БИОФИЗИКА, 2019, том 64, № 3, с. 563-571
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 519.876.5
МОДЕЛИРОВАНИЕ ФЕНОЛОГИИ СОИ С ПОМОЩЬЮ
ИСКУССТВЕННЫХ НЕЙРОННЫХ СЕТЕЙ
© 2019 г. О.Д. Таратухин*, Л.Ю. Новикова* **, И.В. Сеферова**, К.Н. Козлов*
*Санкт-Петербургский политехнический университет Петра Великого,
195251, Санкт-Петербург, Политехническая ул., 29
**Федеральный исследовательский центр «Всероссийский институт генетических ресурсов растений
имени Н.И. Вавилова», 190000, Санкт-Петербург, ул. Большая Морская, 42-44
E-mail: kozlov_kn@spbstu.ru
Поступила в редакцию 11.02.2019 г.
После доработки 11.02.2019 г.
Принята к публикации 25.02.2019 г.
Индикатором климатических изменений и степени пригодности сои к климатическим условиям
является ее фенология, основные регуляторы которой
- температура и длина дня. Для
предсказания времени до цветения у девяти скороспелых образцов сои с пониженной
фотопериодической чувствительностью в условиях Северо-Западного региона России была
адаптирована модель на основе искусственной нейронной сети. Введены масштабирующие
множители для сетевых входов, оптимизированы верхняя и нижняя температурные границы и
базовая длина дня, создана программа на языке Питон с использованием Керас и ТензорФлоу.
Экспериментальные данные, полученные в 1999-2013 гг. в Пушкине (Ленинградская область) и на
Кубани были использованы для обучения 121 параметра модели, по завершению которого
среднеквадратичная ошибка снизилась до 0,026. Образцы характеризовались пониженной верхней
температурной границей по сравнению с литературными данными
(23°С вместо
30°С) и
повышенной нижней температурной границей (12°С вместо 5°С). Увеличение базовой длины дня с
12 до 13 часов подтвердило адаптированность образцов к более длинному дню. Средняя ошибка
прогноза улучшилась примерно на двое суток по сравнению с предыдущей моделью температурных
минимумов. Были сгенерированы ежедневные погодные данные для различных будущих сценариев
выброса парниковых газов и предсказаны времена до цветения девяти образцов сои в условиях
изменения климата в период с 2019 г. до 2030 г. для посевов 1 и 10 мая. Предсказанные значения
уменьшаются до 2030 г. для большинства образцов и сценариев, но могут оставаться постоянными
или в некоторых случаях испытывать колебания. Различие средних между 39,21 суток в данных и
36,33 суток в результатах моделирования для 2030 г. является статистически значимым согласно
одностороннему тесту Манна-Уитни-Уилкоксона (5423,5, P = 0,0097 < 0,01). Таким образом,
результаты подтвердили предсказательную силу разработанной модели.
Ключевые слова: климатичеcкие фактоpы, cоя, искусственные нейронные сети.
DOI: 10.1134/S0006302919030177
Одной из мер минимизации потерь от потеп-
областей умеренных широт. Основными регуля-
ления климата и превращения их в прибыль явля-
торами ее фенологии являются температуры и
ется осеверение сортов [1]. В условиях глобаль-
длина дня. Продвижению культуры на север ме-
ных и региональных климатических трендов на-
шают низкие температуры и длинный день [5]. С
учной основой для изучения механизмов
2000-х годов посевы под соей в России неуклонно
адаптации растений к изменению условий обита-
расширяются (Госкомстат России). В ФИЦ «Все-
ния является математическое моделирование и
российский институт генетических ресурсов рас-
прогнозирование [2,3]. Индикатором климатиче-
тений имени Н.И. Вавилова» (ВИР) много лет
ских изменений и степени пригодности сортов к
проводятся опыты по осеверению сои, выводятся
складывающимся климатическим условиям яв-
сорта с пониженной фотопериодической чув-
ляется фенология растения [4].
ствительностью и пониженными температурным
Соя является одной из наиболее рентабельных
потребностями.
с/х культур, потребность в продуктах переработ-
Разработанные в условиях последней четверти
ки сои продолжает расти. Соя - растение южных ХХ века модели нуждаются в параметризации
563
564
ТАРАТУХИН и др.
Таблица 1. Сравнение точности новой модели с моделью температурных минимумов [11]
Средняя ошибка пpогноза
Средняя ошибка
Коэффициент
R2 - коэффициент
Образец
в модели температурных
пpогноза новой
корреляции Пирсона
детерминации
минимумов, cут
модели, cут
ПЭП 28
4,0
2,4
0,87
0,75
Cветлая
3,9
2,9
0,87
0,75
ПЭП 2
4,1
1,8
0,92
0,84
ПЭП 18
5,9
1,9
0,91
0,84
ПЭП 27
3,9
2,5
0,88
0,78
Fiskeby 1040-4-2
3,6
2,0
0,94
0,87
Окcкая
5,6
2,9
0,86
0,74
Алтом
8,0
3,1
0,83
0,70
KG-20
7,4
2,5
0,94
0,89
Cpеднее
5,2
3,3
0,82
0,69
значение
применительно к новым условиям. Существует
Целью данной работы было исследование
множество моделей фенологии сои. Математиче-
применимости модели нейронных сетей к моде-
ское моделирование важных агрономических по-
лированию фенологии сои в условиях низких
казателей, таких как, например, длина периода
температур и длинного дня Северо-Западного ре-
«всходы-цветение», может осуществляться раз-
гиона России.
личными способами. Строятся регрессионные
модели с различными наборами предикторов [6].
ОБЪЕКТ ИССЛЕДОВАНИЯ
Наиболее распространенный - расчет суммы на-
И ЭКСПЕРИМЕНТАЛЬНЫЕ ДАННЫЕ
копленных «единиц тепла» за сутки. Число дней
между фазами развития находится суммировани-
Были исследованы девять скороспелых образ-
ем вклада каждого дня до достижения суммы, не-
цов сои с пониженной фотопериодической чув-
обходимой для завершения фазы. Скорость раз-
ствительностью, перспективные для продвиже-
вития за день рассчитывается как произведение
ния в северные регионы (таблица). В качестве ис-
функций в основном от длины дня и средней су-
ходных были использованы данные по длине
точной температуры воздуха [7-10]. Для исследо-
периода «всходы-цветение» у этих сортов, полу-
вания развития строятся динамические и имита-
ченные на опытных станциях ВИР в г. Пушкин
ционные модели [11].
(Ленинградская область) и на Кубани в 1999-
2013 гг. Проводились опыты с несколькими посе-
Искусственные нейронные сети широко при-
вами в течение года (от 2 до 15), что позволило
меняются в машинном обучении в различных об-
расширить диапазон условий температуры и фо-
ластях, в том числе для предсказания важных аг-
топериода.
рономических показателей [12-14]. Модели на
основе нейронных сетей основываются на кон-
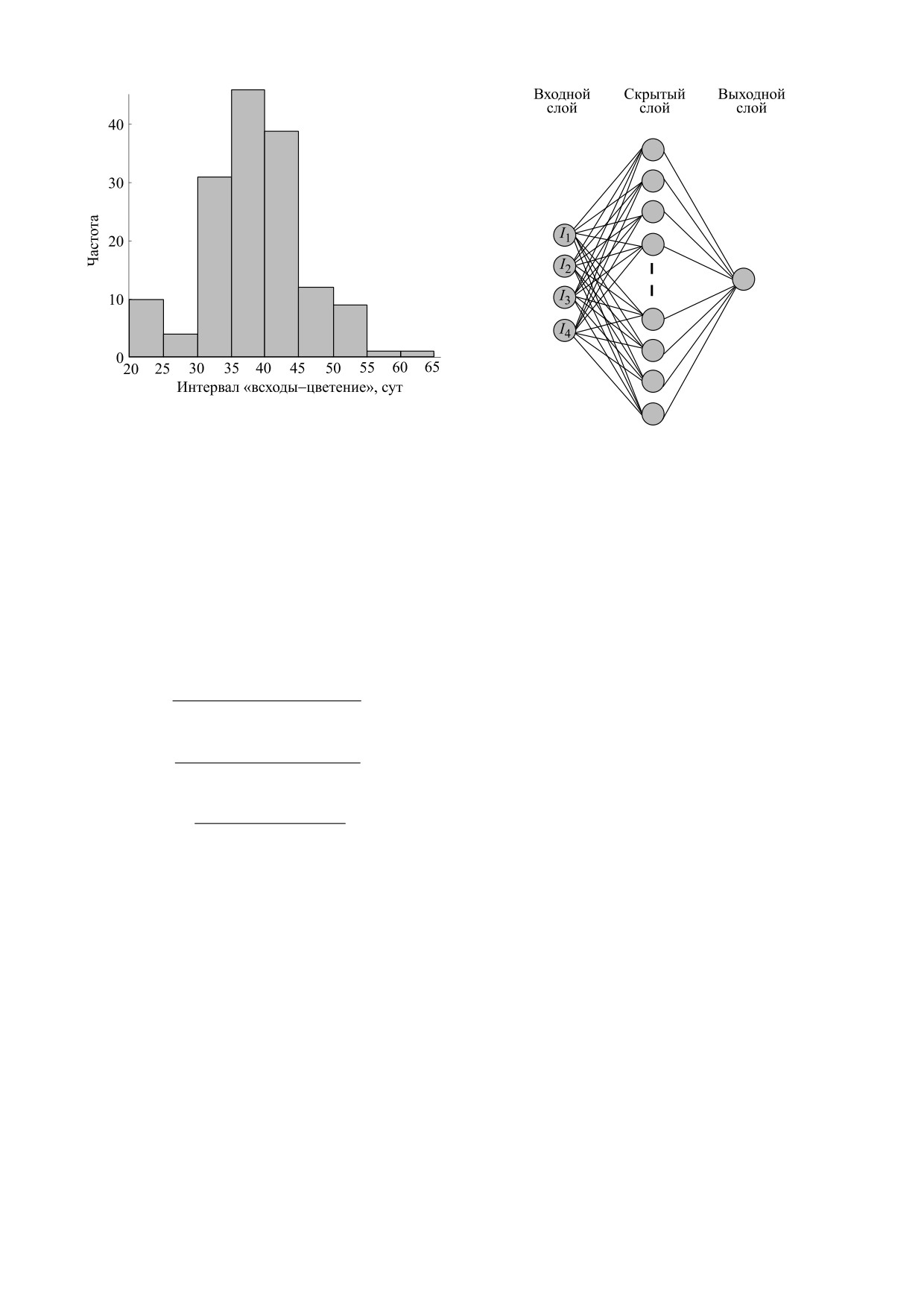
На рис. 1 представлена гистограмма длин пе-
цепции слоев и перцептронов. Нейронная сеть
риода в наборе данных.
состоит из входного слоя, произвольного числа
промежуточных слоев и выходного слоя. Размер-
МОДЕЛЬ ВРЕМЕНИ ЦВЕТЕНИЯ
ности и количество скрытых слоев определяются
НА ОСНОВЕ НЕЙРОННЫХ СЕТЕЙ
исследователем, который учитывает объем имею-
щихся данных и их качество. Модели на основе
В работе [15] предложена модель на основе
нейронных сетей используются и для исследова-
нейронной сети, которая состоит из 4 входных
ния фенологии [15].
нейронов,
20 перцептронов в единственном
БИОФИЗИКА том 64
№ 3
2019
МОДЕЛИРОВАНИЕ ФЕНОЛОГИИ СОИ
565
Рис. 1. Гистограмма длин периода «всходы-цвете-
ние» в наборе данных.
Рис. 2. Нейронная сеть модели времени цветения.
скрытом слое и единственного выходного нейро-
на (см. рис. 2).
Конкретные значения параметров должны
На входы сети I1, I2, I3, I4 подаются результаты
быть выяснены в ходе обучения сети для достиже-
применения специально подобранных в работе
ния наилучших результатов.
[15] функций (1)-(4), которые обобщают влияние
ежедневных показателей максимальной и мини-
РЕЗУЛЬТАТЫ АДАПТАЦИИ МОДЕЛИ
мальной температуры и длины светового дня на
состояние растения:
Модель была запрограммирована на языке
Питон с использованием Keras [16]и TensorFlow
[17] в качестве вычислителя. Данные о ежеднев-
∑H
(T
m,i
−T
m,c
)(T
m,i
−T
m,c
)
I
1
=
,
(1)
ных максимальных и минимальных температу-
μ
m
рах, осадках и длине дня были взяты в открытом
доступе в сети Интернет. Обучение сети состоит в
∑H
(T
x,i
−T
x,c
)(T
x,i
−T
x,c
)
I
=
(2)
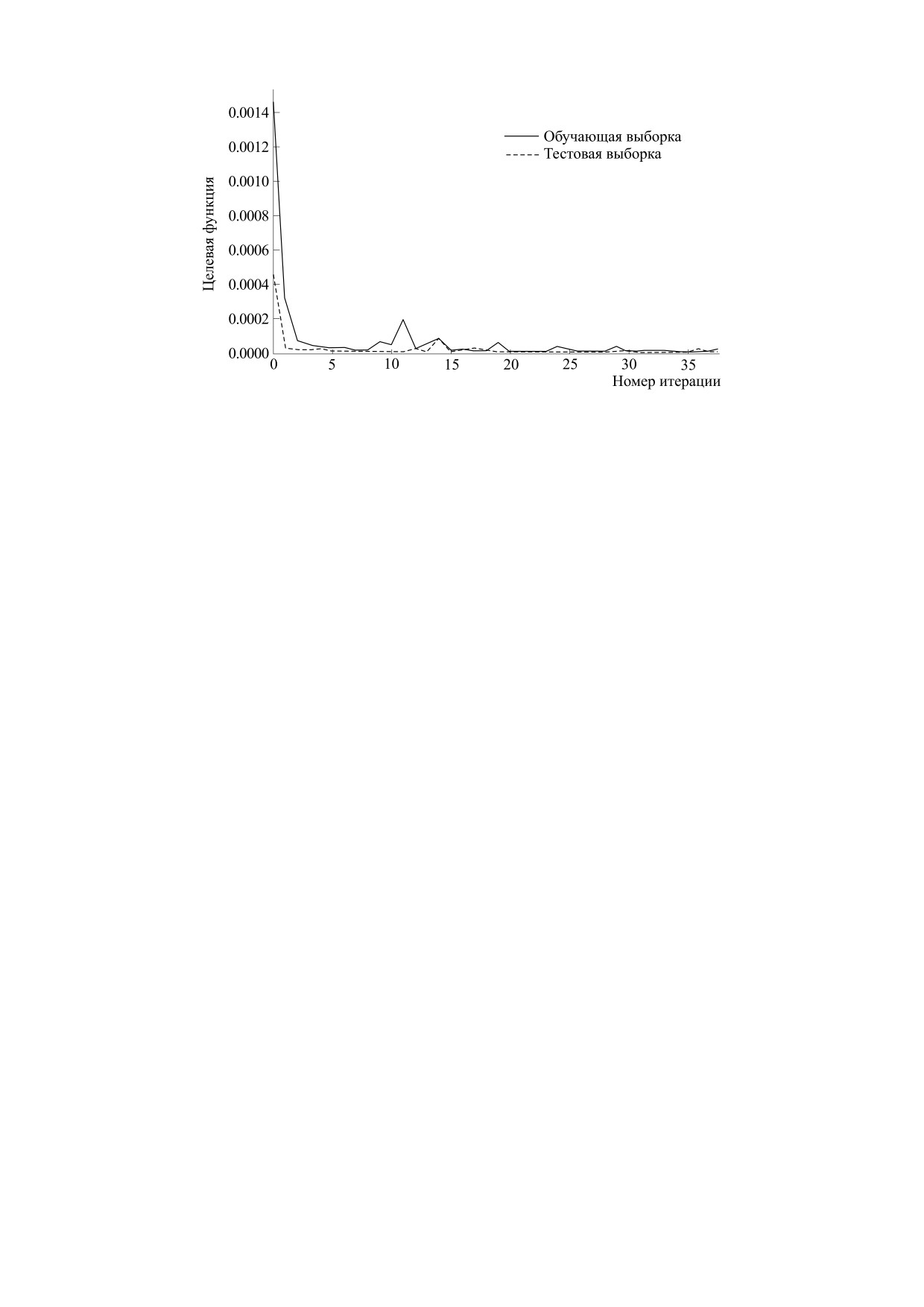
определении 121 параметра. На рис. 3 представ-
2
μ
x
лен график изменения значения функции потерь
при тренировке нейронной сети. Минимальное
∑H
(g
i
−
g
c
)(g
i
−
g
c
)
значение среднеквадратичной ошибки составило
I
3
=
(3)
μ
0,026.
g
В модели [15] для условий, близких к опти-
D
мальным для возделывания сои, был получен оп-
I
4
=
,
(4)
μ
тимальный набор параметров: Tm,c = 5, Tx,c = 30 и
d
= 12. В новых условиях был проведен новый
gc
где H - функция Хевисайда, Tm,c - нижняя гра-
подбор параметров путем перебора с шагом 1 око-
ница роста, Tx,c - верхняя, gc - базовая длина све-
ло предыдущих значений. Полученные для усло-
тового дня, Tm,i, Tx,i, gi — минимальная, макси-
вий Северо-Запада России значения Tm,c = 12,
мальная температура и длина светового дня за
Tx,c = 23 и gc = 13 показывают, что исследованные
день i, а D — номер дня с даты всходов. По срав-
сорта характеризуются более низкой верхней гра-
нению с исходной моделью введены масштабные
ницей температуры роста (23°С вместо 30°С).
константы μm, μx, μg и μd, чтобы не давать заранее
Температурные условия начала вегетации (12°С)
больший вес одному из параметров, но при этом
в регионе определяются началом весенних поле-
не вычислять эти константы для каждого экспе-
вых работ при температурах 10-15°С, связанным
римента и каждого растения.
с избыточной влажностью почвы, что и опреде-
БИОФИЗИКА том 64
№ 3
2019
566
ТАРАТУХИН и др.
Рис. 3. График изменения значения функции потерь.
лило нижнюю границу температурной зоны роста
го перехода температур выше необходимого для
исследованных образцов. Параметр фотоперио-
активного роста сои предела 15°С смещалась на
дической чувствительности gc подтвердил адап-
более ранние сроки на 1,0 сут/год. В 2000-2013 гг.
тированность образцов к более длинному дню
эта дата была 27 мая, к 2030 г. можно прогнозиро-
(13 ч вместо 12 ч).
вать ее наступление в десятых числах мая.
Для масштабных констант были выбраны зна-
С помощью генератора погоды - программы
чения μm = 80, μx = 80, μg = 80 и μd = 36.
МаркСим - были построены прогнозы погоды на
каждый день для Ленинградской области с 2019
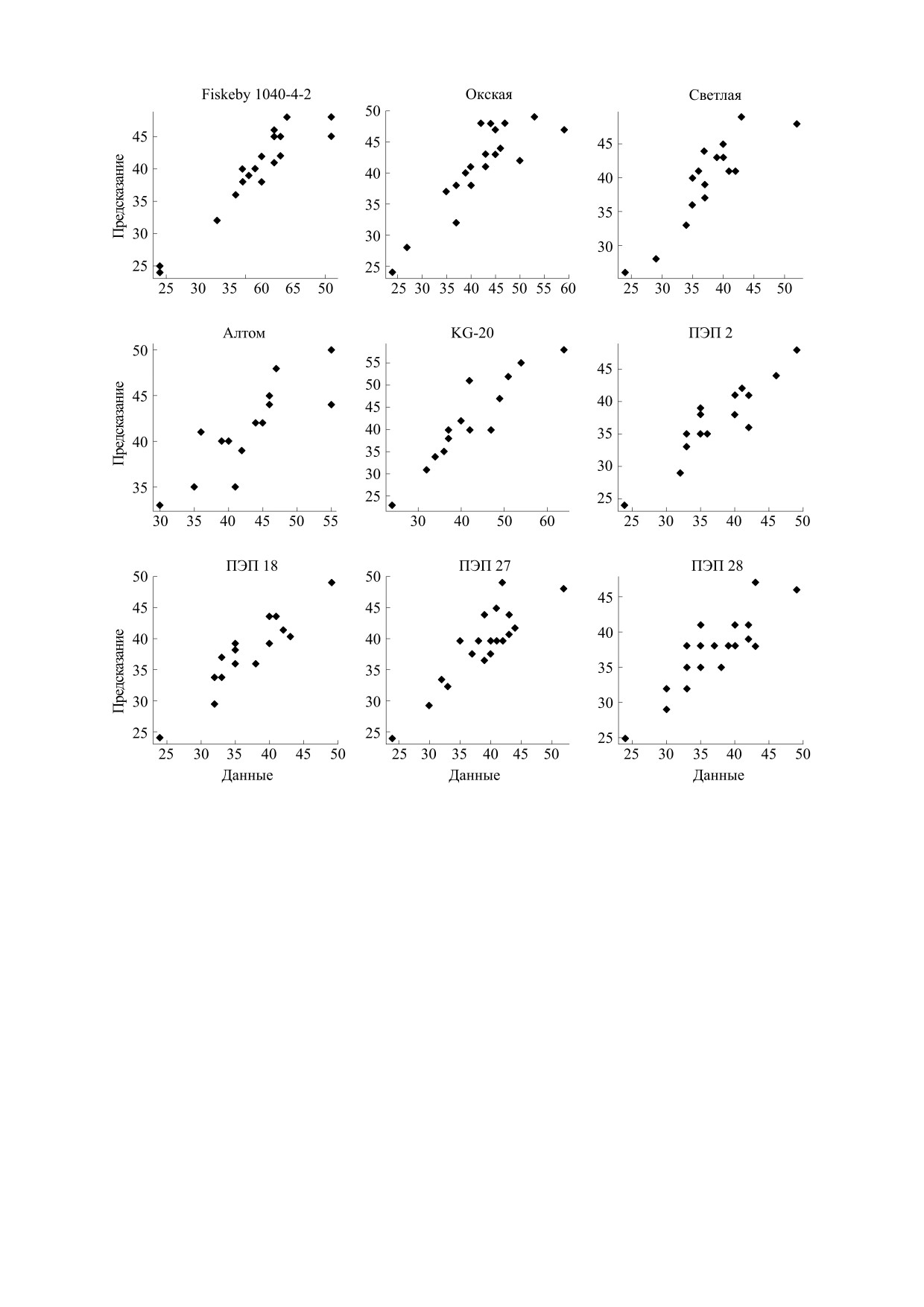
На рис. 4 показано сравнение данных и реше-
до 2030 г. MarkSim был разработан для моделиро-
ния модели. Видно, что модель достаточно точно
вания погоды из известных источников ежеме-
описывает экспериментальные данные.
сячных климатических данных [19-23]. Кроме
В таблице приведено значений сравнение
моделей для прогнозирования климата следует
средней ошибки прогноза, коэффициента корре-
учесть социоэкономические сценарии развития,
ляции и детерминации для новой модели и
которые описываются четырьмя характерными
предыдущей модели температурных минимумов
профилями концентрации углекислого газа
[11] для девяти скороспелых сортов сои. Видно,
(Representative Concentration Pathways, RCPs),
что точность новой модели выше, а средняя
принятые Межправительственной группой экс-
ошибка прогноза улучшилась примерно на двое
пертов по изменению климата (Intergovernmental
суток.
Panel on Climate Change, IPCC) для пятого оце-
ночного отчета (Assessment Report, AR5) в 2014.
Профили соответствуют широкому кругу воз-
РЕЗУЛЬТАТЫ ИСПОЛЬЗОВАНИЯ МОДЕЛИ
можных изменений будущих антропогенных вы-
Актуальной проблемой соеводства являются
бросов парниковых газов и называются rcp26,
ранние и сверхранние посевы [18], позволяющие
rcp45, rcp60 и rcp85 в соответствии с возможными
растениям более эффективно использовать осен-
значениями нарушения радиационного баланса
не-зимние запасы влаги. Проблема ранних посе-
Земли в 2100 г. относительно преиндустриальной
вов сои - замедление развития при более длин-
эры (+2,6, +4,5, +6,0 и +8,5 Вт/м2 соответствен-
ном дне, что сказывается даже на сортах с пони-
но) [24].
женной фотопериодической чувствительностью,
и вероятность заморозков. По модели была ис-
Созданные погодные данные и построенная
следована возможность более раннего посева в
модель были использованы для предсказания
условиях потепления климата. Средняя темпера-
длины периода «всходы-цветение» для девяти
тура посевов в Ленинградской области составила
скороспелых сортов сои в Ленинградской обла-
16,1°С. При потеплении климата дата устойчиво-
сти в 2019-2030 гг.
БИОФИЗИКА том 64
№ 3
2019
МОДЕЛИРОВАНИЕ ФЕНОЛОГИИ СОИ
567
Рис. 4. Сравнение данных и решения модели.
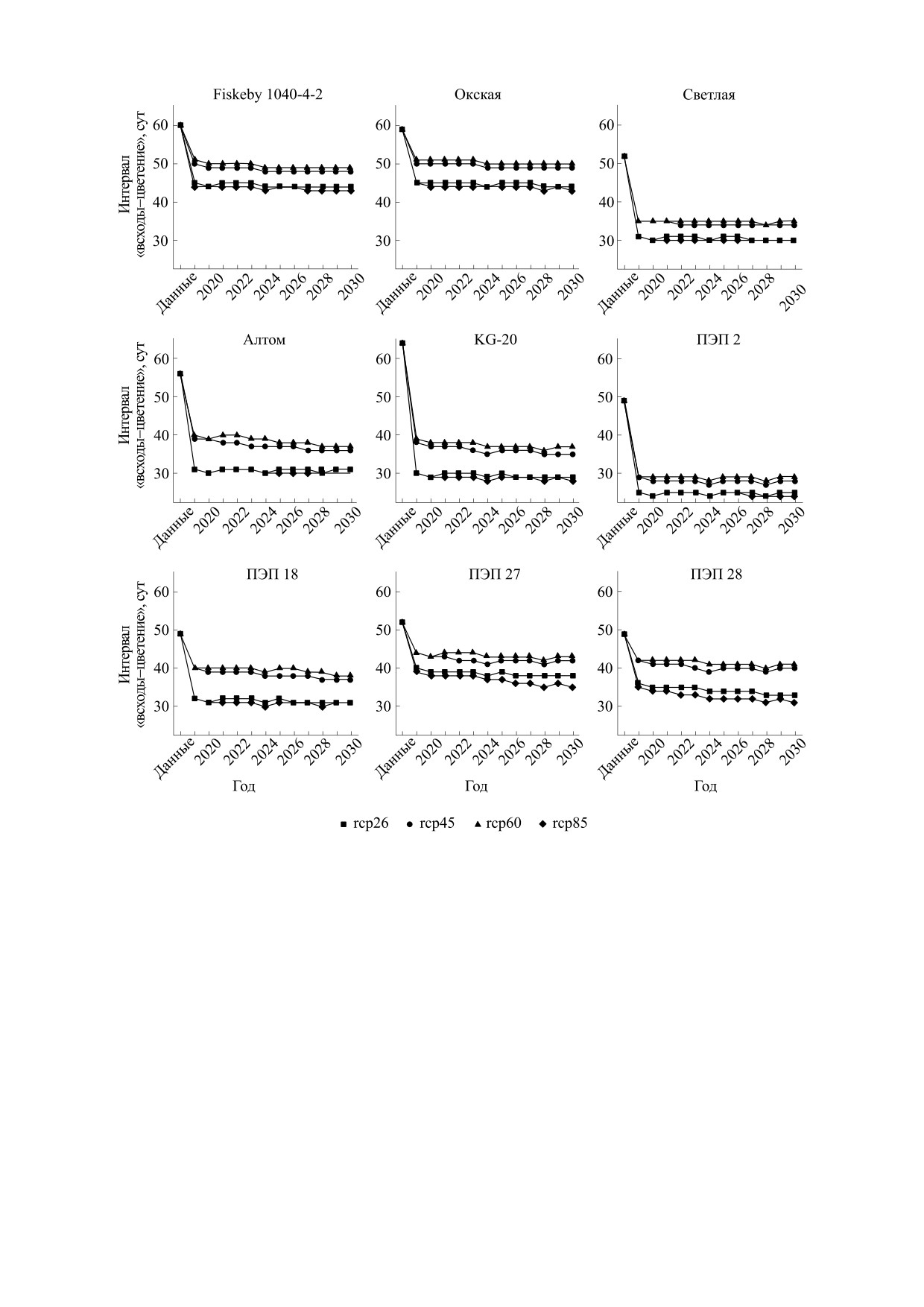
На рис. 5 представлены результаты модельного
риода «всходы-цветение» в контрастных эколо-
исследования посева 1 мая. Результаты модели-
го-географических условиях г. Пушкина и Куба-
рования показывают, что время до цветения
ни, т. е. были наиболее стабильны.
сильно сокращается по сравнению с годами про-
Были модельно исследованы посевы 10 мая в
ведения экспериментов. Для некоторых сортов
условиях потепления (см. рис. 6). Результаты мо-
(Fiskeby
1040-4-2, Окская, Светлая, Алтом,
делирования показывают, что время до цветения
KG-20, ПЭП2) время до цветения будет заметно
сильно сокращается по сравнению с годами про-
сокращаться в период до 2030 г. Для образцов
ведения экспериментов. Для некоторых сортов
ПЭП 27, ПЭП 28, ПЭП 18 возможен вариант, что
(ПЭП 27 и ПЭП 28) время до цветения будет за-
время до цветения существенно не изменится.
метно сокращаться в период до 2030 г. Для
Именно эти образцы (и сорт Светлая) показали
остальных сортов (Fiskeby
1040-4-2, Окская,
наименьшую разницу в продолжительности пе-
Светлая, Алтом, KG-20, ПЭП 2 и ПЭП 18) время
БИОФИЗИКА том 64
№ 3
2019
568
ТАРАТУХИН и др.
Рис. 5. Предсказание интервала «всходы-цветение» для даты посева 1 мая.
до цветения будет оставаться постоянным или не-
пределение модельных результатов представлено на
значительно колебаться.
гистограмме (рис. 7).
Среднее значение длины периода «всходы-цве-
тение» составило для экспериментальных данных
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
39,21 суток, что статистически значимо больше, чем
Математическое моделирование хозяйственно
среднее значение 36,33 суток для результатов моде-
ценных показателей растения, таких как длина
лирования исследуемых образцов в 2030 г. Значение
периода «всходы-цветение» сои, является науч-
одностороннего теста Манна-Уитни-Уилкоксона
ной основой для выведения новых сортов культу-
составило 5423,5, p-значение = 0,009669 < 0,01. Рас-
ры в условиях глобальных изменений климата. В
БИОФИЗИКА том 64
№ 3
2019
МОДЕЛИРОВАНИЕ ФЕНОЛОГИИ СОИ
569
Рис. 6. Предсказание интервала «всходы-цветение» для даты посева 10 мая.
данной работе была построена модель предсказа-
рой модели, так и большим количеством свобод-
ния времени до цветения девяти скороспелых
ных параметров.
сортов сои в условиях Северо-Запада России. Но-
Модельные прогнозы показали перспектив-
вая модель использует специально подобранные
ность сверхранних посевов. Для исследованной
функции погодных данных и нейронную сеть.
группы сортов фактор температур был более важ-
Расчеты по имеющимся экспериментальным
данным показали, что новая модель имеет более
ным, чем фактор длины дня, время цветения сме-
высокую точность по сравнению с моделью тем-
щалось на более ранние сроки, обеспечивая усло-
пературных минимумов. Повышение точности
вия более продолжительного и полного созре-
может объясняться как более сложной структу-
вания.
БИОФИЗИКА том 64
№ 3
2019
570
ТАРАТУХИН и др.
Рис. 7. Гистограмма длин периода «всходы-цветение» в результатах моделирования.
Таким образом, показана пригодность модели
8. T. Hodges and V. French, Agronomy J. 77 (3), 500
фенологии сои, созданной методом искусствен-
(1985).
ных нейронных сетей, для моделирования в нети-
9.
9. P. Pedersen, et al., Agronomy J. 96, 556 (2004).
пичных для культуры условиях.
10. T. D. Setiyono, et al., Field Crops Res. 100 (2-3), 257
Работа выполнена в рамках Федеральной це-
(2007).
левой программы (проект
№14.575.21.0136 от
11. Л. Ю. Новикова, И. В. Сеферова и К. Н. Козлов,
26.09.2017 г., уникальный идентификатор проекта
Биофизика 63 (6), 1182 (2018).
RFMEFI57517X0136). Вычисления были поведе-
12. M. Abdipour, et al., J. Am. Oil Chemists’ Soc. 95 (3),
ны в Суперкомпьютером центре «Политехниче-
283 (2018).
ский» CПбПУ и кластере Университета Южной
13. M. Kaul, R. L. Hill, and C. Walthall, Agricultural Sys-
Калифорнии.
tems 85 (1), 1 (2005).
Исходные данные получены на базе уникаль-
14. A. Bagherzadeh, et al., Modeling Earth Systems and
Environment 2 (2), (2016).
ной научной установки «Коллекция генетиче-
ских ресурсов растений» ВИР и любезно предо-
15. D. A. Elizondo, R. W. McClendon, and G. Hoogen-
ставлены И.В. Сеферовой.
boom, Trans. ASAE 37 (3), 981 (1994).
16. F. Chollet, et al., Keras (GitHub,
2015). URL:
[Электронный ресурс, режим
СПИСОК ЛИТЕРАТУРЫ
доступа: 06.01.2019 г.].
1. А. Г. Папцов и др., Адаптация сельского хозяйства
17. M. Abadi, et al., TensorFlow: A system for large-scale
России к глобальным изменениям климата // Проект
machine learning (2016), p. 21.
независимой
международной
организации
18. С. В. Зеленцов и Е. В. Мошенко, Научный диалог,
№ 7, 40 (2012).
iblock/f96/f9622b41f48549945438f2292f509d14.pdf
[Электронный ресурс, режим доступа: 06.01.2019].
19. M. Srinivasa Rao, et al., PLoS One 10 (2), e0116762
(2015).
2. J. E. Olesen, et al., Eur. J. Agronomy 34 (2), 96 (2011).
20. P. G. Jones and P. K. Thornton, Agricultural and
3. T. Carter and K. Mäkinen, Approaches to climate change
Forest Meteorology 86 (1-2), 127 (1997).
impact, adaptation and vulnerability assessment: towards
a classification framework to serve decision-making: ME-
21. P. G. Jones and P. K. Thornton, Agricultural and
DIATION 2.1 (Finnish Environment Institute (SYKE),
Forest Meteorology 97 (3), 213 (1999).
Helsinki, Finland: 2011).
22. P. G. Jones and P. K. Thornton, Agronomy J. 92, 445
4. A. D. Richardson, et al., Global Change Biology 18
(2000).
(2), 566 (2012).
23. P. G. Jones and A. L. Jones, Centro Internacional de Ag-
5. И. В. Сеферова, Т. В. Мисюрина и М. А. Никиш-
ricultura Tropical. MarkSim: a computer tool that gener-
кина, Сельскохоз. биология, № 5, 42 (2007).
ates simulated weather data for crop modeling and risk as-
6. К. Н. Козлов, Л. Ю. Новикова, И. В. Сеферова и
sessment (CIAT, 2002).
М. Г. Самсонова, Биофизика 63 (1), 175 (2018).
24. D. P. van Vuuren, et al., Climatic Change 109 (1-2), 5
7. D. J. Major, et al., Crop Science 15, 174 (1975).
(2011).
БИОФИЗИКА том 64
№ 3
2019
МОДЕЛИРОВАНИЕ ФЕНОЛОГИИ СОИ
571
Simulation of Soybean Phenology with the Use of Artificial Neural Networks
O.D Taratuhin*, L.Yu. Novikova* **, I.V. Seferova**, and K.N. Kozlov*
*Peter the Great St. Petersburg Polytechnic University, ul. Polytekhnicheskaya 29, St. Petersburg, 195251 Russia
**Federal Research Center “N.I. Vavilov All-Russian Institute of Plant Genetic Resources”,
ul. Bolshaya Morskaya 42-44, St. Petersburg, 190000 Russia
Soybean’s phenology explicitly indicates environmental changes and strongly depends on temperature and
day length. We adapted an artificial neural network model to predict time to flowering in nine early maturing
soybean accessions in the North-West region of Russia. We added scaling constants for network inputs, opti-
mized high and low temperature thresholds and base day length, and implemented a new model written in
Python using Keras and TensorFlow libraries. Experimental data obtained in 1999-2013 in Pushkin (Lenin-
grad region) and Kuban were used for training 121 model parameters and after training the mean-root-square
error became smaller, 0.026. Investigated accessions had the reduced upper temperature threshold as opposed
to literature data (23°С instead of 30°С) and increased low temperature threshold (12°С instead of 5°С). The
extension of day length from 12h to 13h confirmed the adaptation to a longer day. The average prediction error
was improved by approximately 2 days as compared to the previous model of temperature minima. We gen-
erated daily weather for different future greenhouse gas emission scenarios and predicted time to flowering
for nine soybean accessions in changing climate for years 2019-2030 and two planting days - May 1 and May
10. Predicted time to flowering decreases to 2030 for most accessions and scenarios but may stay constant or
f luctuate in several cases. The difference in mean between 39.21 days in experimental data and 36.33 days in
modeling results for 2030 is statistically significant according to Mann-Witney-Wilcoxon criterion (5423.5,
P = 0.0097 < 0.01). Consequently, the results confirmed the predictive power of the developed model.
Keywords: climatic factors, soybeans, artificial neural networks
БИОФИЗИКА том 64
№ 3
2019