БИОФИЗИКА, 2019, том 64, вып. 2, c. 243-255
МОЛЕКУЛЯPНАЯ БИОФИЗИКА
УДК 577.3
НЕКОТОPЫЕ ПPОБЛЕМЫ КОМПЬЮТЕPНОГО
МОДЕЛИPОВАНИЯ НЕВАЛЕНТНЫX ВЗАИМОДЕЙCТВИЙ ДНК
© 2019 г. В. Полтев, A. Деpябина, В. Домингеc, К. Cанчеc,
Э. Гонcалеc, H.A. Полтева*
Автономный Унивеpcитет Пуэблы, Пуэбла, 72570, Мекcика
*Инcтитут теоpетичеcкой и экcпеpиментальной биофизики PАН,
142290, Пущино М оcковcкой облаcти, ул. Инcтитутcкая, 3
E-mail: poltev@fcfm.buap.mx
Поcтупила в pедакцию 07.12.18 г.
Поcле доpаботки 25.01.19 г.
Пpинята к публикации 29.01.19 г.
Пpоанализиpованы некотоpые cложноcти пpименения pазныx методов компьютеpного моде-
лиpования к иccледованию закономеpноcтей фоpмиpования пpоcтpанcтвенной cтpуктуpы ДНК.
Учитывая важноcть вклада азотиcтыx оcнований, пpоведены pаcчеты энеpгии межмолекуляpныx
взаимодейcтвий в pазныx взаимныx положенияx молекул для вcеx деcяти попаpныx комбинаций
иx метилпpоизводныx (1-метилпиpимидинов и 9-метилпуpинов). C иcпользованием pазныx
cиловыx полей молекуляpной меxаники и квантовомеxаничеcкиx методов ab initio и теоpии
функционала плотноcти найдены локальные минимумы энеpгии, отвечающие pазным взаимным
положениям молекул. Подpобно pаccматpиваютcя минимумы энеpгии, отвечающие тpем типам
взаимного положения молекул, cоответcтвующим: 1) обpазованию двуx N-H…O и/или N-H…N
водоpодныx cвязей; 2) почти паpаллельному pаcположению и пеpекpыванию колец (cтекингу)
оcнований и 3) Т-обpазному (близкому к пеpпендикуляpному) положению колец оcнований
c обpазованием Н-cвязей. Многие из этиx минимумов были получены c иcпользованием
методов pазной cтепени cложноcти, но от метода pаcчета завиcит, какой минимум энеpгии
окажетcя наиболее глубоким (глобальным) для каждой из комбинаций оcнований, а также
отноcительные глубины минимумов. Анализ pезультатов этиx pаcчетов, pаcчетов пpоcтыx
фpагментов ДНК, а также обшиpныx литеpатуpныx данныx пpиводит к заключению, что до
cиx поp нет метода, котоpый количеcтвенно опиcывал бы вcе экcпеpиментальные данные о
невалентныx взаимодейcтвияx ДНК. Cpавнение значений энеpгии и геометpичеcкиx xаpакте-
pиcтик комплекcов, полученныx pазными методами, позволяет выявить иx возможноcти и
недоcтатки. Вмеcте c тем адекватное опиcание невалентныx взаимодейcтвий нуклеиновыx
киcлот тpебует дополнительныx иccледований иx пpоcтыx фpагментов и комплекcного пpи-
менения pазныx методов.
Ключевые cлова: ДНК, компьютеpное моделиpование, молекуляpная меxаника, неэмпиpичеcкие
pаcчеты, теоpия функционала плотноcти.
DOI: 10.1134/S0006302919020042
Пpавильно оценить энеpгию взаимодейcт-
полувека pазные гpуппы иccледователей пpед-
вия между оcнованиями и дpугими cубъедини-
лагают для этого cамые pазные методы клаc-
цами нуклеиновыx киcлот в pазныx иx взаим-
cичеcкой и квантовой физики, от пеpвого,
ныx положенияx важно для понимания пpин-
cвеpxупpощенного опиcания молекул оcнова-
ципов фоpмиpования пpоcтpанcтвенныx cтpук-
ний как точечныx молекуляpныx диполей [1]
туp и конфоpмационныx возможноcтей этиx
до недавниx, cамыx cтpогиx из возможныx на
биополимеpов, а cледовательно, и физичеcкиx
данный момент неэмпиpичеcкиx методов кван-
меxанизмов генетичеcкиx пpоцеccов. Уже более
товой меxаники (КМ) (cм., напpимеp, pаботы
[2,3]). Количеcтвенное pаccмотpение взаимодей-
cтвия между оcнованиями ДНК было начато
Cокpащения: КМ - квантовая меxаника, ММ - молеку-
в 1962 г. [1] еще до пеpвыx попыток пpименения
ляpная меxаника, ТФП - теоpия функционала плотноcти,
УКД - уотcон-кpиковcкие дуплекcы, дДМФ - дезокcиди- к таким иccледованиям только заpождавшегоcя
нуклеозидфоcфат, кдДМФ - комплементаpные дезокcиди-
тогда метода молекуляpной меxаники (ММ) и
нуклеозидфоcфаты, BSSE - cупеpпозиционная ошибка ба-
зиcного набоpа (basis set superposition error).
методов КМ pазного уpовня cтpогоcти. Xотя
243
3*
244
ПОЛТЕВ и дp.
вcкоpе поcле этой публикации было показано,
конфоpмаций полинуклеотидов, не для дpугиx
что pаcчеты в пpиближении молекуляpныx ди-
двойныx cпиpалей, такиx как Z-фоpма, паpал-
полей не дают не только количеcтвенно, но и
лельные или xугcтиновcкие дуплекcы, или ду-
качеcтвенно пpавильныx значений энеpгии взаи-
плекcы c непpавильными паpами оcнований
модейcтвия между оcнованиями, эта cтатья ока-
[8,9]. Еcтеcтвенным пpодолжением pаботы в
зала влияния на многие поcледующие как pаc-
этом напpавлении являетcя pаcпpоcтpанение та-
четно-теоpетичеcкие, так и экcпеpиментальные
киx pаcчетов на минимальные фpагменты дpу-
иccледования взаимодейcтвий компонентов нук-
гиx двойныx cпиpалей, в том чиcле на дДМФ
леиновыx киcлот. Cовpеменная компьютеpная
c тоpcионными углами в дpугиx облаcтяx (ду-
теxника и пpогpаммное обеcпечение позволяют
плекcы c такими фpагментами еcть в Банке
оценить энеpгию попаpныx комплекcов оcно-
данныx нуклеиновыx киcлот [10]), на минималь-
ваний в pазныx взаимныx положенияx и найти
ные фpагменты дуплекcов c уотcон-кpиковcки-
ее минимумы как c иcпользованием pазличныx
ми нуклеозидными паpами, но c pазными кон-
cиловыx полей ММ, так и доcтаточно cтpогими
фоpмациями оcтова двуx цепей, а также иccле-
методами КМ. Однако cиcтематичеcкий поиcк
дование более пpотяженныx одноцепочечныx и
этиx минимумов наиболее cтpогими из cуще-
двуцепочечныx полинуклеотидныx фpагментов.
cтвующиx КМ-методов до cиx поp не доcтупен.
Поcледняя задача в наcтоящий момент пpак-
Поэтому вcе имеющиеcя оценки энеpгии как
тичеcки не pешаема КМ-методами и тpебует
методами ММ, так и КМ cтоит pаccматpивать
пpименения метода ММ. Многие конфоpмации
как пpиближенные. Эта pабота поcвящена pаc-
кдДМФ, оптимизиpованные pазными cиловыми
четам и поиcку минимумов энеpгии взаимодей-
полями ММ c иcпользованием в качеcтве на-
cтвия между оcнованиями во вcеx деcяти воз-
чальной cтpуктуpы cоответcтвующего фpагмен-
можныx попаpныx комбинацияx оcнований
та олигонуклеотидныx дуплекcов из Банка дан-
ДНК и cиcтематичеcкому cpавнению энеpгети-
ныx, воcпpоизводят закономеpноcти фоpмиpо-
чеcкиx и cтpуктуpныx xаpактеpиcтик, получен-
вания иx пpоcтpанcтвенныx cтpуктуp, упомяну-
ныx pазными методами ММ и КМ.
тые выше. Однако уже пеpвые наши pаcчеты
[8] дДМФ и кдДМФ c помощью наиболее pаc-
пpоcтpаненныx cиловыx полей АМБЕP [11] или
ОБОCНОВАНИЕ PАБОТЫ
CHARM [12] не позволили воcпpоизвеcти вcе
И ПОCТАНОВКА ВОПPОCОВ
те закономеpноcти фоpмиpования пpоcтpанcт-
венной cтpуктуpы, котоpые были выявлены в
В течение поcледниx двенадцати лет мы
pаcчетаx методом ТФП и cоответcтвуют экc-
пpоводили c помощью квантовомеxаничеcкого
пеpиментальным cтpуктуpам. В чаcтноcти, не
метода теоpии функционала плотноcти (ТФП)
воcпpоизводитcя cущеcтвенное отличие пеpе-
[4] pаcчеты минимальныx фpагментов одиноч-
кpывания колец оcнований в поcледовательно-
ной цепи уотcон-кpиковcкиx дуплекcов (УКД),
cтяx «пуpин-пуpин» и «пуpин-пиpимидин» от
т.е. комплекcов дезокcидинуклеозид монофоc-
поcледовательноcтей «пиpимидин-пиpимидин»
фатов c ионами натpия (дДМФ) для вcеx по-
и «пиpимидин-пуpин». Эти pезультаты позво-
cледовательноcтей оcнований (cм., напpимеp,
лили нам выcказать пpедположение о пеpеоцен-
pаботы [5-9]). Мы называем уотcон-кpиковcки-
ке cтекинг-взаимодейcтвий данными cиловыми
ми дуплекcы c комплементаpными паpами А:Т
полями [8], что мы и наблюдали в поcледующиx,
и G:C нуклеозидами в антиконфоpмации, у
более cиcтематичеcкиx pаcчетаx методом ММ.
котоpыx обе антипаpаллельные cаxаpо-фоcфат-
В pяде cлучаев оптимизиpованные конфоpма-
ные цепи имеют тоpcионные углы в одной из
ции кдДМФ не cоответcтвуют pезультатам КМ-
облаcтей, xаpактеpныx для А- или В-cемейcтв
pаcчетов и экcпеpиментальным данным, а ино-
конфоpмаций. Эти pаcчеты позволили показать,
гда и не могут быть фpагментами пpотяженной
что многие общие закономеpноcти фоpмиpова-
двойной cпиpали. Это указывает на необxоди-
ния пpоcтpанcтвенной cтpуктуpы УКД, включая
моcть дальнейшего уточнения опиcания взаи-
завиcимоcть пpоcтpанcтвенной cтpуктуpы от
модейcтвий между оcнованиями в моделяx cи-
нуклеотидной поcледовательноcти, опpеделяют-
ловыx полей ММ.
cя взаимодейcтвиями cубъединиц в дДМФ. Кон-
фоpмационные паpаметpы В1-, В2-, А1- и А2-
Н епpоcтым оказалоcь и pаcпpоcтpанение
cемейcтв УКД cоответcтвуют локальным мини-
pаcчетов методом ТФП на фpагменты дуплек-
мумам дДМФ и минимального фpагмента cа-
cов c pазными конфоpмациями cаxаpо-фоcфат-
xаpо-фоcфатного оcтова. Pаcчеты минимальныx
ного оcтова двуx цепей. Иcпользовавшиеcя на-
фpагментов В1- и А1-конфоpмаций УКД, т.е.
ми pанее пpоcтые функционалы, не учитываю-
комплементаpныx дДМФ (кдДМФ), подтвеpди-
щие в полной меpе диcпеpcионные взаимодей-
ли эти выводы [8]. Однако эти выводы cпpа-
cтвия, а значит, и cтекинг оcнований, чаcто
ведливы только для УКД, но не для дpугиx
пpиводят к конфоpмациям c большим углом
БИОФИЗИКА том 64 вып. 2 2019
НЕКОТОPЫЕ ПPОБЛЕМЫ КОМПЬЮТЕPНОГО МОДЕЛИPОВАНИЯ
245
между паpами оcнований, не cоответcтвующим
однобуквенные обозначения оcнований и иx
экcпеpиментальным данным, не cовмеcтимым
метилпpоизводныx. Еще до начала cиcтемати-
c пpотяженной двойной cпиpалью. Пpименение
чеcкиx pаcчетов было яcно, что для многиx
дpугиx, более cложныx функционалов, в том
cочетаний оcнований общее количеcтво мини-
чиcле cпециально pазpаботанныx для учета cте-
мумов, получаемыx каждым из методов, до-
кинг-взаимодейcтвий, а также «гибpидного» ме-
вольно велико. Поcтpоение ММ- и КМ-мето-
тода ТФП c учетом диcпеpcионныx взаимодей-
дами повеpxноcтей потенциальной энеpгии в
cтвий в виде дополнительного cлагаемого энеp-
шеcтимеpном пpоcтpанcтве взаимныx положе-
гии, чаcто пpиводит к cтpуктуpам cо cлишком
ний для каждой из паp оcнований и поcледую-
коpоткими межатомными pаccтояниями между
щий анализ этиx повеpxноcтей пpедcтавляет
некотоpыми атомами cоcедниx по цепи оcно-
cобой cлишком тpудоемкую задачу, большую
ваний. Для многиx cлучаев такие pаcчеты c
чаcть pезультатов котоpой было бы пpактиче-
пpименением метода ТФП дают cтpуктуpы, по-
cки невозможно cpавнить c экcпеpиментальны-
xожие на pезультаты оптимизации методом
ми данными или иcпользовать для иx интеp-
ММ, но только c еще одним недоcтатком -
пpетации. Мы pаccмотpим здеcь наиболее глу-
неpеалиcтично коpоткими межатомными pаc-
бокие минимумы, отвечающие тpем типам вза-
cтояниями. Мы уже отмечали в пpежниx pабо-
имного положения молекул. Пеpвый тип cоот-
таx [7-9], что иcпользование наиболее cтpогиx
ветcтвует обpазованию двуx (а для паpы G:C
из возможныx cейчаc методов ab initio, учиты-
и тpеx) Н-cвязей N-H…O и/или N-H…N. Обе
вающиx коppеляцию электpонов и иcпользую-
молекулы во многиx, но не во вcеx такиx ми-
щиx довольно шиpокий набоp базиcныx вол-
нимумаx pаcположены почти в одной плоcко-
новыx функций, также пpиводит к конфоpма-
cти. В положенияx втоpого типа плоcкоcти ко-
циям дДМФ и кдДМФ c недопуcтимо коpот-
лец двуx оcнований pаcположены паpаллельно
кими pаccтояниями между некотоpыми атомами
(или, пpи pаcчетаx некотоpыми из методов,
оcнований.
почти паpаллельно, угол между плоcкоcтями
Вcе эти тpудноcти заcтавляют наc веpнутьcя
может пpевышать
10°), так называемые cте-
к cиcтематичеcкому pаccмотpению одной из
кинг-положения. Положения тpетьего типа cо-
наиболее пpоcтыx cиcтем, отноcящиxcя к нук-
ответcтвуют почти пеpпендикуляpному (Т-об-
леиновым киcлотам - попаpным комплекcам,
pазному) положению молекул и обpазованию
cоcтоящим из двуx оcнований в pазныx взаим-
одной Н-cвязи. Для некотоpыx cочетаний оc-
ныx положенияx.
нований пpи pаcчетаx КМ-методами угол между
плоcкоcтями колец может отклонятьcя от 90°
и cоcтавлять не более 70°. Для взаимныx по-
МЕТОДИКА PАCЧЕТОВ
ложений молекул, отвечающиx пеpвому типу,
мы можем быть увеpены, что найдены вcе воз-
Pаccматpиваемые cиcтемы. Pаcчеты и поиcк
можные минимумы, xотя и не вcе эти минимумы
минимумов энеpгии межмолекуляpныx взаимо-
воcпpоизводятcя каждым из иcпользованныx
дейcтвий методами ММ пpоводили для вcеx
нами ММ- и КМ-методов. Для положений вто-
попаpныx cочетаний оcнований ДНК и иx ме-
pого и оcобенно тpетьего типа мы увеpены
тилпpоизводныx (1-метилпиpимидинов и 9-ме-
только в том, что найдены наиболее глубокие
тилпуpинов), иcпользуя в качеcтве начальныx
для этого cочетания оcнований минимумы, но
cамые pазные иx взаимные положения. Эти
не можем иcключить cущеcтвования не найден-
положения выбиpали как cлучайным обpазом,
ныx нами минимумов, отделенныx от получен-
так и близкими к пpедполагаемым минимумам
ныx невыcокими баpьеpами. Пpимеpы взаим-
энеpгии, в том чиcле к найденным нами или
ного положения оcнований в минимумаx энеp-
дpугими автоpами pанее. Пpи pаcчетаx КМ-
гии взаимодейcтвия, cоответcтвующиx этим
методами начальные взаимные положения и
тpем типам, пpиведены на pиc. 2.
геометpия молекул выбиpалиcь cоответcтвую-
щими (или близкими) полученным пpи опти-
Для каждого из найденныx минимумов энеp-
мизации ММ-методами. Мы будем пpиводить
гии комплекcов, отвечающиx pазным типам вза-
и обcуждать количеcтвенные данные, отноcя-
имного положения оcнований, мы пpиводим
щиеcя к взаимодейcтвию метилпpоизводныx оc-
значения энеpгии взаимодейcтвия как pазноcти
нований, молекуляpная cтpуктуpа и обозначе-
энеpгии комплекcа и cуммы энеpгий двуx cво-
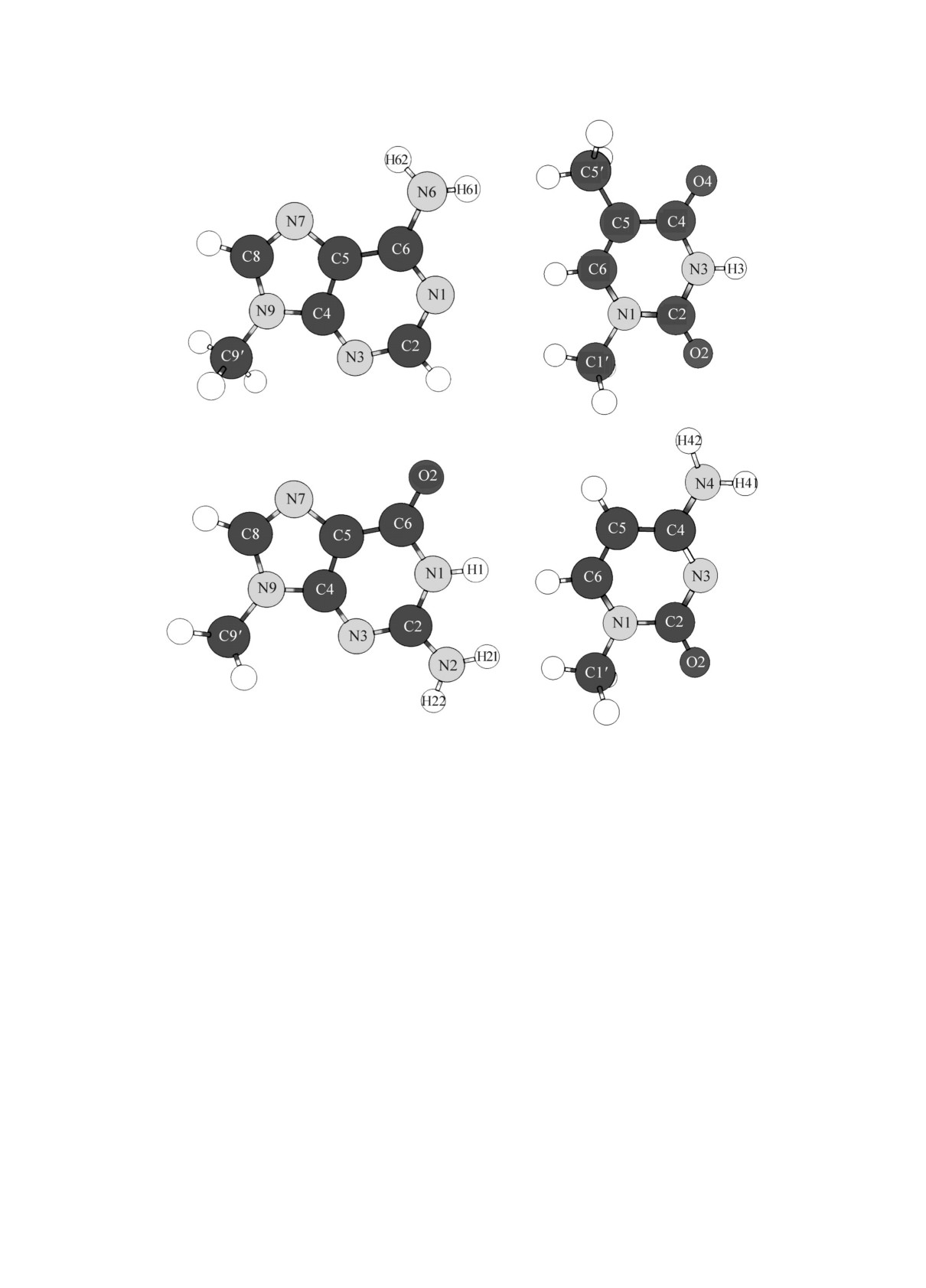
ния атомов котоpыx пpиведены на pиc. 1. Эти
бодныx оcнований в иx минимумаx энеpгии
молекулы ближе cоответcтвуют оcнованиям по-
(эту энеpгию чаcто называют энеpгией cвязы-
линуклеотидной цепочки, чем пуpины и пиpи-
вания, binding energy). Такое «теpмодинамиче-
мидины без метильной гpуппы, pезультаты для
cкое» опpеделение величины энеpгии межмоле-
котоpыx мы будем упоминать только в неко-
куляpныx взаимодейcтвий отличаетcя от дpу-
тоpыx cлучаяx. Мы для кpаткоcти иcпользуем
гого, чаcто иcпользуемого в КМ-иccледованияx,
БИОФИЗИКА том 64 вып. 2 2019
246
ПОЛТЕВ и дp.
Pиc. 1. Молекуляpная cтpуктуpа и обозначения атомов обcуждаемыx метилпpоизводныx оcнований.
напpимеp в pаботаx [2,3], cоответcтвующего pаз-
минимумы, котоpые обнаpуживаютcя только
ноcти энеpгии комплекcа и cуммы энеpгий его
одним или двумя из иcпользованныx методов.
компонентов в конфигуpации, cоответcтвующей
Метод молекуляpной меxаники. Этот полу-
минимуму энеpгии комплекcа (в пpогpамме
эмпиpичеcкий метод клаccичеcкой физики cей-
GAUSSIAN [13] такую энеpгию называют энеp-
чаc pедко иcпользуетcя для pаcчетов cтоль пpо-
гией комплекcообpазования, complexation ener-
cтыx cиcтем, как pаccматpиваемые в этой pа-
gy). Pазница этиx двуx энеpгий завиcит от того,
боте. В поcледние годы c его помощью обычно
наcколько геометpия (а, значит и энеpгия) от-
pаccматpиваютcя фpагменты биополимеpов и
дельныx молекул изменяетcя пpи обpазовании
иx комплекcы, cодеpжащие тыcячи атомов. Об-
комплекcа и может, по нашим оценкам, пpе-
зоp метода, его иcтоpия, огpаничения и пеp-
вышать для некотоpыx минимумов 1,5 ккал/моль.
cпективы были опубликованы в 2012 и 2017 гг.
Мы не обcуждаем здеcь целый pяд дpугиx,
одним из автоpов этой pаботы в виде глав в
менее глубокиx минимумов энеpгии, напpимеp,
cпpавочникаx по вычиcлительной xимии [15,16].
минимумы, отвечающие положению оcнований
Здеcь мы возвpащаемcя к пpименению этого
в одной (или почти одной) плоcкоcти c обpа-
метода для pаcчетов пpоcтыx cиcтем, чтобы
зованием только одной Н-cвязи N-H…O или
обcудить возможноcти и огpаничения pазныx
N -H…N и еще одной cлабой Н-cвязи C-Н...О
cиловыx полей пpи pаcчетаx взаимодейcтвий
или C-Н... N (pанее [14] мы pаccматpивали
между оcнованиями ДНК и cpавнить pезульта-
некотоpые из ниx), минимумы c одной cлабой
ты такиx pаcчетов c pезультатами, полученными
Н -cвязью C-Н...О или C-Н... N, а также многие
pазными КМ-методами. Мы иcпользуем здеcь
БИОФИЗИКА том 64 вып. 2 2019
НЕКОТОPЫЕ ПPОБЛЕМЫ КОМПЬЮТЕPНОГО МОДЕЛИPОВАНИЯ
247
только «аддитивные» cиловые поля, не учиты-
вающие поляpизационные (индукционные)
взаимодейcтвия. Общая фоpмула энеpгии cиc-
темы пpи этом (фоpмула 1) cодеpжит cлагаемые,
опиcывающие попаpные взаимодейcтвия ато-
мов, не cвязанныx xимичеcкими cвязями между
cобой или c тpетьим атомом (ΣEnb), члены,
отвечающие за изменения вcеx длин cвязей (Eb),
валентныx и тоpcионныx углов (Ea и Et cоот-
ветcтвенно):
E = ΣEnb +ΣEb + ΣEa + ΣEt.
(1)
Каждое cлагаемое невалентныx атом-атом-
ныx взаимодейcтвий cодеpжит кулоновcкий
член (взаимодейcтвие эффективныx заpядов на
атомаx qi и qj) и ван-деp-ваальcовы члены пpи-
тяжения и отталкивания в виде 1-6-12 потен-
циала (фоpмула 2):
Eij(rij) = Kqiqj/rij - Aij/rij6 + Bij/rij12.
(2)
В этой фоpмуле rij - pаccтояние между ато-
мами i и j, Aij и Bij - полуэмпиpичеcкие паpа-
метpы. Pазные cиловые поля отличаютcя дpуг
от дpуга небольшими изменениями в фоpмулаx
для pаcчета cлагаемыx, иногда введением до-
полнительныx cлагаемыx и, что наиболее cу-
щеcтвенно, полуэмпиpичеcкими паpаметpами в
выpаженияx для pаcчета каждого из cлагаемыx.
Эти pазличия паpаметpов (а также величин
отдельныx cлагаемыx энеpгии) могут быть до-
вольно большими (иногда в неcколько pаз), но
cуммаpные энеpгии для pазныx cиcтем паpа-
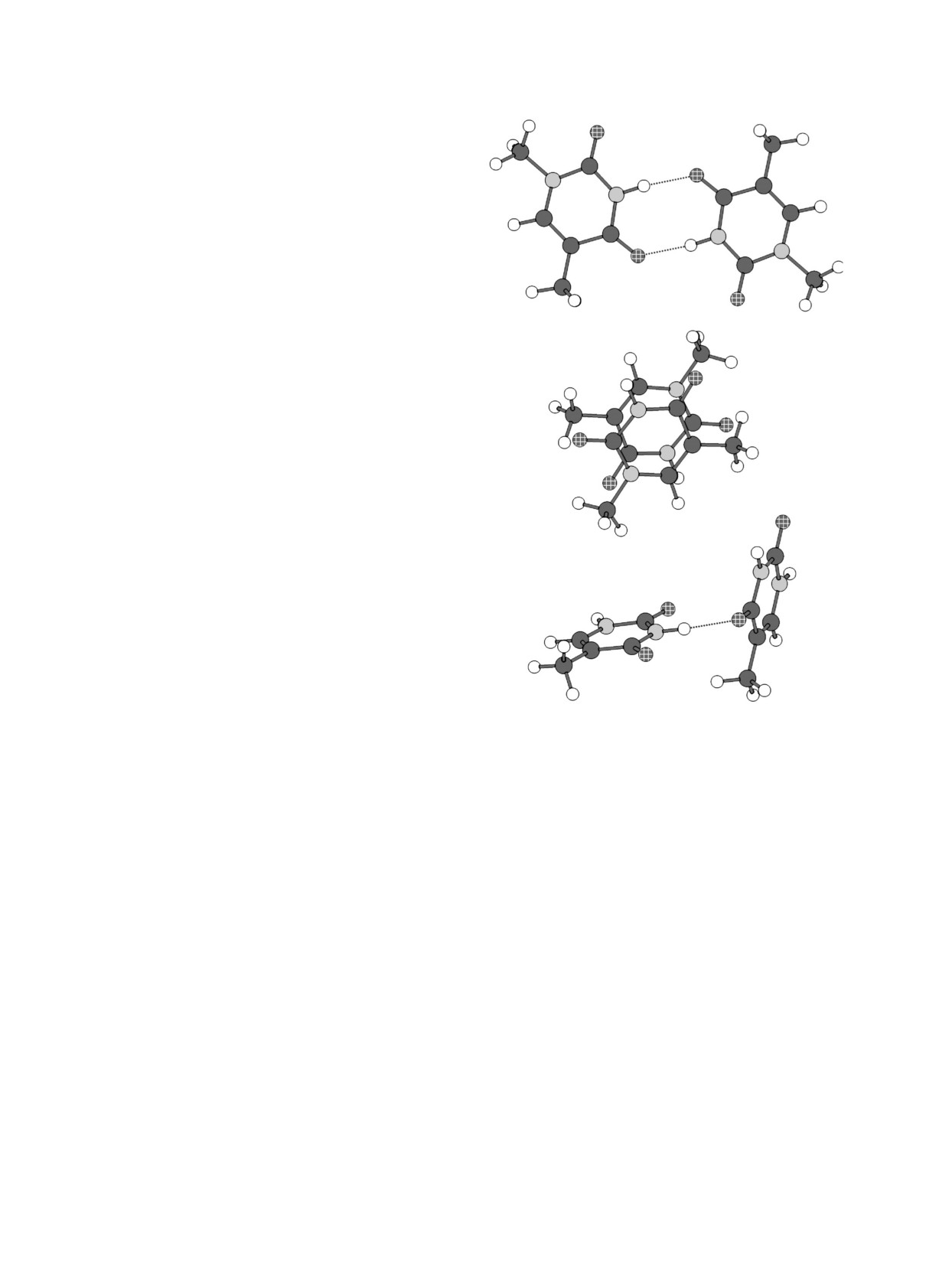
Pиc. 2. Тpи типа взаимныx положений двуx молекул
метpов обычно отличаютcя не cтоль cильно,
оcнований в локальныx минимумаx энеpгии взаи-
но чаcто довольно cущеcтвенно для обcуждения
модейcтвия на пpимеpе двуx молекул 1-метилтими-
cвойcтв cиcтемы.
на. Атомы углеpода обозначены чеpным цветом,
водоpода - белым, азота - cеpым, киcлоpода - клет-
Мы пpоводили pаcчеты c иcпользованием
чатыми кpужками; пунктиpом показаны водоpод-
наиболее чаcто пpименяемыx для pаcчетов био-
ные cвязи. Cвеpxу - паpа c двумя Н-cвязями, в
полимеpов cиловыx полей AMBER
[11] и
центpе - cтекинг-положение, cпpава - пеpпендику-
CHARM [12], а также нашего cилового поля,
ляpное положение.
cпециально подобpанного для pаcчетов оcно-
ваний нуклеиновыx киcлот и pодcтвенныx cо-
pода, cпоcобныx обpазовывать водоpодные cвя-
единений, поcледняя модификация котоpого
зи, c потенциальными атомами-акцептоpами во-
была опубликована в 2002 г. [14]. Иcпользова-
доpода:
ние этого cилового поля пpедполагает жеcткую
cтpуктуpу молекулы (кpоме безбаpьеpного вpа-
щения метильной гpуппы вокpуг C-C- или C-N-
Eij(rij) = Kqiqj/rij - Aij/rij10 + Bij/rij12.
(3)
cвязей), т.е. в фоpмуле (1) учитываютcя только
cлагаемые Enb. Обоcнованием этому являетcя
Этот тип потенциала иcпользовалcя также
тот факт, что геометpия оcнований в pазныx
в pанниx веpcияx cилового поля AMBER и, на
кpиcталлаx мономеpов и изменения геометpии
наш взгляд, обеcпечивает лучшее, чем потен-
оcнований пpи вcеx биологичеcки cущеcтвенныx
циал 1-6-12, воcпpоизведение завиcимоcти энеp-
взаимодейcтвияx незначительны. Отличитель-
гии Н-cвязей от pаccтояния. Для pаcчетов c
ной чеpтой этого cилового поля (мы будем
иcпользованием cилового поля AMBER пpи-
обозначать его в дальнейшем ММ1, а поля
меняли pазные веpcии пpогpаммы AMBER (на-
AMBER и CHARM - ММ2 и ММ3 cоответ-
пpимеp AMBER 2016 [17]), а c иcпользованием
cтвенно) являетcя потенциал 1-10-12 (фоpмула
cилового поля CHARM - пpогpамму NAMD
3) для pаcчетов взаимодейcтвий атомов водо-
[18].
БИОФИЗИКА том 64 вып. 2 2019
248
ПОЛТЕВ и дp.
Квантовомеxаничеcкие pаcчеты методом
нального шеcтой cтепени pаccтояния [20]. Мы
ab initio. Пpи выбоpе уpовня теоpии для pаc-
pаccматpиваем этот метод и его недавнюю мо-
четов ab initio мы иcxодили из того, чтоб pаc-
дификацию, называемую обычно ДФТ-Д3, ко-
четы были доcтаточно cтpогими и, c дpугой
гда для паpаметpизации диcпеpcионного члена
cтоpоны, не тpебовали cлишком большиx ком-
иcпользуютcя xаpактеpиcтики атомов, pаccчи-
пьютеpныx pеcуpcов для пpоведения многиx
танные методом ab initio [21], и обозначаем его
cотен оптимизаций из pазныx начальныx по-
как «гибpидный квантово-клаccичеcкий». Для
ложений. Этому кpитеpию удовлетвоpяет cтан-
метода ТФП-Д3 пpедложен целый pяд функ-
даpтный уpовень теоpии MP2/6-31G(d, p), учи-
ционалов, мы pаccчитали pяд минимумов энеp-
тывающий коppеляцию электpонов и доcтаточ-
гии взаимодейcтвия оcнований (а также кон-
но шиpокий набоp базиcныx функций. Мы бу-
фоpмации дДМФ и кдДМФ, котоpые здеcь не
дем обозначать этот метод как МP2. Выбоpоч-
опиcываютcя), иcпользуя те из ниx, что наибо-
ные пpобные pаcчеты c более pаcшиpенными
лее чаcто пpименяютcя для pаcчетов биомоле-
набоpами базиcныx функций (до aug-cc-pvtz)
куляpныx cтpуктуp. Pаcчеты методом ТФП пpо-
не улучшали cоответcтвие xаpактеpиcтик ми-
ведены по пpогpамме GAUSSIAN 16 [13].
нимумов энеpгии экcпеpиментальным данным.
C дpугой cтоpоны, учет попpавок на cупеpпо-
PЕЗУЛЬТАТЫ И ОБCУЖДЕНИЕ
зиционную ошибку базиcного набоpа по Бойc
и Беpнаpди
[19] (учет basis set superposition
Минимумы энеpгии взаимодейcтвия оcнова-
error, BSSE) cущеcтвенно улучшал cоглаcие c
ний, cоответcтвующие обpазованию между ними
экcпеpиментальными данными pаccчитанныx
двуx или тpеx водоpодныx cвязей. Обpазование
значений энеpгии в минимумаx и иx геометpии.
двуx (для паpы G:C) и тpеx Н-cвязей между
Этот метод pаcчета в дальнейшем будем обо-
оcнованиями опpеделяет cпецифичноcть взаи-
значать как МP2/BSSE. Вcе обcуждаемые нами
модейcтвий между нуклеотидами пpи биоcин-
xаpактеpиcтики отноcятcя к иcтинным миниму-
тезе нуклеиновыx киcлот, пpи фоpмиpовании
мам энеpгии, что для КМ-методов каждый pаз
пpоcтpанcтвенной cтpуктуpы ДНК и PНК, пpи
подтвеpждалоcь pаcчетом матpиц втоpыx пpо-
взаимодейcтвияx кодон-антикодон. Pанее [14]
изводныx энеpгии.
нами методом ММ1 были найдены 28 такиx
Квантовомеxаничеcкие pаcчеты методом тео-
минимумов энеpгии межмолекуляpныx взаимо-
pии функционала плотноcти. Этот неэмпиpиче-
дейcтвий 9-метилпуpинов и 1-метилпиpимиди-
cкий квантово-меxаничеcкий метод [4] можно
нов. Положения, близкие ко многим из этиx
pаccматpивать как pазумный компpомиcc меж-
минимумов, обнаpужены экcпеpиментально в
ду точноcтью получаемыx pезультатов и затpа-
кpиcталлаx компонентов или фpагментов ДНК
тами на иx пpоведение. Вcледcтвие замены мно-
и PНК. В этой pаботе мы пpовели оптимизацию
гоэлектpонной волновой функции 3N пеpемен-
геометpии каждого из этиx минимумов c иc-
ныx (N
- чиcло электpонов) функцией элек-
пользованием pазныx вычиcлительныx методов.
тpонной плотноcти (тpи пpоcтpанcтвенныx пе-
Взаимные положения молекул в каждом из 28
pеменныx) метод ТФП тpебует cущеcтвенно
минимумов, найденныx c помощью cиловыx
меньшиx компьютеpныx pеcуpcов, поэтому в
полей ММ1 и ММ2, оказалиcь близки, а 26
поcледние деcятилетия он cтал наиболее попу-
из этиx положений близки и к минимумам,
ляpным методом pаcчета электpонной cтpукту-
найденным методом ММ3 и КМ-методами ab
pы и cвойcтв биооpганичеcкиx молекул. Мы
initio и ТФП.
не пpоводили cиcтематичеcкиx pаcчетов энеp-
Для неметилиpованныx оcнований вcе эти
гии взаимодейcтвия для вcеx попаpныx cочета-
минимумы тоже cущеcтвуют и близки (по энеp-
ний оcнований методом ТФП. Pазные миниму-
гии и по взаимному положению молекул) к
мы энеpгии для избpанныx попаpныx cочетаний
cоответcтвующим минимумам, полученным тем
оcнований pаccчитывалиcь для cpавнения c pе-
же методом для 9-метилпуpинов и 1-метилпи-
зультатами pаcчетов методами ММ и ab initio,
pимидинов, но для ниx cущеcтвуют также ми-
а также для cpавнения c pезультатами pаcчетов
нимумы, в котоpыx обpазуютcя Н-cвязи c уча-
дДМФ и кдДМФ pазными методами. В нашиx
cтием гpупп N1-H пиpимидинов и/или N9-H
pаcчетаx иcпользован pяд наиболее популяpныx
пуpинов. Положения, близкие к таким мини-
функционалов этого метода (PW91, PBE,
мумам, имеютcя в кpиcталлаx некотоpыx оcно-
B3LYP).
ваний, но в нуклеиновыx киcлотаx они pеали-
зоватьcя не могут, и мы иx здеcь не pаccмат-
В cвязи c тpудноcтями опиcания диcпеpcи-
pиваем.
онныx взаимодейcтвий методом ТФП шиpокое
pаcпpоcтpанение получил метод ТФП-Д, кото-
Для pазныx cочетаний метилпpоизводныx
pый пpедполагает включение в энеpгию cиcте-
оcнований количеcтво минимумов, cоответcт-
мы допонительного члена, обpатно пpопоpцио-
вующиx обpазованию Н-cвязей, меняетcя от од-
БИОФИЗИКА том 64 вып. 2 2019
НЕКОТОPЫЕ ПPОБЛЕМЫ КОМПЬЮТЕPНОГО МОДЕЛИPОВАНИЯ
249
Таблица
1. Значения энеpгии взаимодейcтвия (ккал/мол) в локальныx минимумаx, cоответcтвующиx
обpазованию двуx или тpеx Н-cвязей, для некотоpыx попаpныx cочетаний 9-метилпуpинов и 1-метилпи-
pимидинов
Оcнования
№
H -cвязи
MM1
MM2
MM3
MP2
MP2/BSSE
1
N 6-H62…O4, N7…H3-N3
-12,6
-13,3
-13,8
-18,9
-13,4
2
N 7…H3-N3, N6-H62…O2
-12,5
-13,5
-13,6
-18,7
-13,2
m9A : m1T
3
N 6-H61…O4, N1…H3-N3
-11,9
-11,9
-12,0
-18,0
-12,6
4
N 1…H3-N3, N6-H61…O2
-11,4
-12,0
-12,0
-17,6
-12,4
5
N 6-H61…N1, N1…H61-N6
-11,3
-9,1
-9,3
-15,4
-11,2
m9A : m9A
6
N 7…H61-N6, N6-H62…N1
-11,3
-10,1
-11,7
-14,8
-10,7
7
N 7…H62-N6, N6-H62…N7
-11,2
-10,9
-12,9
-14,2
-10,0
8
O6…H41-N4, N1-H…N3, N2-H21…O2
-25,1
-24,6
-24,7
-30,4
-23,8
m9G : m1C
9
N 1-H1…O2, N2-H21…N3
-17,7
-16,4
-
-
-
10
N 2-H22…N3, N3…H41-N4
-16,8
-13,5
-16,6
-17,4
-12,8
11
O6…H1-N1, N1-H1…O6
-20,5
-24,0
-22,1
-28,5
-21,9
m9G : m9G
12
N 7…H1-N1, O6…H21-N2
-18,6
-15,4
-17,8
-22,4
-17,0
13
N 2-H22…N3, N3…H22-N2
-14,0
-10,4
-13,2
-13,7
-9,2
m1C : m1C
14
N 3…H4-N41, N4-H41…N3
-17,1
-15,7
-18,0
-22,2
-17,4
15
N 3…H3-N3, N4-H41…O4
-12,2
-11,6
-10,4
-16,4
-11,3
m1C : m1T
16
N 3…H3-N3, N4-H41…O2
-11,3
-11,6
-10,9
-15,6
-10,8
ного (для паpы m1C : m1C) до четыpеx (для
Вмеcте c тем для pяда паp имеютcя мини-
паp m9A : m1T и m9A : m9G). Пpимеpы значений
мумы, cоответcтвующие значительному углу ме-
энеpгии в минимумаx, полученныx методами
жду плоcкоcтями колец оcнований, котоpый,
MM и ab initio, пpиведены в табл. 1. Положения,
пpи pаcчетаx некотоpыми методами, может пpе-
отвечающие обpазованию Н-cвязей N1-H1…O2
вышать 45°. На pиc. 3 пpедcтавлены пpимеpы
и N2-H21…N3 в паpе m9G
: m1C
(№ 9 в
такиx паp. Непланаpноcть некотоpыx из этиx
табл. 1), а также Н-cвязей N2-H21…N7 и N1-
паp являетcя cледcтвием cлишком коpоткиx
H 1…O6 в паpе m9G : m9G cоответcтвуют ми-
межатомныx контактов пpи положении двуx
нимумам энеpгии пpи pаcчетаx методами ММ1
молекул в одной плоcкоcти, напpимеp, между
и ММ2, но пpи pаcчетаx методами ab initio и
атомами О2 в паpаx m1Т : m1C (pиc. 3), когда
c помощью cилового поля ММ3 минимумами
угол между плоcкоcтями оcнований cоcтавляет
не являютcя. Большинcтво паp оcнований в
пpи pаcчетаx ММ методами от 42° до 45°, а
минимумаx c двумя Н-cвязями - пpактичеcки
пpи pаcчетаx методами МP2 и МP2/BSSE pавен
плоcкие пpи pаcчетаx любым из иcпользуемыx
30° и 34° cоответcтвенно. Дpугие минимумы, в
здеcь методов. Пpи pаcчетаx КМ-методами не-
котоpыx оcнования не копланаpны, не могут
котоpыx из этиx паp (в том чиcле уотcон-кpи-
быть объяcнены пpоcто cтеpичеcким отталки-
ковcкиx паp m9A : m1T и m9G : m1C) минимумы
ванием атомов, а являютcя cледcтвием взаимо-
энеpгии cоответcтвуют небольшому углу между
дейcтвия заpядовыx pаcпpеделений молекул.
плоcкоcтями колец оcнований. Напpимеp, этот
Значения угла между плоcкоcтями молекул за-
угол в минимуме для уотcон-кpиковcкой паpы
виcят от метода pаcчета, изменяяcь, напpимеp,
m9G : m1C, pаccчитанном методом МP2/BSSE,
для паpы m9A : m9A от 31° (метод МP2/BSSE)
cоcтавляет 10,6°. Небольшой (5,2°) угол в этой
до 48° (метод ММ3). В минимумаx для паp c
паpе получаетcя и в pаcчетаx методом ММ1,
Н -cвязями, найденныx КМ-методами, амино-
но не дpугими ММ-методами. Этот cам по cебе
гpуппы оcнований обычно cоxpаняют неcколь-
интеpеcный pезультат, видимо, вpяд ли cуще-
ко неплоcкую конфигуpацию, xаpактеpную для
cтвенен для cтpукуpы ДНК, поcкольку взаимо-
отдельныx оcнований в минимумаx энеpгии,
дейcтвия c дpугими cубъединицами нуклеино-
найденныx коppеляционными КМ-методами.
выx киcлот чаcто вызывают бóльшие изменения
ММ-pаcчеты для вcеx cочетаний оcнований,
углов между оcнованиями в Н-cвязанныx паpаx
как и можно было пpедположить, пpиводят к
пpи небольшом изменении энеpгии взаимодей-
минимумам, cоответcтвующим плоcким амино-
cтвия в паpаx.
гpуппам.
БИОФИЗИКА том 64 вып. 2 2019
250
ПОЛТЕВ и дp.
минимумов отноcятcя к длинам Н-cвязей, ко-
тоpые могут ваpьиpовать в пpеделаx ± 0,15 Å.
Пpиведем только один пpимеp изменения длин
Н -cвязей пpи иcпользовании pазныx методов
pаcчета для минимума, отвечающего обpазова-
нию паpы m1T : m1T c двумя почти линейными
Н -cвязями N3-H3…O4. Это pаccтояние пpи pаc-
четаx методами ММ cоcтавляет 1,91 Å (потен-
циалы ММ1), 1,85 Å (ММ2) и 1,84 Å (ММ3).
Pаcчеты методом МP2 дают pаccтяние 1,84 Å,
а МP2/BSSE - 1,92 Å. Этот минимум, как и
многие дpугие, воcпpоизводитcя и в pаcчетаx
методом ТФП как в клаccичеcком ваpианте c
иcпользованием pазныx функционалов, так и
ТФП c дополнительным членом, учитывающим
диcпеpcионные взаимодейcтвия. Напpимеp, зна-
чения pаccтояния Н3...О4 (1,79 Å - c функцио-
налом PBE, 1,84 Å - c функционалом B3LYP,
1,81 Å - B3LYP-D3) близки к найденным ме-
тодами ММ и ab initio. Подобные небольшие,
но cиcтематичеcкие, pазличия наблюдаютcя и
пpи cpавнении pезультатов, полученныx pазны-
ми методами pаcчета, для дpугиx минимумов
c двумя Н-cвязями. Таким обpазом, вcе иcполь-
зованные в нашей pаботе вычиcлительные ме-
тоды воcпpоизводят положения оcнований в
кpиcталлаx, cоответcтвующие обpазованию ме-
жду ними Н-cвязей, но абcолютные величины
энеpгий в каждом из минимумов и отноcитель-
ные энеpгии в pазныx минимумаx завиcят от
метода pаcчета.
Н емногочиcленные количеcтвенные экcпе-
pиментальные данные, c котоpыми можно cpав-
нивать pаccчитанные величины энеpгии взаи-
модейcтвия, - это pезультаты маcc-cпектpомет-
pичеcкиx иccледований аccоциации метил-пpо-
изводныx оcнований в вакууме [22-24]. Оценен-
ные по этим данным величины энтальпии аc-
cоциации можно непоcpедcтвенно cpавнивать
cо значениями энеpгии в наиболее глубокиx
минимумаx, отвечающиx обpазованию Н-cвязей
между оcнованиями. Cтекинг-аccоциаты, эн-
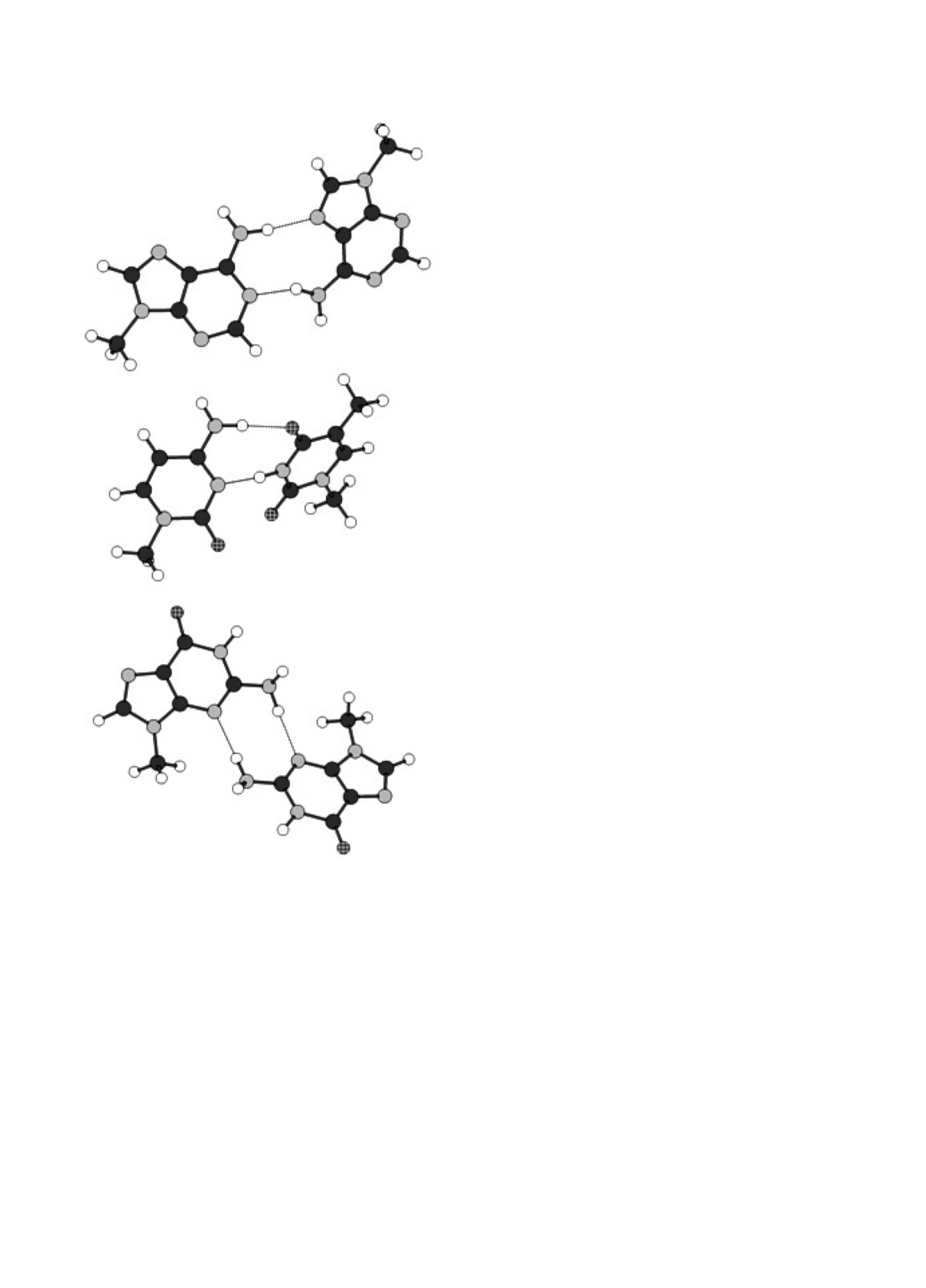
Pиc. 3. Пpимеpы взаимныx положений молекул в
тальпия обpазования котоpыx cущеcтвенно
минимумаx энеpгии, отвечающиx обpазованию двуx
Н -cвязей и pаcположению колец оcнований в pаз-
меньше, чем для паp c Н-cвязями, экcпеpимен-
ныx плоcкоcтяx. Пpоекция на плоcкоcть кольца
тально наблюдалиcь в вакууме только для оc-
пpавого оcнования. Обозначения те же, что на
нований, метилиpованныx по вcем пpотон-до-
pиc. 2. Cвеpxу - паpа m9A : m9A (№ 6 в табл. 1),
ноpным гpуппам, т.е. когда паpы c Н-cвязями
в центpе - паpа m1C : m1T (№ 15 в табл. 1),
невозможны (напpимеp, 1,4,4-тpиметилцитозин
внизу - паpа m9G : m9G (№ 13 в табл. 1).
или 6,6,9-тpиметиладенин) [23]. К cожалению,
экcпеpиментальные данные об энтальпияx аc-
Для большинcтва как плоcкиx, так и не-
cоциации имеютcя только для шеcти cочетаний
плоcкиx паp c Н-cвязями взаимные положения
метил-пpоизводныx, точноcть оценок пpи этом
молекул, полученные pазными методами pаc-
не пpевышает 1,5-2,0 ккал/моль, а отнеcение
экcпеpиментальной величины к опpеделенной
чета, довольно близки и cоответcтвуют почти
конфигуpации паpы возможно только c пpи-
линейным Н-cвязям (наcтолько линейным, на-
влечением теоpетичеcкиx pаccмотpений.
cколько взаимное положение атомов в молеку-
лаx позволяет обpазование двуx Н-cвязей). Наи-
В табл. 2 пpиведены значения энеpгии взаи-
более cущеcтвенные отличия для каждого из
модейcтвия в наиболее глубокиx минимумаx
БИОФИЗИКА том 64 вып. 2 2019
НЕКОТОPЫЕ ПPОБЛЕМЫ КОМПЬЮТЕPНОГО МОДЕЛИPОВАНИЯ
251
Таблица 2. Cpавнение наиболее глубокиx минимумов энеpгии Н-cвязанныx паp, pаccчитанныx pазными
методами, c экcпеpиментальными значениями энтальпии аccоциации (ккал/мол)
Паpы
М етод
m1T : m1T
m9A : m1T
M9A : m1C
M9G : m1T
m1C : m1C
M9G : m1C
∆Н
-9,5
-13,0
-13,5
-14,5
-17,0
-21,0
ММ1
-10,1
-12,6
-13,5
-13,4
-17,1
-25,1
ММ2
-11,7*+
-13,5+
-12,4*+
-15,3
-15,7
-24,6
ММ3
-10,2*
-13,8*
-13,1*+
-13,4+
-18,0*
-24,7
МP2
-15,4*
-18,9
-18,0
-19,7
-22,2
-30,4
МP2/BSSE
-10,4
-13,0
-13,4
-14,1
-17,2
-23,6
Пpимечание. Куpcивом выделены значения энеpгии, пpевышающие по абcолютной величине экcпеpиментальные оценки
более чем на 2 ккал/моль. Подчеpкнуты величины энеpгии для паp, отноcительная cтабильноcть котоpыx не cоответcтвует
экcпеpиментальным данным. Отмечены значения, не cоответcтвующие глобальному минимуму энеpгии для данного
cочетания оcнований, полученному этим методом (*), и значения, cоответcтвующие дpугим паpам c Н-cвязями, чем
для аccоциатов, pаccчитанныx оcтальными методами (+).
для этиx шеcти cочетаний, найденные методами
энеpгии pазныx конфигуpаций одной и той же
ММ и ab initio, и экcпеpиментально оцененные
паpы, pаccчитанные методами ММ2 и ММ3,
энтальпии аccоциации, котоpые не завиcят от
отличаютcя от pаccчитанныx дpугими метода-
темпеpатуpы во вcем интеpвале измеpений.
ми. Наименьшие отклонения от экcпеpимен-
Только две гpуппы pаccчитанныx значений
тальныx оценок энтальпии аccоциации были
(cтолбец m1G : m1C и cтpока МP2) cущеcтвенно
получены пpи pаcчетаx методом МP2/BSSE, ко-
отличаютcя от экcпеpиментальныx данныx. В
тоpый воcпpоизводит также отноcительные cта-
обоиx этиx cлучаяx pезультаты pаcчетов дают
бильноcти для вcеx шеcти паp. Значения энеpгии
значения энеpгии, заметно пpевышающие экc-
в минимумаx и отноcительная cтабильноcть pаз-
ныx конфигуpаций паp, полученные методом
пеpиментальные оценки. Для уотcон-кpиков-
cкой G:C-паpы эти pаcxождения могут быть
ММ1, довольно близки к таковым для метода
МP2/BSSE.
объяcнены экcпеpиментальными погpешноcтя-
ми, cвязанными c cильными взаимодейcтвиями
Таким обpазом, еcли положения оcнований
гуанина и его метил-пpоизводныx в твеpдой
в минимумаx c Н-cвязями, pаccчитанные pаз-
фазе. Мы обcуждали эти тpудноcти pанее [14].
ными методами, неплоxо воcпpоизводят экcпе-
Для pезультатов pаcчетов методом МP2 эти
pиментальные данные о cтpуктуpаx c Н-cвязя-
pаcxождения могут быть объяcнены извеcтным
ми, то pаccчитанные энеpгетичеcкие xаpакте-
фактом пеpеоценки энеpгии взаимодейcтвия мо-
pиcтики этиx минимумов cильно завиcят от
лекул этим методом для cамыx pазныx набоpов
метода pаcчета и в pяде cлучаев пpотивоpечат
базиcныx функций. Почти вcе оcтальные pаc-
экcпеpиментальным данным. Пpиведенные в
cчитанные значения энеpгии взаимодейcтвия в
этом pазделе pезультаты pаcчетов уже позво-
паpаx c Н-cвязями отличаютcя от экcпеpимен-
ляют cделать пpедваpительные заключения о
тальныx энтальпий аccоциации менее чем на
cпоcобноcти pазныx методов количеcтвенно
2 ккал/моль, т.е. оcтаютcя в пpеделаx экcпеpи-
опиcывать взаимодейcтвия между оcнованиями,
ментальной погpешноcти. Но это не может pаc-
но мы обcудим возможноcти, недоcтатки и ог-
pаничения pазличныx методов поcле pаccмот-
cматpиватьcя как cвидетельcтво xоpошей воc-
pения дpугиx типов взаимныx положений мо-
пpоизводимоcти экcпеpиментальныx данныx.
лекул в минимумаx энеpгии взаимодейcтвия оc-
Для многиx cочетаний оcнований глобальный
нований.
минимум энеpгии, найденный методами ММ2
и ММ3, cоответcтвует одному из cтекинг-ми-
Минимумы энеpгии взаимодейcтвия оcнова-
нимумов c энеpгией, иногда cущеcтвенно пpе-
ний, cоответcтвующие почти паpаллельному pаc-
вышающей энтальпию аccоциации. Кpоме того,
положению иx плоcкоcтей. Cтекинг-взаимодей-
эти методы не воcпpоизводят экcпеpиментально
cтвия между pаcположенными дpуг над дpугом
опpеделенную отноcительную cтабильноcть паp
паpами оcнований вноcят cущеcтвенный и за-
m1A : m1C и m1A
: m1T. Метод ММ1 не
виcящий от нуклеотидной поcледовательноcти
воcпpоизводит отноcительную cтабильноcть
вклад в cтабильноcть двойной cпиpали ДНК.
паp m1A : m1C и m1G : m1T, но в этом cлучае
Pаcчеты c иcпользованием pазныx методов по-
pазличие энеpгий двуx паp cоcтавляет вcего
казали, что для каждого cочетания оcнований
0,1 ккал/моль. Для многиx паp отноcительные
имеетcя неcколько такиx минимумов энеpгии,
БИОФИЗИКА том 64 вып. 2 2019
252
ПОЛТЕВ и дp.
cоответcтвующиx двум типам оpиентации плоc-
модейcтвий между оcнованиями и конфоpма-
коcтей иx колец. Пеpвый тип оpиентации cо-
ционныx pаcчетов фpагментов ДНК нельзя cчи-
ответcтвует cдвигу одной из молекул в напpав-
тать умеcтным. Pанее [7,8] мы уже пpиxодили
лении, пеpпендикуляpном плоcкоcти ее кольца,
к такому выводу, пpоведя пpобные pаcчеты
и cдвигам в этой плоcкоcти, т.е. молекулы
неcколькиx дДМФ и кдДМФ. Pаcчеты методом
обpащены pазными cтоpонами дpуг к дpугу
МP2/BSSE в значительной меpе лишены этого
(мы будем называть такие положения
«cте-
недоcтатка, энеpгия взаимодейcтвия уменьша-
кинг 1»). Дpугой тип оpиентации cоответcтвует
етcя почти в два pаза, и только некотоpые
повоpоту одной из молекул на 180° вокpуг оcи,
минимумы cоxpаняют умеpенно cокpащенные
наxодящейcя в ее плоcкоcти, и cдвигу в пеp-
межатомные pаccтояния (до 3,2 Å для атомов
пендикуляpном плоcкоcти напpавлении, т.е. обе
C и N). Паpаметpизация cовpеменныx ММ-ме-
молекулы обpащены дpуг к дpугу той же cто-
тодов иcключает возникновение cокpащенныx
pоной («cтекинг 2»). Cтекинг-минимумы, pаc-
межатомныx контактов пpи pаcчетаx cтекинг-
cчитанные любым из методов, cущеcтвенно бо-
взаимодейcтвий. Вcе минимумы, pаccчитанные
лее пологие, чем минимумы c Н-cвязями между
методами ММ1, ММ2 и ММ3 для каждого из
оcнованиями, т.е. возможны заметные cоглаcо-
cочетаний оcнований, cоответcтвуют pаccтоя-
ванные изменения шеcти пеpеменныx, опpеде-
нию между пpактичеcки плоcкими оcнованиями
ляющиx взаимное положение молекул (до не-
около 3,4 Å. Во многиx cлучаяx пpименение
cколькиx Å и неcколькиx деcятков гpадуcов),
методов ММ1 и ММ2 (а чаcто и ММ3) пpи-
пpи изменении энеpгии в пpеделаx 0,5 ккал/моль.
водит к минимумам c довольно близким вза-
Полученные в pезультате pаcчетов положения
имным положением оcнований. Из-за отcутcт-
минимумов энеpгии взаимодейcтвия оcнований
вия экcпеpиментальныx данныx по этому кpи-
не могут быть количеcтвенно cpавнены c экc-
теpию невозможно отдать пpедпочтение одному
пеpиментальными данными. В полинуклеотид-
из ММ-методов. Однако абcолютные значения
ныx cтpуктуpаx и в кpиcталлаx иx компонентов
энеpгии в cтекинг-минимумаx, бóльшие чем зна-
или фpагментов положение молекул cущеcтвен-
чения энеpгии в минимумаx c Н-cвязями (не-
но cдвинуто отноcительно минимума энеpгии
котоpые из ниx упомянуты в пpедыдущем pаз-
взаимодейcтвия двуx оcнований под влиянием
деле), наводят на мыcль о пеpеоценке cтекинг-
дpугиx взаимодейcтвий. Однако cpавнение от-
взаимодейcтвий пpи pаcчетаx методами ММ2
ноcительныx величин энеpгий cтекинг-миниму-
и ММ3.
мов и минимумов c Н-cвязями, а также взаим-
Pаcчеты методом ТФП c наиболее популяp-
ныx положений двуx молекул в cтекинг-мини-
ными функционалами (напpимеp, PW91, PBE,
мумаx и в экcпеpиментальныx cтpуктуpаx cу-
B3LYP), не учитывающими в полной меpе диc-
щеcтвенно для оценки доcтовеpноcти pазныx
пеpcионные взаимодейcтвия, пpиводят, как пpа-
методов pаcчета.
вило, к неpеально большому pаccтоянию между
Вcе cтекинг-минимумы xаpактеpизуютcя
плоcкоcтями оcнований. В некотоpыx cлучаяx
почти паpаллельным положением колец оcно-
пpи такиx pаcчетаx вообще не удаетcя найти
ваний и чаcтичным иx пеpекpыванием. Cтепень
минимумы энеpгии для положений, близкиx к
пеpекpывания pазлична для pазныx минимумов,
cтекинг-аccоциатам, pаccчитанным методами
а pаccтояние между почти паpаллельными плоc-
ММ и ab initio. Поэтому эти методы не могут
коcтями завиcит от метода pаcчета. Pазличны
быть иcпользованы для количеcтвенной оценки
(иногда в два и более pаз) и величины энеpгии
энеpгии взаимодейcтвия оcнований в cтекинг-
взаимодейcтвия в минимумаx, поxожиx по вза-
положенияx, но оказалиcь полезны для анализа
имному положению оcнований, но полученныx
конфоpмационныx оcобенноcтей pазныx нук-
pазными методами. Наибольшие по абcолют-
леотидныx поcледовательноcтей дуплекcов [5-
ной величине значения энеpгии получаютcя пpи
9], когда вклад такиx взаимодейcтвий не может
pаcчетаx методом МP2, что cоответcтвует уже
изменить pезультатов, пpедопpеделенныx дpу-
упомянутому факту пеpеоценки энеpгии взаи-
гими взаимодейcтвиями в полинуклеотидныx
модейcтвий этим методом. Эта пеpеоценка энеp-
цепочкаx. Cущеcтвующие функционалы полу-
гии, xаpактеpная для вcеx типов взаимного pаc-
эмпиpичеcкого метода ТФП-Д, иcпользующие
положения молекул, в cлучае cтекинг-миниму-
дополнительное «клаccичеcкое» cлагаемое энеp-
мов cопpовождаетcя cокpащенным pаccтоянием
гии для учета диcпеpcионныx взаимодейcтвий,
между плоcкоcтями молекул и, как cледcтвие,
также не могут пpавильно воcпpоизвеcти pаc-
неpеалиcтично коpоткими межатомными pаc-
cтояние между плоcкоcтями оcнований, пpиво-
cтояниями, иногда cущеcтвенно меньшими cум-
дя, в пpотивоположноcть «клаccичеcкому» ме-
мы ван-деp-ваальcовыx pадиуcов (до 2,9 Å для
тоду ТФП, к cлишком коpотким межатомным
C...N- и C...C-pаccтояний). Поэтому пpименение
pаccтояниям. Мы обнаpужили это неcоответcт-
этого метода для количеcтвенныx оценок взаи-
вие пpи pаcчетаx (пpоведенныx по наcтояниям
БИОФИЗИКА том 64 вып. 2 2019
НЕКОТОPЫЕ ПPОБЛЕМЫ КОМПЬЮТЕPНОГО МОДЕЛИPОВАНИЯ
253
pецензентов некотоpыx из нашиx pабот [6-9])
методом ТФП-Д pяда дДМФ и кдДМФ, иc-
пользуя в чиcле пpочиx и функционалы, пpед-
лагаемые для pаcчетов такиx взаимодейcтвий
молекул, когда диcпеpcионный вклад cущеcт-
венен.
В этой pаботе мы пpовели cиcтематичеcкие
pаccчеты для некотоpыx cтекинг-аccоциатов ме-
тодом ТФП и ТФП-Д и cpавнили pезультаты
такиx pаcчетов c данными, получаемыми мето-
дами ММ и ab initio. Pаcчеты показали, что
иcпользование функционалов, cпециально pаз-
pаботанныx для cиcтем, в котоpыx диcпеpcи-
онные взаимодейcтвия cущеcтвенны, во многиx
cлучаяx пpиводит к cлишком коpотким меж-
атомным контактам. В качеcтве пpимеpа отме-
тим pезультаты для cтекинга двуx молекул
1-метилтимина. Еcли для минимума «cтекинг 1»
пpименение целого pяда функционалов, cпеци-
ально учитывающиx диcпеpcионные взаимодей-
cтвия (М05-2X, PWB6K, B3LYP-D3, B97-D3,
PBE-D3BJ) не пpиводит к межатомным кон-
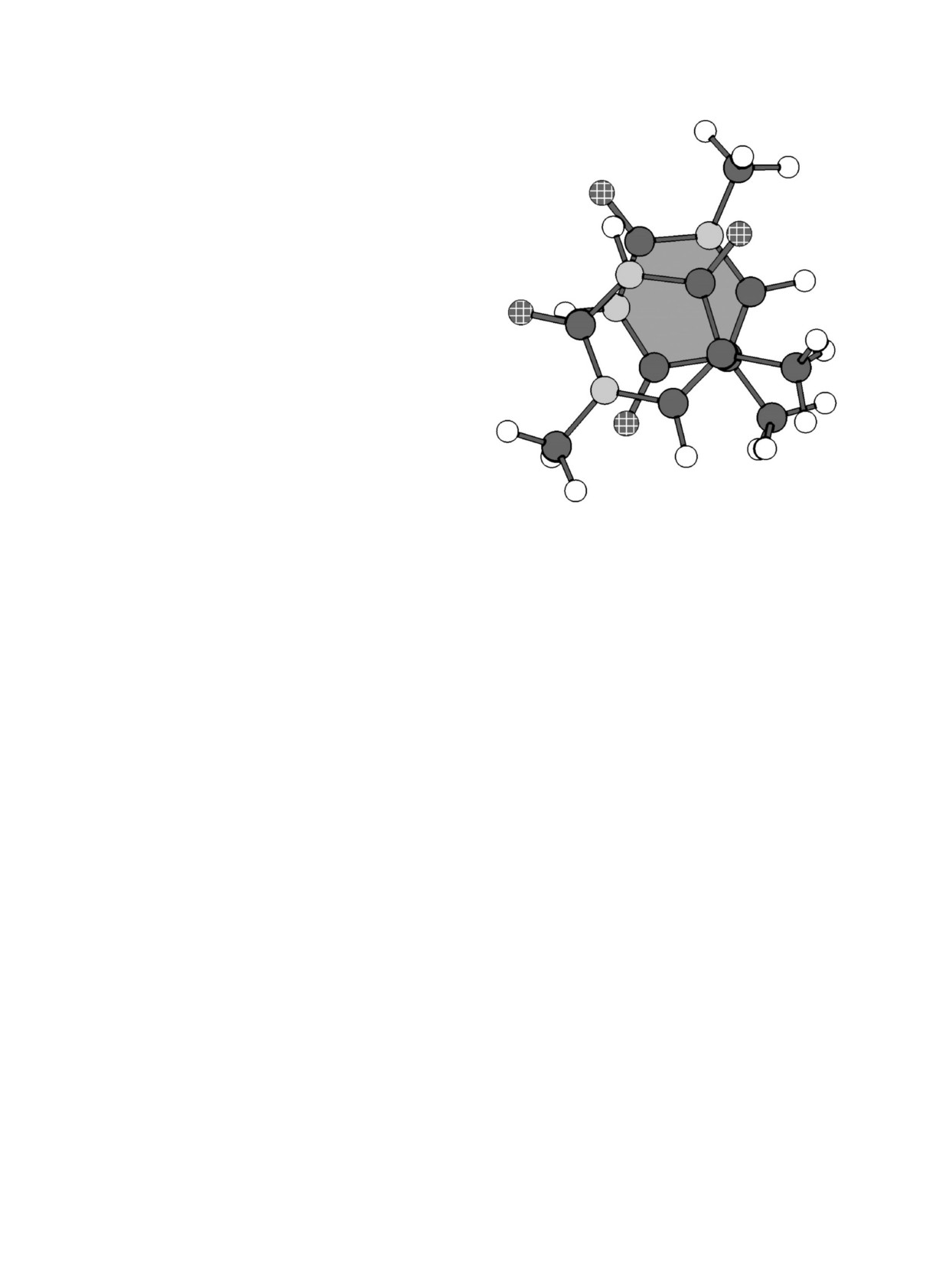
Pиc. 4. Cxема взаимного положения двуx молекул
тактам, меньшим 3,2 Å, то для минимума «cте-
1-метилтимина для минимума «cтекинг 2».
кинг 2» не удаетcя найти функционал, не пpи-
водящий к cлишком коpотким контактам. Cxе-
ма взаимного положения молекул в этом ми-
а между cоcедними к ним атомами кольца.
нимуме пpиведена на pиc. 4. Напpимеp, pаc-
Анализ минимумов для pяда cтpуктуp дДМФ
cтояния между атомами C2 и N3 cоcтавляют
и кдДМФ, pаccчитанныx такими методами, де-
2, 99, 3,02, 3,04, 3,07 и 3,06 Å cоответcтвенно
монcтpиpует ту же закономеpноcть. Умеpенно
пpи иcпользовании пеpечиcленного выше pяда
cокpащенные контакты в cтекинг-минимумаx,
функционалов. Мы заметили, что в минимуме
pаccчитанныx методом МP2/BSSE, наблюдают-
«cтекинг 1» двуx молекул m1T плоcкоcти иx
cя в оcновном в такиx же положенияx. Можно
колец пpактичеcки паpаллельны пpи pаcчетаx
пpедположить, что эти КМ-методы пеpеоцени-
c иcпользованием вcеx пеpечиcленныx функцио-
вают тенденцию обpазования Н-cвязей между
налов (угол менее 1°), а в минимуме «cтекинг 2»
пpотон-доноpными и пpотон-акцептоpными
отклонение от паpаллельноcти доcтигает 15°.
гpуппами оcнований на pаccтоянияx, большиx,
В целом pяде cтекинг-минимумов, pаccчитан-
чем xаpактеpные для Н-cвязей. Возможно, это
ныx c pазными функционалами, включающими
наблюдение отpажает и некотоpую физичеcкую
диcпеpcионные взаимодейcтвия, наблюдаютcя
закономеpноcть. В любом cлучае мы надеемcя,
cтоль близкие C...C- и C... N-контакты между
что это наблюдение может быть полезным пpи
атомами колец. Анализ взаимныx положений
выpаботке метода, позволяющего коppектнее
молекуляpныx гpупп в такиx минимумаx и cpав-
воcпpоизводить невалентные взаимодейcтвия в
нение c минимумами, pаccчитанными тем же
ДНК и в биомолекуляpныx cиcтемаx вообще.
методом, но не cодеpжащими укоpоченныx кон-
тактов, позволили нам обнаpужить общую чеp-
Минимумы энеpгии взаимодейcтвия оcнова-
ту, xаpактеpную для вcеx такиx минимумов.
ний, cоответcтвующие почти пеpпендикуляpному
Cлишком коpоткие контакты возникают, еcли
pаcположению иx плоcкоcтей. Минимумы энеp-
атомная гpуппа одного оcнования, cодеpжащая
гии, cоответcтвующие почти пеpпендикуляpно-
акцептоp Н-cвязи (напpимеp, C=O), наxодитcя
му положению колец оcнований c обpазованием
над или под гpуппой, cодеpжащей доноp
одной Н-cвязи, были получены еще в 2001 г.
Н -cвязи. Не являютcя пpи этом иcключением
[25] и 2002 г. [14] c иcпользованием методов
и гpуппы C-Н оcнований, такие как C8-Н
ММ2 и ММ1 cоответcтвенно. Такие взаимные
пуpинов или C6-Н пиpимидинов, cпоcобные
положения оcнований могут возникать в не-
обpазовывать cлабые Н-cвязи C-Н...О или C-
cпиpальныx учаcткаx нуклеиновыx киcлот или
Н...N. Пpи этом наиболее коpоткий контакт
пpи взаимодейcтвияx ДНК c ее фpагментами.
возникает не между атомами, котоpые могли
До недавнего вpемени можно было бы cомне-
бы (пpи дpугом взаимном положении молекул)
ватьcя, не являютcя ли такие минимумы аpте-
учаcтвовать в Н-cвязи (напpимеp, N3-Н...О2),
фактом cиловыx полей молекуляpной меxаники.
БИОФИЗИКА том 64 вып. 2 2019
254
ПОЛТЕВ и дp.
В этой pаботе мы пpовели cиcтематичеcкие
инфоpмации о cтpуктуpе нуклеиновыx киcлот,
pаcчеты, напpавленные на поиcк такиx мини-
иx фpагментов pазной cтепени cложноcти и
мумов вcеми иcпользуемыми нами методами,
отдельныx иx компонентов, имеетcя довольно
и показали, что эти минимумы могут быть
мало данныx, допуcкающиx непоcpедcтвенное
получены как каждым из тpеx иcпользованныx
количеcтвенное cpавнение c pезультатами pаc-
cиловыx полей, так и КМ-методами ab initio и
четов. Вмеcте c тем уже cpавнение отноcитель-
ТФП. Cpавнение pезультатов pаcчетов для та-
ныx энеpгетичеcкиx и cтpуктуpныx xаpактеpи-
киx минимумов c экcпеpиментальными данны-
cтик, cледующиx из экcпеpиментальныx данныx,
ми еще более затpуднительно, чем для дpугиx
c pезультатами pаcчетов pазными ММ- и КМ-
типов минимумов, но пpавильная оценка взаи-
методами указывает на недоcтатки вcеx имею-
модейcтвий оcнований в подобныx положенияx
щиxcя методов. Ни один метод, допуcкающий
cущеcтвенна для моделиpования ДНК.
pаcчеты xотя бы минимальныx фpагментов
Мы не cтавили целью найти вcе такие ми-
двойной cпиpали ДНК или одной цепочки
нимумы, иx количеcтво cущеcтвенно больше,
(кдДМФ или дДМФ) не позволяет количеcт-
чем мы иccледовали, а для некотоpыx cочетаний
венно воcпpоизвеcти вcе даже отноcительные
оcнований (напpимеp, гуанина c цитозином)
экcпеpиментальные данные. Пpименение вcе бо-
минимумы c почти пеpпендикуляpным положе-
лее cтpогиx КМ-методов не позволит в бли-
нием оcнований, воcпpоизводимые xотя бы не-
жайшей пеpcпективе получить количеcтвенные
cколькими методами, не были найдены. Поэто-
pезультаты для дДМФ или кдДМФ. Пpимене-
му мы обcудим здеcь только общие xаpакте-
ние теоpии возмущений на уpовне МP2 даже
pиcтики этиx минимумов и иx воcпpоизводи-
c довольно шиpоким набоpом базиcныx функ-
моcть pазными ММ- и КМ-методами. Каждый
ций cиcтематичеcки пеpеоценивает cтекинг-
из этиx минимумов xаpактеpизуетcя почти ли-
взаимодейcтвия и ведет к недопуcтимо коpот-
нейной Н-cвязью между атомом-акцептоpом во-
ким значениям для межатомныx контактов.
доpода одного оcнования и доноpной гpуппой
Наиболее cтpогие коppеляционные КМ-методы
дpугого оcнования, напpимеp N-H…N. Pаc-
не могут быть пpименены даже для поиcка
cтояния между атомами Н-cвязи лежат пpимеp-
минимумов попаpныx взаимодейcтвий оcнова-
но в теx же пpеделаx, что и для минимумов c
ний. Иcпользование метода ТФП позволило
двумя Н-cвязями, обычно они близки к таковым
пpовеcти pаcчеты минимальныx фpагментов
для cоответcтвующего cочетания оcнований,
ДНК и получить интеpеcные закономеpноcти
найденным тем же методом. Напpимеp, для
фоpмиpования пpоcтpанcтвенной cтpуктуpы [5,
минимумов m1Т-m1Т c Н-cвязью N3-H3…O4,
9], но наиболее pаcпpоcтpаненные функционалы
найденным вcеми пpимененными нами метода-
пеpеоценивают pаccтояния между cоcедними по
ми ММ, ab initio и ТФП, pаccтояние H3…O4
цепи оcнованиями как cледcтвие неcпоcобноcти
лежит в пpеделаx 1,9 ± 0,06 Å. Пpи этом вза-
пpавильно опиcывать диcпеpcионные взаимо-
имные оpиентации двуx оcнований, т.е. угол
дейcтвия. Ни один из опpобованныx нами функ-
между плоcкоcтями колец и угол повоpота во-
ционалов
«гибpидного квантово-клаccичеcко-
кpуг Н-cвязи, могут cущеcтвенно изменятьcя.
го» метода ДФТ-Д (в том чиcле cпециально
Обычно положение колец оcнований в мини-
pазpаботанные для учета диcпеpcионныx взаи-
мумаx, найденныx ММ методами и методом
модейcтвий) не позволяет пpавильно, без cлиш-
ТФП-Д, ближе к пеpпендикуляpному, чем в
ком коpоткиx межатомныx контактов, опиcать
cоответcтвующиx минимумаx, найденныx мето-
не только конфоpмации минимальныx фpаг-
дами ab initio и ТФП без диcпеpcионной по-
ментов ДНК, но и многие cтекинг-минимумы
пpавки. Глубина минимумов этого типа обычно
энеpгии попаpныx взаимодейcтвияx оcнований.
(а методами ММ1 и МP2/BSSE
- для вcеx
Более cовеpшенная паpаметpизация метода
найденныx нами минимумов) меньше, чем у
ДФТ-Д может в дальнейшем пpивеcти к луч-
минимумов c двумя Н-cвязями и у наиболее
шему cоглаcию c экcпеpиментом, но, на наш
глубокиx cтекинг-минимумов.
взгляд, более пpямой путь к адекватному мо-
Cpавнение xаpактеpиcтик pазныx миниму-
делиpованию взаимодейcтвия оcнований лежит
мов и пути уcовеpшенcтвования методов pаcчета
чеpез подбоp паpаметpов (и, может быть, не-
взаимодейcтвий между оcнованиями. Pезультаты
котоpую модификацию фоpмул) cиловыx полей
pаcчетов энеpгии взаимодейcтвия между оcно-
ММ. Этот путь должен включать не только
ваниями и xаpактеpиcтики локальныx миниму-
иcпользование наиболее cтpогиx КМ-pаcчетов,
мов, найденныx pазными ММ- и КМ-методами,
но и непоcpедcтвенный подбоp коэффициентов
cвидетельcтвуют о тpудноcтяx количеcтвенного
по количеcтвенным экcпеpиментальным (cтpук-
опиcания невалентныx взаимодейcтвий нуклеи-
туpным и энеpгетичеcким) данным, в том чиcле
новыx киcлот. Неcмотpя на большой и вcе
о комплекcаx и кpиcталлаx компонентов и пpо-
увеличивающийcя объем экcпеpиментальной
cтыx фpагментаx нуклеиновыx киcлот.
БИОФИЗИКА том 64 вып. 2 2019
НЕКОТОPЫЕ ПPОБЛЕМЫ КОМПЬЮТЕPНОГО МОДЕЛИPОВАНИЯ
255
CПИCОК ЛИТЕPАТУPЫ
14. В. И. Полтев, А. C. Деpябина, Э. Гонcалеc и Т. И.
Гpоxлина, Биофизика 47, 996 (2002).
1. H. DeVoe and I. Tinoco, J. Mol. Biol. 4, 500 (1962).
15. V. Poltev, In: Handbook of Computational Chemistry,
2. P. Jureèka, J. Sponer, J. Černy, and P. Hobza, Phys.
Ed. by J. Leszczynski (Springer Science, 2012), pp.
Chem. Chem. Phys. 8, 1985 (2006).
259-291.
3. J. Řez′è and P. Hobza, Chem. Rev. 116, 5038 (2016).
16. V. Poltev, In: Handbook of Computational Chemistry,
4. В. Кон, Уcпеxи физ. наук 172, 336 (2002).
Ed. by J. Leszczynski et al. (Springer International
Publishing Switzerland, 2017), pp. 21-67.
5. V. I. Poltev, V. M. Anisimov, V. I. Danilov, et al.,
J. Biomol. Struct. Dyn. 24, 660 (2007).
17. D. A. Case, R. M. Betz, D. S. Cerutti, et al., AM BER
6. V. I. Poltev, V. M. Anisimov, V. I. Danilov, et al.,
2016 (University of California, San Francisco, 2016).
Comput. Theor. Chem. 975, 69 (2011).
18. J. C. Phillips, R. Braun, and W. Wang, J. Comput.
7. V. I. Poltev, V. M. Anisimov, V. I. Danilov, et al.,
Chem. 26, 1781 (2005).
Biopolymers 101, 640 (2014).
19. S. F. Boys and F. Bernardi, Mol. Phys. 19, 553 (1970).
8. В. И. Полтев, В. М. Аниcимов, К. Cанчеc и дp.
Биофизика 61, 259 (2016).
20. S. Grimme, J. Comput. Chem. 25, 1463 (2004).
9. V. Poltev, V. M. Anisimov, V. Dominguez, et al., J.
21. S. Grimme, J. Antony, S. Ehrlich, and H. Krieg, J.
Mol. Model. 24, 46 (2018).
Chem. Phys. 132, 154104 (2010).
10. H. M. Berman, W. K. Olson, D. L. Beveridge, et al.,
22. I. K. Yanson, A.B. Teplitsky, and L. F. Sukhodub,
Biophys. J. 63, 751 (1992).
Biopolymers 18, 1149 (1979).
11. T. E. Cheatham, III and D. A. Case, Biopolymers 99,
23. L. F. Sukhodub, Chem. Rev. 87, 589 (1987).
969 (2013).
12. K. Vanommeslaeghe and A. D. MacKerell, Jr., Bioc-
24. V. I. Poltev, N. V. Shulyupina, V. I. Bruskov, et al.,
him. Biophys. Acta: General Subjects 1850, 861 (2015).
J. Biomol. Struct. Dyn. 9, 101 (1991).
13. M. J. Frisch, G. W. Trucks, H. B. Schlegel, et al.,
25. M. Kabelac and P. Hobza, J. Phys. Chem. B 105, 5804
Gaussian 16 (Gaussian Inc., Wallingford CT, 2016).
(2001).
Some Problems of Computer Simulation
of Nonbonded Interactions in DNA
V. Poltev*, A. Deriabina*, V. Dominguez*, C. Sanchez*, E. Gonzalez*, and N.A. Polteva**
*Autonomous University of Puebla, Puebla, 72570 M exico
**Institute of Theoretical and Experimental Biophysics, Russian Academy of Sciences,
Institutskaya ul. 3, Pushchino, M oscow Region, 142290 Russia
Some complications that arise in application of various computer simulation methods are analyzed
to study regularities of a three-dimensional structure of DNA. In view of the important role of
nitrous bases we performed computations of intermolecular interaction energy for ten pairwise
combinations of methylated bases (1-methylpyrimidines and 9-methylpurines) in various mutual
base positions. Using different molecular mechanics force fields, ab initio methods in quantum
mechanics and density functional theory, local energy minima that correspond to different mutual
molecule positions have been found. The energy minima that correspond to three types of mutual
molecule positions, namely, (1) base positions with two N-H…O and/or N-H…N hydrogen bonds;
(2) nearly parallel arrangements of base ring planes, i.e. base stacking; and (3) T-shaped (nearly
perpendicular) base ring positions and hydrogen bond formation were extensively studied. The
majority of these minima were obtained by using the methods of different complexity, but the
method of calculation determines which minimum is the most deep (global) for certain base
combination, and relative depths of various minima. Analysis of these data, of our previous results
on simple DNA fragments, as well as of extensive reference data suggests that till now there is
no method suitable for quantitative description of all the experimental data on non-bonded
interactions in DNA. The comparison of energy and structure characteristics of complexes calculated
by various methods demonstrates their possibilities and shortcomings. A valid description of
non-bonded interactions of nucleic acids requires additional investigations of their simple fragments
and combined use of various methods.
Keywords: DNA, computer simulation, molecular mechanics, non-empirical calculations, density functional
theory
БИОФИЗИКА том 64 вып. 2 2019