БИОФИЗИКА, 2019, том 64, № 4, с. 754-766
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 574.24
СЕЗОННЫЕ КОЛЕБАНИЯ АТМОСФЕРНОГО ДАВЛЕНИЯ,
ПАРЦИАЛЬНОЙ ПЛОТНОСТИ КИСЛОРОДА И ГЕОМАГНИТНОЙ
АКТИВНОСТИ КАК ДОПОЛНИТЕЛЬНЫЕ СИНХРОНИЗАТОРЫ
ЦИРКАННУАЛЬНЫХ РИТМОВ
© 2019 г. Н.В. Кузьменко* **
*Национальный медицинский исследовательский центр имени В.А. Алмазова МЗ РФ,
197341, Санкт-Петербург, ул. Аккуратова, 2
**Первый Санкт-Петербургский государственный медицинский университет имени академика И.П. Павлова МЗ РФ,
197022, Санкт-Петербург, ул. Льва Толстого, 6/8
Е-mail: nat.kuzmencko2011@yandex.ru
Поступила в редакцию 28.02.2019 г.
После доработки 12.04.2019 г.
Принята к публикации 22.04.2019 г.
Для регионов с разными типами климата проанализирована сезонная динамика атмосферного
давления и геомагнитной активности и рассчитаны сезонные колебания парциальной плотности
кислорода. На основе полученных данных предпринята попытка объяснения возможных причин и
механизмов синхронизации цирканнуальных ритмов в стандартных лабораторных условиях. В ходе
анализа собственных и литературных данных сделано заключение, что атмосферное давление и
парциальная плотность кислорода могут быть рассмотрены в качестве дополнительных
синхронизаторов цирканнуальных ритмов организма, поскольку для них характерна четкая
сезонная динамика. Также показано, что существуют механизмы, позволяющие включать эти
метеорологические факторы в процесс синхронизации биологических ритмов.
Ключевые слова: сезон, атмосферное давление, парциальная плотность кислорода, геомагнитная
активность.
DOI: 10.1134/S0006302919040015Х
зонные колебания гормонов щитовидной железы
Годовой цикл жизнедеятельности представля-
ет собой систему сменяющих друг друга физиоло-
и гипофиза [4,5], сезонная динамика плодовито-
сти у крыс [6], сезонные колебания параметров
гических состояний, связанных с приспособле-
нием организма к меняющимся природным усло-
гемодинамики и активности отделов вегетатив-
виям. Сезонная динамика показана в
ной нервной системы у крыс [7], а также сезон-
функционировании нервной, сердечно-сосуди-
ные изменения чувствительности митохондрий
стой, эндокринной, репродуктивной систем. Ос-
клеток печени крыс к Са2+ [8] и сезонные вариа-
новными факторами синхронизации цирканну-
ции в активности Mg2+-зависимой ATФазы в ми-
альных ритмов являются изменения длины дня и
тохондриях клеток сердца крыс [9]. Кроме того,
температуры воздуха в течение года. Стандартные
было установлено, что время года оказывает вли-
лабораторные условия, в которых поддерживает-
яние на моделирование патологий в стандартных
ся постоянный фотопериод, температура и влаж-
лабораторных условиях. Так, в работе [10] на сам-
ность воздуха должны были бы отменить циркан-
ках крыс линии Sprague-Dawley было обнаруже-
нуальные ритмы. Однако в экспериментальных
но, что при введении канцерогена, способствую-
исследованиях, проведенных в стандартных ла-
щего развитию рака молочной железы, опухоль
бораторных условиях, была показана сезонная
развивается у 61% крыс весной и только у 34%
динамика активности N-ацетилтрансферазы у
осенью, несмотря на постоянный световой ре-
цыплят [1], выработки мелатонина у крыс [2], се-
жим и температуру воздуха в помещении содер-
зонные изменения уровня половых гормонов у
жания животных. Аналогичные результаты были
крыс [3,4] и макак резус (Macaca mulatta) [5], се-
получены в экспериментах, описанных в работе
[11]. Также реноваскулярная гипертензия в стан-
Сокращения: МТ-рецепторы - мелатонинергические ре-
дартных лабораторных условиях в модели «2 поч-
цепторы, СХЯ - супрахиазматическое ядро гипоталамуса.
754
СЕЗОННЫЕ КОЛЕБАНИЯ АТМОСФЕРНОГО ДАВЛЕНИЯ
755
ки, 1 зажим» у крыс линии Wistar развивалась ча-
жет быть рассмотрена в качестве синхронизатора
ще весной, чем осенью и зимой [12].
цирканнуальных ритмов.
Атмосферное давление, температура и влаж-
Существование эндогенных «биологических
ность воздуха определяют парциальную плот-
часов», которые представляют собой генетически
ность кислорода в воздухе, для которой также по-
закрепленную периодичность химических реак-
казана сезонная динамика с максимумом зимой и
ций в организме, не исключает наличие экзоген-
минимумом летом [24-26].
ного компонента-синхронизатора
[13]. Стоит
Цель настоящей работы - в регионах с разны-
разобраться, являются ли на самом деле стандарт-
ми типами климата проанализировать сезонную
ные лабораторные условия неизменными в тече-
динамику атмосферного давления и геомагнит-
ние всего года, возможно, существуют метеоро-
ной активности, рассчитать сезонные колебания
логические факторы, подверженные сезонной
парциальной плотности кислорода и на основе
динамике, от воздействия которых нельзя спря-
полученных данных объяснить результаты экспе-
таться в помещении с искусственным освещени-
риментальных работ, в которых была обнаружена
ем и отоплением. Действительно, геомагнитная
сезонная динамика функционирования организ-
активность и атмосферное давление будут по-
ма в стандартных лабораторных условиях.
прежнему воздействовать на организм, находя-
щийся и в условиях стандартного вивария. На се-
годня установлено, что эти факторы оказывают
МАТЕРИАЛЫ И МЕТОДЫ
влияние на функционирование организма
[7,14,15].
Для исследования были отобраны 12 публика-
ций, в которых авторы сообщали о сезонной за-
Некоторые исследователи предполагают, что
висимости результатов экспериментов, постав-
именно сезонные колебания геомагнитной ак-
ленных в стандартных лабораторных условиях
тивности могут синхронизировать цирканнуаль-
(таблица). Исследования [7,12] из этого перечня
ные циклы в стандартных лабораторных условиях
были проведены автором настоящей публикации
[2]. Расчеты показали, что в среднем геомагнит-
на базе НМИЦ им. В.А. Алмазова.
ная активность ниже в период с декабря по фев-
Для регионов, где проводились исследования,
раль, чем в период с марта по октябрь [7,16-18].
для каждого года помесячно (с января по декабрь)
Кроме того, геомагнитные возмущения являются
за период исследований или за десятилетний пе-
более частыми и сильными в период, близкий к
риод (если период исследований не известен или
датам равноденствий. Предполагают, что в мо-
известен, но нет данных в архиве) рассчитывали
мент равноденствий благодаря ориентации дипо-
среднемесячные показатели температуры (в °С) и
ля Земли к плоскости эклиптики максимальное
относительной влажности воздуха (в %); средне-
количество ионизированных частиц от Солнца
месячные величины атмосферного давления
достигают ионосферы [19-21]. Также помимо го-
(данные в мм рт.ст. были переведены в единицы
довой динамики геомагнитная активность изме-
СИ (гПа)) и их стандартное отклонение (по вели-
няется в связи с 11-летними циклами солнечной
чине стандартного отклонения делался вывод о
активности. Это делает геомагнитную активность
среднемесячной вариабельности атмосферного
нестабильным фактором для синхронизации
давления). Были использованы архивные метео-
цирканнуальных циклов. Так, по данным ИЗ-
рологические данные по температуре воздуха и
МИРАН в 2003 г. было зарегистрировано 109 гео-
фактическому атмосферному давлению на уровне
магнитных бурь, а в 2009 г. - всего 7 [22].
по относительной влажности воздуха - с сайтов
Известно, что сезонные колебания атмосфер-
ного давления тесно связаны с сезонными коле-
weatherbase.com.
баниями температуры воздуха. Зимой, когда по-
На основе среднемесячных величин атмо-
верхность суши остывает, создаются благоприят-
сферного давления, относительной влажности и
ные условия для повышенного атмосферного
температуры воздуха для каждого месяца рассчи-
давления. Летом при прогревании поверхности
тывали парциальную плотность кислорода в воз-
земли атмосферное давление понижается. Кроме
того, зимой суша остывает быстрее, чем море. В
духе (ρO2 в г/м3) по формуле, выведенной
связи с этим, зимой между сушей и морем возни-
В.Ф. Овчаровой из уравнения Менделеева-
кает большой перепад атмосферного давления, к
Клайперона [24]:
лету этот контраст уменьшается. В результате зи-
3
10
·0,232(P − e)
мой наблюдается более частое чередование цик-
ρO
=
,
(1)
2
лонов и антициклонов, чем летом [23]. Сезонная
RT
динамика атмосферного давления достаточно
где 0,232 - массовая доля кислорода в воздухе,
стабильна, поскольку взаимосвязана с сезонной
Р - атмосферное давление в Па, е - парциальное
динамикой температуры воздуха, и поэтому мо-
давление водяного пара в Па, R - удельная газо-
БИОФИЗИКА том 64
№ 4
2019
756
КУЗЬМЕНКО
Экспериментальные исследования, в которых была зафиксирована сезонная динамика в функционировании
организма в стандартных лабораторных условиях
Место исследования,
Период проведения
географические координаты,
Краткая информация об исследовании
исследования, публикация
высота над уровнем моря
Санкт-Петербург, Россия
На самцах крыс стока Wistar установлены
2010-2012 г.г.
59°57′ с.ш., 30°19′ в.д., 18 м н/м
сезонные колебания параметров гемодинамики,
Н. В. Кузьменко и др.,
активности вегетативной нервной системы,
2018 [7], 2017 [12]
а также влияние сезона на частоту развития
реноваскулярной гипертензии
Ганновер, Германия
На самцах крыс стока Sprague-Dawley
1977-1978 г.г.
52°22′ с.ш., 9°44′ в.д., 55 м н/м
установлена сезонная динамика тиреотропного
С. С. Wong et al., 1983 [4]
гормона, пролактина, андрогенов,
трийодотиронина, лютеинизирующего гормона
Ганновер, Германия
На самках крыс стока Sprague-Dawley установ-
Период неизвестен
52°22′ с.ш., 9°44′ в.д., 55 м н/м
лено, что частота возникновения рака молочной
W. Loscher et al., 1997 [10]
железы зависит от сезона
Варшава, Польша
На цыплятах показана сезонная динамика
Период неизвестен
52°13′ с.ш., 21°02′ в.д., 103 м н/м
активности N-ацетилтрансферазы
P. Majewski et al., 2005 [1]
Мюнстер, Германия
На самцах макак-резус (Macaca mulatta)
период неизвестен (4 года)
51°57′ с.ш., 7°37′ в.д., 60 м н/м
показана сезонная динамика половых
E. J. Wickings et al., 1980 [5]
гормонов и гормонов гипофиза
Лодзь, Польша
На самцах крыс стока Wistar показаны сезонные
Период неизвестен
51°47′ с.ш., 19°28′ в.д., 224 м н/м
изменения чувствительности митохондрий
M. Labieniec-Watala et al.,
клеток печени крыс к Са2+
2012 [8]
Кошице, Словакия
На самках крыс стока Sprague-Dawley
Период неизвестен
48°43′ с.ш., 21° в.д., 208 м н/м
установлено, что частота возникновения рака
P. Kubatka et al., 2002 [11]
молочной железы зависит от сезона
Тюбинген, Германия
На самках крыс стока Sprague-Dawley
1997-2006 гг.
48°31′ с.ш., 9° в.д., 341 м н/м
установлена сезонная динамика мелатонина
H. Bartsch et al., 2012 [2]
Братислава, Словакия
На самцах крыс стока Wistar показаны сезонные
Период неизвестен
48° с.ш., 17° в.д., 140 м н/м
вариации в активности Mg2+-зависимой ATФазы
J. Mujkosová et al., 2008 [9]
в митохондриях клеток сердца
Овьедо, Испания
На самках крыс стока Wistar установлена
Осень 2000 г. - зима 2002 г.
43°21′ с.ш., 5°50′ з.д., 232 м н/м
сезонная динамика гонадотропных гормонов
N. Vázquez et al., 2007 [3]
и пролактина
Чикаго, США
На самках крыс стока Sprague-Dawley
Период неизвестен (2 года)
41°54′ с.ш., 87°39′ з.д., 179 м н/м
установлена сезонная динамика плодовитости
T. M. Lee et al., 1986 [6]
БИОФИЗИКА том 64
№ 4
2019
СЕЗОННЫЕ КОЛЕБАНИЯ АТМОСФЕРНОГО ДАВЛЕНИЯ
757
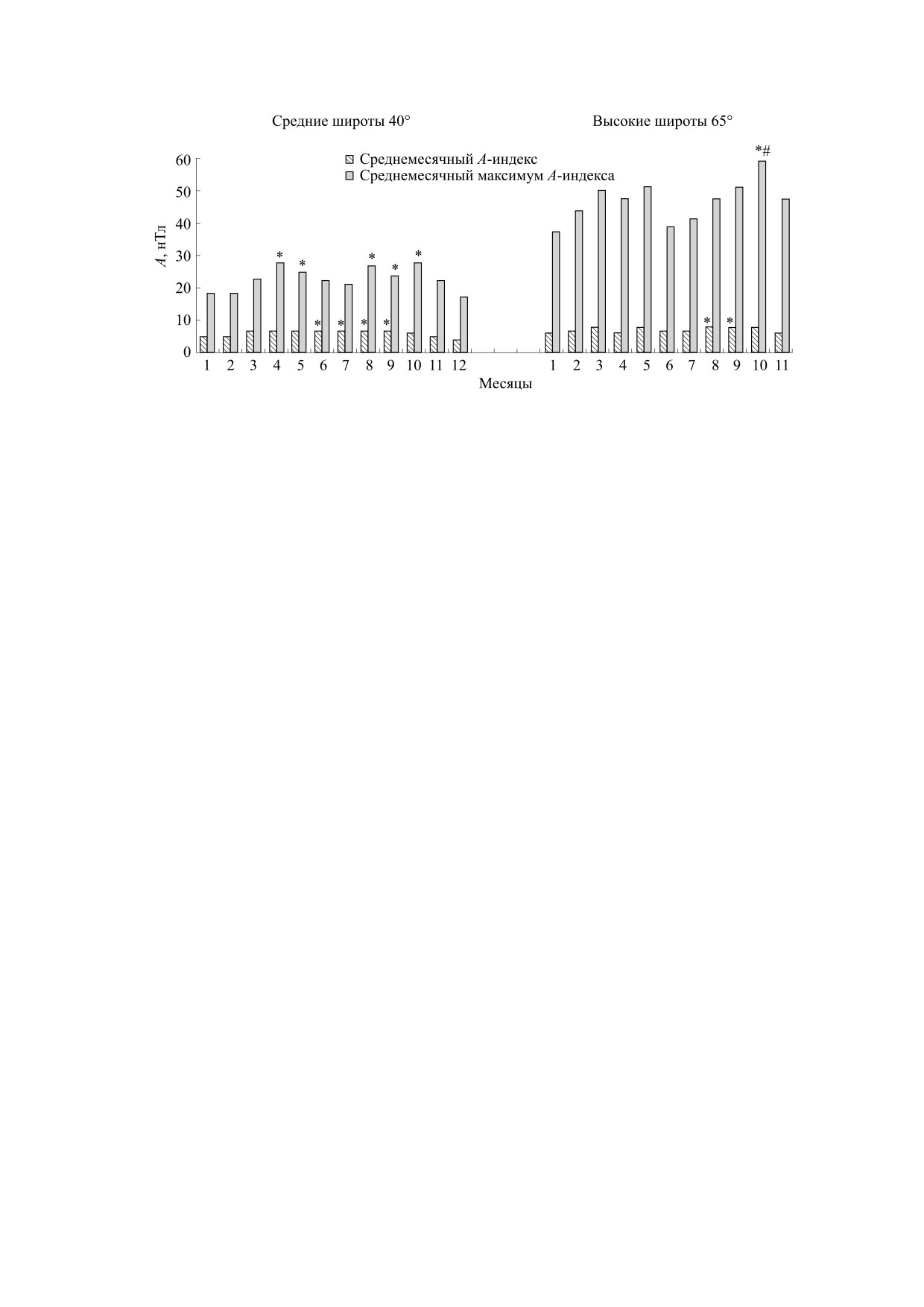
Рис. 1. Сезонные колебания геомагнитной активности для средних и высоких широт (статистика за период 1997-
2016 гг.): * - p < 0,05 - значимость различий относительно зимних месяцев, # - p < 0,05 - значимость различий отно-
сительно июня. А - ежедневный локальный индекс геомагнитной активности.
вая постоянная сухого воздуха, равная
логической организации [29]. Данные по атмо-
287 Дж/(кг · К), Т - температура воздуха в К.
сферному давлению и ρO2 в графиках представле-
Парциальное давление водяного пара рассчи-
ны в виде средних значений. Данные по геомаг-
тывали по формуле:
нитной активности в графиках представлены в
виде медиан, т.к. для этого показателя было уста-
ϕP
e
=
v,
(2)
новлено распределение, отличное от нормально-
100
го (тест Коломогорова-Смирнова). Cравнение
где φ - относительная влажность воздуха в %, Pv -
среднемесячных А-индексов осуществляли с по-
давление насыщенного водяного пара при дан-
мощью U-критерия Манна-Уитни. Статистиче-
ной среднемесячной температуре воздуха в Па
ски значимыми считали различия данных при
[27].
р < 0,05.
В работе [28] было показано вполне удовлетво-
рительное соответствие (в пределах 3 г/м3, или
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
около 1%) результатов измерения ρO2 посред-
Сезонная динамика геомагнитной активности,
ством газоанализаторов и результатов, рассчи-
атмосферного давления и парциальной плотности
танных по формуле В.Ф. Овчаровой.
кислорода. В ходе проведенных исследований бы-
Проводился анализ сезонной динамики сред-
ло подтверждено, что помимо температуры воз-
немесячной величины и вариабельности атмо-
духа и длины дня сезонной динамике подвержена
сферного давления, а также парциальной плотно-
геомагнитная активность, атмосферное давление
сти кислорода в наружном воздухе.
и парциальная плотность кислорода в наружном
Для исследования сезонной динамики геомаг-
воздухе.
нитной активности были взяты архивные данные
Расчеты показали, что геомагнитная актив-
Ар-индексов геомагнитной активности с сайта
ность в среднем ниже в декабре-феврале, чем в
Space Weather Prediction Center (ftp://ftp.swpc.
период с марта по октябрь, а геомагнитные воз-
noaa.gov/pub/indices/old_indices/). Были рассчи-
мущения максимальны вблизи дат равноден-
таны среднемесячные значения и среднемесяч-
ствий. Причем сезонная динамика геомагнитной
ные максимумы локальных ежедневных А-индек-
активности лучше выражена в средних широтах,
сов для средних и высоких широт за период 1997-
чем в высоких (рис. 1), что согласуется с расчета-
2016 гг. Кроме того, были рассчитаны медианы
ми других исследователей [19-21]. Анализ сезон-
Ар-индексов за период 1998-2009 гг., а также за
ной динамики Ар-индексов в годы с высокой и
годы с высокой (2003-2005 гг.) и с низкой (2008,
низкой солнечной активностью показал, что в го-
2009 гг.) геомагнитной активностью.
ды с высокой солнечной активностью может
Статистическую обработку данных по метео-
происходить смещение ежегодного максимума
рологическим параметрам проводили согласно
Ар-индекса, как, например, случилось в 2004 и
«Наставлению по глобальной системе обработки
2005 гг., тогда как в годы с низкой геомагнитной
данных и прогнозирования» Всемирной метеоро-
активностью (2008 и 2009 гг.) сохранялась стан-
БИОФИЗИКА том 64
№ 4
2019
758
КУЗЬМЕНКО
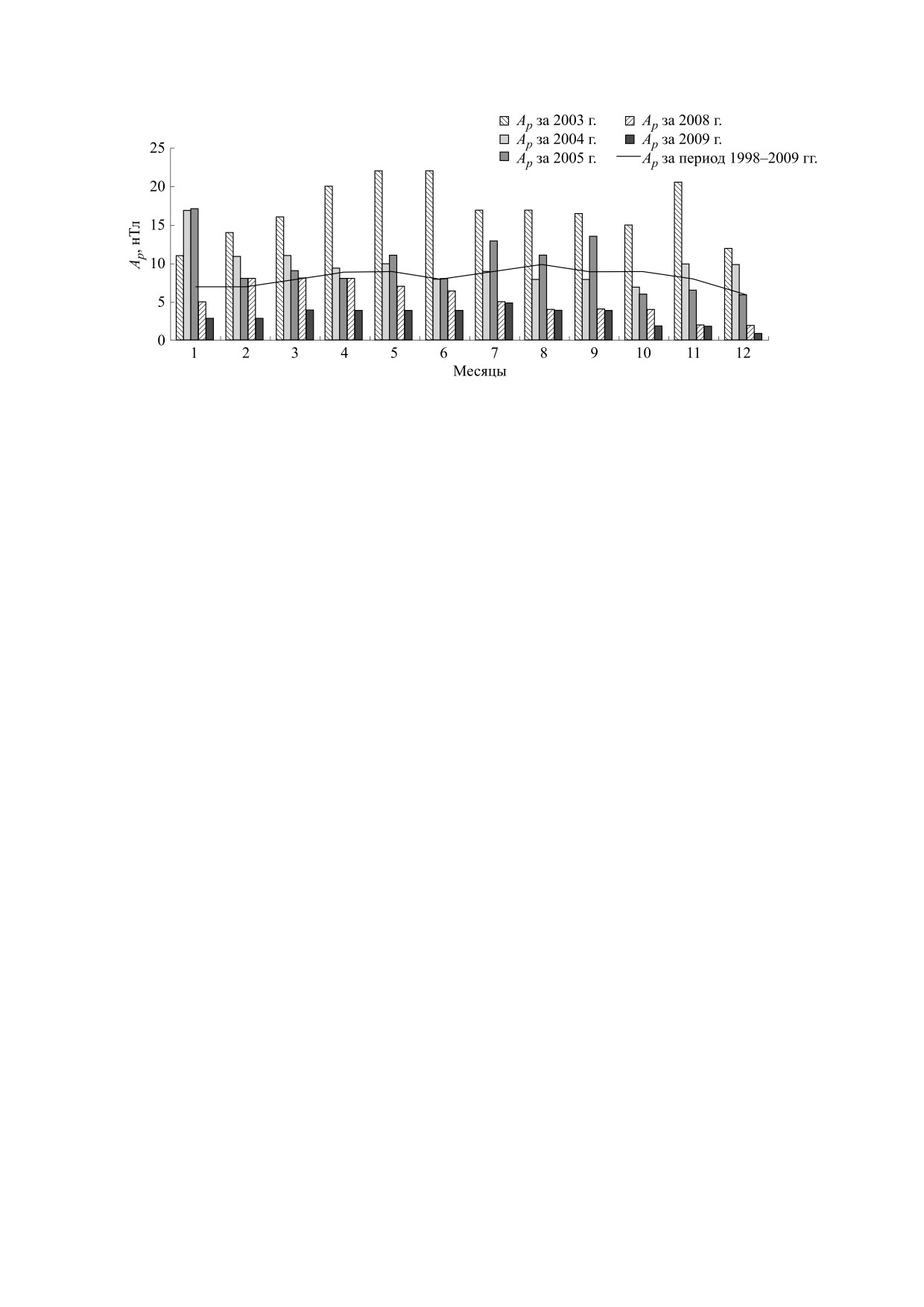
Рис. 2. Сезонные колебания геомагнитной активности в годы с высокой и низкой солнечной активностью. Ар -
ежедневный индекс геомагнитной активности.
дартная сезонная динамика Ар-индекса (рис. 2). В
зонная динамика вариабельности атмосферного
итоге существенные геомагнитные возмущения
давления выражена слабо или не выражена [26].
являются скорее не синхронизаторами, а десин-
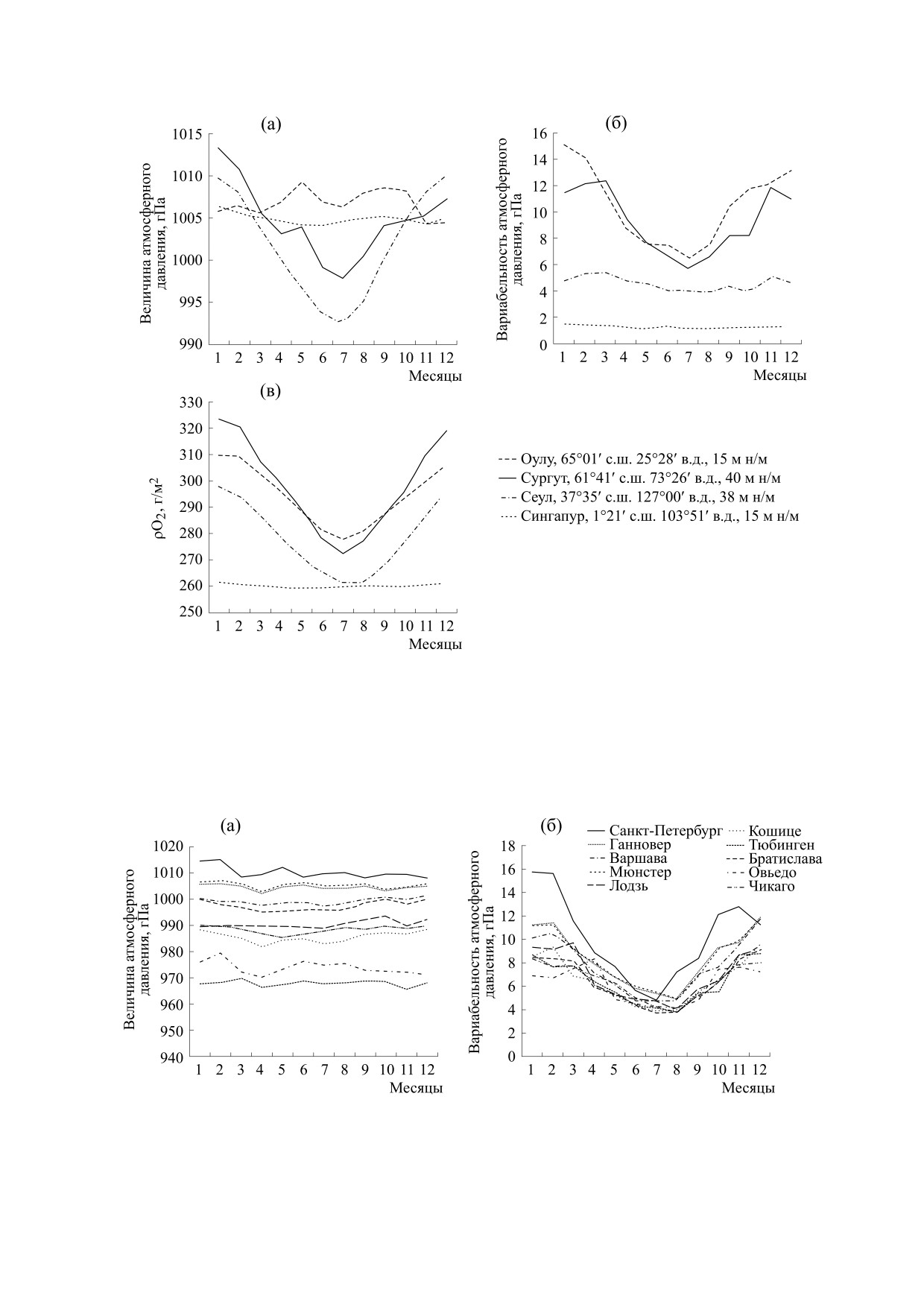
Подобный климат наблюдается, например, в Се-
хронизаторами биоритмов. Роль высокой солнеч-
уле (Южная Корея) (рис. 3). К третьей группе от-
носятся области, где имеет место четко выражен-
ной активности в десинхронизации биологиче-
ских ритмов отмечали и другие авторы [30]. Одна-
ная сезонная динамика как величины, так и вари-
ко поскольку многими исследованиями
абельности атмосферного давления. Такой
климат, например, наблюдается в г. Сургут (Хан-
подтверждается способность геомагнитных воз-
ты-Мансийский автономный округ, Россия)
мущений влиять на функционирование организ-
ма [15], то, вероятно, и этот фактор может изме-
(рис. 3). Четвертый тип климата, в котором сезон-
ная динамика величины и вариабельности атмо-
нить результаты экспериментов, поставленных в
сферного давления не выражена, характерен для
стандартных лабораторных условиях.
регионов, расположенных вблизи экватора, на-
Некоторые исследователи пытались объяс-
пример для Сингапура (рис. 3). Для всех типов
нить цирканнуальные ритмы, зарегистрирован-
климата, кроме экваториального, характерна чет-
ные в стандартных лабораторных условиях, коле-
кая сезонная динамика парциальной плотности
баниями атмосферного давления, но пришли к
кислорода с максимум зимой и минимумом летом
выводу, что атмосферное давление не имеет вы-
(рис. 3). Аналогичная сезонная динамика ρO2 по-
раженной сезонной динамики [6]. Однако про-
казана и другими авторами [25,28]. Ранее были
стые расчеты среднемесячных значений и стан-
опубликованы графики сезонной динамики ве-
дартных отклонений атмосферного давления в
личины и вариабельности атмосферного давле-
регионах с разными типами климата показывают,
ния, а также парциальной плотности кислорода в
что эти исследователи ошибались. По характеру
21 регионе земного шара с различными типами
сезонной динамики атмосферного давления ре-
климата (от субарктического до тропического)
гионы можно разделить на четыре группы. К пер-
[26].
вой группе относятся регионы, в которых годовая
динамика среднемесячной величины атмосфер-
Результаты экспериментов, поставленных в
ного давления четко не выражена, но наблюдает-
стандартных лабораторных условиях. Дополни-
ся хорошо выраженная сезонная динамика вари-
тельные синхронизаторы сезонных ритмов. Все ре-
абельности атмосферного давления, которая уве-
гионы, где зафиксирована сезонная динамика в
личивается от лета к зиме с годовым минимумом
функционировании организма в стандартных ла-
в летние месяцы и с годовым максимумом в де-
бораторных условиях (таблица), по характеру се-
зонной динамики атмосферного давления отно-
кабре-марте [26]. Такой тип климата характерен,
например, для Оулу (Финляндия) (рис. 3; н/м -
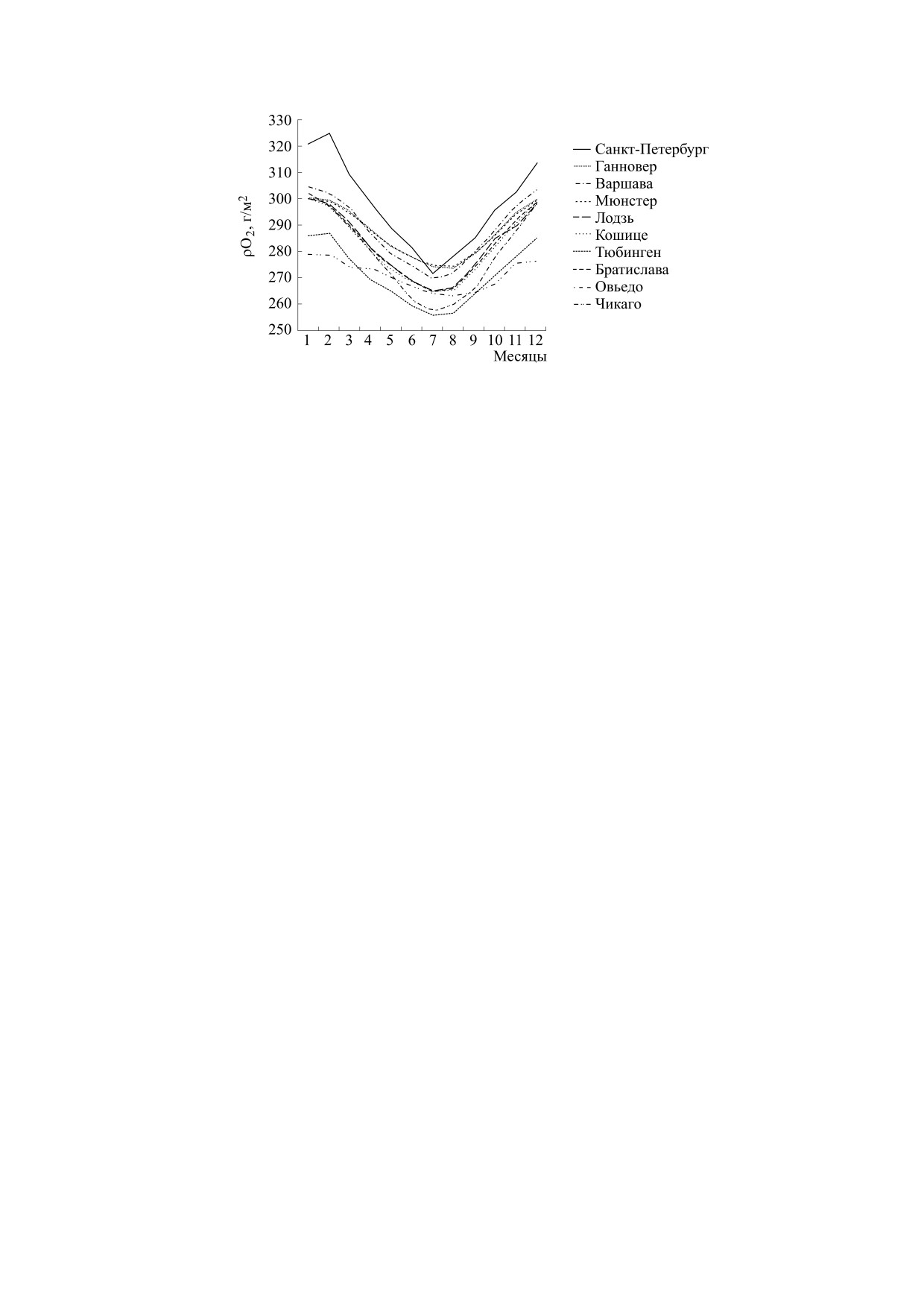
сятся к первой группе (рис. 4). Также в этих горо-
над уровнем моря). Ко второй группе относятся
дах имеет место четко выраженная сезонная
динамика парциальной плотности кислорода с
регионы, в которых четко выражена сезонная ди-
намика среднемесячной величины атмосферного
амплитудой колебаний «зима-лето» - 23 ± 7 г/м3
давления с годовым минимумом в летние месяцы
(рис. 5). В лабораториях, расположенных в обла-
и с годовым максимумом в зимние. При этом се-
стях с подобным типом климата, синхронизиру-
БИОФИЗИКА том 64
№ 4
2019
СЕЗОННЫЕ КОЛЕБАНИЯ АТМОСФЕРНОГО ДАВЛЕНИЯ
759
Рис. 3. Варианты сезонной динамики атмосферного давления и парциальной плотности кислорода (статистика за
период 2007-2016 гг.).
Рис. 4. Сезонная динамика атмосферного давления в регионах, где были зафиксированы цирканнуальные колебания
в функционировании организма в стандартных лабораторных условиях.
БИОФИЗИКА том 64
№ 4
2019
760
КУЗЬМЕНКО
Рис. 5. Сезонная динамика парциальной плотности кислорода в регионах, где были зафиксированы цирканнуальные
колебания в функционировании организма в стандартных лабораторных условиях.
ющими факторами сезонных ритмов могут вы-
эти причины и объясняют более высокие значе-
ступать сезонная динамика вариабельности атмо-
ния артериального давления (127 мм рт. ст. по
сферного давления, а также сезонные изменения
сравнению с 107 мм рт. ст.) и маркеров активно-
качества воздуха, поступающего в лабораторию
сти симпатической нервной системы в спектре
при вентилировании.
вариабельности сердечного ритма (НЧ/ВЧ 0,28
по сравнению с 0,19), которые наблюдались в
В исследованиях по моделированию гипертен-
стандартных лабораторных условиях у нормотен-
зии «2 почки, 1 зажим», проведенных в Санкт-
зивных животных в зимний период по сравнению
Петербурге, было показано, что реноваскулярная
с летним [7].
гипертензия развивается чаще всего весной (в
Кроме того, о влиянии сезонных изменений
56% случаев) и реже всего зимой (только в 20%
ρO2 на организмы свидетельствует сезонная ди-
случаев) [12]. Скорее всего, это объясняется тем,
намика в функционировании митохондрий, на-
что зимой наблюдается ежегодный максимум
блюдавшаяся в городах Братислава [9] и Лодзь [8].
ρO2, который мог нивелировать влияние ишемии
В исследованиях в Братиславе была показана по-
почечной артерии. Также сезонная динамика ρO2
вышенная активность Mg2+-зависимой ATФазы
объясняет сезонные изменения параметров гемо-
в митохондриях клеток сердца крыс зимой и вес-
динамики в стандартных лабораторных условиях
ной в стандартных лабораторных условиях. Это
[7], которые через хеморецепторы реагируют на
согласуется с результатами другой работы, в кото-
газовый состав вдыхаемого воздуха. Известно,
рых наблюдали увеличение ATФазной активно-
что гипоксия и гипероксия оказывают влияние на
сти в митохондриях легких крысы в условиях ги-
функционирование сердечно-сосудистой систе-
пероксии [38].
мы [31-33]. Обнаружено угнетающее действие
кислорода на эндотелиальные вазодилататорные
В ряде исследований в стандартных лабора-
факторы (оксид азота и простагландины) [34] и
торных условиях была установлена сезонная ди-
его стимулирующее действие на выделение ка-
намика в функционировании репродуктивной и
техоламинов надпочечниками [35]. Кроме того,
эндокринной систем, характерная для естествен-
зимой атмосферное давление более вариабельно,
ных условий [3-6]. В экспериментах на крысах
а исследования показали, что колебания атмо-
наблюдали увеличение пролактина и андрогенов
сферного давления от 755-756 мм рт. ст. (1004-
весной, тироксина - летом, фолликулостимули-
1005,5 гПа) как в сторону увеличения, так и в сто-
рующего гормона - летом и осенью [3,4], сниже-
рону уменьшения способствуют подъему артери-
ние плодовитости зимой [5]. Сезонная динамика
ального давления у крыс [7]. К тому же зимы в
репродуктивной функции может оказывать влия-
Санкт-Петербурге в период проведения этих экс-
ние на результаты экспериментов по моделирова-
периментов сопровождались высокой среднеме-
нию гормонозависимых патологий, например,
сячной величиной атмосферного давления. Из-
рака молочной железы [10,11]. Кроме того, были
вестно, что атмосферное давление, помимо влия-
показаны сезонные колебания в продукции мела-
ния на ρO2 воздуха согласно формуле
(1),
тонина в стандартных лабораторных условиях [2].
определяет давление всех жидкостей в организме
Возникает закономерный вопрос: «Могут ли се-
и, следовательно, может вызвать изменение
зонные колебания ρO2 и атмосферного давления
трансмурального давления сосудов [36,37]. Все
оказывать влияние на синхронизацию функцио-
БИОФИЗИКА том 64
№ 4
2019
СЕЗОННЫЕ КОЛЕБАНИЯ АТМОСФЕРНОГО ДАВЛЕНИЯ
761
нирования всего организма, и какие для этого су-
интактными МТ1-рецепторами - сохранялось
ществуют механизмы»?
[51]. Это связывают с разной локализацией МТ1-
и МТ2-рецепторов в головном мозге. Так, в pars
Мелатонин и сезонная динамика атмосферного
tuberalis гипофиза были обнаружены МТ1-рецеп-
давления и парциальной плотности кислорода. Ос-
торы, но не МТ2-рецепторы [52]. Тиреоидэкто-
новным синхронизатором суточных и сезонных
мия (удаление щитовидной железы) блокировала
ритмов является гормон мелатонин, синтезирую-
сезонные репродуктивные циклы у овец [53], так-
щейся пинеальной (шишковидной) железой.
же как и делеция рецепторов, чувствительных к
Взаимосвязь мелатонина и тиреотропного гор-
тиреотропному гормону гипофиза, блокировала
мон гипофиза, обеспечивая переключение генов
переключение DIO2/DIO3 у мышей при введе-
DIO2/DIO3 (активирующего/деактивирующего
нии экзогенного мелатонина [50]. В итоге, хотя
гормоны щитовидной железы), модулирует се-
литературные данные противоречивы, наличие
зонные ритмы в репродуктивном поведении и
сезонной динамики мелатонина и функциониру-
метаболизме позвоночных
[39]. Несмотря на
ющей щитовидной железы, скорее всего, необхо-
большую роль мелатонина в биологических рит-
димы для сохранения цирканнуальных ритмов,
мах, в опытах на мышах с делецией мелатонинер-
по крайней мере, репродуктивной функции.
гических рецепторов (МТ-рецепторов) было по-
казано, что интактная мелатонинергическая си-
Мелатонин синтезируется у млекопитающих
из серотонина под воздействием фермента N-аце-
стема не требуется для генерации суточной
тилтрансферазы, активность которого повышает-
ритмичности у мышей [40]. Также у людей, в ред-
ся в темное время суток под воздействием стиму-
ких случаях отсутствия ритмической секреции
ляции пинеальных адренергических рецепторов
мелатонина пинеальной железой циркадный
цикл сна/бодрствования сохраняется [41]. В опы-
норадреналином [54,55]. В опытах на хомяках
(Cricetus cricetus) было показано повышение син-
тах на крысах симпатэктомия (эктомия шейного
теза мелатонина после инъекций агонистов β-ад-
симпатического ганглия и 6-гидроксидофамино-
ренергических рецепторов [56]. К тому же в экс-
вая симпатэктомия) отменяла циркадные колеба-
периментах на мышах наблюдали выраженную
ния мелатонина [42] и артериального давления, а
также уменьшала суточные флуктуации сердеч-
экспрессию часовых генов после введения адре-
налина, норадреналина или агонистов адренер-
ного ритма и локомоторной активности [42,43].
гических рецепторов. Считается, что эти эффек-
Кроме того, было отмечено отсутствие сезонных
ты вызывает цАМФ-зависимый транскрипцион-
ритмов репродуктивной функции у диких кроли-
ный фактор CREB, который может оказывать
ков после симпатэктомии [44] и у крыс после пи-
неалэктомии [45]. В противоположность этому,
влияние на экспрессию часовых генов Per1 и Per2
[57]. В ряде исследований сообщается о сезонной
симпатэктомия не отменяла сезонный пик пине-
динамике концентрации катехоламинов с макси-
ального пептида у крыс [46]. Также реакцию мас-
мумом зимой [58,59]. Кроме того, обнаружено
сы тела и репродуктивной функции на изменение
стимулирующее действие кислорода и атмосфер-
фотопериода наблюдали у хомяков (Cricetus crice-
tus) после удаления шишковидной железы [47].
ного давления на выделение адреналина и норад-
реналина надпочечниками [35], из чего следует,
Пинеалэктомия не изменяла репродуктивные
что сезонная динамика синтеза катехоламинов,
циклы у волков в естественной среде обитания,
возможно, будет сохраняться и в стандартных ла-
из чего был сделан вывод, что для синхронизации
бораторных условиях. Можно предположить, что
сезонного размножения существуют и другие
сигналы кроме сезонной динамики мелатонина
катехоламины, модулируя экспрессию часовых
генов и синтез мелатонина, способны синхрони-
[48]. Однако следует учесть, что помимо шишко-
зировать биологические ритмы при фиксирован-
видной железы, мелатонин может синтезировать-
ном фотопериоде. Это подтверждает и сезонная
ся экстропинеально (кожей, сетчаткой глаза, ти-
динамика активности N-ацетилтрансферазы в
мусом, селезенкой, сердцем, мышцами, печенью,
желудком, поджелудочной железой, кишечни-
шишковидной железе цыплят, показанная в
стандартных лабораторных условиях [1].
ком, плацентой, яичками, яичником, костным
мозгом, корой головного мозга и стриатумом), к
Однако даже при фиксированной длине дня не
тому же мелатонин легко проникает через гемато-
стоит полностью пренебрегать ролью фоторецеп-
энцефалический барьер [49]. Кроме того, у нока-
торов в сезонной динамике мелатонина. Так, ис-
утных мышей с генетическим дефицитом мелато-
следования показали, что внутриглазное давле-
нина экспрессия генов DIO2, DIO3 не реагирова-
ние подвержено сезонным колебаниям с макси-
ла на изменение фотопериода [50]. В экспе-
мумом зимой и минимумом летом
[60], в
риментах при введении экзогенного мелатонина
результате чего, возможно, изменится чувстви-
мышам с делецией мелатонинергических рецеп-
тельность фоторецепторов сетчатки, что может
торов 1 типа (МТ1) и интактными МТ2-рецепто-
оказать влияние на синтез мелатонина. Рассуж-
ров переключение DIO2/DIO3 генов полностью
дая о факторах, которые могут модулировать
отсутствовало, а при делеции МТ2-рецепторов с
внутриглазное давление, авторы перечисляют
БИОФИЗИКА том 64
№ 4
2019
762
КУЗЬМЕНКО
солнечный свет, температуру воздуха и даже ко-
го и темного времени суток, которое через СХЯ
лебания геомагнитной активности [60]. Однако
посредством транскрипционной/трансляцион-
атмосферное давление, которое определяет дав-
ной обратной связи, состоящей из часовых генов
ление всех жидкостей в организме, в первую оче-
и их белковых продуктов, регулирует выработку
редь участвует в изменении внутриглазного дав-
мелатонина и синхронизирует работу всего орга-
ления. Действительно, исследования показали,
низма [70,71]. Кроме того, ритмическая экспрес-
что колебания именно атмосферного давления, а
сия часовых генов показана во всех органах [71].
не температуры воздуха оказывают влияние на
Основными часовыми генами являются Per1,
внутриглазное давление [61,62]. Интересно отме-
Per2, Cry1 и Cry2, которые контролируются тран-
тить, что амплитуда сезонных колебаний внут-
скрипционными факторами CLOCK, NPAS2 (па-
риглазного давления зависит от типа климата
ралогичен CLOCK) и BMAL1 [72]. Поскольку
[63]. Кроме того, установлена прямая корреляция
CLOCK, NPAS2 и BMAL1 представляют собой
между атмосферным давлением и внутричереп-
транскрипционные факторы с доменом PER-
ным давлением, что напрямую может воздейство-
ARNT-SIM (PAS), воздействие внешних стиму-
вать на функционирование различных структур
лов включает часовые гены. Установлено, что до-
головного мозга, в том числе, и связанных с био-
мен PAS проявляет чувствительность не только к
логическими ритмами [64].
свету, но также к изменениям окислительно-вос-
становительного потенциала, колебаниям кисло-
Несмотря на то что есть механизмы, которые
рода, лигандов и общего энергетического уровня
могут вызвать сезонные колебания мелатонина и
[57,73]. Это делает теоретически возможным син-
при фиксированном фотопериоде, вероятно, от-
хронизацию биологических ритмов посредством
сутствие сезонной динамики длины дня приведет
атмосферного давления и парциальной плотно-
к уменьшению выраженности цирканнуального
сти кислорода. На сегодня известно, что такой
профиля мелатонина.
фактор, как температура воздуха, может легко
Часовые гены и сезонная динамика атмосферно-
увлечь экспрессию часовых генов практически во
го давления и парциальной плотности кислорода.
всех тканях организма, кроме СХЯ, в котором
Известно, что основным регулятором циркадных
экспрессия часовых генов сохранит свой суточ-
ритмов у млекопитающих является супрахиазма-
ный профиль даже при существенных колебани-
тическое ядро гипоталамуса (СХЯ). У других по-
ях температуры [74].
звоночных основной осциллятор (пинеалоцит)
К настоящему времени известно об изменении
также расположен в гипоталамусе, причем у всех
экспрессии часовых генов в течение суток, в то же
позвоночных (кроме млекопитающих) пинеало-
время участие часовых генов в сезонных ритмах
цит содержит фоторецепторы, циркадные часы и
изучено в меньшей степени. Исследования экс-
синтезирует мелатонин [39]. У млекопитающих
прессии часовых генов в течение года, проведен-
циркадные часы находятся в СХЯ, а мелатонин
ные на японской лесной мыши (Apodemus Spe-
синтезируется пинеальной (шишковидной) же-
ciosus), показали существенное снижение экс-
лезой [39]. В СХЯ млекопитающих показана вы-
прессии BMAL1 и Cry1 в переходный период
сокая плотность МТ1-рецепторов, при этом
между репродуктивным и нерепродуктивным се-
МТ2-рецепторы не обнаружены [52]. Поврежде-
зонами [75]. Сезонная ритмичность часовых ге-
ние СХЯ нарушает циркадный профиль мелато-
нов также была установлена у рыб [76] и у мигри-
нина [42], а также устраняет эффект фотопериода
рующих птиц [77]. Кроме того, было обнаружено,
на нейрональные и сердечно-сосудистые реак-
что генетические вариации в ARNTL(BMAL1) и
ции [65]. Результаты исследований по значению
NPAS2 ассоциированы с выраженностью сезон-
СХЯ в сезонных ритмах репродуктивной функ-
ных колебаний пищевого и социального поведе-
ции противоречивы. Так, у пятнистых скунсов с
ния у людей [78]. В ряде экспериментальных ра-
поврежденными СХЯ отсутствовало изменение
бот наблюдали зависимость экспрессии часовых
размера семенников на естественную сезонную
генов от фотопериода [79-82]. Так, при длинном
динамику фотопериода [66]. В экспериментах на
дне отмечалась повышенная экспрессия Per1 в
хомяках после двустороннего разрушения СХЯ не
момент акрофазы [80,81]. Ген Per1 также ассоци-
было зарегистрировано изменения массы тела и
ирован с повышением уровня альдостерона и
гонад на введение экзогенного мелатонина и на
удержанием ионов Na+. У мышей, нокаутных по
длину дня [67,68]. В противоположность этому,
этому гену, наблюдается низкий уровень альдо-
исследования на норке показали, что поврежде-
стерона и артериального давления [83]. Рост ча-
ние СХЯ не оказывает влияния на ответ репро-
стоты развития реноваскулярной гипертензии,
дуктивной функции на изменение фотопериода
который наблюдался весной в стандартных лабо-
при интактной пинеальной железе или инфузии
раторных условиях [7], также может быть связан с
экзогенного мелатонина [69].
весенним увеличением экспрессии Per1, если до-
Самым сильным сигналом для синхронизации
пустить возможность синхронизацию цирканну-
биологических часов служит чередование светло-
альных ритмов функционирования всего орга-
БИОФИЗИКА том 64
№ 4
2019
СЕЗОННЫЕ КОЛЕБАНИЯ АТМОСФЕРНОГО ДАВЛЕНИЯ
763
низма посредством сезонной динамики атмо-
температуры воздуха в среднем на 10°С вызыва-
сферного давления и парциальной плотности
ют, соответственно, уменьшение и увеличение
кислорода. Исследования на цыплятах показали,
уровня HIF-1α в крови [94]. В связи с этими фак-
что содержание в условиях постоянного освеще-
тами можно предположить, что HIF-α участвует в
ния не отменяет сезонных колебаний суточных
синхронизации цирканнуальных циклов в ответ
профилей часовых генов, но изменяет их по срав-
на сезонные колебания ρO2 и атмосферного дав-
нению с животными, содержащимися при фик-
ления. Действительно, в экспериментах наблю-
сированном коротком или длинном дне [82]. В
дали изменение экспрессии часовых генов при
опытах на мышах было показано, что дефицит од-
увеличении HIF-1α в ответ на гипоксию [95,96].
ного из часовых транскрипционных факторов
Также установлено, что стабилизация HIF-1α
(CLOCK, NPAS2) вызывает тонкие циркадные
может увеличивать экспрессию DIO3 и снижать
дефекты, а повреждение двух транскрипционных
продукцию гормонов щитовидной железы [97].
факторов уже приводят к существенным наруше-
ниям в ритмичности, до полной аритмичности
Роль дополнительных синхронизаторов циркан-
при возмущениях фотопериода [72]. Мутация
нуальных ритмов в лабораторных и естественных
BMAL1 приводит к полной потере циркадного
условиях. Многочисленные эксперименты с мо-
ритма [84].
делированием фотопериода, проведенные на жи-
вотных, у которых репродуктивная активность
Есть предположения, что в процессах пере-
зависит от длины дня, показали, что именно фо-
ключения фаз сезонных циклов большую роль
топериод является главным синхронизатором се-
играет обратимое метилирование промоторных
зонных ритмов [39,66-69,85]. Однако атмосфер-
участков генов, связанных с биологическими
ное давление и парциальная плотность кислорода
ритмами. В исследованиях на хомяках было пока-
также могут быть рассмотрены в качестве допол-
зано, что зимой в результате ингибирования ме-
нительных синхронизаторов цирканнуальных
латонином ДНК-метилтрансферазы в гипотала-
ритмов организма, поскольку для них характерна
мусе уменьшается метилирование гена DIO3, что
четкая сезонная динамика, а также существуют
повышает его экспрессию, обеспечивая регрес-
механизмы, позволяющие включать эти метеоро-
сию гонад [85]. Также в экспериментах на мышах
логические факторы в процесс синхронизации
была обнаружена роль микроРНК-132/212 в сбро-
биологических ритмов. Тем не менее из-за отсут-
се часов в СХЯ [86]. Процессы метилирования
ствия колебаний длины дня, температуры возду-
ДНК могут регулироваться не только колебания-
ха, количества и качества кормов сезонная дина-
ми мелатонина. Сегодня уже установлено влия-
мика функционирования организма в лаборатор-
ние кислорода на активность ДНК-метилтранс-
ных условиях будет существенно отличаться от
феразы и экспрессию микроРНК [87-90].
той, что наблюдается в естественной среде обита-
Организм позвоночных чутко реагирует на со-
ния. Известно, что в лабораторных условиях
держание кислорода во вдыхаемом воздухе.
нельзя вызвать оцепенение у гибернирующих в
Практически каждая клетка может активировать
природе млекопитающих без изменения фотопе-
эволюционно консервативный путь гипоксиче-
риода и понижения температуры, а также предва-
ского ответа, регулируемый гипоксически-инду-
рительного накопления животными жира [98,99].
цируем фактором (HIF-1), который является чле-
Также, в лабораторных условиях у диких свиней
ном семейства транскрипционных факторов с
(Tayassu tajacu) и крыс отмечалось увеличение ча-
PAS. HIF-1 контролирует экспрессию большин-
стоты сердечных сокращений зимой по сравне-
ства генов, участвующих в адаптации к гипокси-
нию с летом [7,100]. В противоположность этому,
ческим условиям. HIF-1 представляет собой гете-
в естественных условиях (или в условиях близких
родимер, состоящий из кислородо-лабильных
к естественным) установлено снижение частоты
HIF-1α и конститутивно-экспрессируемых HIF-
сердечных сокращений зимой у копытных и у
1β-субъединиц. Кислородозависимая регуляция
кроликов [101-104]. Это явление принято связы-
HIF-1α является многоступенчатым процессом,
вать со снижением метаболизма, которое вызы-
который включает деградацию при нормоксии и
вают низкая амбиентная температура и скудная
стабилизацию с активацией транскрипционной
кормовая база [102]. При этом эксперименты на
функции в гипоксических условиях [91,92]. HIF-
кастрированных оленях (Cervus elaphus) показали,
2α паралогичен HIF-1α. Стабилизация HIF-α
что ежедневное введение мелатонина отменяет
происходит, когда ρO2 в ткани органа становится
сезонную динамику сердечного ритма даже при
ниже 5%, в то же время при нормоксии в боль-
естественных флуктуациях длины дня и темпера-
шинстве тканей ρO2 находится в пределах от 3 до
туры воздуха [105].
10% [93]. Из этого следует, что HIF-α достаточно
В естественных условиях атмосферное давле-
чувствителен к изменениям ρO2 воздуха, а также
ние и парциальная плотность кислорода как до-
к факторам, которые вызывают сдвиг ρO2. В опы-
полнительные синхронизаторы цирканнуальных
тах на мышах установлено, что падение и подъем
ритмов, вероятно, играют роль в модуляции се-
БИОФИЗИКА том 64
№ 4
2019
764
КУЗЬМЕНКО
зонных реакций у животных, впадающих в спяч-
ровании организмов, помещенных в стандартные
ку. В экспериментах на ехидне (Tachyglossus acu-
лабораторные условия. Это может быть причиной
leatus) в фазе гибернации было установлено, что
различий в результатах экспериментов, постав-
гипоксия и гипероксия существенно влияют на
ленных в разные сезоны.
картину дыхания [106]. В исследованиях на жи-
5. Высказывается предположение, что в син-
вотных, впадающих в зимнюю спячку, наблюда-
хронизацию цирканнуальных ритмов посред-
ли увеличение кислородной сатурации гемогло-
ством атмосферного давления и парциальной
бина в период гибернации [107,108]. Известно,
плотности кислорода могут быть вовлечены меха-
что сезонная спячка прерывается эпизодами
низмы обратимого метилирования DIO2/DIO3 и
кратковременного возбуждения и нормализации
часовых генов, транскрипционные факторы
температуры тела. До сих пор непонятно, какие
CLOCK (NPAS2), BMAL1 и HIF-α, имеющие до-
внешние или внутренние стимулы вызывают эти
мен PAS, а также катехоламины, способные вы-
периоды возбуждения, поскольку они наблюда-
зывать экспрессию часовых генов и воздействую-
ются и при моделировании гибернации в лабора-
щие на синтез мелатонина через модулирование
торных условиях с поддержанием постоянной
активности N-ацетилтрансферазы. Кроме того,
низкой амбиентной температуры [109].
нельзя исключить влияние на синтез мелатонина
Также дополнительные факторы синхрониза-
изменений внутриглазного и внутричерепного
ции сезонных ритмов могут быть важны для со-
давления, происходящих под действием колеба-
временного человека, изменяющего микрокли-
ний атмосферного давления.
мат помещений с помощью искусственного осве-
щения, отопления или кондиционирования. В
СПИСОК ЛИТЕРАТУРЫ
исследованиях на людях с использованием пуль-
совой оксиметрии авторы работы [14] отмечали
1.
P. Majewski, I. Adamska, J. Pawlak, et al., J. Pineal
прямую корреляцию между среднесуточным зна-
Res. 39 (1), 66 (2005).
чением атмосферного давления и кислородной
2.
H. Bartsch, D. Mecke, H. Probst et al. Chronobiol Int.
сатурацией гемоглобина. В клиническом иссле-
29 (8), 1048 (2012).
довании [110] индекс апноэ у взрослых пациентов
3.
N. Vázquez, E. Díaz, C. Fernández, et al., Physiol Res.
56 (1), 79 (2007).
имел сезонную динамику, при этом обратно кор-
4.
C. C. Wong, K. D. Dehler, M. J. Atkinson, et al., J.
релировал с температурой воздуха и прямо корре-
Endocr. 97, 179 (1983).
лировал с величиной атмосферного давления и
5.
E. J. Wickings and E. Nieschlag, Int. J. Androl. 3 (1), 87
относительной влажности воздуха. Мета-анализ
(1980).
клинических исследований сезонной динамики
6.
T. M. Lee and M. K. McClintock, J. Reprod. Fertil. 7
артериального давления в регионах с разным ти-
(1), 51(1986).
пом климата показал, что выраженность сезон-
7.
Н. В. Кузьменко, Н. С. Рубанова, М. Г. Плисс и
ных колебаний артериального давления у людей
В. А. Цырлин, Рос. физиол. журн. 104 (4), 477
сильнее в регионах с сезонной динамикой сред-
(2018).
немесячной величины атмосферного давления,
8.
M. Labieniec-Watala and K. Siewiera, CellBio. 2 (1),
чем в регионах, где имеет место сезонная динами-
20 (2013).
ка вариабельности атмосферного давления [26].
9.
J. Mujkosová, M. Ferko, P. Humeník, et al., Physiol.
Res. 57 (2),75 (2008).
10.
W. Loscher, M. Mevissen, and M. Häussler, Pharma-
ВЫВОДЫ
col. Toxicol. 81 (6), 265 (1997).
11.
P. Kubatka, E. Ahlersová, I. Ahlers, et al., Physiol. Res.
1. Геомагнитная активность, подчиненная
51 (6), 633(2002).
одиннадцатилетним циклам, вероятно, не может
12.
Н. В. Кузьменко, М. Г. Плисс и В. А. Цырлин,
рассматриваться в качестве фактора, синхрони-
Артериальная гипертензия 23 (6), 561 (2017).
зирующего цирканнуальные ритмы в стандарт-
13.
С. Н. Ежов, Вестн. ТГЭУ 2, 104 (2008).
ных лабораторных условиях.
14.
C. A. Pope, D. W. Dockery, R. E. Kanner, et al., Am. J.
Respir. Crit. Care Med. 159 (2), 365 (1999).
2. Атмосферное давление имеет четкую сезон-
15.
S. J. Palmer, M. J. Rycroft, and M. Cermack, Surv.
ную динамику, но характер ее зависит от особенно-
Geophys. 27 (5), 557 (2006).
стей климата региона. Вблизи экватора сезонная
16.
A. Clua de Gonzalez, V. M. Silbergleit, W. D. Gonza-
динамика атмосферного давления отсутствует.
lez, and B. T. Tsurutani, J. Atmos. Sol. Terr. Phys. 63
3. Парциальная плотность кислорода имеет чет-
(4), 367 (2001).
кую сезонную динамику с зимним максимумом и
17.
Н. М. Руднева, Е. А. Гинзбург, Л. А. Дремухина и
А. А. Нусинов, Гелиогеофизические исследования
летним минимумом в регионах с любым типом кли-
13, 97 (2005).
мата, за исключением экваториального.
18.
W. Lyatsky and A. Tan, J. Geophys. Res. 108 (A5), 12
4. Сезонная динамика атмосферного давления
(2003).
и парциальной плотности кислорода могут вы-
19.
E. W. Cliver, Y. Kamide, and A. G. Ling, J. Atmospher.
звать цирканнуальные колебания в функциони-
Solar-Terrestr. Physics 64 (1), 47 (2002).
БИОФИЗИКА том 64
№ 4
2019
СЕЗОННЫЕ КОЛЕБАНИЯ АТМОСФЕРНОГО ДАВЛЕНИЯ
765
20. D. H. McIntosh, Phil. Trans. Roy. Soc. Lond. 251
51. S. Yasuo, T. Yoshimura, S. Ebihara, and H. W. Kolf, J.
(1001), 525 (1959).
Neurosci. 29 (9), 2885 (2009).
21. B. R. Boller and R. Stolov, J. Geophys. Res. 75 (1), 92
52. B. Lacoste, D. Angeloni, S. Dominguez-Lopez, et al.,
(1970).
J. Pineal Res. 58 (4), 397(2015).
22. В. Н. Обридко, Х. Д. Канониди, Т. А. Митрофа-
53. 53. S. M. Moenter, C. J. Woodfill, and F. J. Karsch,
нова и Б. Д. Шельтинг, Геомагнетизм и аэрономия
Endocrinology 128 (3), 1337 (1991).
53 (2), 157 (2013).
54. D. C. Klein, D. Sugden, and J. L. Weller, Proc. Natl.
23. J. F. Petersen, D. Sack, and R. E. Gabler, Physical Ge-
Acad. Sci. USA 80 (2), 599 (1983).
ography (Cengage Learning, 2016).
55. R. J. Reiter, Endocr. Rev. 12 (2), 151 (1991).
24. В. Ф. Овчарова, Вопросы курортологии, физиоте-
56. M. L. Garidou, B. Vivien-Roels, P. Pevet, et al., Am. J.
рапии и лечебной физкультуры 2, 29 (1981).
Physiol. Regul. Integr. Comp. Physiol. 284 (4), R1043
25. В. Ф. Овчарова, в сб. Труды междунар. симпозиума
(2003).
ВМО/ВОЗ/ЮНЕП СССР «Климат и здоровье чело-
57. Y. Tahara and S. Shibata, Free Radic. Biol. Med. 119,
века» (Гидрометеоиздат, Ленинград, 1988).
129 (2018).
26. Н. В. Кузьменко, М. Г. Плисс и В. А. Цырлин, Рос.
58. T. Tsuchihashi, K. Uezono, I. Abe, et al., Hypertens.
кардиол. журн. 24 (1), 80 (2019).
Res. 18 (3), 209 (1995).
27. А. А. Васильев, В. Е. Федоров и Л. Д. Храмов, Фи-
59. H. J. Kruse, I. Wieczorek, H. Hecker, et al., J. Labor.
зика: учебное пособие для СПО, 2-е изд., испр. и доп.
Clin. Med. 140 (4), 236 (2002).
(Юрайт, М., 2018).
60. J. Cheng, M. Xiao, H. Xu, et al., Medicine (Baltimore)
28. A. S. Ginzburg, A. A. Vinogradova, and E. I. Fedorova,
95 (31), e4453 (2016).
Izvestiya, Atmospheric and Oceanic Physics 50 (8), 782
61. S. Van de Veire, P. Germonpre, C. Renier, et al., Invest.
(2014).
Ophthalmol. Vis. Sci. 49 (12), 5392 (2008).
29.
World Meteorological Organization, Manual on the global
62. O. Brinchmann-Hansen and K. Myhre, Aviat. Space
data-processing and forecasting system. V. 1. Global As-
Environ. Med. 60 (10, Pt 1), 970 (1989).
pects (Geneva, 2010).
63. S. K. Gardiner, S. Demirel, M. O. Gordon, and M.
30.
Ф. И. Комаров, С. И. Рапопорт, Т. К. Бреус и
A. Kass, Ophthalmology 120 (4), 724 (2013).
С. М. Чибисов, Клинич. медицина
95
(6),
502
64. L. Herbowski, Int. J. Biometeorol. 61 (1), 181 (2017).
(2017).
65. T. Mutoh, S. Shibata, H. W. Korf, and H. Okamura, J.
31.
R. J. Julian, Poult. Sci. 86 (5),1006 (2007).
Physiol. 547 (Pt 1), 317 (2003).
32.
R. van der Bel, M. Çalişkan, R. A. van Hulst, et al.,
66. M. Berria, M. DeSantis, and R. A. Mead, J. Exp. Zool.
Front. Physiol. 30 (8), 186 (2017).
255 (1), 72 (1990).
33.
L. E. Stuhr, G. W. Bergø, and I. Tyssebotn, Aviat.
67. T. J. Bartness, B. D. Goldman, and E. L. Bittman, Am.
Space Environ. Med. 65 (6), 531 (1994).
J. Physiol. 260 (1, Pt 2), R102 (1991).
34.
F. Yamazaki, K. Takahara, R. Sone, et al., J. Appl.
68. G. A. Eskes, M. Wilkinson, W. H. Moger, and B. Ru-
Physiol. 103 (6), 2026 (2007).
sak, Biol. Reprod. 30 (5),1073 (1984).
35.
T. Nakada, H. Koike, T. Katayama, et al., Hinyokika
69. D. L. Maurel, M. M. Ben Saad, G. Roch, and P. Siaud,
Kiyo 30 (10), 1357 (1984).
J. Pineal Res. 32 (1), 15 (2002).
36.
N. V. Mel’nikov, V. I. Poliakov, S. G. Krivoshchekov,
70. Э. Б. Арушанян и А. В. Попов, Успехи физиол.
et al., Aviakosm. Ekolog. Med. 48 (3), 51 (2014).
наук 42 (4), 39 (2011).
37.
P. A. Doriot, J. Theor. Biol. 221 (1), 133 (2003).
71. И. В. Курбатова, Л. В. Топчиева и Н. Н. Немов,
38.
Z. L. Harris, K. M. Ridge, B. Gonzalez-Flecha, et al.,
Труды Карельского науч. центра РАН 5, 3 (2014).
Eur. Respir. J. 9 (3), 472 (1996).
72. J. P. Debruyne, J. Genet. 87 (5), 437 (2008).
39.
Y. Nakane and T. Yoshimura, Front. Neurosci. 8, 115
73. B. L. Taylor and I. B. Zhulin, Microbiol. Mol. Biol.
(2014).
Rev. 63 (2), 479 (1999).
40.
M. Pfeffer, H. W. Korf, and H. Wicht, Int. J. Mol. Sci.
18 (3), Е530 (2017).
74. E. D. Buhr, S. H. Yoo, and J. S.Takahashi, Science 330
41.
J. Lipton, J. T. Megerian, S. V. Kothare, et al., Neurol-
(6002), 379 (2010).
ogy 73 (4), 323 (2009).
75. M. Akiyama, S. Takino, Y. Sugano, et al., J. Biol. Reg-
42.
S. Perreau-Lenz, A. Kalsbeek, M. L. Garidou, et al.,
ul. Homeost. Agents 29 (3), 589 (2015).
Eur. J. Neurosci. 17 (2), 221 (2003).
76. 76. M. J. Herrero and J. M. Lepesant, Gen. Comp. En-
43.
S. A. Briaud, B. L. Zhang, and F. Sannajust, J. Cardio-
docrinol. 208, 30 (2014).
vasc. Pharmacol. Ther. 9 (2), 97(2004).
77. 77. D. Singh, A. K. Trivedi, S. Rani, et al., FASEB J. 29
44. M. M. Ben Saad and D. Maurel, Reproduction 121(2),
(10), 4248 (2015).
323 (2001).
78. L. Kovanen, S. T. Saarikoski, A. Aromaa, et al., PLoS
45.
X. Y. Liu, Y. T. Xu, Q. Shi, et al., Endocrinol. Lett. 34
One 5 (4), e10007 (2010).
(2), 143 (2013).
79. G. Lincoln, S. Messager, H. Andersson and D. Ha-
46.
M. M. Prechel, T. K. Audhya, R. Swenson, et al., Life
zlerigg, Proc. Natl. Acad. Sci. USA 99 (21), 13890
Sci. 44 (2), 103 (1989).
(2002).
47.
S. Monecke, D. Sage-Ciocca, F. Wollnik, and P. Pévet,
80. A. Sumová, M. Sládek, M. Jác, and H. Illnerová, Brain
J. Biol. Rhythms 28 (4), 278 (2013).
Res. 947 (2), 260 (2002).
48.
C. S. Asa, U. S. Seal, M. Letellier, et al., Biol. Reprod.
81. S. Messager, D. G. Hazlerigg, J. G. Mercer, and
37 (1), 14 (1987).
P. J. Morgan, Eur. J. Neurosci. 12 (8), 2865 (2000).
49.
D. Acuña-Castroviejo, G. Escames, C. Venegas, et al.,
82. E. Turkowska, P. M. Majewski, S. Rai, and K. Skwarlo-
Cell Mol. Life Sci. 71 (16), 2997 (2014).
Sonta, Chronobiol. Int. 31 (1), 134 (2014).
50.
H. Ono, Y. Hoshino, S. Yasuo, et al., Proc. Natl. Acad.
83. J. Richards, K. Y. Cheng, S. All, et al., Am. J. Physiol.
Sci. USA 105 (47),18238 (2008).
Renal. Physiol. 305 (12), F1697 (2013).
БИОФИЗИКА том 64
№ 4
2019
766
КУЗЬМЕНКО
84. M. K. Bunger, L. D. Wilsbacher, S. M. Moran, et al.,
98. M. Trefna, M. Goris, C. M. Thissen, et al., J. Comp.
Cell 103 (7), 1009 (2000).
Physiol. B 187(5-6), 725 (2017).
85. T. J. Stevenson and B. J. Prendergast, Proc. Natl. Acad.
99. M. Kart Gür, R. Refinetti, and H. Gür, J. Comp.
Sci. USA 110 (41), 16651 (2013).
Physiol. B 179 (2), 155 (2009).
86. L. Mendoza-Viveros, C. K. Chiang, J. K.Ong, et al.,
100.S. M. Zervanos, Comp. Biochem. Physiol. A. Comp
Cell Rep. 19 (3),505 (2017).
Physiol. 50 (2), 365 (1975).
87. J. Nanduri, Y. J. Peng, N. Wang, et al., J. Physiol. 596
101. W. Arnold, T. Ruf, L. E. Loe, et al., Sci. Rep. 8 (1),
(15), 3087 (2018).
14466 (2018).
88. M. R. Alivand, Z. S. Soheili, M. Pornour, et al., J. Cell
Biochem. 118 (10), 3193 (2017).
102.C. Turbill, T. Ruf, T. Mang, and W. Arnold, J. Exp.
Biol. 214 (Pt 6), 963 (2011).
89. Q. Yang, X. Wu, J. Sun, et al., Mol. Neurobiol. 53 (1),
436 (2016).
103.N. M. Baldock, R. M. Sibly, and P. D. Penning, Ani-
mal Behaviour 36 (1), 35(1988).
90. L. Xiong, F. Wang, X. Huang, et al., FEBS J. 279 (23),
4318 (2012).
104.K. Eisermann, B. Meier, M. Khaschei, and D. von
91. Chavez, L. F. Miranda, P. Pichiule, and J. C. Chavez,
Holst, Physiol. Behav. 54 (5), 973 (1993).
Ann. NY Acad. Sci. 1147, 312 (2008).
105.B. M. Domingue, P. R. Wilson, D. W. Dellow, and
92. U. Berchner-Pfannschmidt, S. Frede, C. Wotzlaw, and
T. N. Barry, Br. J. Nutr. 68 (1), 77 (1992).
J. Fandrey, Eur. Respir J. 32 (1), 210 (2008).
106.S. Nicol and N. A. Andersen, Comp. Biochem.
93. A. Carreau, B. El Hafny-Rahbi, A. Matejuk, et al., J.
Physiol. A. Mol. Integr. Physiol. 136 (4), 917 (2003).
Cell Mol. Med. 15 (6), 1239 (2011).
107. I. G. Revsbech and A. Fago, J. Comp. Physiol. B 187
94. S. Zhang, Z. Kuang, and X. Zhang, Int. J. Environ.
(5-6), 847 (2017).
Res. Public Health 12 (6), 5743 (2015).
108.L. A. Maginniss and W. K. Milsom, Respir. Physiol. 95
95. C. Yu, S. L. Yang, X. Fang, et al., Mol. Med. Rep. 11
(2), 195 (1994).
(5), 4002 (2015).
109.T. Ikeno, C. T. Williams, C. L. Buck, et al., J. Biol.
96. M. Egg, L. Köblitz, J. Hirayama, et al., Chronobiol.
Int. 30 (4), 510 (2013).
Rhythms 32 (3), 246 (2017).
97. W. S. Simonides, M. A. Mulcahey, E. M. Redout, et al.,
110. C. M. Cassol, D. Martinez, F. A. Boeira Sabino da Sil-
J. Clin. Invest. 118 (3), 975 (2008).
va, et al., Chest. 42 (6), 1499 (2012).
Seasonal Variations in Atmospheric Pressure, Partial Oxygen Density and Geomagnetic
Activity as Additional Synchronizers of Circannual Rhythms
N.V. Kuzmenko* **
*Almazov National Medical Research Centre, ul. Akkuratova 2, St. Petersburg, 197341 Russia
** Pavlov First St. Petersburg State Medical University, ul. Lva Tolstogo 6/8, St. Petersburg, 197022 Russia
The seasonal dynamics of atmospheric pressure and geomagnetic activity in regions with different types of
climate were analyzed to calculate the seasonal variations in the partial density of oxygen. On the basis of the
data obtained, the attempt was undertaken to explain the possible causes and mechanisms of synchronization
of circannual rhythms in standard laboratory conditions. In the analysis of our own and literature data, it was
concluded that atmospheric pressure and partial oxygen density can be considered as additional synchroniz-
ers of circannual rhythms of the body, since they are characterized by clear seasonal dynamics. It has also been
shown that there are mechanisms that allow these meteorological factors to be included in the process of syn-
chronizing biological rhythms.
Keywords: season, atmospheric pressure, partial oxygen density, geomagnetic activity
БИОФИЗИКА том 64
№ 4
2019