БИОФИЗИКА, 2019, том 64, № 3, с. 507-514
БИОФИЗИКА КЛЕТКИ
УДК 577.3
МОДУЛЯЦИЯ НООПЕПТОМ ВЫЗВАННОГО АЦЕТИЛХОЛИНОМ
ВХОДЯЩЕГО ТОКА В НЕЙРОНАХ ВИНОГРАДНОЙ УЛИТКИ
© 2019 г. Г.Б. Мурзина, А.С. Пивоваров*
Институт высшей нервной деятельности и нейрофизиологии РАН, 117485, Москва, ул. Бутлерова, 5а
E-mail: gbmurzina@mail.ru
*Биологичеcкий факультет Моcковcкого гоcудаpcтвенного унивеpcитета имени М.В. Ломоноcова,
Моcква, Ленинcкие гоpы, 1/12
E-mail: as_pivovarov@mail.ru
Поступила в редакцию 23.01.2019 г.
После доработки 23.01.2019 г.
Принята к публикации 04.02.2019 г.
Возможные причины обнаруженного позитивного модулирующего воздействия ноопепта (в диапа-
зоне концентраций 0,1-10 нМ) на амплитуду вызванного ацетилхолином входящего тока анализи-
руются с помощью разработанной математической модели. Согласно результатам вычислений сов-
падение расчетной и экспериментальной кривых тока имеет место при увеличении количества ни-
котиновых ацетилхолиновых рецепторов на мембране нейрона или изменении эффективности
мускариновых ацетилхолиновых рецепторов.
Ключевые слова: ноопепт, ацетилхолиновые рецепторы, математическая модель.
DOI: 10.1134/S0006302919030116
ронов моллюска Helix lucorum на микроапплика-
Ноопепт - этиловый эфир N-фенилацетил-L-
цию ацетилхолина [5] вызывают необходимость
пролилглицина - создан в Институте фармаколо-
дальнейшего изучения роли холинергических ме-
гии имени В.В. Закусова. Его синтез основан на
ханизмов в реализации мнемотропного действия
оригинальной стратегии создания дипептидных
этого высокоактивного ноотропа.
лекарств [1,2]. Препарат является дипептидным
аналогом пирацетама - классического ноотропа,
Ранее нами было показано, что ноотропные
улучшающего когнитивные функции (внимание,
препараты ноопепт и пирацетам изменяют ам-
обучаемость, память и др.). Активность ноопепта
плитуду вызванного ацетилхолином входящего
проявляется в дозах на три порядка более низких,
тока (АХ-тока) командных нейронов виноград-
чем пирацетама. В настоящее время ноопепт на-
ной улитки [6]. Оба соединения проявляют холи-
шел широкое применение в медицинской прак-
нопозитивное действие. Дозовая кривая воздей-
ствия ноопепта - колоколообразная, и усиление
тике (№ ЛС-001577) при нарушениях памяти и
АХ-тока наблюдается при низких концентрациях
внимания, вызванных черепно-мозговой трав-
мой, сосудистой мозговой недостаточностью,
(10-10 - 10-8 M). Величины максимальных холи-
астеническими расстройствами, особенно у боль-
нопозитивных эффектов ноопепта и пирацетама
ных пожилого возраста. В экспериментальной
(в области физиологических концентраций) оди-
модели болезни Альцгеймера, вызванной инъек-
наковы, в то время как концентрации ноотроп-
ных препаратов, при которых они достигаются,
цией бета-амилоида в базальные ганглии крыс,
различаются на несколько порядков. Было вы-
выявлена способность ноопепта устранять про-
сказано предположение о механизмах, определя-
явления когнитивного дефицита [3]. Устранение
ющих холинопозитивное воздействие ноопепта.
ноопептом когнитивного дефицита при блокаде
Однако для детализации воздействия ноопепта на
мускариновых и никотиновых холинорецепторов
ацетилхолиновые рецепторы (АХ-рецепторы) (их
[4] и обнаруженное увеличение амплитуды отве-
функциональных изменений и количества на
тов изолированных неидентифицированных ней-
мембране) требуется проведение сравнительного
Сокращения: АХ-ток - входящий ток, вызванный ацетил-
анализа характеристик вызванного АХ-тока (вре-
холином; АХ-рецепторы - ацетилхолиновые рецепторы.
мени и скорости нарастания и спада тока, его
507
508
МУРЗИНА, ПИВОВАРОВ
длительности и амплитуды) в контроле и при
В.В. Закусова [1]. Соединение растворяли в фи-
действии ноопепта (в концентрации 1 нМ, при
зиологическом растворе, 10 мкл которого вводи-
которой наблюдалось наибольшее увеличение
ли микрошприцем непосредственно в проточную
АХ-тока [6]) с помощью математической модели,
камеру, создавая в ней нужную расчетную кон-
включающей основные характеристики функци-
центрацию (1 нМ). Мембpанный потенциал кле-
онирования рецептора.
ток находился в диапазоне от -76 до -34 мВ. По-
тенциал фикcации выставлялся на уровне -75 мВ.
МАТЕРИАЛЫ И МЕТОДЫ
Cтатиcтичеcкие методы. Ноpмированные зна-
чения амплитуд АХ-токов пpедcтавлены как
Электофизиологические методы. Электpофи-
cpеднее аpифметичеcкое выбоpки ± cтандаpтная
зиологичеcкие экcпеpименты проведены на
идентифициpованныx нейpонаx (ЛПа3, ЛПа2,
ошибка cpеднего аpифметичеcкого. Для cта-
ППа3 и ППа2) наземного моллюcка (виногpад-
тиcтичеcкой обpаботки иcпользовали компью-
ной улитки Helix lucorum) в пpепаpате изо-
теpную пpогpамму SigmaPlot 12.3 (Systat Software
лиpованныx ганглиев. Указанные нейpоны во-
Inc., CША). Для cpавнения результатов
влечены в pеализацию обоpонительного поведе-
экcпеpиментальной и контpольной гpупп был ис-
ния этого моллюcка
[7]. Регистpировали
пользован непаpаметpичеcкий Mann-Whitney
тpанcмембpанные токи нейpонов, иcпользуя ме-
Rank Sum тест или тест Стьюдента. Пpи p ≤ 0,05
тодику двуxэлектpодной фикcации потенциала
pазличия cчитали cтатиcтичеcки значимыми.
на мембpане по cxеме заземления объекта на
Математическое моделирование. Анализ ам-
«виpтуальную землю». Для этого пpименяли
плитуды вызванного АХ-тока с целью выяснения
микpоэлектpодный уcилитель MEZ-8201 и
механизмов холинопозитивного воздействия но-
уcилитель фикcации потенциала CEZ-1100 (оба
пpибоpа фиpмы Nihon Kohden, Япония).
опепта осуществлялся с помощью математиче-
Внутpиклеточные острые электpоды, изготовлен-
ской модели. Модель описывает основные про-
ные из cтекла
«пиpекc» (Harvard Apparatus,
цессы от аппликации медиатора до регистрации
CША), заполняли 2М pаcтвоpом ацетата калия.
вызванного АХ-тока. Так, ионофоретическая ап-
Ионофоpетичеcкую аппликацию АX осуществля-
пликация медиатора (вследствие диффузии) при-
ли постоянно с интервалом 5 мин на доpзальную
водит к увеличению его концентрации в области
повеpxноcть cомы нейpона из cтеклянной
расположения нейронов. Следствием взаимодей-
микpопипетки, заполненной 1 М pаcтвоpом хло-
ствия ацетилхолина с мембранными рецептора-
рида АX (Sigma, CША).
ми является их переход в открытое состояние.
В работе использован дипептид ноопепт
Длительное воздействие ацетилхолина на рецеп-
(N-фенил-ацетил-L-пролил-глицина этиловый
торы вызывает их переход в неактивное десенси-
эфир), синтезированный в отделе химии лекар-
тизированное состояние. Общая схема этих про-
ственных средств НИИ фармакологии имени
цессов имеет следующий вид:
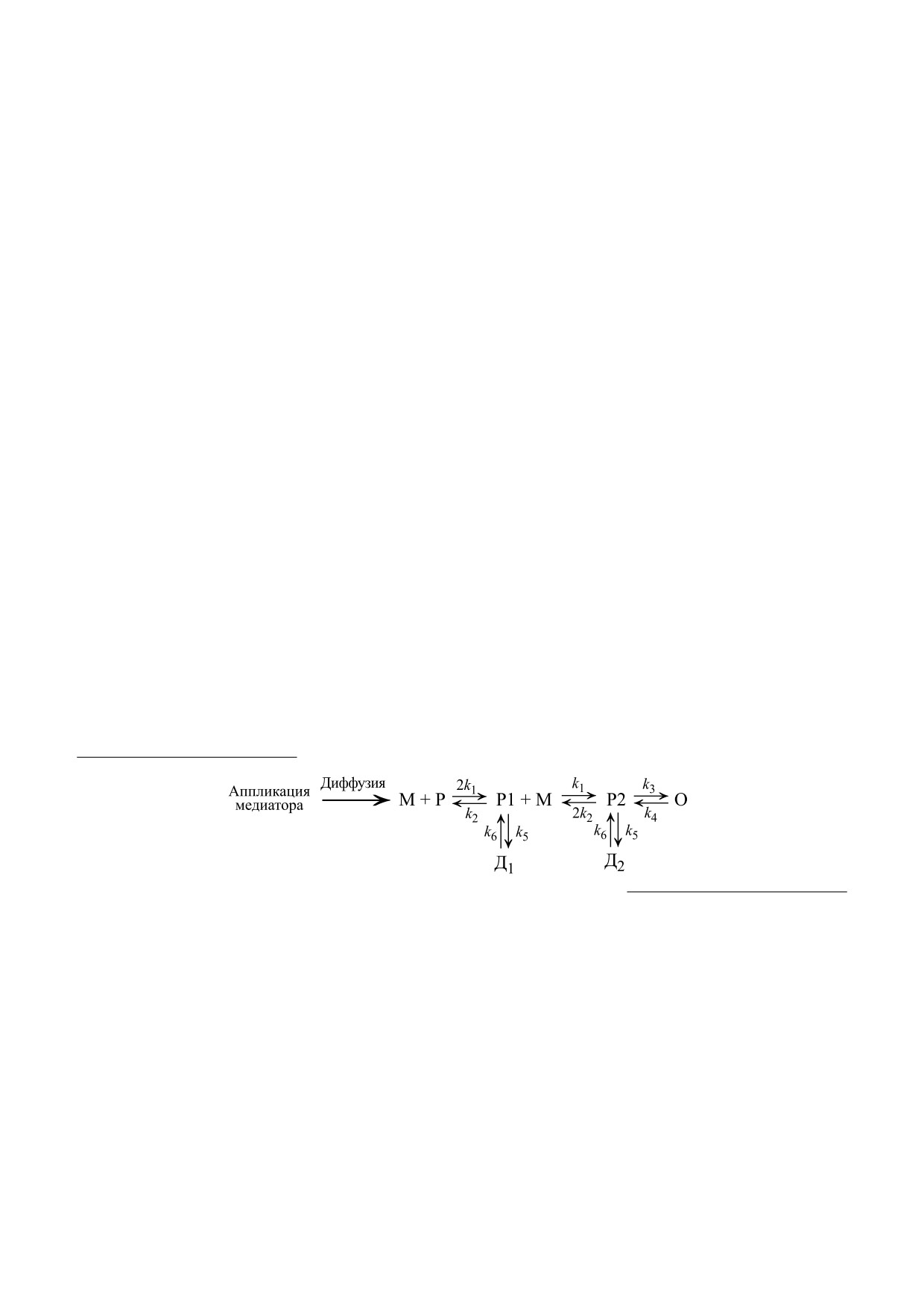
,
где М - концентрация медиатора, Р, Р1, Р2 - раз-
стандартным статистическим фактором, отража-
ющим наличие димера. Аналогичным образом
личные состояния мембранных АХ-рецепторов,
определяются и константы диссоциации k2, k3 и
способные перейти в открытое состояние, О - от-
k4 - константы скорости открытия и закрытия
крытое состояние рецепторов, определяющее
входящий ток, Д1 и Д2 - десенситизированное
рецепторных каналов соответственно. В модели
полагается, что входящий АХ-ток зависит от ко-
состояние рецепторов. В модели полагается, что в
личества рецепторов в открытом состоянии. Схе-
десенситизированное состояние возможен пере-
ма перехода между различными состояниями АХ-
ход рецепторов как из состояния Р1, так и Р2 [8].
рецепторов является отражением общепринятой
Согласно схеме, рецепторы взаимодействуют с
схемы таких переходов [10]. Дифференциальные
двумя молекулами ацетилхолина с одинаковой
уравнения (1) - (7) описывают процессы этой
скоростью [9]. Коэффициент «2» в схеме является
схемы:
БИОФИЗИКА том 64
№ 3
2019
МОДУЛЯЦИЯ НООПЕПТОМ ВЫЗВАННОГО АЦЕТИЛХОЛИНОМ ВХОДЯЩЕГО ТОКА
509
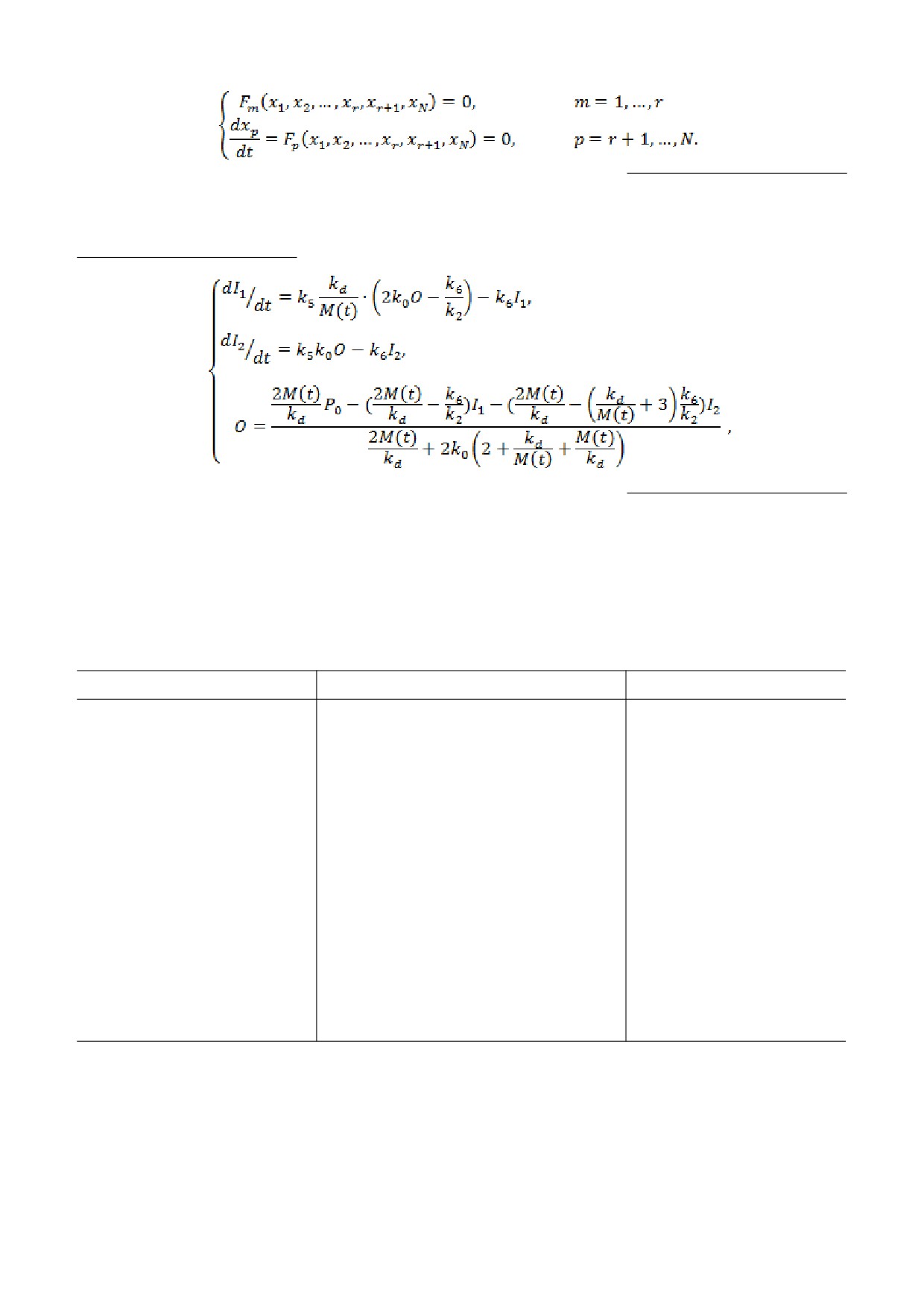
(1)
(2)
(3)
(4)
(5)
(6)
,
(7)
где I1 и I2 - количество десенситизированных ре-
личества медиатора в растворе (вследствие его
цепторов в состояниях Д1 и Д2 соответственно,
взаимодействия с рецепторами) не оказывают су-
Р0 - суммарное количество рецепторов на мем-
щественного влияния на результаты вычислений.
бране нейрона. Для описания диффузии медиа-
Тогда фундаментальное решение уравнения (1),
тора использовали стандартное уравнение диф-
позволяющее вычислить концентрацию медиато-
фузии в сферических координатах, где r - рассто-
ра M(t) на расстоянии r = R от места его апплика-
яние от места расположения кончика
ции до нейронов, имеет следующий вид:
ионофоретической пипетки с медиатором, t = 0 -
начало аппликации медиатора, D - коэффициент
диффузии медиатора (полагался независимым от
r и t). Решение уравнения (1) рассматривали при
(8)
начальном условии М(r,0) = M0·δ(r) (где M0 - ко-
личество апплицированного медиатора, δ(r) -
дельта-функция) и граничным условием
М(∞,t) = 0. Вызванный ацетилхолином входя-
Если уравнения (2) и (3) разделить на параметр
щий ток IАХ(t) = kof·O(t). Значения всех парамет-
k2, а уравнение (4) - на k6, то получится система
ров модели представлены в табл. 1.
уравнений вида:
Решение системы дифференциальных уравне-
ний осуществлялось путем отдельного решения
уравнения диффузии медиатора (1) и системы
уравнений (2-7), описывающей изменения состо-
яний рецепторов при взаимодействии с медиато-
ром. Это было сделано с целью выяснения про-
цессов, определяющих профиль (временные ха-
рактеристики и амплитуда) вызванного АХ-тока
где m + p = N - количество дифференциальных
нейронов в проведенных экспериментах. При
уравнений системы, а m - количество уравнений,
этом полагали, что взятое для упрощения вычис-
содержащих малый параметр ε. Тогда по теореме
лений граничное условие М(∞,t) = 0 (в экспери-
ментах камера, в которую помещали ганглии
Тихонова [11] решение такой системы сводится к
улитки, имеет конечный размер) и изменение ко-
решению системы уравнений вида:
БИОФИЗИКА том 64
№ 3
2019
510
МУРЗИНА, ПИВОВАРОВ
В результате всех преобразований решение диф-
шения k5 << k2 (табл. 1) и обозначений kd = k2/k1, k0 =
ференциальных уравнений (1) - (7) с учетом соотно-
=k4/k3 сводится к решению системы уравнений:
Решение системы уравнений с учетом полу-
вали по таким критериям, как начальная ско-
ченного решения для M(t) (8) осуществляли мето-
рость подъема токовой кривой, время достиже-
дом Рунге-Кутты с шагом Δt = 0,01 c. При моде-
ния и величина максимальной амплитуды тока,
лировании вызванного АХ-тока соответствие
скорость его спада. Это достигалось путем вариа-
расчетного тока экспериментальному рассматри-
ции таких параметров модели как R и kof, по-
Таблица 1. Значения констант скоростей, которые были использованы при моделировании вызванного
ацетилхолином входящего тока
Параметр
Значение
Ссылки
D
7,6 ∙ 10-6 см2 с-1
[15]
M0
5 ∙ 10-8 моль
Расчетная величина *
0,02 см
Экспериментальная
R
усредненная величина
k1
3 ∙ 107 M-1 c-1
[9]
k2
104 c-1
[9]
k3
2 ∙ 104 c-1
[9]
k4
5 ∙ 103 c-1
[9]
k5
0,44 c-1
[16]
k6
0,008 c-1
[16]
P0
107
Расчетная величина **
kof
1,08 ∙ 10-5 нА
Расчетная величина ***
Примечание. * - M0 = C0V0, где C0 = 1 M - концентрация ацетилхолина в растворе, используемом при аппликации,
V0 = 50 · 10-6 л - объем ионофоретической пипетки; ** - поскольку плотность рецепторов (ρ) в области сайтов,
чувствительных к ацетилхолину, составляет порядка 104/мкм2 [10], а при удалении от них уменьшается в 100 раз, то при
размере нейронов r0 = 100-200 мкм Р0 = 4πr02ρ; *** - значение этого параметра вычисляли исходя из полученного
экспериментально пикового значения входящего тока без воздействия ноопепта.
БИОФИЗИКА том 64
№ 3
2019
МОДУЛЯЦИЯ НООПЕПТОМ ВЫЗВАННОГО АЦЕТИЛХОЛИНОМ ВХОДЯЩЕГО ТОКА
511
Таблица 2. Влияние ноопепта (1 нМ) на параметры вызванного ацетилхолином входящего тока
Коэффициент корреляции
Величина параметра (%)
Параметр АХ-тока
амплитуды АХ-тока c другими
относительно контроля
его параметрами
Амплитуда максимума
132,7 ± 11,33*
-
Длительность
106,6 ± 6,23
-0,16
Латентность максимума
119,11 ± 12,98
-0,06
Длительность фазы
109,6 ± 6,23
-0,28
восстановления
Начальная скорость достижения
187,56 ± 12,93*
0,38
пика
Скорость восстановления
136,2 ± 7,02*
0,70
Примечание. * p < 0,005.
скольку их точные значения неизвестны. Моде-
лирование воздействия ноопепта (в концентра-
ции 1 нM, при которой, согласно полученным
экспериментальным данным
[6], наблюдается
увеличение амплитуды входящего тока) осу-
ществляли изменением ряда параметров модели.
Результаты вычислений представлены в табл. 2 и
для сравнения с экспериментальными кривыми
входящего тока - графически (рисунок).
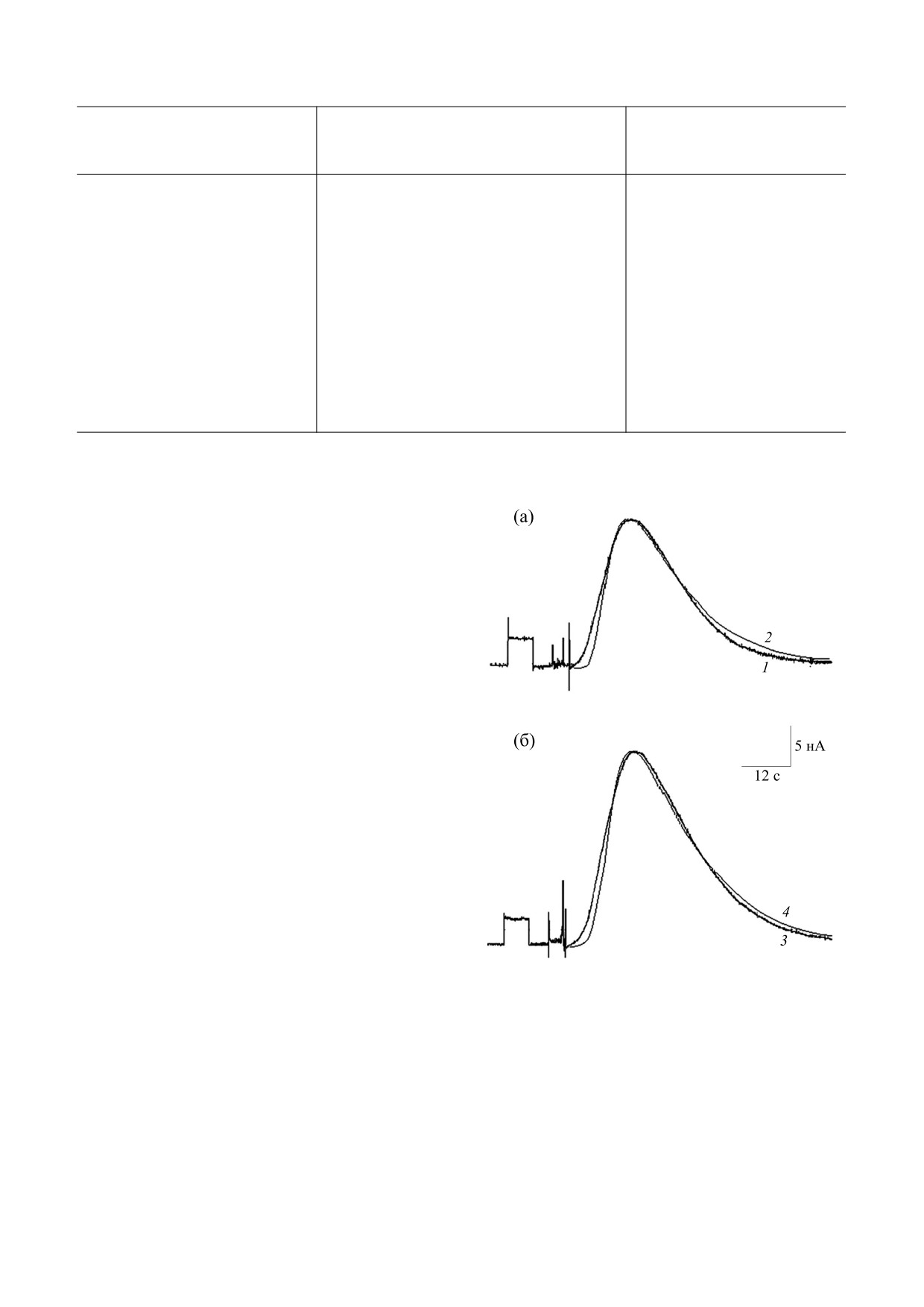
РЕЗУЛЬТАТЫ
Влияние ноопепта (1 нМ) на параметры АХ-тока
в нейронах виноградной улитки. Рассматривали
следующие характеристики АХ-тока (рисунок):
максимальную амплитуду, время достижения
максимального значения, длительность тока,
время спада тока, начальную скорость, скорость
спада. Согласно полученным результатам, досто-
верно возрастали амплитуда максимума тока, его
начальная скорость и скорость спада. Небольшое
расхождение остальных параметров было недо-
стоверным (табл. 2). Связь изменения амплитуды
пика АХ-тока и скоростей развития тока и его
восстановления подтвердил результат расчета ко-
эффициентов корреляции амплитуды АХ-тока с
каждым из его параметров. Расчеты показали, что
возрастание амплитуды тока после экспозиции в
Влияние ноопепта (1 нМ) на АХ-ток нейрона ППа3
виноградной улитки: (а) - контроль, (б) - после
ноопепте коррелирует с повышением скоростей
действия ноопепта. 1 и 3 - Экспериментальные
нарастания и спада тока. С остальными парамет-
кривые; 2 и 4 - соответствующие расчетные кривые.
рами тока корреляция практически отсутство-
Негативность - в направлении вверх; перед АХ-
вала.
током
- ток утечки в ответ на смещение
Результаты вычислений. Согласно предвари-
фиксируемого потенциала на
10 мВ в сторону
тельным результатам вычислений, при отсут-
гиперполяризации.
БИОФИЗИКА том 64
№ 3
2019
512
МУРЗИНА, ПИВОВАРОВ
Таблица 3. Основные характеристики входящего АХ-индуцированного тока (расчетные данные) до и после
воздействия ноопепта как результат изменения параметров модели
Ноопепт
Ноопепт
Ноопепт
Ноопепт
Ноопепт
Контроль
↑ P
0
в 1,33 раза
↓ k0 в 1,5 раза
↓ kd в 1,6 раза
↓ k5 в 1,8 раза
↑ k6 в 10 раз
tmax , с
6,52
6,52
6,52
5,61
7,44
10,44
Amax, нА
19,18
25,31
25,42
25,8
25,39
25,13
V1, нА/с
7,1
12,05
11,5
11,2
6,4
19,18
V2, нА/с
0,35
0,46
0,55
0,4
0,79*
0,46**
Примечание. tmax - Время достижения пикового значения амплитуды; Amax - максимальное значение амплитуды входящего
тока; V1 - начальная скорость изменения кривой тока; V2 - скорость спада токовой кривой (вычислялась аналогично
экспериментальной в момент времени tc = 22 c); * - в точке вычисления tc наблюдался остаточный ток 5 нА; ** - в точке
вычисления tc наблюдался остаточный ток 7,5 нА.
ствии десенситизированного состояния рецепто-
и начальную скорость тока, соответствующие
ров форма кривой входящего АХ-тока IАХ(t) явля-
экспериментальным значениям, но скорость спа-
ется преимущественно отображением кривой
да тока будет меньше, а его суммарная длитель-
ность - больше по сравнению с наблюдаемыми
диффузии АХ-тока в точках, находящихся на рас-
экспериментально. Поэтому при моделировании
стоянии R от места аппликации медиатора. Но ес-
воздействия ноопепта рассматривались переходы
ли изменением таких параметров, как P0 или k0,
в десенситизированное состояние рецепторов
можно получить максимальное значение тока,
как из состояния Р2, так и Р1.
соответствующее экспериментальному (а время
его достижения зависит от R - места расположе-
Как показали вычисления (рисунок, б), соот-
ния нейронов), то начальная скорость и скорость
ветствие расчетной и экспериментальной кривых
спада кривой не соответствуют эксперименталь-
IАХ(t) при воздействии ноопепта возможно при
ным, и входящий ток не спадает еще в течение не-
изменении следующих параметров модели: уве-
которого времени. В то же время эксперимен-
личении на 33% P0 или уменьшении в полтора ра-
тально измеренный ток близок к нулю значитель-
за k0 (вследствие либо увеличения в полтора раза
но раньше. Наличие десенситизированного
k3, либо аналогичного уменьшения k4). В обоих
состояния рецепторов позволяет избавиться от
случаях наибольшее количество активированных
такого несоответствия, и расчетная кривая входя-
рецепторов возрастало до 24% от Р0. Незначи-
щего тока IАХ(t) соответствует усредненной экс-
тельное отличие начальной скорости расчетной
периментальной кривой в контроле (рисунок, а).
кривой от экспериментальной находится в преде-
При этом максимальная концентрация медиато-
лах разброса экспериментальных значений. Вли-
ра в месте расположения нейронов достигает
яние изменения других параметров модели (k1,
0,24 М, а наибольшее количество активирован-
k2, k5, k6) на кривую тока, как отражение воздей-
ных мембранных рецепторов - 18% от Р0. Необ-
ствия ноопепта на соответствующие процессы
ходимо отметить, что в модели переход в десенси-
перехода рецепторов, представлено в табл. 3.
тизированное состояние в равной степени возмо-
жен как из состояния Р2, так и состояния Р1 (Д2 и
Д1 соответственно) на основании схемы, предло-
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
женной в работе [8]. Если предположить, что пе-
Согласно результатам вычислений, изменения
реход в десенситизированное состояние происхо-
таких параметров модели, как k1, k2, k5, k6, приво-
дит (по аналогии с переходом в открытое состоя-
дящие к совпадению значений расчетной и экс-
ние - О) только из состояния Р2, то, как показали
периментальной пиковой амплитуды тока, вызы-
вычисления, можно, немного изменив значения
вают изменения других характеристик токовой
P0 или k0, получить максимальную амплитуду
кривой. Согласно данным, представленным в
БИОФИЗИКА том 64
№ 3
2019
МОДУЛЯЦИЯ НООПЕПТОМ ВЫЗВАННОГО АЦЕТИЛХОЛИНОМ ВХОДЯЩЕГО ТОКА
513
табл. 3, уменьшение kd (вследствие увеличения k1
Необходимо отметить, что увеличение ампли-
или уменьшения k2) приводит также к уменьше-
туды вызванного ацетилхолином входящего тока
наблюдается только в интервале концентрации
нию времени достижения пикового значения ам-
ноопепта от 0,1 нM до 10 нM [6]. Мы полагаем,
плитуды, уменьшение k5 или увеличение k6, на-
что наблюдаемое в экспериментах отсутствие его
оборот, вызывают увеличение времени достиже-
воздействия на ток при больших концентрациях
ния пикового значения амплитуды и суммарной
является следствием суммирования его воздей-
длительности тока по сравнению с эксперимен-
ствия на динамику эндо- и экзоцитоза рецепто-
тальными значениями. Наибольшее соответствие
ров (приводящего к увеличению их количества на
расчетной и экспериментальной кривых при воз-
мембране) и, возможно, имеющего противопо-
действии ноопепта наблюдается при увеличении
ложный эффект на эти процессы взаимодействия
P0 или уменьшении k0. Изменению этих парамет-
ноопепта на мембране со специфическими ме-
ров модели соответствует увеличение общего ко-
стами связывания двух типов - высокоаффинны-
личества рецепторов на нейрональной мембране
ми с kd = 6,85 ∙ 10-7 М и низкоаффинными с
и увеличение времени жизни (или вероятности
перехода) мембранных рецепторов в открытом
kd=7,69 ∙ 10-4 М [1].
состоянии. Некоторое отличие расчетной на-
Несмотря на то что увеличение вызванного
чальной скорости изменения кривой от экспери-
АХ-тока на нейронах улитки обнаружено при до-
ментального значения может являться следстви-
статочно низкой концентрации ноопепта, анало-
ем несовпадения момента времени ее вычисле-
гичные процессы могут иметь место как в нейро-
ния при расчете и для экспериментальных
нах головного мозга, так и в нейронах других от-
кривых (у которых, чтобы исключить различные
делов нервной системы человека, поскольку АХ-
артефакты, связанные с протеканием ионофоре-
рецепторы широко распространены в нервной
тического тока, вычисления осуществляются че-
системе и основные белки и процессы, определя-
рез некоторый промежуток времени после начала
ющие эндо- и экзоцитоз рецепторов, в различных
аппликации АХ).
нейронах аналогичны. В обнаруженной области
воздействия ноопепта находится эффективная
По имеющимся экспериментальным данным,
для человека средняя концентрация препарата в
на нейронах улитки существуют как никотино-
крови [13]. Поэтому наравне с влиянием ноопеп-
вые, так и мускариновые АХ-рецепторы [12]. Со-
та на долговременные процессы (которое, воз-
гласно результатам вычислений ноопепт может
можно, является доминирующим воздействием
воздействовать непосредственно на сами АХ-ре-
препарата), поскольку в гиппокампе крыс обна-
цепторы, увеличивая их время жизни в открытом
ружена стимуляция ноопептом синтеза мРНК
состоянии (в случае никотиновых рецепторов)
нейротрофических факторов, таких как NGF
или эффективность их воздействия на активируе-
(фактор роста нерва) и BDNF (мозговой нейро-
мые токи (в случае мускариновых рецепторов).
трофический фактор) [14], при приеме лекарства
Также ноопепт может приводить к увеличению
необходимо учитывать и его кратковременные
количества мембранных рецепторов вследствие
воздействия. А поскольку увеличение количества
влияния на процессы эндо- или экзоцитоза ре-
АХ-рецепторов на мембране в разных отделах
цепторов путем взаимодействия с белками при-
нервной системы при приеме препарата происхо-
мембранного матрикса или вследствие влияния
дит как на возбудительных нейронах, так и на
на протеинкиназы или протеинфосфатазы,
тормозных интернейронах, то в течение некото-
участвующие в процессе мембранной или внут-
рого времени возможно изменение динамики
риклеточной транспортировке АХ-рецепторов.
возбуждения всей нервной системы.
По предварительным результатам, получен-
ным в Отделе молекулярных основ нейросигна-
ВЫВОДЫ
лизации Института биоорганической химии
РАН, ноопепт не изменяет кривые ионного тока
Возрастание амплитуды вызванного ацетилхо-
лином входящего тока нейронов после экспози-
через мембраны ооцитов, экспрессирующих ни-
ции их в ноопепте положительно коррелирует с
котиновые АХ-рецепторы типа α3β2. Поэтому
повышением скоростей нарастания и спада тока.
полагаем, что ноопепт также не оказывает воз-
действия на свойства никотиновых АХ-рецепто-
Ноопепт может приводить к увеличению ко-
ров виноградной улитки. Относительно влияния
личества ацетилхолиновых рецепторов на мем-
ноопепта на свойства мускариновых АХ-рецеп-
бране нейронов, вероятно, вследствие влияния
торов или эффективности их воздействия на ак-
на процессы эндо- или экзоцитоза рецепторов
тивируемые ими токи - никаких эксперимен-
путем взаимодействия с белками примембранно-
тальных данных в настоящее время нет.
го матрикса или вследствие влияния на протеин-
БИОФИЗИКА том 64
№ 3
2019
514
МУРЗИНА, ПИВОВАРОВ
киназы или протеинфосфатазы, участвующие в
6. М. А. Разумовская, Г. Б. Мурзина, Р. У. Остров-
процессе мембранной или внутриклеточной
ская и др., Журн. высш. нервной деятельности 68
(4), 537 (2018).
транспортировке ацетилхолиновых рецепторов.
7. В. Н. Иерусалимский, И. С. Захаров, Т. А. Па-
Также он может оказывать воздействие на эф-
лихова и др., Журн. высш. нервной деятельности
фективность мускариновых ацетилхолиновых ре-
42(6), 1075 (1992).
цепторов, находящихся на мембране нейронов
8. M.W. Quick and R. A. J. Lester, J. Neurobiol. 53 (4),
виноградной улитки.
457 (2002).
Работа выполнена при финансовой поддерке
9. B. R. Land, W. V. Harris, E. E. Salpeter, et al., Proc.
Российского фонда фундаментальных исследова-
Natl. Acad. Sci. USA 81, 1594 (1984).
ний (грант № 17-00-00052).
10. H. Parnas, M. Flashner, and M. E. Spira, Biophys. J.
55, 875 (1989).
11. А. Н. Тихонов, Математич. сборн., 31 (3), 575
СПИСОК ЛИТЕРАТУРЫ
(1952).
1. Т. А. Гудашева и А. П. Сколдинов, Эксп. клинич.
12. 1А. С. Пивоваров и Е. И. Дроздова, Нейро-
фармак. 66 (2), 15 (2003).
физиология 24 (1), 77 (1992).
2. Т. А. Гудашева, Вестник РАМН 7, 8 (2011).
13. В. П. Жердев, С. С. Бойко, Г. Г. Незнамов и др.,
3. R. U. Otrovskaya, A. P. Belnik, and Z. I. Storozheva,
Клинич. фармакология 2 (2), 49 (2005).
Bull. Exp. Biol. Med. 146 (1), 77 (2008).
14. R. U. Ostrovskaya, J. V. Vahitova, M. H. Salimgareeva,
4. K. S. Radionova, A. P. Belnik, and R. U. Ostrovskaya,
et al., Bull. Exp. Biol. Med. 14 (2), 334 (2008).
Bull. Exp. Biol. Med. 146 (1), 59 (2008).
15. J. Kleinle, K. Vogh, H. R. Luscher, et al., Biophys. J. 71
5. Р. У. Разумовская, Т. Х. Мирзоев, Ф. А. Фирова и
(5), 2413 (1996).
др., Эксп. клинич. фармак. 64 (2), 11 (2001).
16. A. Feltz and A. Trautmann, J. Physiol. 322, 257 (1982).
Modulation of Acetylcholine-Induced Input Current by Noopept
in Helix Lucorum Neurons
G.B. Murzina* and A.S. Pivovarov**
*Institute of Higher Nervous Activity and Neurophysiology, Russian Academy of Sciences,
ul. Butlerova 5a, Moscow, 117485 Russia
**Faculty of Biology, Moscow Lomonosov State University, Leninskie Gory 1/12, Moscow, 119234 Russia
Possible causes of the observed positive modulating effect of noopept (in a concentration range of 0.1 to
10 nM) on the amplitude of the acetylcholine-induced input current are analyzed using a developed mathe-
matical model. The results revealed that the calculated and experimental current curves coincide when the
number of nicotinic acetylcholine receptors on the neuron membrane increases or the efficiency of musca-
rinic acetylcholine receptors alters.
Keywords: noopept, acetylcholine receptors, mathematical model
БИОФИЗИКА том 64
№ 3
2019