БИОФИЗИКА, 2019, том 64, № 4, с. 747-753
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 577.3
АНАЛИЗ ВЗАИМНЫХ КОРРЕЛЯЦИЙ МЕЖДУ КОЛЕБАНИЯМИ ОБИЛИЯ
ПОПУЛЯЦИЙ ПЛАНКТОНА И КОРРЕЛЯЦИЙ МЕЖДУ КОЛЕБАНИЯМИ
ПЛАНКТОННОГО ОБИЛИЯ И ВАРИАЦИЯМИ ТЕМПЕРАТУРЫ
НА ПРИМЕРЕ ЭКОСИСТЕМЫ НАРОЧАНСКИХ ОЗЕР
© 2019 г. Д.А. Тихонов* **, А. Б. Медвинский*
*Институт теоретической и экспериментальной биофизики РАН,
142290, Пущино Московской области, ул. Институтская, 3
**Институт математических проблем биологии РАН - филиал Института прикладной математики
им. М.В. Келдыша
РАН, 142290, Пущино Московской области, ул. Профессора Виткевича, 1
E-mail: alexander_medvinsky@yahoo.com
Поступила в редакцию 08.04.2019 г.
После доработки 08.04.2019 г.
Принята к публикации 11.04.2019 г.
Демонстрируется эффективность использования кросс-корреляционной энтропии Шеннона,
представляющей собой обобщение широко применяемой в научных исследованиях спектральной
энтропии, как метода анализа взаимных корреляций между колебаниями планктонного обилия и
вариациями температуры. На примере анализа временных рядов, характеризующих динамику зоо-
планктона, фитопланктона, бактериопланктона и вариации температуры воды, показывается, в
частности, что наиболее значимыми являются взаимные корреляции между температурой и бакте-
риопланктоном.
Ключевые слова: кросс-корреляционная энтропия Шеннона, планктон, температура.
DOI: 10.1134/S0006302919040148
Колебания численности популяций в боль-
ходе полевых измерений (мониторинга экоси-
шинстве случаев характеризуются нерегулярно-
стем), представляют существенный интерес.
стью и ограниченной предсказуемостью [1,2]. В
Мониторинг экосистем предполагает проведе-
качестве причины такой нерегулярности часто
ние одновременных измерений биотических и
рассматривают динамический хаос, т.е. неустой-
абиотических факторов (см. например, работу
чивость динамики, независимую от внешних
[7]), что позволяет сопоставлять между собой
влияний. Подобная неустойчивость во многих
cоответствующие временные ряды и, в частности,
математических моделях темпоральных вариаций
оценивать степень их сопряженности. Так, в ре-
популяционного обилия проявляется в виде зави-
зультате анализа данных полевых исследований
симости нерегулярных временных рядов от на-
экосистемы Нарочанских озер (Белоруссия) бы-
чальных условий [2-4]. Необходимо, однако,
ло показано [8], что уровень фазовой синхрониза-
учитывать то, что реальные, а не модельные, по-
ции колебаний температуры воды и колебаний
пуляции связаны между собой, образуя сложные
биомассы фитопланктона связан со степенью со-
сети трофических взаимодействий (см., напри-
пряжения колебаний популяционного обилия
мер, [5]), и потому не являются независимыми.
фитопланктона и бактериопланктона.
Кроме того, все природные популяции подверже-
ны воздействию внешних абиотических факто-
Отметим, что адекватность оценки степени со-
ров, таких как: температура, солнечная радиация,
пряжения планктонных осцилляций и их синхро-
ветер, - что далеко не всегда учитывается в ходе
низации с вариациями температуры в значитель-
математического моделирования [6]. Поэтому
ной мере зависят от объема и качества данных,
оценки влияния динамики внешних абиотиче-
полученных в ходе полевых наблюдений. В этой
ских факторов на динамику популяций, наряду с
связи использование таких методов оценки взаи-
исследованиями межпопуляционных взаимодей-
мосвязи между колебаниями обилия популяций
ствий, базирующиеся на данных, полученных в
планктона и связи планктонной динамики с ва-
747
748
ТИХОНОВ, МЕДВИНСКИЙ
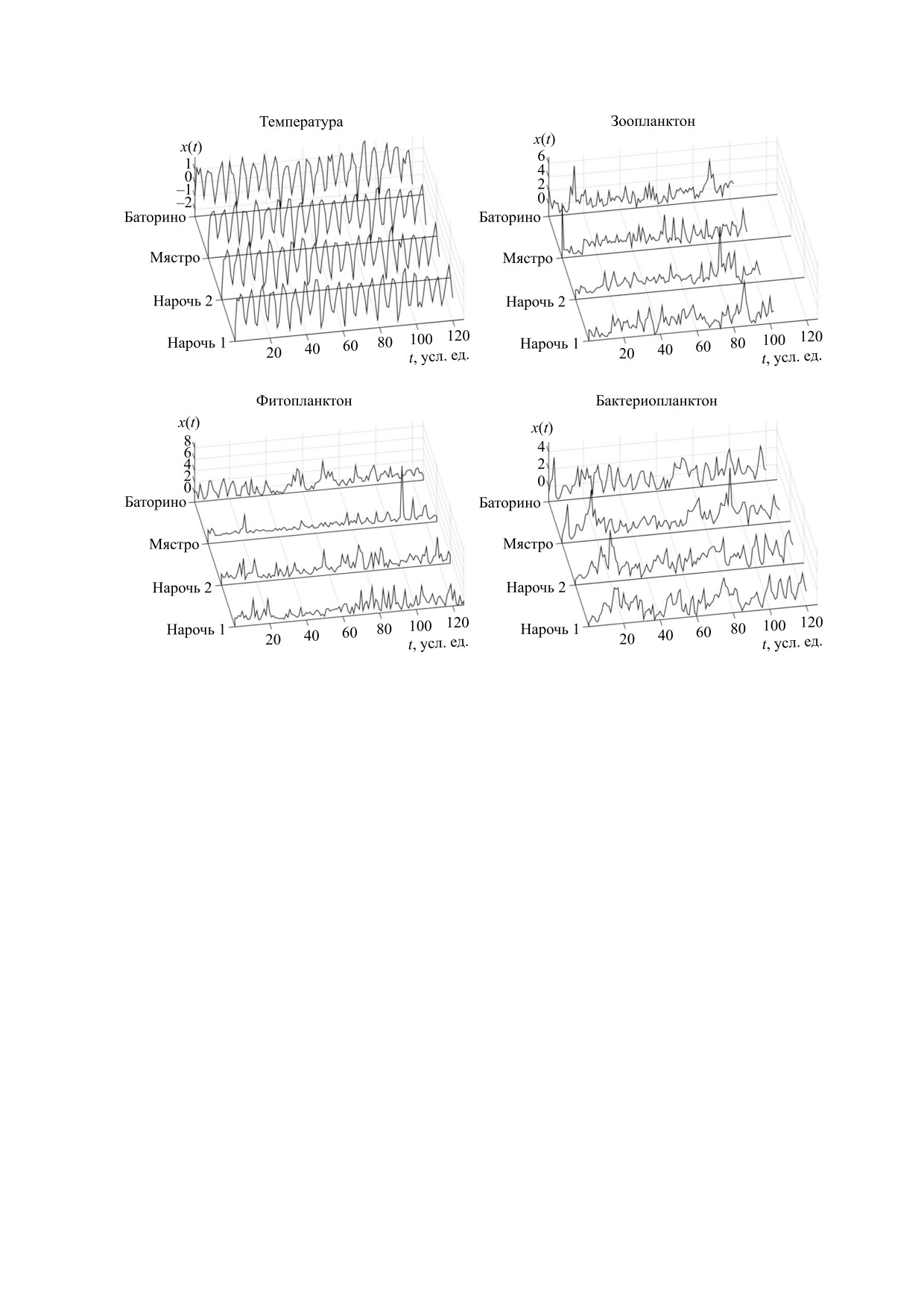
Рис. 1. Временные ряды, иллюстрирующие колебания обилия планктона и вариации температуры воды в каждом из
водоемов, входящих в систему Нарочанских озер. Один шаг по времени соответствует одному месяцу. Обзначения:
Нарочь 1 - Малый плес оз. Нарочь, Нарочь 2 - Большой плес оз. Нарочь.
риациями температуры, которые отличны и неза-
полного водообмена, трофический статус - при-
висимы от методов, применявшихся ранее в ра-
ведены в работе [7].
боте [8], представляется оправданным. Целью на-
Временные ряды, характеризующие колеба-
стоящей работы является установление
ния планктонного обилия и вариации температу-
корреляций между спектрами мощности, кото-
ры воды в Нарочанских озерах, показаны на
рые характеризуют динамику популяций фито-
рис. 1.
планктона, зоопланктона, бактериопланктона, а
также колебания температуры в озерах Нарочан-
Исходные данные, на основании которых по-
ской группы.
строены эти временные ряды, приведены в рабо-
тах [8,9]. Необходимо отметить, что в исходных
данных наличествовало некоторое количество
МАТЕРИАЛЫ И МЕТОДЫ
пропусков, связанных с теми сравнительно ко-
роткими, временными интервалами, в течение
Нарочанские озера, расположенные в северо-
которых измерения не проводились. Нами в ходе
западной части Белоруссии, представляют собой
предварительной обработки такие пропуски за-
систему трех связанных между собой протоками
полнялись средними величинами, исходя из дан-
водоемов: оз. Нарочь, которое, в свою очередь,
ных, которые были получены в соответствующие
подразделяется на Большой плес и Малый плес,
месяцы, но в другие годы. В связи с тем, что ис-
оз. Мястро и оз. Баторино. Основные характери-
ходные данные содержат различное число точек
стики этих водоемов - площадь, объем водной
(120 точек для температуры, 102 точки для зоо-
массы, средняя и максимальная глубины, время
планктона, 126 точек для фитопланктона и 120 то-
БИОФИЗИКА том 64
№ 4
2019
АНАЛИЗ ВЗАИМНЫХ КОРРЕЛЯЦИЙ
749
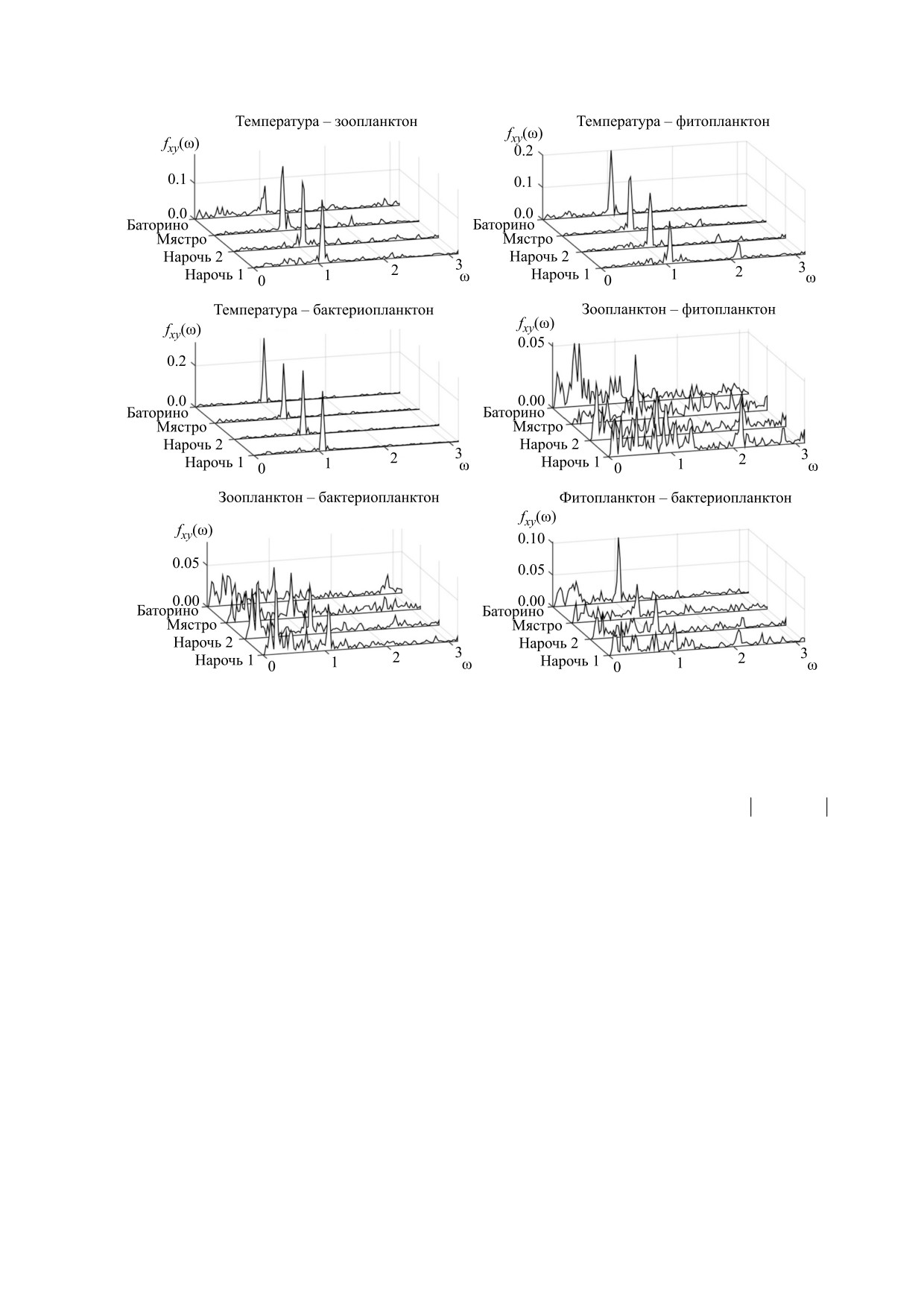
Рис. 2. Спектры взаимных корреляций временных рядов, представленных на рис. 1. Обзначения: Нарочь 1 - Малый
плес оз. Нарочь, Нарочь 2 - Большой плес оз. Нарочь.
чек для бактериопланктона), нами было проведе-
ный спектр (fxy(ω)) взаимных корреляций вре-
но унифицирование длины временных рядов: мы
менных рядов x(t) и y(t):
f
=λ
x ω)y(ω
,
(ω)
)
xy
дополняли нулями недостающие значения так,
где
и
- фурье-преобразования рядов
x ω)
ŷ ω)
чтобы длина каждого временного ряда составляла
x(t) и y(t), а λ - нормировочная константа, кото-
126 точек. Такая унификация может вносить по-
рая выбирается исходя из следующего условия:
грешности в том случае, если исследуются фазо-
ω
max
вые сдвиги между временными рядами. Однако
f
(ω)dω
=
1,
предметом нашего исследования являются вза-
∫
xy
ω
min
имные корреляции временных рядов, при вычис-
лении которых сдвиг фазы колебаний усредняет-
где ωmin и ωmax - верхняя и нижняя границы
ся, поэтому проведенная унификация не влияет
спектров. В соответствии с теоремой Найквиста
существенно на результат вычислений. Во всех
границы спектров заданы, коль скоро задан вре-
менной шаг и число отсчетов временных рядов. В
временных рядах перед проведением анализа
диагональном случае fxx(ω) является нормиро-
корреляций удаляли линейный тренд, кроме то-
ванным спектром автокорреляционной функции
го, все временные ряды приводили к нулевому
для временного ряда x(t) или нормированным
среднему и единичной дисперсии.
сигнала x(t). Подроб-
спектром мощности |
|2
x ω)
Для анализа степени сопряжения временных
ное описание методов анализа степени сопряже-
рядов в данной работе используется нормирован- ния временных рядов приведено в работе [10].
БИОФИЗИКА том 64
№ 4
2019
750
ТИХОНОВ, МЕДВИНСКИЙ
Кросс-корреляционная энтропия Шеннона спектров взаимных корреляций
Нарочь 1
Нарочь 2
Мястро
Баторино
Температура-зоопланктон
3.757
3.816
3.719
4.372
Температура-фитопланктон
4.002
3.793
3.774
3.730
Температура-бактериопланктон
3.252
3.134
3.505
3.149
Зоопланктон-фитопланктон
4.361
4.459
4.565
4.362
Зоопланктон-бактериопланктон
4.106
4.178
4.433
4.393
Фитопланктон-бактериопланктон
4.291
4.310
4.533
4.140
Примечание. Нарочь 1 - Малый плес оз. Нарочь, Нарочь 2 - Большой плес оз. Нарочь.
РЕЗУЛЬТАТЫ
пряженности соответствующих временных ря-
дов. В таблице представлены численные значе-
На рис. 2 показаны спектры взаимных корре-
ния кросс-корреляционной энтропии Шеннона
ляций fxy(ω) для всех парных сочетаний времен-
для каждого из водоемов системы Нарочанских
ных рядов, представленных на рис. 1. Видно, что
озер. Из таблицы видно, что кросс-корреляцион-
спектры взаимных корреляций с участием темпе-
ная энтропия спектров взаимных корреляций,
ратуры характеризуются хорошо выраженной
характеризующихся непрерывной структурой,
прерывной структурой, т.е. наличием отчетливых
выше, чем кросс-корреляционная энтропия
обособленных высокоамплитудных пиков. Такая
спектров, которые имеют прерывную структуру
структура отражает регулярность вариаций тем-
(ср. с рис. 2).
пературы (рис. 1), связанную с сезонностью. В то
же время спектры взаимных корреляций колеба-
Представляет интерес сравнение полученных
ний планктонного обилия характеризуются не-
нами значений кросс-корреляционной энтропии
прерывной структурой. Такая особенность этих
с величиной энтропии случайного сигнала дли-
спектров обусловливается нерегулярной и, как
ной n, для которого на всех частотах (с учетом
показано в работах [8,9], хаотической природой
нормировки) спектр f = 1/n. Для такого случайно-
планктонной динамики.
го сигнала энтропия Шеннона определяется как
Для количественной оценки степени инфор-
1
мативности спектров взаимных корреляций мы
S =−n
ln
=
ln n)
(
(1))
n n
вводим численное значение энтропии Шеннона.
В диагональном случае, когда спектр представля-
При n = 126 S = 4,836. Сравнение этой величи-
ет собой спектр мощности, такая мера хорошо из-
ны с числами, приведенными в таблице, показы-
вестна. Она получила название спектральной эн-
вает, что энтропия случайного сигнала превыша-
тропии и широко используется в различных при-
ет значения энтропии спектров взаимных корре-
ложениях, таких как: распознавание речи,
ляций.
медицинская энцефалографии, механика и кли-
матические исследования [11-14]. Обобщение
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
понятия спектральной энтропии, кросс-корреля-
ционная энтропия Шеннона, позволяет числен-
В данной работе мы вводим кросс-корреляци-
но характеризовать упорядоченность спектров
онную энтропию Шеннона как численную харак-
взаимных корреляций и тем самым - степень со-
теристику сопряженности временных рядов [8,9],
БИОФИЗИКА том 64
№ 4
2019
АНАЛИЗ ВЗАИМНЫХ КОРРЕЛЯЦИЙ
751
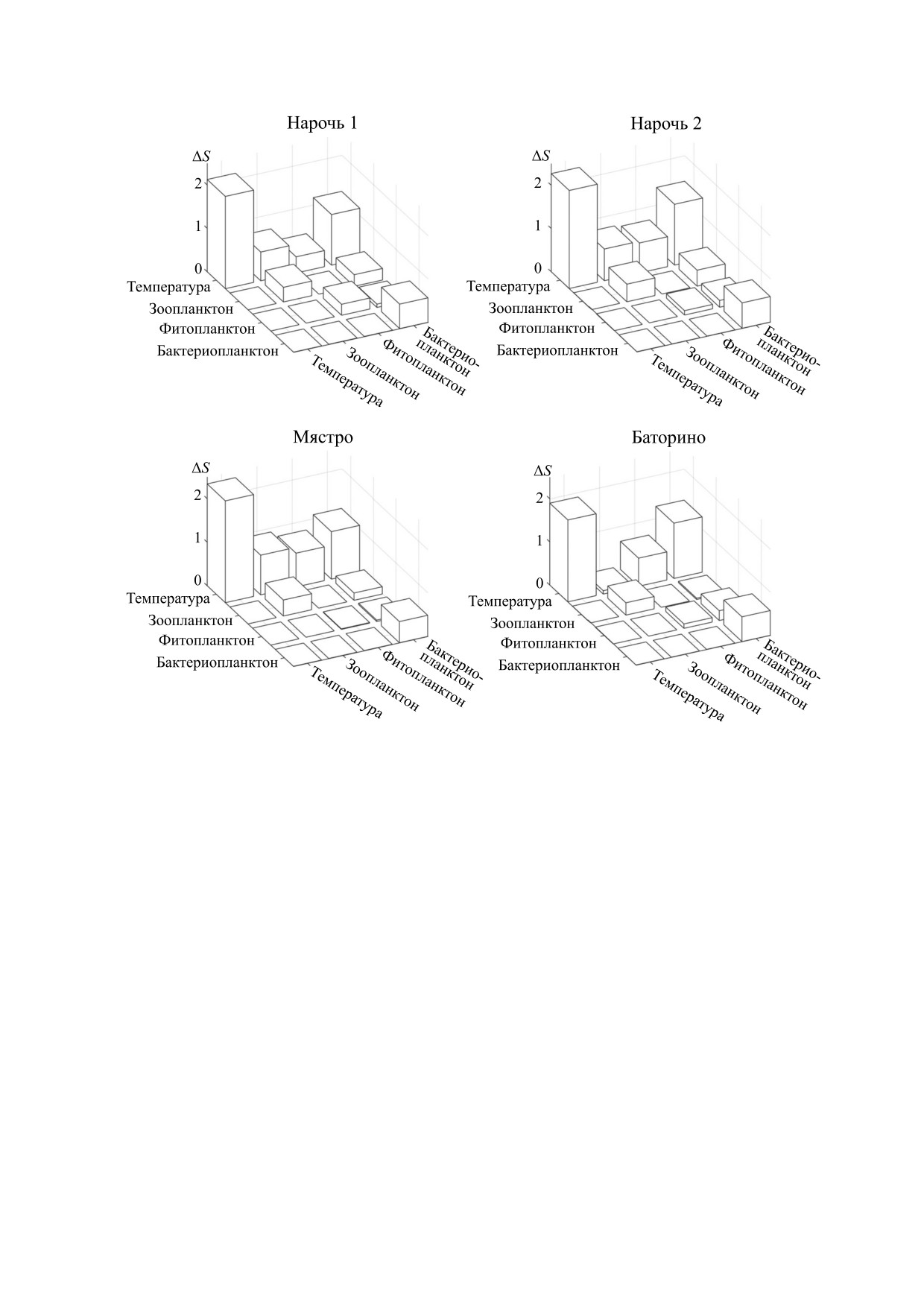
Рис. 3. Гистограммы, представляющие разницу ΔS численных значений кросс-корреляционной энтропии Шеннона,
для спектров взаимной корреляции относительно спектров корреляции спектров колебаний обилия зоопланктона и
фитопланктона для озер Нарочь, Мястро и Баторино (диагональные гистограммы представляют относительные зна-
чения спектральной энтропии). Ввиду того, что матрицы разностей ΔS симметричны, показаны гистограммы только
для диагональных и выше элементов этих матриц. Обзначения: Нарочь 1 - Малый плес оз. Нарочь, Нарочь 2 - Боль-
шой плес оз. Нарочь, темпер. - температура, зоопл. - зоопланктон, фитопл. - фитопланктон, бактпл. - бактерио-
планктон.
полученных в результате длительного монито-
реляциями спектров колебаний обилия планкто-
ринга озерной экосистемы. На рис. 3 представле-
на в разных водоемах. Видно, что наиболее значи-
ны гистограммы, характеризующие разности ве-
мыми являются автокорреляции температуры и
личин кросс-корреляционной энтропии (ΔS) от-
взаимные корреляции между температурой и бак-
носительно базового уровня, в качестве которого
териопланктоном. Отметим, что столь высокие
выбрано значение энтропии для самого неупоря-
корреляции спектров вариаций температуры и
доченного спектра взаимных корреляций спек-
колебаний обилия бактериопланктона естествен-
тров анализируемых нами временных рядов. Для
ным образом связаны с тем фактом, что колеба-
всех водоемов системы Нарочанских озер таким
ния бактериопланктона и вариации температуры
спектром является спектр взаимных корреляций
существенно синхронизованы по фазе [8].
колебаний обилия зоопланктона и фитопланкто-
Необходимо отметить, что взаимная корреля-
на (см. таблицу).
ция временных рядов далеко не всегда сопрягает-
Представленные на рис. 3 гистограммы позво-
ся с фазовой синхронизацией. Например, из рис.
ляют судить об отличиях между взаимными кор-
3 следует, что колебания фитопланктона значимо
БИОФИЗИКА том 64
№ 4
2019
752
ТИХОНОВ, МЕДВИНСКИЙ
коррелированы с вариациями температуры. При
является необходимым условием прогресса в ис-
этом синхронизация колебаний фитопланктона с
следовании механизмов, обусловливающих
вариациями температуры имеет место только в
сложный характер популяционных процессов.
эвтрофированном оз. Баторино [8]. Такую син-
Исследование выполнено при финансовой
хронизацию связывают скорее не с непосред-
поддержке Российского фонда фундаментальных
ственным влиянием температуры на динамику
исследований (грант 17-04-00048).
фитопланктона, а со взаимосвязью динамики
фитопланктона и динамики бактериопланктона;
эта взаимосвязь, в свою очередь, зависит от кон-
СПИСОК ЛИТЕРАТУРЫ
центрации биогенов (в первую очередь, азота и
1.
B. A. Maurer, Untangling Ecological Complexity (Uni-
фосфора) в воде [8]. В озерах Нарочь и Мястро
versity of Chicago, Chicago, 1999).
концентрации биогенов не обусловливают син-
2.
R. V. Solé and J. Bascompte, Self-Organization in Com-
хронизацию колебаний обилия фитопланктона и
plex Ecosystems (Princeton University, Princeton,
2006).
температуры [8]. Тем не менее эти колебания ха-
3.
A. B. Medvinsky, S. V. Petrovskii, I. A. Tikhonova,
рактеризуются существенным уменьшением
et al., SIAM Rev. 44, 311 (2002).
кросс-корреляционной энтропии Шеннона по
4.
А. Б. Медвинский, Н. И., Нуриева, А. В. Русаков и
сравнению с базовым уровнем (таблица). Такой
Б. В. Адамович, Биофизика 62, 107 (2017).
факт может свидетельствовать о том, что помимо
5.
концентраций биогенов в воде водоемов на кор-
TURES/L24/trophic.jpg.
реляцию колебаний фитопланкона с вариациями
6.
P. Turchin, Complex Population Dynamics. A Theoreti-
температуры могут влиять другие факторы (на-
cal/Empirical Synthesis (Princeton University, Prince-
ton, 2003).
пример, трофические связи фитопланктона с
7.
B. V. Adamovich, A. B. Medvinsky, L. V. Nikitina,
зоопланктоном).
et al., Ecol. Indic. 97, 120 (2019).
8.
A. B. Medvinsky, B. V. Adamovich, R. R. Aliev, et al.,
Обращает на себя внимание тот факт, что в
Ecol. Complex. 32, 90 (2017).
озерах Нарочь и Мястро величина ΔS, характери-
9.
A. B. Medvinsky, B. V. Adamovich, A. Chakraborty,
зующая корреляцию спектров колебаний темпе-
et al., Ecol. Complex. 23, 61 (2015).
ратуры и плотности популяций зоопланктона,
10.
Д. А. Тихонов и А. Б. Медвинский, О корреляции
выше, чем в оз. Баторино (рис. 3). Это может обу-
временных рядов в экологии аквасистем (Препринт
словливаться тем обстоятельством, что в эвтро-
№ 49 ИПМ им. М. В. Келдыша, Москва, 2019)
фированном оз. Баторино обилие фитопланктона
11.
V. Sharma and A. Parey, Procedia Engineer. 144, 253
(2016).
настолько велико, что его колебания (синхрони-
12.
J. Shen, J. Hung, and L. Lee, in: Intern. Conf. on Spoken
зованные с температурой [8]) слабо влияют на ко-
Language Processing 3, 1015 (ASSTA, Sydney, 1998).
лебания зоопланктона. В результате корреляции
13.
X. Li, H. Liu, Y. Zheng, B. Xu, in: Bio-Inspired Compu-
спектров вариаций температуры и колебаний
tational Intelligence and Application (Springer, Berlin,
зоопланктона оказываются ослабленными.
Heidelberg, 2007), pp. 36-45.
14.
W. Yi, S. Qiu, H. Qi, et al., J. Neuroeng. Rehabil. 10,
Результаты представленного в данной работе
106 (2013).
анализа временных рядов показывают, что ис-
15.
В. Д. Фёдоров и Т. Г. Гильманов, Экология (Изд-во
пользование кросс-корреляционной энтропии
МГУ, Москва, 1980).
Шеннона является эффективным методом, поз-
16.
M. Kot, Elements of Mathematical Ecology (Cambridge
воляющим приблизиться к пониманию тех меха-
University, Cambridge, 2001).
17.
Г. Ю. Ризниченко и А. Б. Рубин, Биофизическая
низмов, которые обусловливают характер коле-
динамика продукционных процессов (Институт
бательных процессов. Результативность этого ме-
компьютерных исследований, Москва-Ижевск,
тода повышается при его сочетании с другими
2004).
подходами к исследованию таких механизмов. В
18.
S. P. Otto and T. Day, A Biological Guide to Mathemat-
частности, при изучении динамики популяций
ical Modeling in Ecology and Evolution (Princeton Uni-
гидробионтов, как показано в данной работе, по-
versity, Princeton, Oxford, 2007).
19.
H. Kantz and T. Schreiber, Nonlinear Time Series Anal-
лезно сравнивать взаимные корреляции времен-
ysis (Cambridge University, Cambridge, 1997).
ных рядов с оценками их фазовой синхрониза-
20.
D. A. Tikhonov, J. Enderlein, H. Malchow, and
ции. Накопление данных в ходе длительного мо-
A. B. Medvinsky, Chaos, Solitons and Fractals 12, 277
ниторинга экосистем
[7] наряду с матема-
(2001).
тическим моделированием динамики популяций
21.
D. Tikhonov and H. Malchow, Chaos, Solitons and
[15-18] и анализом временных рядов [3,19-21]
Fractals 16, 287 (2003).
БИОФИЗИКА том 64
№ 4
2019
АНАЛИЗ ВЗАИМНЫХ КОРРЕЛЯЦИЙ
753
Analysis of Mutual Correlations between Fluctuations in Plankton Population
Abundances and Correlations between Plankton Abundance Fluctuations
and Temperature Variations as Applied to the Ecosystem of Naroch Lakes
D.A. Tikhonov* ** and A.B. Medvinsky*
*Institute of Theoretical and Experimental Biophysics, Russian Academy of Sciences,
Institutskaya 3, Pushchino, Moscow Region, 142290 Russia
**Institute of Mathematical Problems of Biology - Branch of Keldysh Institute of Applied Mathematics
, ul. Professora Vitkevicha 1, Pushchino, Moscow Region, 142290 Russia
The paper demonstrates the effectiveness of the use of the cross-correlation Shannon entropy, which is a gen-
eralization of the spectral entropy widely used in scientific research, as a method of the analysis of mutual
correlations between plankton abundance fluctuations and temperature variations. The analysis of time series
characterizing the dynamics of zooplankton, phytoplankton, bacterioplankton and water temperature varia-
tions shows, in particular, that the most significant are the mutual correlations between temperature and bac-
terioplankton.
Keywords: cross-correlation Shannon entropy, plankton, temperature
БИОФИЗИКА том 64
№ 4
2019