БИОФИЗИКА, 2019, том 64, № 3, с. 572-577
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 577.3:612.8
ЭЛЕКТРОХИМИЧЕСКАЯ ОБРАТНАЯ СВЯЗЬ, КАК ОДИН
ИЗ ВОЗМОЖНЫХ МЕХАНИЗМОВ ГЕНЕРАЦИИ НИЗКОЧАСТОТНОЙ
СОСТАВЛЯЮЩЕЙ БИОЭЛЕКТРИЧЕСКОЙ АКТИВНОСТИ МОЗГА
© 2019 г. Е.Л. Машеров
Национальный медицинский исследовательский центр нейрохирургии имени акад. Н.Н. Бурденко,
125047, Москва, ул. 4-я Тверская-Ямская, 16
E-mail: EMasherow@nsi.ru
Поступила в редакцию 29.10.2018 г.
После доработки 24.12.2018 г.
Принята к публикации 24.01.2019 г.
Предложена модель формирования электроэнцефалограммы, учитывающая, помимо суммирова-
ния постсинаптических потенциалов, колебания в процессе регулирования уровня постоянного
потенциала и объясняющая ряд феноменов, наблюдаемых преимущественно в области низкоча-
стотной части спектра электроэнцефалограмм.
Ключевые слова: электроэнцефалограмма, автоматическое регулирование, обратная связь.
DOI: 10.1134/S0006302919030189
личные частоты и формируют разное простран-
Существующая теория, представляющая гене-
ственное распределение электрического потен-
рацию электроэнцефалограммы (ЭЭГ) суммиро-
циала. Такая мысль была высказана ранее в рабо-
ванием постсинаптических потенциалов [1], объ-
те [2], где предлагалось рассматривать диапазоны
ясняя ряд наблюдаемых феноменов, в то же время
1-10 Гц, 10-100 Гц и более 100 Гц как различные
не дает удовлетворительного объяснения ряду
по механизмам формирования (указанная работа
других явлений, обнаруживаемых на ЭЭГ. В их
была сосредоточена на диапазоне 10-100 Гц и ме-
числе сама форма электроэнцефалографического
ханизме порождения ЭЭГ суммированием пост-
сигнала, для ряда ритмов достаточно близкая к
синаптических потенциалов). Диапазон более
синусоидальной; частотный состав с выраженны-
низких частот, однако, представляет большой
ми в его спектре пиками; различия частотного
клинический интерес, включая в себя диагности-
состава ЭЭГ и электрокортикограммы; рост ам-
чески значимые δ-, θ- и α-активности, и если ме-
плитуды ЭЭГ в патологических состояниях, со-
ханизм формирования для них отличен от более
провождающий замедление колебаний биопо-
высокочастотных диапазонов, это позволит раз-
тенциалов; проявление ритмов, обычно трактуе-
рабатывать новые методы анализа электроэнце-
мых, как признак нормального состояния, при
фалограммы или более глубоко понимать суще-
патологических изменениях, например, α-коме и
ствующие. Указанные границы, разумеется, не-
некоторые другие.
возможно рассматривать как точные величины,
Объяснением этому может быть наличие не-
это не более чем указание на порядок значений, и
скольких принципиально различных механизмов
они могут сдвигаться в зависимости от возраста
формирования биоэлектрической активности,
пациента, его состояния, действия препаратов и
действующих одновременно, один из которых, в
т.п. Кроме того, диапазоны эти в значительной
настоящее время рассматривающийся как един-
степени могут пересекаться по частоте.
ственный, связан с суммированием возбуждаю-
щих постсинаптичесих потенциалов (ВПСП) и
тормозящих постсинаптических потенциалов
МАТЕМАТИЧЕСКАЯ МОДЕЛЬ
(ТПСП), другие же могут иметь иную природу.
Предлагаемая попытка объяснения одного из
При этом различные механизмы генерируют раз-
механизмов формирования ЭЭГ опирается на
высказанное в работе [3] утверждение о роли ко-
Сокращения: ЭЭГ - электроэнцефалограмма, ВПСП - воз-
буждающие постсинаптичесие потенциалы, ТПСП - тор-
лебаний уровня постоянного потенциала (пони-
мозящие постсинаптические потенциалы.
мая под этим потенциал покоя, формируемый на
572
ЭЛЕКТРОХИМИЧЕСКАЯ ОБРАТНАЯ СВЯЗЬ
573
мембране нейрона) в биоэлектрической активно-
Таким образом, для реализации подобного ре-
сти. Потенциал этот, формирующийся между
гулирования требуется промежуточное звено
внутренней и внешней сторонами мембраны
управления, роль которого может играть какое-
нейрона, составляет около 70 мВ и обусловлен
либо вещество. Если отклонение регулируемой
разницей концентраций ионов натрия и калия. В
величины приводит к его накоплению, что можно
формировании его принимает участие калий-на-
приблизить интегралом от отклонения, то затем
триевый насос, выводящий из клетки натрий в
концентрация промежуточного вещества влияет
обмен на калий и создающий градиент концен-
на изменение регулируемой величины. В каче-
трации. Затем в ходе формирования потенциала
стве такого промежуточного элемента предлага-
действия (нервного импульса) открываются на-
ется рассмотреть ион Ca2+, предполагая, что по-
триевые каналы и концентрация ионов выравни-
ступление его определяется потенциал-зависи-
вается [4]. Оксюморон, порожденный сочетани-
мым кальциевым насосом, а калий-натриевый
ем «колебаний» и «постоянного» снимается, если
насос регулируется концентрацией иона кальция
вспомнить, что постоянство в условиях измене-
[6,7]. Концентрация кальция вне клетки состав-
ния условий функционирования организма тре-
ляет, согласно работе [7], 2,28 ммоль/литр (для
бует наличия регулятора, но при этом «нет регу-
сравнения: для калия - 2,86 ммоль/литр), его
лятора без колебания» [5]. Последний тезис сле-
внутриклеточная концентрация в состоянии по-
дует понимать так, что регулятор, способный
коя мала, однако после генерации потенциала
выдерживать требуемый уровень при различных
действия существенно возрастает, а затем каль-
значениях нагрузки (астатический), должен учи-
ций выводится из клетки. Функции, описываю-
тывать значения отклонения не только в текущий
момент, но и в прошлые моменты времени, и не-
щие влияние уровня потенциала на изменение
избежно будет порождать колебания. Простей-
концентрации кальция и влияние концентрации
ший тип такого регулятора - интегральный, в ко-
кальция на работу калий-натриевого насоса, во-
тором величина отклонения интегрируется по
обще говоря, нелинейны. Однако, ограничиваясь
времени, а изменение регулируемой величины
в качестве первого приближения линейной зави-
является функцией от этого интеграла.
симостью, получим:
dK
2+
Ca
dP
=
f P)
≈-a(P
−
π)
+
b(K
2+
−κ);
=
g
(
K
2+
)
≈
c(K
2+
−κ)
Ca
Ca
Ca
dt
dt
где K - концентрация ионов кальция, P - внекле-
где ω есть мнимая часть, а - δ - действительная
точный потенциал, κ, π, g, h - параметры линеа-
часть корней вспомогательного уравнения
ризованной модели (κ и π - значения концентра-
z2 = bz - w2, коэффициенты же A и φ задаются на-
ции кальция и потенциала при нормальных усло-
чальными условиями (формально решением
виях, a и c - воздействие отклонений значений
может быть и незатухающая, и даже имеющая
потенциала и концентрации кальция на скорость
экспоненциально возрастающую амплитуду си-
изменения концентрации кальция и потенциала
нусоида, но такие решения не выглядят осмыс-
соответственно, b - диффузия кальция в сосед-
ленными физиологически).
ние структуры мозга в ходе его выведения). Эта
Возникающие при этом колебания предпола-
система дифференциальных уравнений диффе-
гаются источником низкочастотной (предполо-
ренцированием второго и подстановкой в него
жительно до
10-15 Гц) части спектра ЭЭГ.
первого уравнения сводится к линейному диффе-
Спектр их имеет форму достаточно острого пика
ренциальному уравнению второго порядка, где
(в зависимости от декремента затухания δ - чем
p = P - π:
ниже затухание, тем острее пик спектра). Частота
же зависит от коэффициентов модели, а они
2
d
p
2
dp
определяются, с одной стороны, наличием ис-
=
aс(P
−π)
+
bc(K
2+
−
κ)
=-w
p+ b
,
2
Ca
dt
dt
пользуемой калий-натриевым насосом АТФ,
на которое, в свою очередь, влияет оксигенация
решением которого является экспонента с ком-
соответствующих структур мозга, с другой сторо-
плексным показателем, в действительной обла-
ны - от концентрации ионов кальция. Как пока-
сти в силу тождества Эйлера eix = cosx + isinx
зывают клинические наблюдения [1], как недо-
представляемая как затухающая синусоида:
статок кислорода, так и гипокальциемия могут
проявляться медленноволновой активностью
p(t) = Re(Aeiωt-δ) = Acos(ωt + φ)e-δt,
или комплексами «пик-медленная волна». На-
БИОФИЗИКА том 64
№ 3
2019
574
МАШЕРОВ
личие указанной зависимости может позволить
Описание картины электрического поля, фор-
оценивать по ЭЭГ эти существенные характери-
мирующегося в процессе регулирования посто-
стики ткани мозга, однако математический аппа-
янного потенциала, включает, помимо диполь-
рат для такого оценивания в значительной степе-
ных компонент, также и монопольные компо-
ни нужно создавать заново.
ненты, так что убывание потенциала в
зависимости от расстояния до источника не столь
Чтобы колебания, имеющие затухающий ха-
быстрое, как в случае чисто дипольного источни-
рактер, наблюдались достаточно длительное вре-
ка (обратно к расстоянию в отличие от обратно-
мя, требуется наличие возбуждающего сигнала.
квадратичной зависимости для диполей), и не за-
Если при этом поток возбуждающих сигналов яв-
висит от ориентации диполя по отношению к
ляется пуассоновским, то есть появление очеред-
приемнику, что позволяет объяснить успешность
ного возбуждающего импульса не зависит от по-
регистрации сигнала от глубинных источников.
явления прочих, частота полученного сигнала
Для дипольных источников приходится, чтобы
будет соответствовать частоте отдельной затухаю-
объяснить наблюдаемые на скальпе значения ам-
щей синусоиды. Для непуассоновского потока
плитуды, либо отказываться от доказавшей свою
такого прямого соответствия не будет, частотная
практическую полезность модели объемного
характеристика полученного сигнала будет опре-
проведения [9], либо предполагать чрезвычайно
деляться частотным составом исходного возбуж-
высокие, не подтверждающиеся внутримозговы-
дающего сигнала, пропущенного через фильтр с
ми измерениями, значения потенциала вблизи
амплитудно-частотной характеристикой, соот-
источников. В литературе отмечалось, что вкла-
ветствующей амплитудно-частотной характери-
дом монопольных источников нельзя пренебре-
стике данного регулирующего механизма. По-
гать [10].
скольку поток сенсорной информации заведомо
будет непуассоновским, а в отсутствие такого по-
АНАЛИЗ ДАННЫХ И ЧИСЛЕННОЕ
тока отдельные спонтанно возникающие импуль-
МОДЕЛИРОВАНИЕ
сы будут независимы и могут быть приближены
пуассоновским потоком, так может объясняться
Утверждения, что представленная модель спо-
появление α-ритма при закрывании глаз и его
собна описывать некоторые ритмы ЭЭГ, как и то,
смена высокочастотной активностью при откры-
что существуют проявления биоэлектрической
вании, появление роландического (μ-) ритма при
активности, не вполне укладывающиеся в объяс-
расслаблении контралатеральной руки и его ис-
нение суммированием ВПСП и ТПСП, требуют
чезновение при сжимании, а также явление α-ко-
экспериментального подтверждения. Поскольку
мы при тяжелых стволовых поражениях: этот
данные, записываемые электроэнцефалографом,
крайне неблагоприятный прогностически при-
дискретизируются по времени, от приведенного
знак тогда может трактоваться как прекращение,
выше дифференциального уравнения необходи-
в силу поражения стволовых структур, сенсорно-
мо перейти к уравнению в конечных разностях:
го притока в относительно сохранную кору.
yt = a1yt-1 + a2yt-2 + ε,
Замедление ритмики с одновременным ро-
где yt - наблюдаемый биопотенциал, a1 и a 2 - ко-
стом амплитуды колебаний в патологических со-
эффициенты модели, ε - возбуждающий сигнал.
стояниях (замедление частоты α-ритма вплоть до
Для записанного в отведении O2 α-ритма была
перехода в область частот θ-ритма, появление δ-
получена следующая оценка:
активности в области травмы, перифокальной зо-
yt = 1,629469yt-1 - 0,928725yt-2 + ε,
не опухоли, при нарушениях мозгового кровооб-
ращения), а также при некоторых функциональ-
по которой были вычислены частота ω = 8,99 Гц и
ных пробах (тест Матаса с пережатием сонной ар-
декремент затухания δ = 0,0369.
терии, где появление медленноволновой
С использованием в качестве возбуждающего
активности свидетельствует о недостаточности
сигнала последовательности нормально распре-
кровоснабжения полушария через соединитель-
деленных случайных чисел была смоделирована
ные артерии) тогда означает, что регулятор не
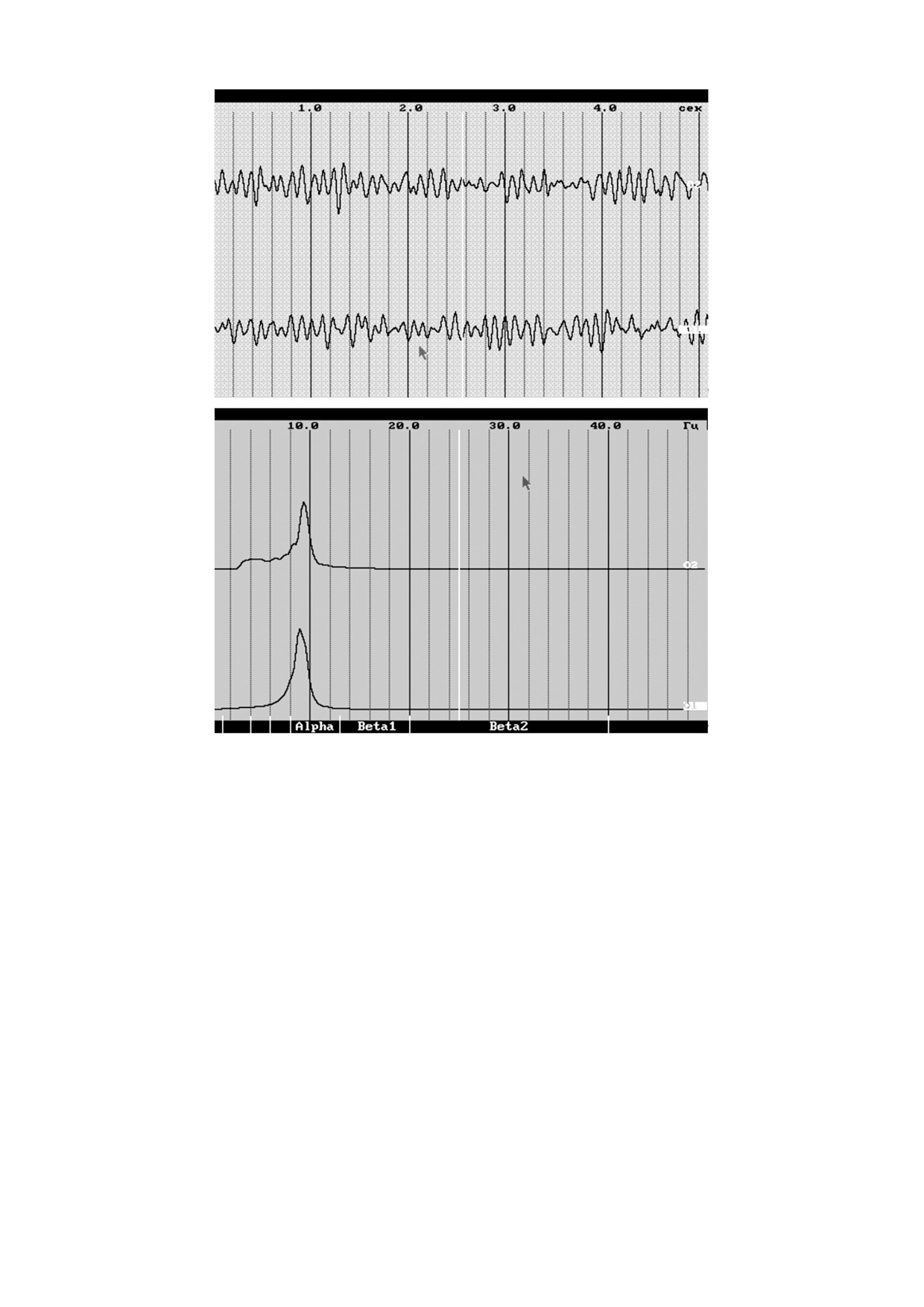
ЭЭГ, представленная на рис. 1а (вверху - натив-
вполне справляется с регулированием, так что от-
ная ЭЭГ, внизу - смоделированная). Были рас-
клонения устраняются медленнее, а их величина
считаны спектры мощности реальной и смодели-
(амплитуда патологической активности) возрас-
рованной кривых (рис. 1б). На обоих четко выде-
тает. С другой стороны, увеличение поступления
ляется пик в полосе α-ритма. В то же время
кислорода к тканям мозга, например, в ходе ги-
видно, что как нативная кривая, так и в особен-
пербарической оксигенации, как отмечалось в
ности ее спектр существенно богаче смоделиро-
работе [8], приводит к увеличению частоты α-
ванной. Это прежде всего может быть обусловле-
ритма.
но тем, что в модели предполагается единствен-
БИОФИЗИКА том 64
№ 3
2019
ЭЛЕКТРОХИМИЧЕСКАЯ ОБРАТНАЯ СВЯЗЬ
575
Рис. 1. Нативная (верхняя кривая) и смоделированная (нижняя кривая) ЭЭГ (а) и их спектры мощности (б).
ный источник, тогда как запись реальной ЭЭГ
баний указанных ритмов. Для проверки этого бы-
отражает суммарную активность различных
ла записана активность в состоянии покоя с за-
структур мозга, включая более отдаленные от ме-
крытыми глазами в отведении O1, разделенная
ста записи.
цифровой фильтрацией на диапазоны α (8-13 Гц)
Модель, в которой суммировались возбуждае-
и β (15-45 Гц). Колебания, соответствующие по
мые в случайные моменты постсинаптические
длительности и форме ВПСП и ТПСП, должны
потенциалы описанной в литературе формы (вос-
проявиться в частотной полосе β-ритма. Была
ходящая фаза 2-3 мс, нисходящая фаза 10-12
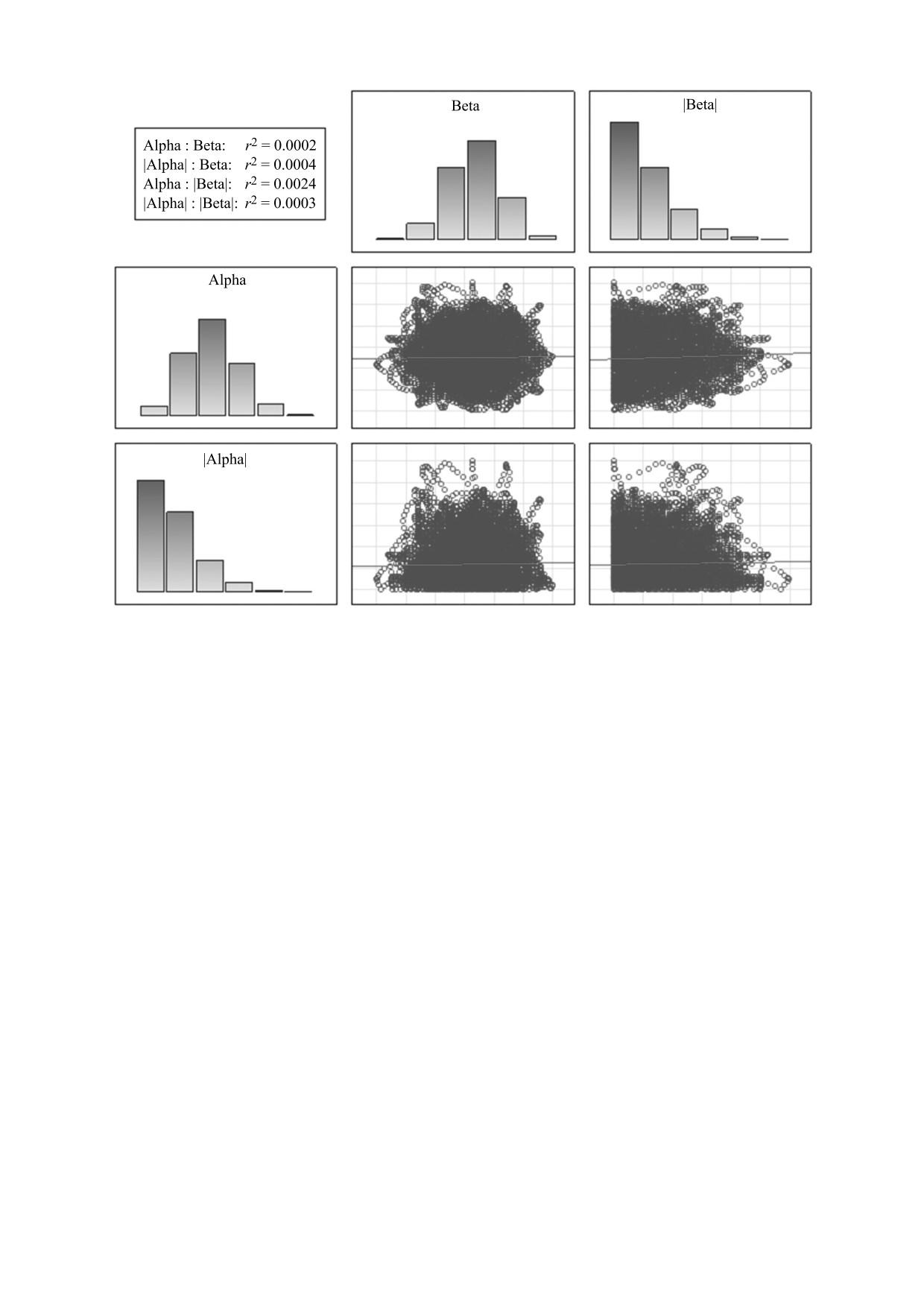
вычислена огибающая β-активности, и затем рас-
мс), дала картину, совершенно не схожую с пери-
смотрена корреляция, с одной стороны, сигнала в
одической активностью в составе ЭЭГ, однако
имеющую визуальное сходство с β-активностью и
полосе α и его абсолютной величины, а с другой
близкий к ней спектральный состав.
стороны, огибающей β-ритма и ее абсолютной
величины. Как представлено на рис. 2, значимой
Однако остается возможность того, что коле-
связи между этими величинами выявлено не бы-
бания в диапазоне δ-, θ- и α-ритмов формируют-
ло, что, разумеется, не доказывает отсутствия
ся не случайно и независимо возникающими
ВПСП и ТПСП, а пачками импульсов, и частота
связи между различными ритмами ЭЭГ, но явля-
поступления этих пачек определяет частоту коле-
ется аргументом против утверждения о формиро-
БИОФИЗИКА том 64
№ 3
2019
576
МАШЕРОВ
Рис. 2. Взаимосвязь сигнала в полосе α-ритма (Alpha) и огибающей β-ритма (Beta) и абсолютных величин этих
сигналов (|Alpha| и |Beta| соответственно).
вании низкочастотных ритмов суммированием
более высоких частот (β- и γ-ритмов и «острых
постсинаптических потенциалов.
волн») существующая модель ЭЭГ, объясняемая
суммированием ВПСП и ТПСП, представляется
вполне адекватной.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Практическая ценность такой модели форми-
Данная модель предлагается отнюдь не в каче-
рования низкочастотной части ЭЭГ обусловлена,
стве альтернативы существующей модели, объяс-
в частности, тем, что частотный состав ЭЭГ с са-
няющей формирование потенциала ЭЭГ сумми-
мого начала клинической электроэнцефалогра-
рованием дипольных потенциалов, связанных с
фии рассматривался как характеристика состоя-
ВПСП и ТПСП, а как ее дополнение, позволяю-
щее объяснить прежде всего особенности частот-
ния пациента. Появление медленных ритмов или
замедление существующих может свидетельство-
ного состава ЭЭГ. Даваемое принятой в настоя-
щее время моделью объяснение появления низ-
вать о патологических изменениях или же о про-
ких частот вследствие того, что суммируется
явлениях изменения физиологического состоя-
большое число отдельных источников импульсов
ния (сон, анестезия). В связи с этим нашли при-
малой продолжительности, противоречит линей-
менение такие показателя, как средняя или
ности преобразования Фурье, исходя из которой
медианная частота, частоты спектрального среза
комплексный спектр суммы источников должен
(на 90% или 95% мощности спектра), а также эф-
быть равен сумме спектров отдельных источни-
фективная частотная полоса [11-15]. Однако они
ков. Однако длительность постсинаптических
были введены как эмпирические характеристики
потенциалов составляет порядка 10-20 мс, что
биоэлектрической активности мозга, и модель,
соответствует колебаниям с частотой порядка де-
объясняющая связь физиологического состояния
сятков герц, и частота суммарного сигнала будет
тканей мозга и показателей спектра ЭЭГ, может
близка к средней частоте этих источников. Для
представить практический интерес, позволяя
БИОФИЗИКА том 64
№ 3
2019
ЭЛЕКТРОХИМИЧЕСКАЯ ОБРАТНАЯ СВЯЗЬ
577
усовершенствовать методику получения таких
3. В. С. Русинов, Доминанта (Медицина, М., 1969).
показателей.
4. Й. Дудель, Й. Рюэгг, Р. Шмидт и др., Физиология
человека (Мир, М., 2007).
Несмотря на крайне упрощающий характер
предположений, на которых основывается дан-
5. W. F. Boron and M. D. Boulpaep, Medical Physiology:
A Cellular and Molecular Approach (Elsevier, Boston,
ная модель, она способна объяснить ряд наблю-
2017).
даемых феноменов электроэнцефалограммы. В
6. Cerebrospinal fluid in clinical practice, Ed. by
то же время появление сигналов с выраженной
D. N. Irani (Elsevier, Boston, 2009)
формой, такой как аркообразная для μ-ритма или
7. Дж. К. Максвелл, И. А. Вышнеградский, А. Стодо-
пик-волновые комплексы, наблюдаемые при
ла, Теория автоматического регулирования (Изд-во
эпилепсии, она объяснить не может, однако это
АН СССР, М., 1949).
представляется возможным при введении в мо-
8. Руководство по гипербарической оксигенации, под
дель нелинейностей. Остается открытым и во-
ред. С. Н. Ефуни (Медицина, М., 1986).
прос взаимодействия между собой различных ис-
9. В. В. Гнездицкий, Обратная задача ЭЭГ и клиниче-
точников биоэлектрической активности, в том
ская электроэнцефалография (картирование и лока-
числе и их самосинхронизация. Предложенная
лизация источников электрической активности
модель не может рассматриваться иначе как пер-
мозга) (Медпрессинформ, М., 2004.
вое приближение, однако требуемое для повыше-
10. A. Varsavsky, I. Mareels, and M. Cook, Epileptic sei-
ния реалистичности ее усложнение может быть
zures and the EEG: measurement, models, detection and
сделано лишь после проведения дополнительных
prediction (Taylor & Francis, Boca Raton, 2011).
исследований, включающих одновременную ре-
11. T. W. Boonstra, S. Nikolin, A.-C. Meisener, et al.,
гистрацию разнородных параметров (ЭЭГ, транс-
Frontiers Human Neurosci. 10, 270 (2016).
краниальной оксигенации, скорости кровотока)
12. N. Accornero, M. Capozza, L. Pieroni, et al., J. Neu-
и сопоставления с клиническими данными.
rophysiol. 112 (6), 1367 (2014).
Работа выполнена при финансовой поддержке
13. A. Moungou, J.-L. Thonnard, and A. Mouraux, Sci.
Российского фонда фундаментальных исследова-
Reports 6, 20738 (2016). DOI: 10.1038/srep20738.
ний (грант 18-013-00967а).
14. О. Б. Сазонова, Е. М. Трошина, Е. Л. Машеров и
А. Ю. Беляев, в сб. Материалы Международной
конференции «Новые информационные технологии в
СПИСОК ЛИТЕРАТУРЫ
медицине, биологии, фармакологии и экологии» (Ин-
1. J. S. Ebersole, A. M. Husain, and D. R. Nordli Jr., Cur-
ститут новых информационных технологий, М.,
rent Practice of Clinical Electroencephalography (Wolters
2016).
Kluwer, Philadelphia, 2014)
15. А. В. Рылова, О. Б. Сазонова, А. Ю. Лубнин и
2. А. М. Гутман, Биофизика внеклеточных полей мозга
Е. Л. Машеров, Анестезиология и реаниматоло-
(Наука, М., 1980).
гия, № 2, 31 (2010).
Electrochemical Feedback as a Possible Mechanism for Generation of Low-Frequency
Component of Brain Bioelectric Activity
E.L. Masherov
Burdenko National Scientific and Practical Center for Neurosurgery,
4-ya Tverskaya-Yamskaya ul. 16, Moscow, 125047 Russia
We propose a model of electroencephalogram recording that includes, in addition to a summation of post-
synaptic potentials, the fluctuations while controlling the constant potential level and explains a number of
phenomena observed predominantly in the low-frequency spectrum of electroencephalogram.
Keywords: electroencephalogram, automatic control, feedback
БИОФИЗИКА том 64
№ 3
2019