БИОФИЗИКА, 2019, том 64, № 3, с. 578-586
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 577.3
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ НЕОДНОРОДНОГО
ИНТАКТНОГО СИНОАТРИАЛЬНОГО УЗЛА СЕРДЦА КРОЛИКА
С ИСПОЛЬЗОВАНИЕМ МЕТОДОВ ТЕОРИИ ПОДОБИЯ
© 2019 г. В.В. Галанин
Медицинский университет «Реавиз», 443001, Самара, ул. Чапаевская, 227
E-mail: galanin_v@hotmail.com
Поступила в редакцию 10.12.2018 г.
После доработки 10.12.2018 г.
Принята к публикации 18.03.2019 г.
В работе на основе детального подхода моделируется электрическая активность неоднородного ин-
тактного синоатриального узла при его взаимодействии с сократительным миокардом предсердия.
Осуществлена редукция данной модели с использованием методов теории подобия. Проведено
сравнение результатов расчета редуцированной модели с решениями, полученными в исходной мо-
дели. Также в модели установлено, что увеличение числа невозбудимых участков в миокарде пред-
сердия, пограничном пейсмекеру, вызывает замедление общего ритма электрической активности.
Ключевые слова: синоатриальный узел, предсердие, кардиомиоцит, математическое моделирование,
теория подобия, сложные системы.
DOI: 10.1134/S0006302919030190
математически моделируется синхронизация и
Синоатриальный узел (САУ) сердца кролика
распространение возбуждения в САУ в норме и
содержит порядка полумиллиона кардиомиоци-
под действием ацетилхолина. В работе [6] рас-
тов [1], обладающих пейсмекерной активностью
сматривается 1D-модель САУ с учетом неодно-
и связанных друг с другом электротонически с
родности его структуры. Данная модель содержит
помощью щелевых контактов. Пейсмекерная
200 пейсмекерных и типичных предсердных эле-
ткань узла по своим морфологическим и электро-
ментов. 2D-модель в исследовании [7] включает
физиологическим свойствам является неодно-
уже 91 × 28 элементов. Однако работа с таким
родной [1-3]: центральные клетки отличаются от
множеством элементов требует значительных вы-
периферийных малым размером, слабо развитым
числительных ресурсов.
сократительным аппаратом, отсутствием быстро-
го деполяризующего натриевого тока и более
В предыдущей нашей статье [8] была представ-
продолжительной фазой медленной диастоличе-
лена модель функционирования группы клеток
ской деполяризации. Также необходимо учиты-
центральной части САУ с учетом физического
вать, что узловая ткань взаимодействует с много-
подобия одно- и многопейсмекерного клеточно-
кратно превосходящим ее объемом сократитель-
го образования. Предметом настоящего исследо-
ных клеток миокарда предсердия, который
вания является возможность применения теории
оказывает замедляющее влияние на ритм пейсме-
подобия для моделирования электрической ак-
кера САУ.
тивности разнородного по своей структуре ин-
Столь неоднородная тканевая структура, со-
тактного САУ при его взаимодействии с типич-
стоящая из множества взаимодействующих меж-
ным миокардом предсердия. Данный подход поз-
ду собой кардиомиоцитов, может быть описана в
воляет существенно упростить описание
рамках классического подхода Ходжкина-Хакс-
процессов в САУ, уменьшив количество рассмат-
ли [4], в котором каждая клетка моделируется
риваемых элементов, и, в то же время, сохранить
системой нелинейных дифференциальных урав-
наиболее важные особенности моделей, у кото-
нений. К настоящему времени разработано мно-
рых число используемых уравнений гораздо
жество детальных моделей электрической актив-
больше. Также для сокращения числа дифферен-
ности САУ на основе данного подхода. Напри-
циальных уравнений мембранного потенциала
мер, представляет интерес работа [5], в которой
электровозбудимых клеток в данной работе при-
менен метод разделения кинетических перемен-
Сокращение:САУ - синоатриальный узел.
ных ионных токов на быстрые и медленные.
578
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ
579
МОДЕЛЬ НЕОДНОРОДНОГО ИНТАКТНОГО
СИНОАТРИАЛЬНОГО УЗЛА
Представим интактный САУ и окружающий
его рабочий миокард предсердия в виде плоского
диска, состоящего из M коаксиальных цилиндри-
ческих слоев пейсмекерных клеток и N слоев ти-
пичных клеток предсердия (рис. 1). Электрофи-
зиологические параметры мембраны пейсмекер-
ных клеток внутри каждого слоя постоянные, но
отличаются у соседних слоев в соответствии с вы-
ражениями, предложенными в работе [6]:
Рис. 1. Схематическое изображение неоднородного
0,04s
−1,0
интактного САУ и окружающего его сократительно-
⎡
⎛
⎞⎤
C
=
C
+
C
−C
1+
exp
−
,
го миокарда предсердия в виде плоского диска (s -
s
c
(
p
c
)
⎢
⎜
⎟⎥
⎣
⎝
0,2
⎠⎦
пейсмекерные слои САУ, a - предсердные слои со-
кратительного миокарда; c, p - центральный и пери-
(
C
p
−C
s
)
ферийный слои САУ соответственно).
g
=
g
+
(
g
−
g
)
,
(1)
is
ic
ip
ic
(
C
p
−C
c
)
c
c
c
⎡
⎛
0,04s
−
1,0⎞⎤
смекерных клеток САУ, представлены в
g
=
g
+
g
1
+
exp
−
,
s
c
p
⎢
⎜
⎟⎥
⎣
⎝
0,2
⎠⎦
Приложении 1.
где s - номер пейсмекерного слоя (s = s1, ..., sM);
Электроемкость Ca, электропроводимость
ионных каналов gia мембраны типичных клеток
Cs, gis, gcs - электроемкость, ионная электропро-
водимость через каналы мембраны кардиомио-
предсердия, а также электропроводимость меж-
цита и электропроводимость межклеточных кон-
клеточных контактов gca от слоя к слою не меня-
тактов для s-го слоя; Cc,p, gic,p, gcc,p - электроем-
ются (a - номер предсердного слоя, a = a1, ..., aN).
кость, ионная электропроводимость мембраны и
Описание типичных клеток предсердия следует
электропроводимость межклеточных контактов в
модели, развитой в работе [11].
центре (c) и на периферии (p) САУ соответ-
Кроме того, поскольку миокард предсердия
ственно.
содержит невозбудимые участки соединительной
Для описания колебаний мембранного потен-
ткани [2,3], блокирующей распространение воз-
циала центральных и периферийных пейсмекер-
буждения в тангенциальном направлении, разде-
ных клеток САУ упростим математическую мо-
лим околопейсмекерную область типичного мио-
дель, представленную в работах [9,10]. Для этого
карда на несколько электрически несвязанных
наиболее быстрые кинетические переменные
секторов, например на три (рис. 1).
ионных токов заменим на их квазистационарные
Выбранная в модели радиальная структура
значения аналогично тому, как это было сделано
предполагает эквипотенциальность каждого от-
в работе [8]. Натриевый ток пейсмекеров, грани-
дельно взятого клеточного слоя. Вследствие этого
чащих с предсердными слоями, задается выраже-
система дифференциальных уравнений для по-
ниями модели в работе [11]. Подробные уравне-
тенциала мембраны цилиндрического слоя кар-
ния, определяющие модель изолированных пей-
диомиоцитов имеет следующий вид:
dV
s
⎡
c
c
⎤
C
=-
V
−
E
)
−
g
(V
−V
)
−
g
(V
−V
)
,
s
⎢
∑gis(
s
i
s−1
s−1
s
s
s+1
s
⎥
dt
⎣
i
⎦
(2)
dV
a
⎡
c
c
⎤
C
a
=-
g
a a
V -E
i
)
−
g
a−1
(V
a−1
−V
a
)
−
g
(
a a
V
+1
−V
a
)
,
⎢
⎥
dt
⎣
i
⎦
где Vs и Va - мембранный потенциал s-го пейсме-
ных уравнений модели можно найти в Приложе-
нии 2.
керного и a-го предсердного клеточных слоев со-
ответственно; Ei - равновесный потенциал i-го
В результате модель интактного САУ и связан-
ионного тока. Полную систему дифференциаль- ного с ней миокарда предсердия в виде плоского
БИОФИЗИКА том 64
№ 3
2019
580
ГАЛАНИН
Рис. 2. Схема разветвленной цепочки пейсмекерно-предсердных элементов.
диска эквивалентна цепочке из sM пейсмекерных
поскольку он характеризуется достаточно устой-
элементов, соединенной с тремя цепочками, в
чивым регулярным ритмом электрической актив-
каждой из которых содержится aN предсердных
ности - качеством, неприсущим отдельным пей-
элементов (рис. 2).
смекерным клеткам. Следовательно, если мы ме-
няем способ соединения элементов или число
На рис. 3 и 4 представлены профили потенци-
межэлементных связей в модели узла, то систему
алов действия, рассчитанные в соответствии с
дифференциальных уравнений необходимо пре-
данной моделью (M = 55, 3N = 30). Период следо-
вания спонтанных потенциалов действия в дан-
образовать так, чтобы сохранить основные харак-
ной модели равен 324 мс. Рис. 3 и 4 иллюстрируют
теристики ритма САУ (период, фазу, профили
прогрессивное изменение формы потенциала
потенциалов действия). Далее будет продемон-
действия от центра САУ к предсердию.
стрирован способ, в соответствии с которым при-
Компьютерное моделирование показывает,
менение методов теории подобия позволяет до-
что увеличение числа ветвей предсердных эле-
статочно эффективно выполнить редукцию ис-
ментов, взаимодействующих с периферийным
ходной математической модели САУ.
пейсмекером, сопровождается значительным
Итак, упростим исходную модель «САУ-пред-
удлинением периода следования электрических
импульсов (таблица, рис. 5). При достаточно
сердие», т.е. уменьшим число дифференциаль-
большом количестве таких разветвлений (в рас-
ных уравнений в системе (2), используя теорию
сматриваемой модели - больше пяти, рис. 5б) пе-
подобия. Сначала оставим в цепочке пейсмекер-
редача возбуждения на миокард предсердия не
ных и предсердных элементов только каждый пя-
происходит.
тый элемент: s′ = 1, ..., 11; a′ = 1, 2. В этом случае
электроемкость и электропроводимость межкле-
точных контактов будут определяться следующи-
РЕДУКЦИЯ МОДЕЛИ ИНТАКТНОГО
ми соотношениями:
СИНОАТРИАЛЬНОГО УЗЛА
С ИСПОЛЬЗОВАНИЕМ
C
=
C
+
(
C
−C
) [
1+
exp
(
−s′
+
5,8
)]
,
МЕТОДОВ ТЕОРИИ ПОДОБИЯ
s′
c
p
c
(3)
c
c
c
САУ - сложная тканевая система, основной
g
=
g
+
g
[
1+
exp
(
−s
′
+
5,8
)]
s′
c
p
структурной единицей которой является пейсме-
керная клетка. Благодаря взаимной электротони-
В результате получается укороченная цепочка
ческой синхронизации пейсмекерных клеток
клеточных элементов, описываемая следующей
САУ как целое представляет собой нечто большее,
системой дифференциальных уравнений (штри-
чем просто совокупность клеточных элементов,
хи над индексами s и a опущены):
dV
s
′
⎡
c
c
⎤
C
=-
V′-
E
)
−
g′
(V′
−V′)
−
g′
(
V′
−V′)
,
s
⎢
∑gis(
s
i
s−1
s−1
s
s
s+1
s
⎥
dt
⎣
i
⎦
(4)
dV
a
′
⎡
c
c
⎤
C
=-
g
V′−E
)
−
g′
(V′
−V′)
−
g′
(V
′
−V
′)
a
⎢
a a
i
a−1
a−1
a
a
a+1
a
⎥
dt
⎣
i
⎦
Поскольку модель САУ представляет собой
ношений пропорциональности между их сход-
систему нелинейных дифференциальных уравне-
ственными параметрами (электроемкостями,
ний, условием подобия исходной (2) и редуциро-
электропроводимостями, мембранными потен-
ванной (4) моделей является существование соот-
циалами, ионными токами и т.д.):
БИОФИЗИКА том 64
№ 3
2019
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ
581
Рис. 3. Изменение формы потенциала действия вдоль цепочки пейсмекерно-предсердных элементов. Граница
между пейсмекерными и предсердными элементами отмечена стрелкой.
V = KV V ′,
t = Ktt′,
c
c
g
=
K
c
g′
,
(5)
s−1
g
s−1
s−1
c
g
=
K
g′
c,
s
g
c
s
s
(V
−V
)
=
K
(V′
−V′),
s−1
s
(V
−V
s )
s−1
s
s−1
(V
-V
)
=
K
(V′
−V′),
s+1
s
(V
V
s )
s+1
s
s+1-
где KV, Kt,
K
c
,
K
c
,K
c
,
K
,
K
-
g
g
g
(V
−V
s )
(V
−V
s )
s−1
s
s
s−1
s+1
так называемые масштабные коэффициенты, со-
ответствующие мембранному потенциалу, време-
ни, электропроводимость межклеточных соеди-
нений и падению мембранного потенциала вдоль
цепочки клеточных элементов.
Преобразованная система отличается от ис-
ходной только членами уравнений, определяю-
щими связь между клеточными элементами, т.е.
токами через клеточные контакты. В случае подо-
бия электрических процессов контактные токи
исходного и редуцированного уравнений должны
быть равны:
c
c
g
(V
−V
)
=
g
′
(V′
−V′),
s−1
s−1
s
s−1
s−1
s
(6)
c
c
g
(V
−
V
)
=
g′
(V′
−V′
),
s
s+1
s
s
s+1
s
следовательно, комбинации масштабных коэф-
фициентов сходственных параметров редуциро-
ванной модели равны единице:
c
=
1,
g
V
−
V
)
s−1
s−1
s
Рис. 4. Колебания мембранного потенциала вдоль
(7)
цепочки пейсмекерных (а) и предсердных
c
=
1.
g
V
−
V
)
(б) элементов.
s−1
s−1
s
БИОФИЗИКА том 64
№ 3
2019
582
ГАЛАНИН
Период следования спонтанных потенциалов действия в модели интактного синоатриального узла при
увеличении числа ветвей предсердных элементов
Число ветвей предсердных
1
2
3
4
5
6
элементов
T, мс
209
267
324
377
595
-
Рис. 5. Изменение формы потенциала действия вдоль цепочки пейсмекерно-предсердных элементов (а - в модели
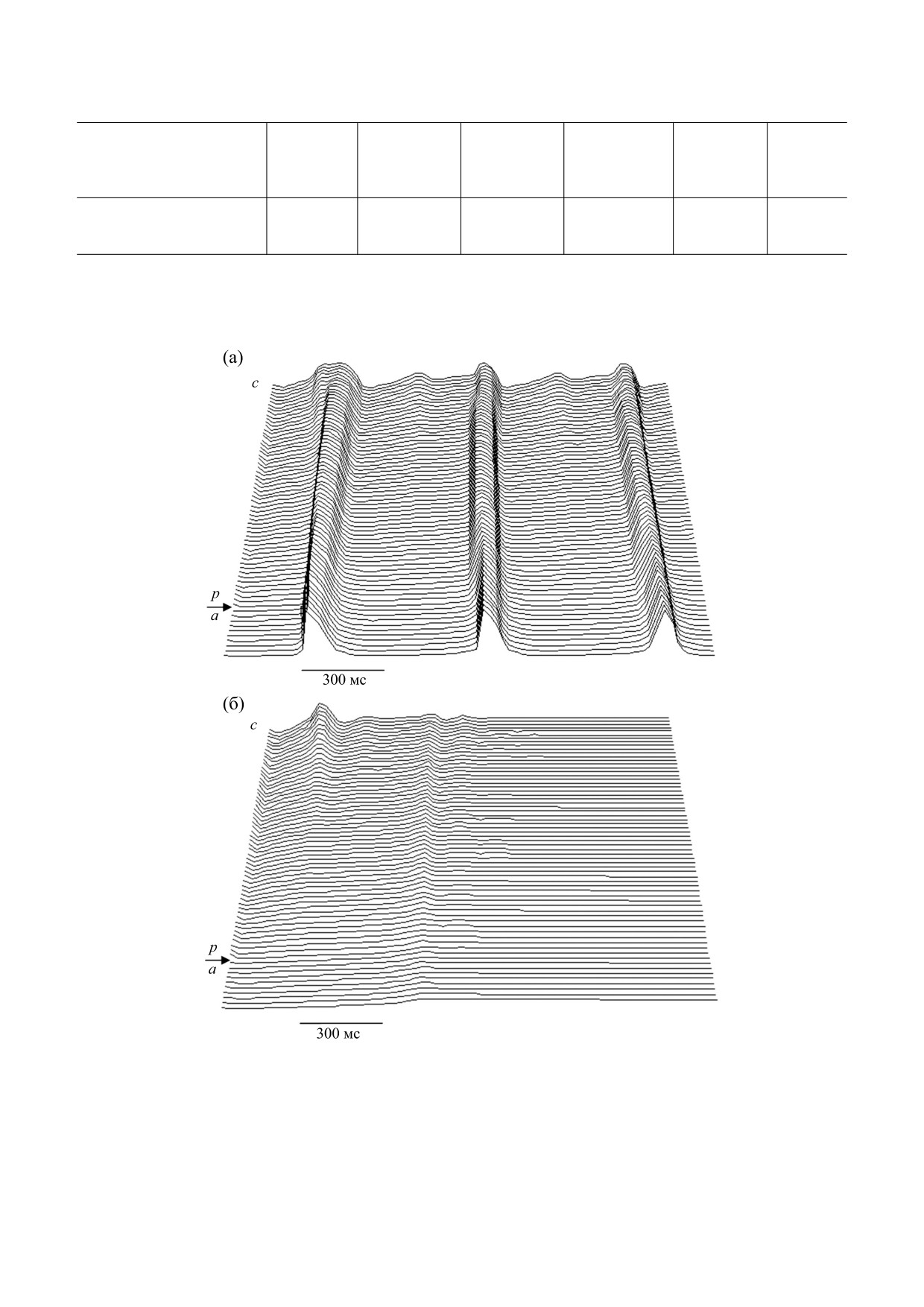
задействовано пять предсердных ветвей; б - в модели задействовано шесть предсердных ветвей).
БИОФИЗИКА том 64
№ 3
2019
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ
583
Рис.
6. Изменение формы потенциала действия вдоль цепочки пейсмекерно-предсердных элементов,
соответствующей редуцированной модели неоднородного интактного САУ
С точки зрения физики изучаемых процессов это
имущество применения метода подобия состоит
означает, что увеличение падения потенциала
в том, что удалось значительно сократить число
(Vs-1 - Vs) и (Vs+1 - Vs) вдоль цепочки за счет изъ-
используемых дифференциальных уравнений,
ятия части ее элементов следует компенсировать,
оставаясь при этом в рамках детального подхода к
уменьшив электропроводимости межклеточных
моделированию электрической активности ти-
соединений в то же число раз. Аналогичная про-
пичных и атипичных кардиомиоцитов. В резуль-
цедура проводится для уравнений, соответствую-
тате исключается необходимость вычисления по-
щих предсердным слоям.
тенциалов действия для всех клеток, входящих в
САУ и типичный миокард предсердия. Данный
Численные оценки показали, что наиболее
подход позволяет проводить численный экспери-
подходящими для масштабных коэффициентов
мент быстрее, удобнее и экономнее.
являются следующие значения:
Также в рамках рассмотренной математической
а) проводимость межклеточных контактов:
модели было показано, что рост числа невозбуди-
K
c
=
20,0;
g
s
мых участков в миокарде предсердия, окружающего
б) мембранный потенциал: KV = 1;
в) время: Kt = 0,975.
При этих оценках редуцированная система
уравнений (4) дает решение, мало отличающееся
от того, которое получалось при использовании
исходной модели. Рис. 6-8 представляют профи-
ли потенциалов действия редуцированной
(сплошные линии) и исходной (пунктирные ли-
нии) моделей. Как видно из рис. 8, редуцирован-
ная модель достаточно хорошо согласуется с ис-
ходной моделью.
ЗАКЛЮЧЕНИЕ
Таким образом, в настоящей работе предложе-
на детальная математическая модель неоднород-
ного интактного САУ и взаимодействующего с
Рис. 7. Колебания мембранного потенциала вдоль
ним типичного миокарда предсердия с использо-
цепочки пейсмекерных элементов редуцированной
ванием методов теории подобия. Основное пре-
модели интактного САУ.
БИОФИЗИКА том 64
№ 3
2019
584
ГАЛАНИН
Рис. 8. Сравнение решений, соответствующих редуцированной (сплошная линия) и исходной (пунктирная линия)
моделей интактного САУ: (а) - s′ = 1, s = 1; (б) - s′ = 7, s = 35; (в) - s′ = 11, s = 55.
пейсмекер, приводит к существенному замедлению
и анатомические особенности изучаемой си-
ритма его электрической активности.
стемы.
В будущем на основе предложенного метода
Автор выражает особую признательность
становится возможным разрабатывать програм-
к.б.н. А.И. Сироте за многочисленные обсуж-
мы для более сложных 3D-моделей САУ и связан-
ного с ним предсердного миокарда, которые бы
дения, полезные советы и дружескую под-
более полно учитывали электрофизиологические
держку.
БИОФИЗИКА том 64
№ 3
2019
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ
585
ПРИЛОЖЕНИЕ 1
МАТЕМАТИЧЕСКАЯ МОДЕЛЬ ПЕЙСМЕКЕРНОЙ КЛЕТКИ
СИНОАТРИАЛЬНОГО УЗЛА
Мембранный потенциал V изолированной пейсмекерной клетки, а также переменные активации и
инактивации Z, f, qt, rt, q, r, y ионных каналов определяются следующей системой дифференциальных
уравнений [8,9-11]:
dV
dZ
d f
dq
q
−
q
d r
r
−
r
d q
d y
Z - Z
f
−
f
t
t
t
t
t
t
q - q
y − y
С
= -I
tot
,
=
,
=
,
=
,
=
,
=
,
=
,
(8)
dt
dt
τ
dt
τ
dt
τ
dt
τ
dt
τ
dt
τ
Z
f
qt
rt
q
y
в которой полный мембранный ток:
(
0,015V
+1,8
)
I
tot
=
I
Na
+
I
Ca
+
I
to
+
I
sus
+
I
K
(1
+
0,6
e
)
+
I
f
+
I
b
(9)
образован суммой следующих токов: натрие-
прямления IK, активируемого при гиперпо-
вого INa, кальциевого ICa, 4-AP-чувствитель-
ляризации If и фонового Ib токов. Данные
ных Ito и Isus, калиевого задержанного вы-
токи задаются формулами:
I
Na
=σ
Na Na
g P Z
(V -E
Na
),
I
Ca
=σ
Ca Ca
g
d f
(
V -E
Ca
),
I
=
1,03
g
q
r
(V -E
),
I
=
1,03
g
r
(V -E
),
to
to
t
t
K
sus
sus
t
K
(10)
I
=σ
g r q
(V -E
)
,I
=
1,2
g
y
(
V -E
)
,
K
K K
K
f
f
f
I
b
=
g
b
(V -E
b
).
Соответствующие ионным токам квазистационарные значения кинетических переменных и их по-
стоянные времени имеют вид:
−
(
0,2
V
+15
)
−
(
0,2
V
+15
)
Z
=
1
−
1
(
1
+
e
)
,
τ
Z
=
0,011,
P
=
1
(
1
+
e
)
,
(
0,2
V
+9
)
−
(
0,143
V
+3,3
)
f
=
0,57
1
+
e
,
τ
=
0,043,
d
=
1
1
+
e
,
(
)
fL
(
)
(
0,077
V
+
4,532
)
q
=
1
1
+
e
,
t
(
)
−3
−
3
−
(
0,080
V
+
3,920
)
(
0,270
V
+13,75
)
τ
qt
=
10,1⋅
10
+
65,17
⋅
10
(
0,57
e
+
0,72
e
)
,
−
(0,056
V
−
0,278)
−
(
0,122
V
−0,
611
)
q
=
1,620
e
(
1
+
e
)
,
(11)
⎛
8
-
(
0,032
V
−
0,968
)
⎞
τ
=
0,108
+
e
+
0,007,
q
⎜
−
(
0,500
V
+4,693
)
⎟
⎝
1
+
e
⎠
(
0,111
V
+1,778
)
r
=
1
1
+
e
,
(
)
−
(
0,047
V
+
3,536
)
−
(
0,085
V
+
6,500
)
y = e
(1
+
e
)
,
(
0,085
V
+
6,500
)
τ
y
=
1
(
1
+
e-
)
,
Множители, входящие в формулы ионных то-
Электроемкость мембраны C и равновес-
ков, имеют следующие значения: для перифери-
ные электропроводимости gi ионных каналов
ческих клеток САУ - σNa = 1, σCa = 2,2, σK = 2,5;
для центральных и периферийных клеток уз-
для центральных клеток САУ - σNa = 0, σCa = 1,15,
ла соответствуют значениям, приведенным в
σK = 2,8.
работе [9].
БИОФИЗИКА том 64
№ 3
2019
586
ГАЛАНИН
ПРИЛОЖЕНИЕ 2
ПОЛНАЯ СИСТЕМА ДИФФЕРЕНЦИАЛЬНЫХ УРАВНЕНИЙ
ИСХОДНОЙ МОДЕЛИ ИНТАКТНОГО САУ
И ВЗАИМОДЕЙСТВУЮЩЕГО С НИМ МИОКАРДА ПРЕДСЕРДИЯ
Пейсмекерные слои:
dV
⎡
⎤
s
1
c
s
=
s
:
C
=-
g
(
V
−
E
)
−
g
(
V
−V
)
,
1
s
1
⎢
∑
is
1
s
1
i
s
1
s
2
s
1
⎥
dt
⎣
i
⎦
dV
s
⎡
c
c
⎤
s
≤
s
≤
s
:
C
=-
V
−
E
)
−
g
(V
−V
)
−
g
(V
−V
)
,
2
M–1
s
⎢
∑gis(
s
i
s−1
s−1
s
s
s+1
s
⎥
dt
⎣
i
⎦
dV
s
M
⎡
c
C
=-
g
(
V
−
E
)
−
g
(
V
−V
)
−
M
s
⎢
∑
is
M
s
M
i
s
M
−1
s
M
−1
s
M
s
=
s
:
dt
⎣
M
i
c
c
c
−
g
(
V
−V
)
−
g
(
V
−
V
)
−
g
V
−
V
⎤
s
M
a
1
s
M
s
M
a
N
+1
s
M
s
M
(
a
2
N
+1
s
M
)
⎦
Предсердные слои:
dV
a
1
⎡
c
c
⎤
a=a:
C
=-
g
(
V
−
E
)
−
g
(
V
−
V
)
−
g
(
V
−V
)
,
1
a
1
⎢
∑
ia
1
a
1
i
s
M
s
M
a1
a
1
a
2
a
1
⎥
dt
⎣
i
⎦
dV
a
⎡
c
c
⎤
a
2
≤
a≤a
N-1
:
C
a
=-
g
a a
V -E
i
)
−
g
a−1
(
V
a
−
1
−
V
a
)
−
g
(
a a
V
+1
−V
a
)
,
⎢
⎥
dt
⎣
i
⎦
dV
⎡
⎤
aN
c
a=a
:
C
=-
g
(
V
−
E
)
−
k
g
(
V
−
V
)
N
a
N
⎢
∑
ia
N
a
N
i
a a
N
−1
a
N
−1
a
N
⎥
dt
⎣
i
⎦
Аналогичные уравнения имеют место для
4. A. L. Hodgkin and A. F. Huxley, J. Physiol. (Lond.)
предсердных слоев с номерами aN + 1 ≤ a ≤ a2N и
117, 500 (1952).
a2N ≤ a ≤ a3N.
5. Р. А. Сюняев и Р. Р. Алиев, Биофизика 55 (6), 1132
(2010).
6. S. Inada, H. Zhang, J. O. Tellez, et al., PLoS One 9 (4),
СПИСОК ЛИТЕРАТУРЫ
e94565 (2014). DOI: 10.1371/journal.pone.0094565.
7. H. Zhang, Y. Zhao, M. Lei, et al., Am. J. Physiol. Heart
1. M. R. Boyett, H. Honjo, and I. Kodama, Cardiovasc.
Circ. Physiol. 292, 165 (2007).
Res. 47, 658 (2000).
8. В. В. Галанин, Биофизика 63 (1), 134 (2018).
2. E. E. Verheijck, A. Wessels, A. C. van Ginneken, et al.,
9. H. G. Zhang, A. V. Holden, I. Kodama, et al., Am. J.
Circulation 97, 1623 (1998).
Physiol. Heart Circ. Physiol. 279, 397 (2000).
3. Д. В. Абрамочкин, Г. С. Сухова и Л. В.
10. D. G. Tsalikakis, H. G. Zhang, et al., Computers in Bi-
Розенштраух, Успехи физиол. наук
40
(4),
21
ology and Medicine 37, 8 (2007).
(2009).
11. B. Wohlfart and G. Ohlen, Clin. Physiol. 19, 11 (1999).
Mathematical Simulation of the Rabbit Heterogeneous Intact Sinoatrial Node
Using Similitude Theory
V.V. Galanin
Medical University “Reaviz”, Chapayevskaya ul. 227, Samara, 443001 Russia
In this work, the electrical activity of the heterogeneous intact sinoatrial node at interaction with the atrial
myocardium is simulated based on a detailed approach. The reduction of this model using the methods of the
theory of similarity. The results of the calculation of the reduced model are compared with the solutions ob-
tained in the initial model. The model also found that an increase in the number of non-excitable sites in the
atrial myocardium bordering the pacemaker causes a slowdown in the overall rhythm of electrical activity.
Keywords: sinoatrial node, atrium, cardiomyocyte, mathematical simulation, similarity theory, complex systems
БИОФИЗИКА том 64
№ 3
2019