БИОФИЗИКА, 2019, том 64, № 4, с. 661-667
МОЛЕКУЛЯРНАЯ БИОФИЗИКА
УДК 577.32
ОПРЕДЕЛЕНИЕ АФФИННОСТИ СВЯЗЫВАНИЯ ВИРИОНОВ
ПАНДЕМИЧЕСКОГО ВИРУСА ГРИППА A/California/7/2009 (H1N1pdm09)
С ФЕТУИНОМ И МУЦИНОМ КОНКУРЕНТНЫМ
И КИНЕТИЧЕСКИМ МЕТОДАМИ
© 2019 г. Г.С. Онхонова, С.В. Мальцев, А.Б. Рыжиков
Государственный научный центр вирусологии и биотехнологии “Вектор” Роспотребнадзора,
630559, р.п. Кольцово, Новосибирская область, Россия
E-mail: vector@vector.nsc.ru
Поступила в редакцию 20.12.2017 г.
После доработки 29.04.2019 г.
Принята к публикации 30.04.2019 г.
Изучена аффинность связывания пандемического вируса гриппа A/California/7/2009 (H1N1pdm09)
с фетуином и муцином. Проведено сравнение трех методов на основе иммуноферментного анализа
для количественного определения рецепторной специфичности штаммов вируса гриппа - конку-
рентного метода Диксона в прямых координатах, конкурентного метода с использованием концен-
трации полумаксимального ингибирования IC50 и кинетического метода. Обнаружено, что конку-
рентные методы существенно искажают результат из-за неспецифического связывания конкуриру-
ющих белков, в то время как кинетический метод исключает подобное влияние и в будущем может
быть широко использован для точного измерения рецепторной специфичности вируса гриппа с ре-
цепторами типа α2-3 и α2-6.
Ключевые слова: рецепторная специфичность, равновесная константа диссоциации, сиаловая кислота.
DOI: 10.1134/S0006302919040033
цифичности связывания различных субтипов ви-
Грипп - это острая вирусная инфекция, кото-
руса гриппа с клеточными рецепторами.
рая, циркулируя по всему миру, может заражать
Существует две основные химические формы си-
млекопитающих, птиц и людей. Являясь одной из
аловой кислоты (α2-3 и α2-6) и разные штаммы
основных причин заболеваемости и смертности,
вируса гриппа отличаются сродством к ним [1].
грипп вызывает ежегодные эпидемии и даже пан-
Штаммы вируса птичьего гриппа используют в
демии и представляет серьезную проблему для
качестве рецептора сиаловые кислоты, прикреп-
здравоохранения и экономики. В процессе жиз-
ленные к следующему остатку сахара посред-
ненного цикла все вирионы проходят обязатель-
ством связи α2-3 (Neu5Acα2-3Gal) [2]. Рецепто-
ную стадию - прикрепление к поверхности клет-
ры такого типа главным образом содержатся на
ки-мишени. Для каждого вируса имеется специ-
эпителиальных клетках пищеварительного трак-
фический рецептор на поверхности чув-
та птиц. Штаммы вируса человеческого гриппа
ствительных клеток, и в свою очередь у вируса
преимущественно прикрепsляются к сиаловым
есть белок, посредством которого он связывается
кислотам со связью α2-6 (Neu5Acα2-6Gal) [3].
с клеточным рецептором. У вируса гриппа это бе-
Такие рецепторы являются основными для кле-
лок гемагглютинин, а рецепторы на поверхности
ток эпителия верхних дыхательных путей челове-
клетки - это гликолипиды или гликопротеины,
ка. При этом небольшое количество α2-3 рецеп-
терминальным фрагментом которых является си-
торов присутствует на реснитчатом эпителии и
аловая кислота (в основном N-ацетилнейрами-
эпителиальных клетках нижних дыхательных пу-
новая кислота (Neu5Ac)).
тей человека. По этой причине существует риск
Структурное различие остатков сиаловых кис-
инфицирования людей вирусом высокопатоген-
лот, прикрепленных к гликановым цепям, явля-
ного птичьего гриппа, и такие ситуации для чело-
ется ключевым фактором для определения спе-
века, как правило, сопровождаются осложнения-
ми вплоть до летального исхода.
Сокращения: ИФА - иммуноферментный анализ, ФСБ -
фосфатно-солевой буфер, ФСБ-Т - фосфатно-солевой
Одним из методов для оценки рецепторной
буфер с добавлением Tween-20.
специфичности вирусов гриппа является имму-
661
662
ОНХОНОВА и др.
ноферментный анализ (ИФА). Для оценки ре-
константа скорости прямой реакции, k- - кон-
зультатов, полученных в ИФА, используют ста-
станта скорости обратной реакции, t - время. По-
ционарные конкурентные и кинетический мето-
казатель экспоненты k подбирается путем ап-
ды. Не смотря на простоту и относительно
проксимации экспериментальной зависимости
низкую стоимость постановки, ИФА имеет ряд
концентрации лиганд-рецепторных комплексов
недостатков. Во-первых, для всех видов анализа
от времени функцией (1). Если k определено при
характерен довольно длительный период прове-
двух и более концентрациях рецептора, то, по-
дения эксперимента, что для большой выборки
строив зависимость рассчитанных параметров k
бывает неудобно. Во-вторых, при полуколиче-
от концентрации добавленных рецепторов, ре-
ственном методе ИФА невозможно сравнить зна-
грессионными методами можно получить урав-
чения между различными штаммами вируса
нение прямой, которое позволяет определить k+ и
гриппа. При количественных конкурентных ме-
k-, а следовательно, и равновесную константу
тодах существует проблема неспецифического
диссоциации Kd.
связывания конкурирующих белков, что в неко-
Конкурентные методы определения константы
торых случаях приводит к значительной погреш-
диссоциации. При конкурентном связывании в
ности измерения. Кроме того, конкурентные ме-
нашем случае два типа рецепторов взаимодей-
тоды корректны только при определенных строго
ствуют с лигандом по следующей схеме [6]:
подобранных соотношениях конкурирующих мо-
лекул и их констант диссоциации. Подобрать ра-
[L] [
R
]
0
1
0
бочие соотношения и концентрации бывает до-
[
L
R
]
=
,
(5)
1
[
R
]
статочно трудоемко, и поэтому этот этап часто
2 0
[
R
1
]
+
K
d1
(1
+
)
игнорируется исследователями. Кинетический
0
K
d2
метод ИФА, опробованный в данной работе, ли-
где [L1R] - концентрация комплексов, [L]0 - кон-
шен перечисленных недостатков полуколиче-
ственного и конкурентного анализа и предлагает-
центрация лиганда, [R1]0, [R2]0 и Kd1, Kd2 - концен-
ся как наиболее оптимальный метод для измере-
трации и константы диссоциации конкурирую-
ния рецепторной специфичности штаммов
щих рецепторов.
вируса гриппа.
В случае конкурентного метода Диксона кон-
стантой диссоциации является значение абсцис-
сы точки пересечения кривых вытеснения одного
МАТЕРИАЛЫ И МЕТОДЫ
рецептора вторым при различных концентрациях
Кинетический метод определения константы
первого рецептора. По теории точка пересечения
диссоциации. Кинетика взаимодействия вируса
кривых вытеснения лежит в отрицательной обла-
гриппа с рецепторами может быть описана кине-
сти и является результатом графического реше-
тикой лиганд-рецепторного взаимодействия.
ния системы уравнений с двумя неизвестными,
Моделируя процесс инфицирования, мы предпо-
одна из которых и есть равновесная константа
лагаем, что вирус в малом количестве попадает на
диссоциации.
эпителий дыхательных путей, покрытый избыт-
Еще одним конкурентным методом измерения
ком рецепторов. Выражение для концентрации
константы диссоциации является метод с исполь-
образовавшегося комплекса в условиях малости
зованием концентрации полумаксимального ин-
концентрации лиганда по сравнению с концен-
гибирования IC50. Равновесную константу диссо-
трацией рецепторов, в которых система находи-
циации в данном методе можно определить сле-
лась в данном исследовании, выглядит следую-
дующим образом:
щим образом [5]:
kt
IC
50
[LR]
=
A(1−
e-
),
(1)
K
=
,
(6)
d2
[
R]
10
1+
[R] [L]
0
0
K
A
=
,
(2)
d1
K
d
+
[
R
]
0
где IC50 - концентрация второго рецептора, ин-
k = k R]0
+
k
,
(3)
гибирующая связывание первого рецептора на
+
-
50%.
k
-
K
d
=
,
(4)
В данной работе в качестве первого рецептора
k+
(конкурента) использовали белки муцин и фету-
ин, в качестве второго - те же белки, меченные
где [LR] - концентрация комплексов лиганд-ре-
биотином. Считая, что константы диссоциации
цептор, [R]0 - начальная концентрация рецепто-
для меченого белка и конкурента равны, полу-
ров, [L]0 - начальная концентрация лиганда (ан-
чаем:
тигена), Kd - равновесная константа диссоциа-
ции лиганд-рецепторных комплексов, k+
-
Kd = IC50 - [R1]0.
(7)
БИОФИЗИКА том 64
№ 4
2019
ОПРЕДЕЛЕНИЕ АФФИННОСТИ СВЯЗЫВАНИЯ
663
Характеристики использованного в работе
ну. Далее проводили подбор кинетических моде-
штамма. Для исследования был выбран вирусный
лей для анализа результатов ИФА и затем -
штамм A/California/7/2009 (H1N1pdm09). Суб-
непосредственное измерение аффинности связы-
тип H1N1 стал причиной пандемий в 1918 и
вания выбранного штамма вируса гриппа тремя
2009 гг., а также причиной эпидемии в России в
методами - методом Диксона в прямых коорди-
2016 г. Штамм H1N1pdm09, ставший серьезной
натах, методом с использованием концентрации
проблемой для здравоохранения, впервые был за-
полумаксимального ингибирования IC50 и с по-
фиксирован в Северной Америке в апреле 2009 г.
мощью кинетического метода.
По официальным данным, распространившись
Очистка вируса гриппа. Вирусную аллантоис-
на более чем 200 стран, он стал причиной более
ную жидкость (100 мл) с инактивированным ма-
18000 смертей по всему миру. Пандемический ви-
териалом фильтровали через фильтр с диаметром
рус H1N1 распространяется среди людей как ви-
пор
0,22 мкм и центрифугировали
3 ч при
рус обычного сезонного гриппа и имеет схожие
20000 об/мин на центрифуге Sigma 3-30K с рото-
симптомы. Однако при тяжелой форме течения
ром 12150-H. Надосадочную жидкость сливали, а
болезни развивается вирусная пневмония, без ле-
бляшку растворяли в 0,5 мл фосфатно-солевого
чения которой смертность очень высока. В ряде
буфера (ФСБ). Полученный концентрат хромато-
исследований [7,8] была изучена рецепторная
графически очищали методом гель-фильтрации
специфичность
связывания
штамма
на колонке с объемом смолы 20 мл (Sepharose-4B,
H1N1(pdm2009) и было выяснено, что он предпо-
Pharmacia, Швеция). Колонку уравновешивали
чтительно связывается с рецепторами типа α2-6.
буфером ФСБ. Концентрацию белка в итоговом
Однако по данным работы [9] утверждается так-
растворе вирусных частиц измеряли методом
же, что этот штамм может связываться с рецепто-
Бредфорда.
рами типа α2-3, но с меньшей аффинностью.
Конкурентный метод с использованием ИФА.
Модели рецепторов. В качестве моделей рецеп-
Очищенный вирус в ФСБ сорбировали на стан-
торов на поверхности чувствительных клеток бы-
дартный 96-луночный планшет для проведения
ли использованы белки фетуин и муцин. Вирус
иммунологических реакций (SPL Life Science,
гриппа имеет сродство к данным белкам через
Корея) в течение ночи при комнатной температу-
взаимодействие с сиаловыми кислотами. Из ли-
ре с концентрацией 10 мкг/мл по 50 мкл до пол-
тературных данных известно, что фетуин содер-
ного высыхания антигена. Планшет промывали
жит α2-3 и α2-6 рецепторы [10], муцин - преиму-
раствором ФСБ с добавлением 0,05% Tween-20
щественно α2-6 рецепторы [11,12], хотя количе-
(ФСБ-Т) с помощью устройства для промывки
ство и тип сиаловых кислот зависит от вида
микропланшет PW40 (Bio-Rad, США). Далее
организма и разные типы муцина могут содер-
планшет заполняли биотинилированным муци-
жать различные типы рецепторов [13,14]. Моле-
ном/фетуином (Sigma-Aldrich, США) в растворе
кулярная масса фетуина, использованного в дан-
для разведения сыворотки (ЗАО «Эпитек», Ново-
ной работе, составляет 48,4 кДа. В отличие от фе-
сибирск, Россия) и проводили серийные двукрат-
туина, муцин сильно дифференцирован по
ные разведения по горизонтали, начиная с кон-
молекулярной массе. По имеющимся на сего-
центраций 0,2 мкг/мл и 10 мкг/мл соответствен-
дняшний день данным приводятся ее оценки в
но. Также проводили разведения по вертикали
очень широком диапазоне - от 10 до 1000 кДа.
немеченого муцина/фетуина, начиная с концен-
Мы дополнительно провели электрофорез в де-
трации 500 мкг/мл. Смесь раскапывали в двух по-
натурирующих условиях, который показал, что
вторениях: в четыре столбца с присушенным ви-
минимальную молекулярную массу муцина мож-
русом и четыре пустых столбца для контроля. Да-
но считать примерно равной массе использующе-
лее все действия выполняли согласно
гося в работе фетуина. В итоге массу используе-
стандартному протоколу: планшет промывали,
мого в работе муцина оценили как среднее гео-
заполняли раствором конъюгата (стрептавидин-
метрическое
известного
из
литературы
полипероксидаза хрена, ЗАО «Биосан», Новоси-
диапазона, т.е. 100 кДа, что не противоречит дан-
бирск, Россия) в ФСБ-Т с концентрацией
ным электрофореза.
1 : 10000, выдерживали 40 ми при комнатной тем-
Иммуноферментный анализ. Для определения
пературе и снова промывали. После промывки от
рецепторной специфичности был выбран имму-
конъюгата планшет заполняли по 100 мкл в лунку
ноферментный анализ. Первым этапом работы
смесью (1:30) тетраметилбензидина и буферного
стала оптимизация условий прямого и конку-
раствора для субстрата (ЗАО «Эпитек», Новоси-
рентного взаимодействия вируса гриппа приме-
бирск, Россия) и инкубировали 30 мин в темноте
нительно к модельным белкам - фетуину и муци-
при комнатной температуре. После этого добав-
БИОФИЗИКА том 64
№ 4
2019
664
ОНХОНОВА и др.
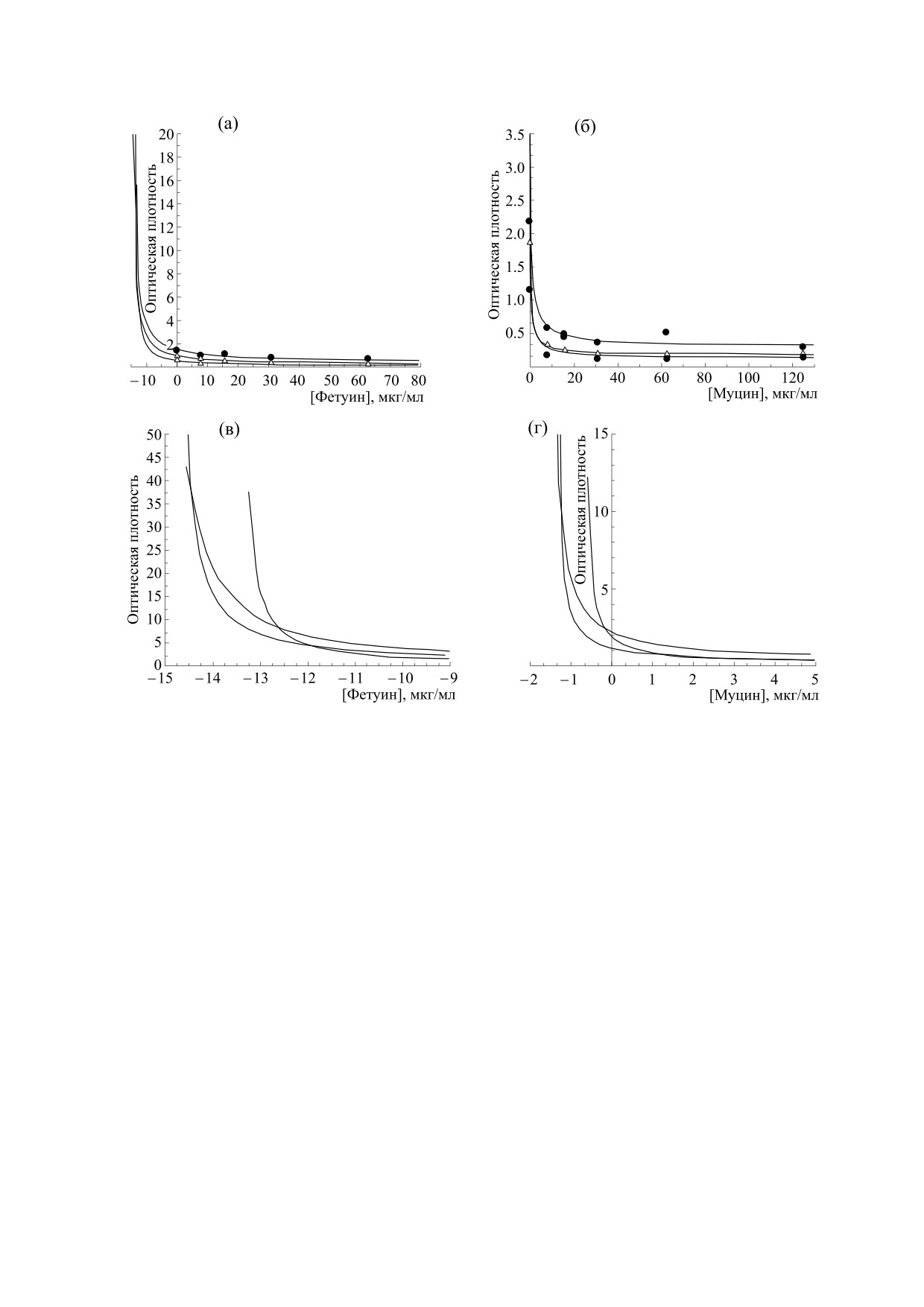
Рис. 1. Зависимости оптической плотности от концентрации конкурентного белка фетуина (а) и муцина (б). Три
кривых вытеснения соответствуют трем различным разведениям биотинилированного белка для штамма вируса
гриппа A/California/7/2009 (H1N1pdm09). На графиках (в) и (г) представлены соотвествующие зависимости в
увеличенном масштабе.
ляли по 50 мкл стоп-реагента (H2SO4, 5%) в лунку
истечении 120 мин планшет промывали, заполня-
и измеряли поглощение на фотометре Multiskan
ли раствором конъюгата в ФСБ-Т с концентраци-
ей 1:10000, выдерживали 40 мин при комнатной
EX (Labsystems, Финляндия) на длине волны 450
температуре и снова промывали. Далее планшет
нм.
заполняли по 100 мкл в лунку смесью (1 : 30) тет-
Кинетический метод с использованием ИФА.
раметилбензидина и буферного раствора для суб-
Для эксперимента очищенный вирус сорбирова-
страта и инкубировали 30 мин в темноте при ком-
ли на планшет по 50 мкл с концентрацией по об-
натной температуре. Для остановки реакции до-
щему белку 10 мкг/мл в течение ночи при комнат-
бавляли по 50 мкл стоп-реагента (H2SO4, 5%) в
ной температуре до полного высыхания антигена.
лунку и измеряли поглощение на фотометре Mul-
После сорбции лунки промывали раствором
tiskan EX на длине волны 450 нм.
ФСБ-Т. Муцин и фетуин, меченные биотином,
разводили в растворе для разведения сыворотки и
проводили серийные двукратные разведения по
РЕЗУЛЬТАТЫ
вертикали в четыре лунки, начиная с 0,5 мкг/мл и
10 мкг/мл соответственно. Каждые 10 мин один
По результатам конкурентных ИФА были по-
стрип снимали с планшета, промывали раство-
лучены кривые вытеснения для штамма вируса
ром ФСБ-Т и убирали в холодильник (4°C). По
гриппа A/California/7/2009 (H1N1pdm09) (рис. 1)
БИОФИЗИКА том 64
№ 4
2019
ОПРЕДЕЛЕНИЕ АФФИННОСТИ СВЯЗЫВАНИЯ
665
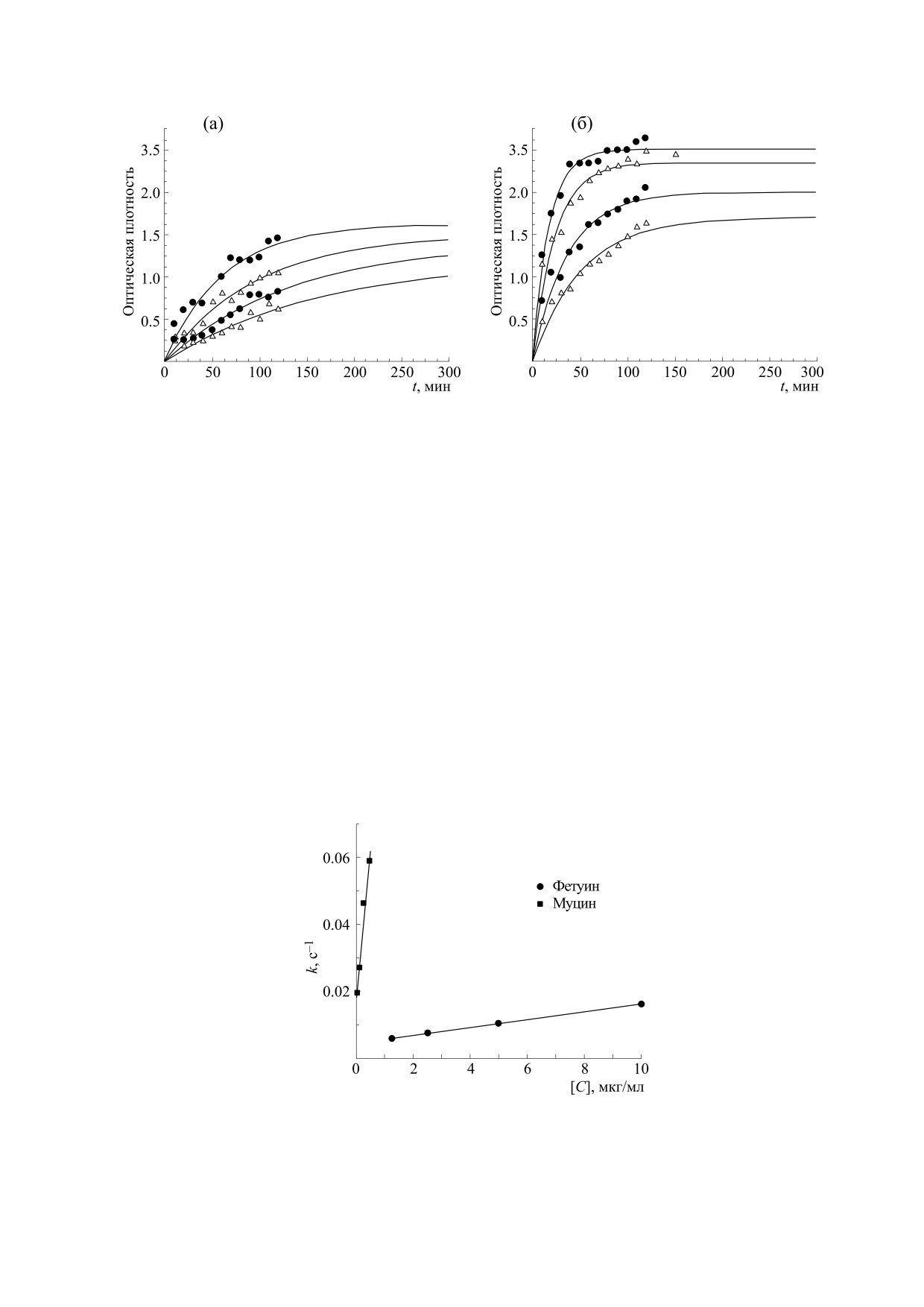
Рис. 2. Кинетика связывания штамма вируса гриппа A/California/7/2009 (H1N1pdm09) с фетуином (а) и муцином (б).
Четыре кривые соответствуют четырем различным разведениям биотинилированного белка.
и рассчитаны равновесные константы диссоциа-
ми вычислены равновесные константы диссоци-
ции методом Диксона в прямых координатах и
ации в соответствии с формулой (4).
методом с использованием концентрации полу-
В таблице содержатся значения равновесных
максимального ингибирования IC50. Значения
констант диссоциации для выбранного штамма
точек пересечения кривых и параметра IC50 были
вируса гриппа и двух белков, рассчитанные тремя
определены с помощью пакета программ
методами: методом Диксона в прямых координа-
тах, методом с использованием концентрации
Scilab 4.0.
полумаксимального ингибирования IC50 и с по-
Экспериментальные точки кинетического
мощью кинетического метода.
ИФА аппроксимировали функцией (1) при помо-
щи пакета программ Origin 8 (рис. 2) в целях вы-
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
числения коэффициентов k в показателе экспо-
ненты при различных начальных концентрациях
Значения константы аффинности вируса
фетуина и муцина. Далее были построены зави-
гриппа A/California/7/2009 (H1N1pdm09) с фету-
симости рассчитанных параметров k от началь-
ином, равные обратным значениям равновесной
ной концентрации добавленного белка (фетуина
константы диссоциации, измеренные методом
или муцина) (рис. 3), и регрессионными метода-
Диксона в прямых координатах и методом IC50,
Рис. 3. Зависимости рассчитанных параметров k от концентрации фетуина и муцина для штамма вируса гриппа
A/California/7/2009 (H1N1pdm09).
БИОФИЗИКА том 64
№ 4
2019
666
ОНХОНОВА и др.
Таблица 1. Равновесные константы диссоциации для штамма вируса гриппа A/California/7/2009 (H1N1pdm09) с
фетуином и муцином
Kd, нМ
Метод
Фетуин
Mуцин
Метод Диксона в прямых координатах
272.9 ± 12.2
12.2 ± 5.9
Метод с использованием концентрации
192.9 ± 43.7
49.1 ± 2.3
полумаксимального ингибирования (IC50)
Кинетический метод
81.6 ± 74.9
1.68 ± 0.38
отличаются в среднем не более чем на
ным белком, что в особенности было продемон-
1,5 мкмоль-1. Измерения кинетическим методом
стрировано в данной работе на примере муцина.
демонстрируют бóльшую аффинность в сравне-
нии с конкурентными методами - средняя разни-
СПИСОК ЛИТЕРАТУРЫ
ца составляет 6,8-8,3 мкмоль-1. Значение аффин-
ности вируса с муцином, измеренное кинетиче-
1.
Т. Н. Ильичева, С. В. Нетесов и В. Н. Гуреев,
ским методом, также превышает значения,
Вирусы
гриппа:
методическое
пособие.
полученные конкурентными методами. Однако
(Новосибирский государственный университет,
разница между кинетическим и конкурентными
Новосибирск, 2014), ч. 1.
методами по сравнению с фетуином более суще-
2.
A. S. Gambaryan, S. S. Iamnikova, D. K. L’vov, et al.,
ственна - в среднем 513,2-574,8 мкмоль-1. По-
Mol. Biol. 36, 542 (2002).
добная разница между конкурентными и кинети-
ческим методами объясняется влиянием неспе-
3.
K. Shinya, M. Ebina, S. Yamada, et al., Nature 440,
цифического связывания конкурирующих
435 (2006).
белков при конкурентном измерении в ИФА, что
4.
R. McBride, J. C. Paulson, and R. P. de Vries, J. Vis.
занижает реальное значение константы диссоци-
Exp., No 111, e53847, DOI: 10.3791/53847 (2016).
ации. Полученные результаты указывают на то,
5.
С. Д. Варфоломеев и К. Г. Гуревич, Биокинетика:
что влияние неспецифического связывания кон-
курирующих белков на результат очень суще-
практический курс (ФАИР-ПРЕСС, М., 1999).
ственно в отношении муцина и умеренно для фе-
6.
С. Д. Варфоломеев и С. В. Зайцев, Кинетические
туина.
методы в биохимических исследованиях (Изд-во
Моск. ун-та, М., 1982).
ВЫВОДЫ
7.
T. R. Maines, A. Jayaraman, J. A. Belser, et al., Science
325, 484 (2009).
Широко использующиеся конкурентные ме-
8.
R. Xu, R. McBride, C. M. Nycholat, et al., J. Virol. 86,
тоды ИФА для измерения аффинности вируса
гриппа относительно адекватно работают с фету-
982 (2012).
ином, но существенно искажают результат при
9.
R. A. Childs, A. S. Palma, S. Wharton, et al., Nat. Bio-
использовании муцина. Отработанный в рамках
technol. 27, 797 (2009).
настоящей работы кинетический метод имеет
10.
J. U. Baenziger and D. Fiete, J. Biol. Chem. 254 (3),
преимущество перед конкурентными методами в
789 (1979).
случае наличия проблемы неспецифического
связывания для некоторых белков.
11.
S. Mizan, A. Henk, A. Stallings, et al., J. Bacteriol. 182,
6874 (2000).
Таким образом, кинетический метод может
применяться для измерения рецепторной специ-
12.
H. Schachter, E. J. McGuire, and S. Roseman, J. Biol.
фичности вирусов гриппа А наряду с конкурент-
Chem. 246, 5321 (1971).
ными методами в ИФА, а также остальными су-
13.
S. C. Baos, D. B. Phillips, L. Wildling, et al., Biophys.
ществующими методами. Кроме того, это наибо-
J. 102, 176 (2012).
лее оптимальный метод ИФА для измерения
аффинности связи вируса с белками, имеющими
14.
M. C. Rose and J. A. Voynow, Physiol. Rev. 86 (1), 245
природное свойство связываться с себе подоб-
(2006).
БИОФИЗИКА том 64
№ 4
2019
ОПРЕДЕЛЕНИЕ АФФИННОСТИ СВЯЗЫВАНИЯ
667
Determination of the Binding Affinity of Pandemic Influenza Virus A/California/7/2009
(H1N1pdm09) to Fetuin and Mucin by Competitive and Kinetic Methods
G.S. Onkhonova, S.V. Maltsev, and A.B. Ryzhikov
State Research Center of Virology and Biotechnology “Vector”, Koltsovo, Novosibirsk Region, 630559 Russia
The binding affinity of pandemic influenza virus A/California/7/2009 (H1N1pdm09) to fetuin and mucin
was studied. Three methods - a competitive Dixon method in direct coordinates, a competitive method with
the use of the half-maximal inhibitory concentration (IC50) and a kinetic method were used for quantitative
determination of receptor specificity of influenza virus strains. ELISA assay results were compared to those
obtained from each of three methods. It has been found that since non-specific binding of competing proteins
has been observed from each of the competitive methods, there are significant differences in the results, while
the kinetic method excludes such an effect and in the future it can be widely used to measure accurately the
receptor specificity of the influenza virus with sialic acids α2-3 and α2-6.
Keywords: receptor specificity, equilibrium dissociation constant, sialic acid
БИОФИЗИКА том 64
№ 4
2019