синхронизованных между собой осцилля-
139
торов, имеющих разные частоты. В качестве одного осциллятора выступает
одна из ячеек Ридера РA (например, ячейка № 8, если в Антенне установилась
AP мода). А в качестве второго осциллятора выступает один из осцилляторов
пары импульсно связанных ингибиторной связью осцилляторов № 13 и № 14.
Благодаря правильно подобранным параметрам ингибиторной связи эти ос-
цилляторы колеблются в противофазе. Осциллятор № 13 посылает импульсы
на ячейку “Pro”, а осциллятор № 14 — на ячейку “Contra”, которые прихо-
дят на эти ячейки в противофазе, т.е. в разные моменты времени. Второй
импульс, необходимый для активации ячеек “Pro” и “Contra”, поступает от
осциллятора № 8 (в случае AP моды в Антенне). Заметим, что осциллятор
№ 8 начинает работать в случайный момент времени, определяемый случай-
ным моментом прихода на Антенну внешнего импульса S. Поэтому одновре-
менный приход двух возбуждающих импульсов на ячейку “Pro” или “Contra”
является случайным событием.
Если одна из ячеек “Pro” или “Contra” возбудилась (что равносильно при-
нятию решения), то нужно эту возбужденную ячейку перевести в колебатель-
ную моду, которая должна существовать некоторое время до момента испол-
нения принятого решения. Поэтому мы вводим для ячеек “Pro” и “Contra”
дополнительные возбуждающие импульсы самоактивации с амплитудой I,
которые периодически вызывают спайки в этих ячейках после первой их ак-
тивации. Период самоактивации ячеек “Pro” и “Contra”, который определяет-
ся временем задержки импульса самоактивации, подобран так, чтобы он был
приблизительно равен периоду синфазных колебаний в блоке Антенна. Если
возбудилась одна из ячеек “Pro” или “Contra”, то активация второй ячейки
должна стать невозможной. Для выполнения этого условия мы вводим меж-
ду ячейками “Pro” и “Contra”двустороннюю импульсную ингибиторную связь
большой амплитуды [22].
После того как все необходимые переключения завершены и случайно при-
нятое решение выполнено, наша сеть должна вернуться в исходное состояние,
в котором она была бы готова принимать новые случайные решения, т.е. все
ячейки кроме ячеек блоков ЦГР, “Р” и ячеек № 13 и № 14 должны оказаться
в стационарном возбудимом состоянии. Эту работу выполняют ячейки № 9
и №12, посылая ингибиторные импульсы на ячейки “Pro” и “Contra” блока ПР
и на ячейку № 6 блока Антенна. Ингибиторные импульсы можно посылать и
на ячейку № 5 блока Антенна, но это не имеет значения.
Возможен также вариант, когда возбуждаются ячейки “Pro” или “Contra”,
а изначальный ритм блока ЦГР уже аналогичен или противоположен, со-
ответственно, ритму Антенны. В этом случае переключения моды ЦГР не
происходит. Вместо этого на Антенну и блок ПР отправляются от ячеек № 9
или № 12 ингибиторные импульсы обратных связей, которые возвращают эти
блоки в их исходное состояние.
Таким образом, сеть импульсно связанных ячеек, представленная на
рис. 1, получая внешний сигнал S, случайным образом решает, установить
ли в ЦГР ритм, аналогичный или противоположный ритму Антенны.
140
3. Эксперимент
В качестве БЖ микроосцилляторов и возбудимых микроячеек мы исполь-
зуем микросферы ионообменной смолы Dowex
50WX2 с иммобилизован-
ным в них светочувствительным катализатором БЖ реакции, Ru(bpy)2+3, так
называемые БЖ микросферы [23]. Метод приготовления микросфер Dowex
50WX2 с иммобилизованным Ru(bpy)2-3 приведен в ссылке [24]. БЖ микро-
сферы, имеющие средний диаметр около 100 микрометров (см. рис. 2), по-
мещаются в обращенную микроэмульсию АOT [24], наполненную всеми ре-
агентами БЖ реакции, но без катализатора. Для приготовления АОТ мик-
роэмульсии [24] в качестве непрерывной гидрофобной фазы использовался
тетрадекан. Концентрация Ru(bpy)2+3в микросферах была высчитана рав-
ной 3 мМ. Водные нанокапли АОТ микроэмульсии с БЖ реагентами по-
стоянно подпитывают БЖ микросферы в силу диффузионного обмена со-
держимым между нанокаплями и БЖ микросферами.
При достаточно большом расстоянии между БЖ микросферами они ве-
дут себя независимым образом. Мы помещали БЖ микросферы в реактор
(см. рис. 2), в качестве которого служит склеенная из предметных стеклы-
шек прямоугольная ванночка глубиной 1 мм. После наполнения БЖ микро-
эмульсией и БЖ микросферами ванночка накрывалась оптическим стеклом,
чтобы предотвратить возможное испарение тетрадекана.
14
Contra
1
2
8
10
13
9
Pro
6
7
5
11
3
4
12
200 мкм
Рис. 2. Снимок используемых в эксперименте БЖ микросфер, находящихся
в реакторе. Квадратными рамками отмечены границы областей облучения
актиничным светом всех БЖ микросфер, используемых для реализации сети,
представленной на рис. 1. Номера рядом с БЖ микросферами соответствуют
номерам ячеек на схеме рис. 1. Не отмеченные номерами БЖ микросферы во
время эксперимента находятся в колебательном состоянии и никак не влияют
на динамику исследуемых БЖ микросфер из-за их удаленного расположения,
исключающего действие диффузионных связей. Две длинные черные линии
на рисунке являются “верхней” и “нижней” границами реактора (правая и
левая границы находятся за пределами рисунка).
141
Камера
ПК
М
ПЗ
510 нм
Подсветка
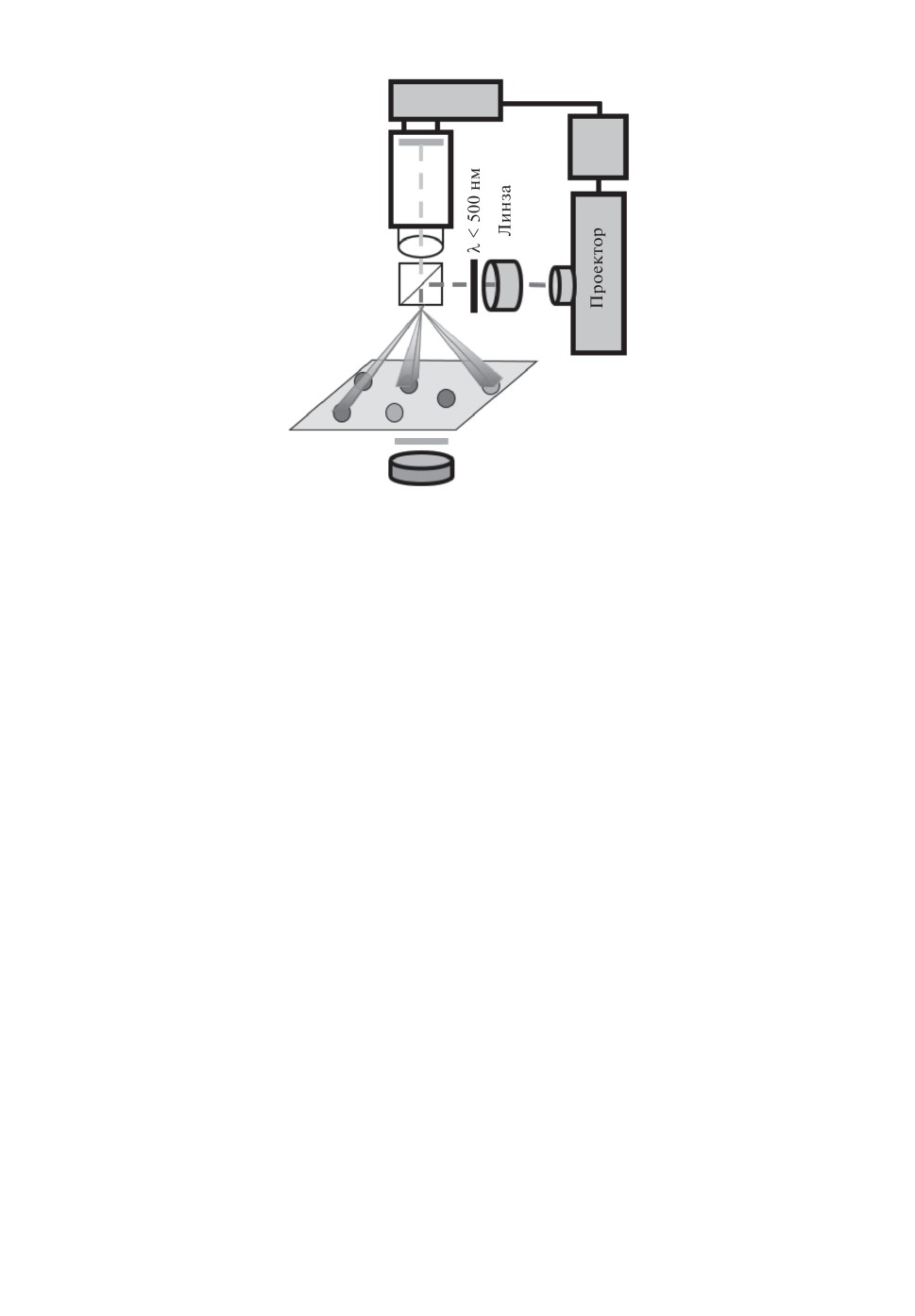
Рис. 3. Схема лабораторной установки. М - микроскоп; ПЗ - полупрозрачное
зеркало с соотношением отражение/пропускание, равным 50/50; ПК - пер-
сональный компьютер. Серыми пластинками отмечены интерференционные
фильтры с длиной волны λ = 510 нм и шириной пропускания Δλ = 10 нм.
Черной пластинкой отмечен коротковолновый фильтр с границей отрезания
на длине волны λ = 500 нм. Серыми и светло-серыми шариками представле-
ны БЖ микросферы в реакторе. Проектор преобразует динамическую маску,
контролируемую персональным компьютером, в лучи актиничного света, а
линза фокусирует эти лучи на микросферы. Максимально возможная интен-
сивность актиничного света Iмакс = 2,07 клк (килолюкса). Анализирующий
свет подсветки проходит сквозь микросферы и регистрируется камерой. Рез-
кое увеличение яркости микросферы отмечается программой LabVIEW как
спайк осциллятора.
Микроэмульсия АОТ описывается двумя физическими параметрами:
ω = [H2O]/[AOT] и объемной долей нанокапелек ϕd
[25]. Величина ω
определяет радиус водного ядра нанокапелек Rw (нм) приблизительно
как Rw = 0,17ω. Для нашего случая ω = 10, а ϕd = 0,45. Концентрации
БЖ реагентов в водной псевдофазе АОТ микроэмульсии: [MA] = 0,36 M,
[H2SO4] = 0,5 M, [NaBrO3] = 0,27 M, [NaBr] = 0,07 M, где МА — это мало-
новая кислота. Эти концентрации поддерживают в БЖ микросферах коле-
бания с периодом около 40 с, которые являются устойчивыми на протяжении
не менее 100 мин.
С помощью видеокамеры, оборудованной на окуляре стереомикроскопа,
регистрируется изменение яркости проходящего сквозь БЖ микросферы све-
та с длиной волны λ = 510 нм (применяется интерференционный фильтр)
(см. рис. 3). Свет этой длины волны не влияет на динамику фоточувстви-
тельной БЖ реакции.
142
Для создания импульсных связей мы используем актиничный свет с дли-
ной волны λ < 500 нм (используется коротковолновый фильтр). В условиях
нашей реакции импульсы такого света увеличивают период БЖ колебаний,
т.е. являются ингибиторными импульсами. Увеличение амплитуды импульса
(интенсивности освещенности) приводит к большему ингибированию. Сфо-
кусированные лучи актиничного света, падающие на индивидуальные мик-
росферы, создаются мощной линзой и проектором, в котором устанавливает-
ся нужная геометрическая маска, контролируемая компьютером. Необходи-
мые области пикселей в маске принимают определенную степень яркости в
нужные моменты времени, т.е. маска является динамической. Компьютерная
программа, разработанная авторами в среде LabVIEW, анализирует состоя-
ние осцилляторов и контролирует все параметры импульсных связей, такие
как амплитуда, длительность и задержка импульса, и создает динамическую
маску, которая проецируется линзой и полупрозрачным зеркалом (ПЗ) на
микросферы.
Исходно все БЖ микросферы находятся в колебательном состоянии при
выбранных концентрациях БЖ реагентов. Для создания из них возбудимых
ячеек мы используем постоянное облучение светом определенной интенсивно-
сти I, при которой БЖ микросферы ингибируются и переходят в возбудимое
состояние. Известно, что если на короткий промежуток времени Δt прервать
это ингибиторное облучение, то возбудимая ячейка даст спайк [26]. Таким об-
разом, импульсное прекращение постоянного облучения используется нами в
качестве возбуждающего импульса — это так называемый “отрицательный”
импульс. Для реализации возбуждающих импульсов разной амплитуды мы
понижаем интенсивность ингибиторного освещения возбудимых ячеек на ам-
плитуду I для случая, если к этой ячейке на рис. 1 идет жирная черная стре-
лочка, и на амплитуду I/2, если к этой ячейке на рис. 1 идет тонкая черная
стрелочка. Если интенсивность освещения возбудимой ячейки понижается до
нуля (что соответствует амплитуде отрицательного возбуждающего импуль-
са равной I), то такая ячейка дает спайк в случае продолжительности этого
импульса не менее Δt = 10 c, но для получения стабильного результата вы-
брано Δt = 15 c. Приход на ячейку одного отрицательного возбуждающего
импульса с амплитудой I/2 и длительностью Δt = 15 c понижает порог возбу-
димости ячейки. Поэтому для ее активации достаточно прихода второго им-
пульса, который пересекается во времени с первым уже не в течение 10-15 с,
а в течение Δtx = 5 с (или больше).
Любой импульс в нашей сети, вышедший из одной ячейки и идущий к
другой ячейке, характеризуется тремя параметрами: (1) время задержки τ
между моментом рождения импульса (= момент времени появления спай-
ка в ячейке) и моментом прихода этого импульса к адресату, т.е. к другой
ячейке, (2) длительность импульса Δt, (3) амплитуда импульса I∗. Инги-
биторная связь, например, между ячейками № 1 и № 2 создается положи-
тельными импульсами света со следующими параметрами: τ = 0 с, Δt = 3 с
и I∗ = 0,94 клк. При этих параметрах устанавливаются устойчивые проти-
143
вофазные или синфазные колебания в зависимости от начальных условий.
Напомним, что колебательные ячейки № 1 и № 2 не освещаются постоянным
светом. Если из ячейки выходит несколько импульсов, то к параметрам им-
пульсов добавляется подстрочный индекс “n_m”, где n - это номер ячейки, из
которой выходит импульс, а m - это номер ячейки, к которой устремляется
импульс.
4. Результаты эксперимента
Используя БЖ-микроосцилляторы, показанные на рис. 2, нашу програм-
му по управлению импульсами света (написанную в среде LabVIEW) и уста-
новку, схема которой показана на рис. 3, мы экспериментально воссоздали
иерархическую сеть спайковых микроосцилляторов и возбудимых микрояче-
ек, представленную на рис. 1. Работа этой сети в виде динамики ячеек всех
ее блоков представлена на рис. 4 для случая, когда случайно активировалась
ячейка “Pro”.
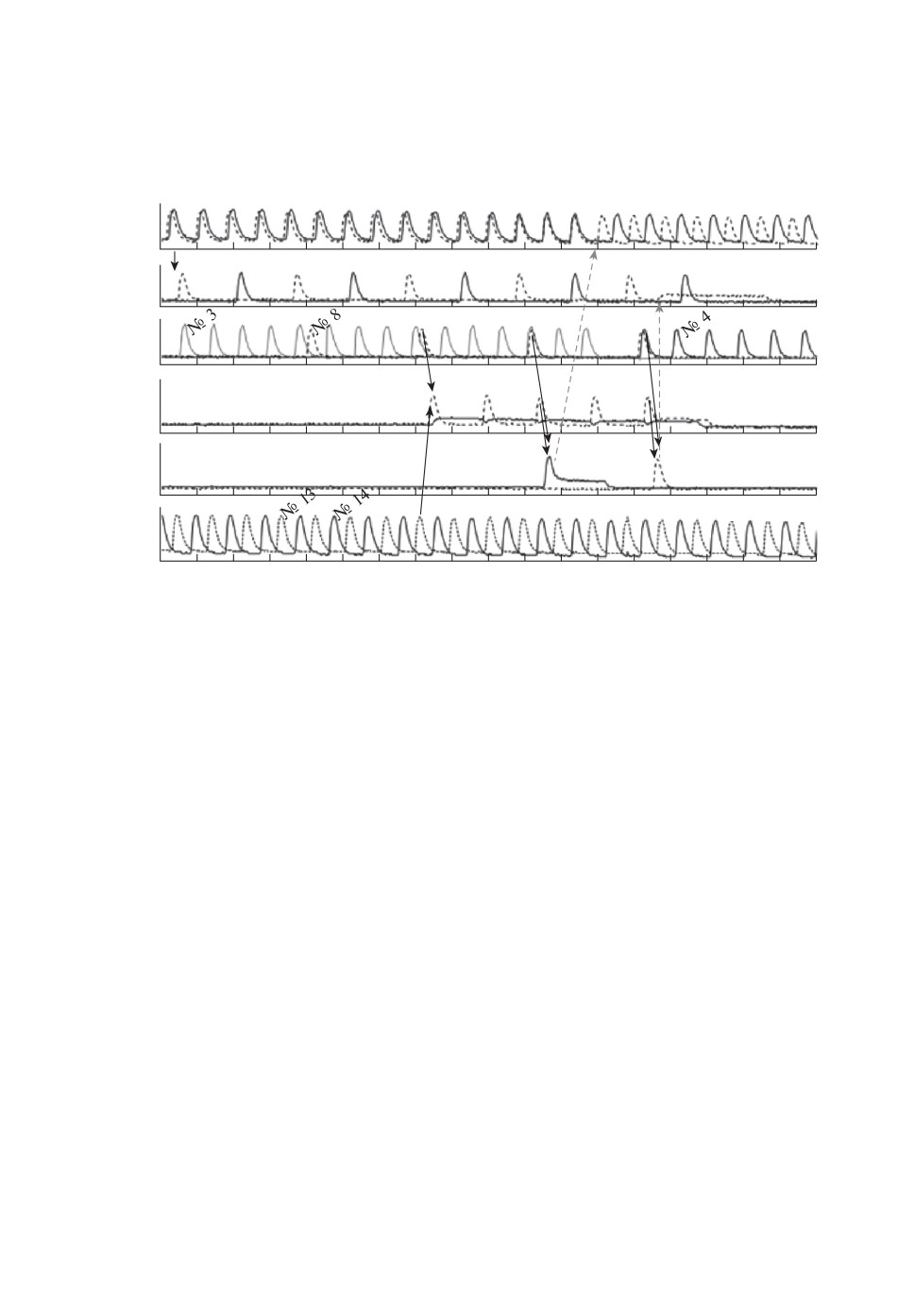
Исходно ритм блока ЦГР является синфазным в рассматриваемом при-
мере, и ячейка № 3 периодически генерирует спайки благодаря одновре-
менно приходящим на нее импульсам от осцилляторов № 1 и № 2. Ячейки
№ 13 и № 14 блока ПР постоянно колеблются в противофазе, что достига-
ется обоюдными ингибиторными импульсами со следующими параметрами:
τ13_14 = 0 с, Δt13_14 = 3 с и I13_14 = 0,94 клк. Все остальные ячейки сети
исходно находятся в возбудимом стационарном состоянии. Взаимные ингиби-
торные импульсы между ячейками Pro и Contra характеризуются следующи-
ми параметрами: τ = 0 с, Δt = 70 с и I∗ = 1,88 клк.
В случайный момент времени на Антенну поступает внешний сигнал S
(черная вертикальная стрелочка на рис. 4) и активирует одну из ее ячеек
(№ 5 или № 6), запуская в Антенне противофазный ритм. Ячейка № 8 Ридера
РA начинает генерировать спайки, регистрируя тем самым противофазный
ритм Антенны (кинетика ячейки № 8 представлена пунктирной кривой на
панельке “P”, PA рис. 4). Ячейки № 8 и № 13 отправляют возбуждающие им-
пульсы на ячейку “Pro”, а ячейки № 8 и № 14 — на ячейку “Contra”. В момент
t = 1650 c два импульса случайно приходят на ячейку “Pro” одновременно и
возбуждают ее. Это означает, что блок ПР случайно выбрал решение “Pro”.
Далее в момент времени t= 1820 с возбуждающие импульсы от ячеек № 3
и “Pro” одновременно поступают на ячейку № 10 и активируют ее. Она в свою
очередь c временной задержкой τ10_2 отправляет переключающий ингиби-
торный импульс на ячейку № 2 блока ЦГР. Величина τ10_2 выбрана равной
60 c в силу того, что при такой задержке переключающий импульс приходит
на ячейку № 2 в тот момент, когда фаза синфазных колебаний ϕIP находится
в той области, в которой синфазный режим весьма чувствителен к внешним
ингибиторным импульсам и может легко переключиться в противофазный
режим. В нашем случае ϕIP = 0,7 (фаза меняется от нуля до единицы). Если
фаза ϕIP будет еще ближе к единице, то чувствительность еще больше повы-
шается, но стабильность переключения понижается. На рис. 4 видно, что пе-
144
реключающий импульс переводит блок ЦГР в противофазную моду в момент
времени t= 1900 с. Параллельно с отправкой переключающего импульса на
ячейку № 2 ячейка № 10 самоингибируется.
ЦГР
Антенна
“P”, PA
“Pro”
“Pro”
“Contra”
№ 10
№ 12
Исп
№ 13 14
1300
1400
1500
1600
1700
1800
1900
2000
2100
2200
Время, с
Рис. 4. Динамика ячеек сети при случайной активации ячейки “Pro”. Черны-
ми сплошными линиями со стрелочками отмечены возбуждающие импульсы,
а серыми пунктирными — ингибиторные. Интенсивность постоянного освеще-
ния для всех возбудимых ячеек I = 0,19 клк. Параметры импульсов. Ингиби-
торные импульсы:
τ10_2 = 60 c, τ11_2 = 30 c, Δt10_2 = Δt11_2 = 3 c,
10_2
= I∗11_2 = 0,94 клк;
τ12_6 = τ9_6 = 0 c, Δt12_6 = Δt9_6 = 150 c,
12_6
= I∗9_6 = 1,88 клк,
τ10_10 = τ11_11 = 0 c, Δt10_10 = Δt11_11 = 60 c,
10_10
= I∗11_11 = 1,88 клк.
Возбуждающие импульсы:
τ5_6 = τ6_5 = 60 c, Δt5_6 = Δt6_5 = 15 c, τ1_3 = τ2_3 = τ5_7 = τ6_7 = 0 c,
Δt1_3τ8_Pro = τ8_Contra = 0 c, Δt8_Pro = Δt8_Contra = 15 c;
τ13_Pro = τ14_Contra = 0 c, Δt14_Contra = Δt13_Pro = 15 c;
τPro_Pro = τContra_Contra = 60 c, ΔtPro_Pro = ΔtContra_Contra = 15 c;
τPro_10 = 0 c, ΔtPro_10 = 15 c, τPro_12 = 0 c, ΔtPro_12 = 15 c,
τContra_11 = 0 c, ΔtContra_11 = 15 c, τContra_9 = 0 c, ΔtContra_9 = 15 c,
τ3_10 = 0 c, Δt3_10 = 15 c, τ3_9 = 0 c, Δt3_9 = 15 c, τ4_11 = 0 c,
Δt4_11 = 15 c, τ4_12 = 0 c, Δt4_12 = 15 c.
В блоке ЦГР период синфазных колебаний TIP = 38 c, а период противо-
фазных колебаний TAP = 44 c. Период противофазных колебаний в Антенне
TAP = 151 c. Период противофазных колебаний в паре осцилляторов № 13-
№ 14 T13_14 = 48 c.
145
Если ячейка № 4 определила новый противофазный ритм ЦГР, она отправ-
ляет возбуждающий импульс на ячейку № 12 блока Исполнитель. В момент
времени t= 1970 с одновременно с импульсом от ячейки № 4 на ячейку № 12
приходит импульс от ячейки “Pro”, вызывая в ней спайк. Возбужденная ячей-
ка № 12 отправляет ингибиторный импульс на ячейку № 6 Антенны. Этот
импульс переводит все ячейки Антенны в стационарное возбудимое состоя-
ние, в котором Антенна готова принимать новые внешние сигналы. Ячей-
ка № 12 посылает также ингибиторный импульс на ячейку “Pro”, переводя ее
в возбудимое состояние, что позволяет блоку ПР вновь принимать случайные
решения.
Аналогичный эксперимент при случайной активации ячейки “Contra” так-
же дал ожидаемый результат.
Оценим теперь численно вероятность активации ячеек “Pro” и “Contra”
при случайном приходе внешнего сигнала S на Антенну. Для этого мы про-
моделировали работу нашей сети с момента tS случайного прихода единич-
ного импульса S и до момента активации одной из ячеек “Pro” или “Contra”.
Условие “единичного импульса” означает, что в Антенне возбуждается про-
тивофазная мода и, следовательно, в Ридере РA всегда будет возбуждаться
ячейка № 8, настроенная на эту моду. Так как исходно все ячейки Антенны
и Ридера РA находятся в стационарном состоянии, то “случайность” прихода
сигнала S надо соотносить с начальной фазой ϕ колебаний пары осциллято-
ров № 13-№ 14.
Если tS - это время прихода внешнего импульса S на одну из ячеек Ан-
тенны, то tS + τS_8 - это время первой активации ячейки № 8 (= t80)), где
временной интервал τS_8 складывается из времени, необходимого для акти-
вации второй ячейки Антенны, и из времени, требуемого на приход двух им-
пульсов от ячеек Антенны на ячейку № 8. Конкретная величина времени τS_8
зависит от того, на какую ячейку, № 5 или № 6, приходит сигнал S. Из рис. 4
можно видеть, что время τS_8 = 190 c. Однако знание точного значения вели-
чины τS_8 не требуется для определения вероятности активации ячеек “Pro”
и “Contra”. Моменты первого и последующих спайков ячейки № 8, t(n)8, опре-
деляются как:
(1)
t(n)8 = t(0)8 + TAP
× n,
где n - целое число от нуля до бесконечности, TAP = 151 с (см. рис. 4). Пред-
положим, что в момент времени tS пара осцилляторов № 13-№ 14 находится
в произвольной фазе ϕ0, где фаза ϕ меняется от 0 до 1, а осциллятор № 13
производит спайк при ϕ = 0 и ϕ = 1 (при ϕ = 1 фаза обнуляется до ϕ = 0), а
осциллятор № 14 производит спайк при ϕ = 1/2 (так как осцилляторы № 13
и № 14 колеблются в противофазе). Тогда спайки в ячейке № 13 происходят
в следующие моменты времени t(m)13:
(2)
t(m)13 = tS + (1 - ϕ0 + m) × T13_14,
146
Pro
Сontra
0
0,2
0,4
0,6
0,8
1,0
0
Рис. 5. Зависимость исхода активации ячеек “Pro” и “Contra” от начальной
фазы колебаний ϕ0 пары осцилляторов № 13-№ 14. Черный цвет соответствует
активации ячейки “Pro”, а серый — активации ячейки “Contra”.
где m - целое число от 0 до бесконечности, а период T13_14 мы положили
равным 48 с, как было измерено в наших экспериментах. Моменты спайков
в ячейке № 14, t(m)14, можно задать через моменты t(m)13:
(3)
t(m)14 = t(m)13 - T13_14
/2.
Так как τ13_Pro = τ8_Pro = 0 с, то мы полагаем, что ячейка “Pro” должна
активироваться, если моменты времени t(m)13 и t(n)8 совпадают при каких-то
целых n и m с точностью до времени перекрытия импульсов активации, при-
чем мы учитываем только наименьшие значения n и m. Так как для акти-
вации ячейки “Pro” импульсы возбуждения могут перекрываться всего лишь
на время Δtx (= 5 c), то для активации ячейки “Pro” должно выполняться
следующее неравенство:
m)
(4)
t(
-t(n)8 < Δt - Δtx,
13
где Δt ≡ Δt8_Pro = Δt13_Pro (= 15 с). Если же выполняется условие:
m)
(5)
t(
-t(n)8 < Δt - Δtx,
14
причем раньше, чем условие (4), то должна активироваться ячейка “Contra”.
m)
Так как выражение
t(
-t(n)8 в (4) после подстановки в него выраже-
13
ний (1) и (2) преобразуется к виду |(1 - ϕ0 + m) × T13_14 - τS_8 - TAP × n|, в
котором уже нет времени tS, но есть начальная фаза ϕ0 (то же самое проис-
ходит и с разностью (t(m)14 - t(n)8), то при моделировании мы изменяли фазу ϕ0
от нуля до единицы. Заметим, что случайный момент времени tS определяет
начальную фазу ϕ0. На рис. 5 мы отмечаем черным цветом и ромбами исход
опыта, если активировалась ячейка “Pro”, и серым цветом и кружочками, ес-
ли активировалась ячейка “Contra”. Длины черной и серой линий оказались
равны друг другу, что доказывает, что активация ячеек “Pro” и “Contra” про-
исходит равновероятно. Такой же равновероятный исход должен быть и при
полностью случайной активации ячеек “Pro” и “Contra”. Однако наша “слу-
чайность” весьма специфична, а именно, существуют две компактные зоны
фаз ϕ0, в которых исход события предопределен, а вероятность попадания в
одну из этих зон одинакова.
147
5. Заключение
В данной статье мы продемонстрировали способ случайного принятия ре-
шения сетью импульсно связанных спайковых осцилляторов. Этот способ,
основанный на случайном совпадении во времени фаз или импульсов от ос-
цилляторов с разной частотой, не является, конечно, единственным. Можно,
например, использовать для принятия решения локальную подсеть из двух-
трех осцилляторов, которая дает хаотические колебания [27]. Такой способ,
вероятно, привел бы к более выраженной “случайности” при принятии ре-
шения. Однако нашей целью является разработка такого блока принятия
решений, который сможет осуществлять выбор решения не только случай-
ным образом, но и целенаправленно, учитывая полученный ранее опыт при
принятии решений. Для этих целей мы планируем разработать блок памя-
ти и синхронизировать его работу с другими блоками сети. Рассмотренный
же в данной статье метод случайного принятия решений является лишь пер-
вым шагом на пути создания “разумного” микроробота, который не всегда
работает по заданной программе.
Отметим, что полное математическое описание единичного БЖ микроос-
циллятора дается четырьмя дифференциальными уравнениями [18, 20]. Им-
пульсная связь с временной задержкой между двумя микроячейками также
описывается четырьмя дифференциальными уравнениями. Однако исполь-
зовать столь точный математический аппарат для описания происходящих в
сети событий совсем не обязательно. БЖ осцилляторы можно заменить фазо-
выми осцилляторами [18], а импульсное воздействие описывать при помощи
кривых переустановки фазы [26, 28]. Возбудимые ячейки легко заменяются
триггерными элементами с задаваемым порогом возбудимости. Временные
задержки можно реализовать при помощи химических (или электрохимиче-
ских) волн, распространяющихся с конечной скоростью по узким каналам
между заданными микроячейками [29]. Такие замены говорят о том, что
предложенный нами метод случайного принятия решений может быть уни-
версален и применим к другим спайковым сетям.
Мы благодарим И.Л. Мальфанова за приготовление БЖ микросфер и
микроэмульсии.
СПИСОК ЛИТЕРАТУРЫ
1. Klinshov V.V. Collective Dynamics of Networks of Active Units with Pulse Coupling:
Review
// Izvestiya Vysshikh Uchebnykh Zavedeniy-Prikladnaya Nelineynaya
Dinamika. 2020. V. 28. P. 465-490.
Клиньшов В.В. Коллективная динамика сетей активных элементов с импульс-
ными связями: Обзор // Изв. высш. уч. заведений. Прикладная нелинейная
динамика. 2020. Т. 28. № 5. С. 465-490.
2. Buzsaki G., Freeman W. Editorial overview: Brain rhythms and dynamic coordina-
tion // Current Opinion in Neurobiology. 2015. V. 31. P. V-IX.
3. Neves F.S., Timme M. Reconfigurable Computation in Spiking Neural Networks //
IEEE Access. 2020. V. 8. P. 179648-179655.
148
4.
Ryu H., Campbell S.A. Stability, Bifurcation and Phase-Locking of Time-Delayed
Excitatory-Inhibitory Neural Networks // Mathematical Biosciences and Engineer-
ing. 2020. V. 17. P. 7931-7957.
5.
Liang X., Zhang X.C., Xia J., Ezawa M., Zhao Y.L., Zhao G.P., Zhou Y. A Spiking
Neuron Constructed by the Skyrmion-Based Spin Torque Nano-oscillator // Appl.
Phys. Lett. 2020. V. 116. #122402.
6.
Spaeth A., Tebyani M., Haussler D., Teodorescu M. Spiking Neural state Machine
for Gait Frequency Entrainment in a Flexible Modular Robot // Plos One.
2020.
V. 15. P. e0240267.
7.
Kadhim K.L., Hermundstad A.M., Brown K.S. Structured Patterns of activity in
Pulse-coupled Oscillator Networks with Varied Connectivity // Plos One.
2021.
V. 16. P. e0256034.
8.
Bazhanova M.V., Krylova N.P., Kazantsev V.B., Khramov A.E., Lobov S.A. Syn-
chronization in a Network of Spiking Neural Oscillators with Plastic Connectivity //
Radiophysics and Quantum Electronics. 2020. V. 63. P. 298-309.
Бажанова М.В., Крылова Н.П., Казанцев В.Б., Храмов А.Е., Лобов С.А. Син-
хронизация в сети импульсных нейронных генераторов с пластичными связя-
ми // Изв. вузов. Радиофизика. 2020. Т. 63. № 4. С. 330-343.
9.
Afifurrahman, Ullner E., Politi A. Collective Dynamics in the Presence of Finite-
width Pulses // Chaos. 2021. V. 31. #043135.
10.
Galinsky V.L., Frank L.R. Collective Synchronous Spiking in a Brain Network of
Coupled Nonlinear Oscillators // Phys. Rev. Lett. 2021. V. 126. #158102.
11.
Gast R., Knosche T.R., Schmidt H. Mean-field Approximations of Networks of Spik-
ing Neurons with Short-term Synaptic Plasticity // Phys. Rev. E. 2021. V. 104.
#044310.
12.
Kuramoto Y. Phase- and Center-manifold Reductions for Large Populations of Cou-
pled Oscillators with Application to Non-locally Coupled Systems // Int. J. Bifur-
cation and Chaos. 1997. V. 7. P. 789-805.
13.
Velichko A., Putrolaynen V., Belyaev M. Higher-order and Long-range Synchro-
nization Effects for Classification and Computing in Oscillator-based Spiking neural
Networks // Neural Computing & Applications. 2021. V. 33. P. 3113-3131.
14.
Taylor A.F., Tinsley M.R., Wang F., Huang Z.Y., Showalter K. Dynamical Quorum
Sensing and Synchronization in Large Populations of Chemical Oscillators // Science.
2009. V. 323. P. 614-617.
15.
Belousov B.P. Collection of Short Papers on Radiation Medicine. Moscow: Medgiz,
1959. P. 145-152.
Белоусов Б.П. В кн. Сборник рефератов по радиационной медицине за 1958 г.
М.: Изд-во Медгиз, 1959. С. 145.
16.
Zhabotinsky A.M. Periodic liquid phase reactions // Proc. Acad. Sci. USSR. 1964.
V. 157. P. 392-395.
Жаботинский А.М. Периодические окислительные реакции в жидкой фазе //
Докл. АН СССР. 1964. Т. 157. № 2. С. 392-395.
17.
Proskurkin I.S., Smelov P.S., Vanag V.K. Experimental Verification of an Opto-
chemical “neurocomputer” // Phys. Chem. Chem. Phys. 2020. V. 22. P. 19359-
19367.
18.
Vanag V.K., Smelov P.S., Klinshov V.V. Dynamical Regimes of Four Almost Identi-
cal Chemical Oscillators Coupled via Pulse Inhibitory Coupling with Time Delay //
Phys. Chem. Chem. Phys. 2016. V. 18. P. 5509-5520.
149
19.
Feudel U., Pisarchik A.N., Showalter K. Multistability and Tipping. From Math-
ematics and Physics to Climate and Brain-Minireview and Preface to the Focus
Issue // Chaos. 2018. V. 28. #033501.
20.
Smelov P.S., Proskurkin I.S., Vanag V.K. Controllable Switching between Stable
Modes in a Small Network of Pulse-coupled Chemical Oscillators // Phys. Chem.
Chem. Phys. 2019. V. 21. P. 3033-3043.
21.
Кузнецов О.П. Ограниченная рациональность и принятие решений // Искус-
ственный интеллект и принятие решений. 2019. № 1. C. 3-15.
22.
Mysore S.P., Kothari N.B. Mechanisms of competitive selection: A canonical neural
circuit framework // Elife. 2020. V. 9. P. e51473.
23.
Taylor A.F., Tinsley M.R., Showalter K. Insights into Collective cell Behaviour from
Populations of Coupled Chemical oscillators // Phys. Chem. Chem. Phys. 2015.
V. 17. P. 20047-20055.
24.
Mallphanov I.L., Vanag V.K. Distance Dependent Types of Coupling of Chemical
Micro-oscillators Immersed in a Water-in-Oil Microemulsion // Phys. Chem. Chem.
Phys. 2021. V. 23. P. 9130-9138.
25.
De T.K., Maitra A. Solution behaviour of Aerosol OT in non-polar solvents // Adv.
Colloid Interface Sci. 1995. V. 59. P. 95-193.
26.
Proskurkin I.S., Vanag V.K. New Type of Excitatory Pulse Coupling of Chemical
Oscillators via Inhibitor // Phys. Chem. Chem. Phys. 2015. V. 17. P. 17906-17913.
27.
Stankevich N., Volkov E. Evolution of Quasiperiodicity in Quorum-sensing Coupled
Identical Repressilators // Chaos. 2020. V. 30. #043122.
28.
Canavier C.C., Achuthan S. Pulse Coupled Oscillators and the Phase Resetting
curve // Math. Biosci. 2010. V. 226. P. 77-96.
29.
Safonov D.A., Vanag V.K. Oscillatory Microcells Connected on a Ring by Chemical
Waves // Chaos. 2021. V. 31. #063134.
Статья представлена к публикации членом редколлегии О.П. Кузнецовым.
Поступила в редакцию 26.11.2021
После доработки 17.01.2022
Принята к публикации 26.01.2022
150