БИОФИЗИКА, 2019, том 64, № 5, с. 854-860
МОЛЕКУЛЯРНАЯ БИОФИЗИКА
УДК 577.353
ТРЕХСУТОЧНАЯ ФУНКЦИОНАЛЬНАЯ РАЗГРУЗКА СОПРОВОЖДАЕТСЯ
УВЕЛИЧЕНИЕМ ЭКСПРЕССИИ ГЕНА TTN В m. soleus КРЫСЫ
ПРИ ОТСУТСТВИИ ИЗМЕНЕНИЙ В АЛЬТЕРНАТИВНОМ СПЛАЙСИНГЕ
В ДИАПАЗОНЕ ЭКЗОНОВ С 50-ГО ПО 111-Й
© 2019 г. А.Д. Уланова*, Ю.В. Грицына*, В.К. Жалимов**, Л.Г. Бобылёва*, С.П. Белова***,
Т.Л. Немировская***, Б.С. Шенкман***, И.М. Вихлянцев* ****
*Инcтитут теоретической и экспериментальной биофизики РАН,
142290, Пущино Московской области, Институтская ул., 3
**Институт биофизики клетки РАН - обособленное подразделение ФИЦ ПНЦБИ РАН,
142290, Пущино Московской области, Институтская ул., 3
***ГНЦ РФ - Институт медико-биологических проблем РАН, 123007, Москва, Хорошевское шоссе, 76а
****Пущинcкий государственный естественно-научный институт,
142290, Пущино Московской области, просп. Науки, 3
E-mail: ivanvikhlyantsev@gmail.com
Поступила в редакцию 22.05.2019 г.
После доработки 11.07.2019 г.
Принята к публикации 12.07.2019 г.
Исследованы изменения экспрессии гена титина и альтернативный сплайсинг пре-мРНК титина в
диапазоне экзонов с 50-го по 111-й в m. soleus крыс после трехсуточной функциональной разгрузки.
Методом ПЦР с обратной транскрипцией в режиме реального времени обнаружено увеличение в
1,81 раза (p ≤ 0,01, n = 6) экспрессии гена титина в m. soleus крыс группы функциональной разгрузки
в сравнении с контрольной группой (n = 7). Показано наличие всех экзонов мРНК титина в
исследуемом диапазоне в m. soleus крыс двух исследуемых групп. Таким образом, трехсуточная
функциональная разгрузка сопровождается увеличением экспрессии гена титина в m. soleus крысы
при отсутствии изменений альтернативного сплайсинга в диапазоне экзонов с 50-го по 111-й.
Ключевые слова: функциональная разгрузка, ген титина, альтернативный сплайсинг, m. soleus.
DOI: 10.1134/S000630291905003Х
диске саркомера некоторые участки титиновой
Функциональная разгрузка мышц приводит к
молекулы могут взаимодействовать с актиновы-
развитию атрофии и снижению мышечного тону-
са [1]. При этом мышца теряет эластичность, че-
ми (тонкими) нитями [10], однако большая часть
му во многом может способствовать снижение
его молекулы в этой зоне проходит свободно, со-
содержания гигантского эластичного белка тити-
единяя концы миозиновых нитей с Z-диском.
на, повышенный протеолиз которого происходит
Расположение титина во всех зонах саркомера,
в условиях функциональной разгрузки [2-6].
эластические свойства и взаимодействие со мно-
Титин (тайтин/коннектин) - самый большой
гими белками создают основу для полифункцио-
из известных на сегодняшний день белков. В сар-
нальности этого белка. Показано, что титин
комерах сердечной и скелетных мышц позвоноч-
необходим для сборки миозиновых нитей и сар-
ных титин является третьим по количеству (после
комеров [11]; участвует в поддержании высоко-
актина и миозина) белком. Его молекулы длиной
упорядоченной саркомерной структуры
[12];
около 1 мкм и диаметром 3-4 нм [7] перекрывают
участвует в регуляции актин-миозинового взаи-
половину саркомера от М-зоны до Z-диска, фор-
модействия и вносит вклад в развитие мышечно-
мируя третий тип нитей [8], получивших назва-
го сокращения [10]. Предполагается, что титин
ние эластичных. В А-зоне саркомера титин свя-
играет роль сенсора растяжения и напряжения и в
зан с миозиновыми (толстыми) нитями [9]. В I-
комплексе с сигнальными белками участвует в
процессах внутриклеточной сигнализации, в
Сокращения: TTN - ген титина, ТЦМ - тяжелые цепи
частности, в регуляции экспрессии генов мышеч-
миозина, ОТ-ПЦР - полимеразная цепная реакция с
обратной транскрипцией.
ных белков и белкового обмена в саркомере
854
ТРЕХСУТОЧНАЯ ФУНКЦИОНАЛЬНАЯ РАЗГРУЗКА
855
[10,13-15]. Таким образом, титин является инте-
лярной массой 3,7 МДа [18,22-24]. В других ске-
ресным объектом для изучения при различных
летных мышцах, состоящих преимущественно из
функциональных состояниях, в том числе при
волокон, содержащих «быстрые» изоформы II
функциональной разгрузке.
ТЦМ, экспрессируются N2A-изоварианты тити-
на с более короткой последовательностью из
Показано, что развитие мышечной атрофии в
условиях моделируемой гравитационной разгруз-
Ig-доменов (молекулярная масса 3,3-3,6 МДа)
[18,22-24].
ки в течение семи и более суток сопровождается
повышенным кальций-зависимым протеолизом
Показано, что одним из ключевых событий,
молекул интактного титина, уменьшением его
происходящих в разгруженной мышце, является
содержания и увеличением содержания его про-
изменение экспрессии генов ТЦМ, сопровожда-
теолитических фрагментов [2-5,16]. После трех-
ющееся изменением содержания их мРНК
суточной функциональной разгрузки при нали-
[25,26]. Эти изменения наблюдаются уже после
чии некой тенденции к снижению содержания
первых суток функциональной разгрузки.
нативного титина достоверных изменений его со-
В нашей работе исследована динамика экс-
держания не обнаружено [17]. Исследования из-
прессии гена титина, а также состав экзонов в
менений экспрессии гена титина (TTN) при этом
диапазоне от 50-го до 111-го экзона в m. soleus
не проводились. Однако известны данные об уве-
крысы после трехсуточной функциональной раз-
личении экспрессии гена TTN в сердечной и трех
грузки.
скелетных мышцах мышей после тридцатисуточ-
ного космического полета [6]. Эти результаты
представляют несомненный интерес, однако сле-
МАТЕРИАЛЫ И МЕТОДЫ
дует понимать, что помимо функциональной раз-
Моделирование функциональной разгрузки. В
грузки вклад в повышение экспрессии гена TTN
эксперименте использовали самцов крыс линии
могли внести другие факторы, в частности, такие
Wistar массой 200-220 г. Животные были разделе-
как ионизирующая радиация в условиях косми-
ны на две группы - контрольная группа («С»,
ческого полета и двенадцатичасовое послеполет-
n = 7) и группа трехсуточного вывешивания
ное пребывание животных в условиях земной
(«HS», n = 6). Моделирование функциональной
гравитации. Таким образом, вопросы о том, про-
разгрузки проводили по стандартной методике
исходят ли изменения экспрессии гена TTN в
Ильина-Новикова в модификации Morey-
условиях функциональной разгрузки, когда они
Holton [27,28]. После завершения эксперимента
начинаются и могут ли наряду с повышенным
животных взвешивали и выводили из экспери-
протеолизом титина вносить вклад в снижение
мента путем эвтаназии сверхдозой авертина
содержания этого белка, остаются открытыми.
(5 мл/кг). Камбаловидную мышцу изолировали,
Поэтому актуальность исследования экспрессии
взвешивали, замораживали в жидком азоте и хра-
гена TTN в условиях моделируемой гравитацион-
нили при -85°С. Все процедуры с участием жи-
ной разгрузки не вызывает сомнений. Понима-
вотных соответствовали этическим стандартам
ние молекулярных механизмов протеолиза и син-
учреждения для таких исследований.
теза титина при функциональной разгрузке
мышц важно для разработки средств/методов
Выделение мРНК и проведение ПЦР с обратной
профилактики мышечной атрофии.
транскрипцией и ПЦР с обратной транскрипцией в
реальном времени. Суммарную РНК экстрагиро-
Ген титина у млекопитающих содержит
вали из 4-6 мг замороженной m. soleus при помо-
364 экзона, из которых 363 являются кодирую-
щи набора RNeasyMicroKit (QIAGEN, Германия)
щими [18-20] и способны потенциально кодиро-
по протоколу изготовителя. Определение кон-
вать белок с молекулярной массой 4,2 МДа, в со-
центрации РНК проводили на спектрофотометре
став которого могут входить: 152 иммуноглобу-
UV 2450 (SHIMADZU, Япония). Показания сни-
лин-подобных домена (Ig), 132 фибронектин III-
мали в диапазоне от 200 до 320 нм. Пробу перед
подобных домена (FnIII), протеинкиназный до-
измерением разводили в 21 раз в TE-буфере
мен, 31 домен PEVK, 7 Z-повторов и 33 неиденти-
(10 мМ трис, 1 мМ ЭДТА, pH 8,0). Измерение
фицированных домена (по базе UniProt). Альтер-
каждой пробы проводили не менее трех раз в кю-
нативный сплайсинг пре-мРНК титина является
вете для микрообъемов. Чистоту образцов оцени-
основой разнообразия изоформ этого белка. Мо-
вали исходя из соотношений показателей погло-
лекулярные массы известных изоформ титина со-
щения при различных длинах волн. Соотноше-
ставляют ~ 3,0-3,9 МДа в поперечно-полосатых
ние А260/А230 экспериментальных образцов
мышцах и ~ 2 МДа в гладких мышцах млекопита-
было > 2,0, это указывало на то, что они являлись
ющих и человека [18-21]. Обнаружено, что в ске-
достаточно чистыми от углеводов, пептидов, фе-
летной мышце m. soleus, состоящей преимуще-
нолов или ароматических соединений [29].
ственно из волокон, содержащих «медленную»
изоформу I тяжелых цепей миозина (ТЦМ), экс-
Для проведения ПЦР с обратной транскрип-
прессируется вариант N2A-изоформы c молеку-
цией (ОТ-ПЦР) использовали компоненты фир-
БИОФИЗИКА том 64
№ 5
2019
856
УЛАНОВА и др.
Таблица 1. Пары праймеров для проведения ОТ-ПЦР в режиме реального времени
Праймер
Нуклеотидная последовательность
Размер продукта, п.н.
TTN F
CAGCAGCCAAGAAAGCCGCT
71
TTN R
CACCACTCTGATACTCTGAGGCTCTG
GAPDH F
GCAAGAGAGAGGCCCTCAG
74
GAPDH R
TGTGAGGGAGATGCTCAGTG
Примечание. F - прямой праймер, R - обратный праймер.
мы «Синтол» (Россия). Для синтеза кДНК вод-
Режим ПЦР был следующим:
1)
«горячий
ный раствор, содержащий 1 мкг суммарной РНК,
старт» - 95°С, 5 мин; 2) денатурация - 92°С, 10 с;
30 мкМ случайных гексануклеотидов и 17,4 мкМ
3) отжиг праймеров - 60°С, 30 с; 4) элонгация -
олиго-d(T)15, инкубировали в течение трех минут
68°С, 1 мин. Этапы 2-4 повторяли 30 раз. Изме-
при 70°С и немедленно переносили на лед. Далее
нение экспрессии генов рассчитывали по методу
к смеси добавляли 11,5 мкл мастер-микса (1,3 мМ
2-ΔΔСt cогласно [30]. Значения ΔΔСt рассчитыва-
dNTP,
0,02 ед./мкл ингибитора РНКазы,
лись по формуле ΔΔСt = ΔСt (контроль) - ΔСt
6 ед./мкл M-MLV-ревертазы, 4 мкл 5×-буфера
(опыт), каждое значение ΔСt рассчитывали по
для M-MLV-ревертазы). После этого пробы по-
формуле ΔСt = Сt (ген белка) - Сt (референсный
мещали в амплификатор (iQ5 Multicolor Real-
Time PCRDetectionSystem, Bio-Rad Laboratories,
ген).
США) для проведения обратной транскрипции:
При исследовании изменений альтернативно-
10 мин при 25°С, 60 мин при 37°С, 5 мин при
го сплайсинга (проверки наличия или отсутствия
95°С, 30 мин при 4°С.
экзонов в диапазоне от 50-го до 111-го экзона
включительно) проводили ПЦР с праймерами
Полученную кДНК использовали для прове-
[31], синтезированными фирмой «Евроген» (Рос-
дения ПЦР в реальном времени с праймерами,
сия) (табл. 2). Проведение ПЦР с данными прай-
специфичными к генам исследуемых белков (ти-
мерами позволяет выявлять наличие или отсут-
тину и GAPDH - референсному гену) (табл. 1).
ствие в мРНК титина экзонов (в диапазоне от
50-го до 111-го), кодирующих самый вариабель-
Подбор праймеров проводили с помощью
ный участок молекулы скелетного титина в I-зоне
программы Vector NTI Advance 11. Праймеры бы-
саркомера. ПЦР проводили на амплификаторе
ли синтезированы фирмой «Евроген» (Россия).
SimplyAmp c использованием готовой смеси для
ОТ-ПЦР в реальном времени проводили на ам-
ПЦР qPCRmix-HS SYBR («Евроген», Россия).
плификаторе ДТ-322 («ДНК-Технология», Рос-
Режим ПЦР был следующим: 1) денатурация -
сия) c использованием ДНК полимеразы Tersus
95°С, 8 с; 2) отжиг праймеров - 57°С, 12 с;
(«Евроген», Россия) и SYBR Green I (Invitrogen,
3) элонгация - 72°С, 80 с. Этапы 1-3 повторяли
США) в качестве флуоресцентного красителя.
35 раз.
Продукты ОТ-ПЦР и ОТ-ПЦР в реальном
времени были анализированы в 1%-м агарозном
геле. Ампликоны выделяли из геля согласно про-
токолу Cleanup Standard («Евроген», Россия).
Фрагменты ДНК были секвенированы в ЗАО
«Евроген».
Статистическую обработку данных по экс-
прессии гена титина проводили с использовани-
ем непараметрического U-критерия Манна-
Уитни.
РЕЗУЛЬТАТЫ
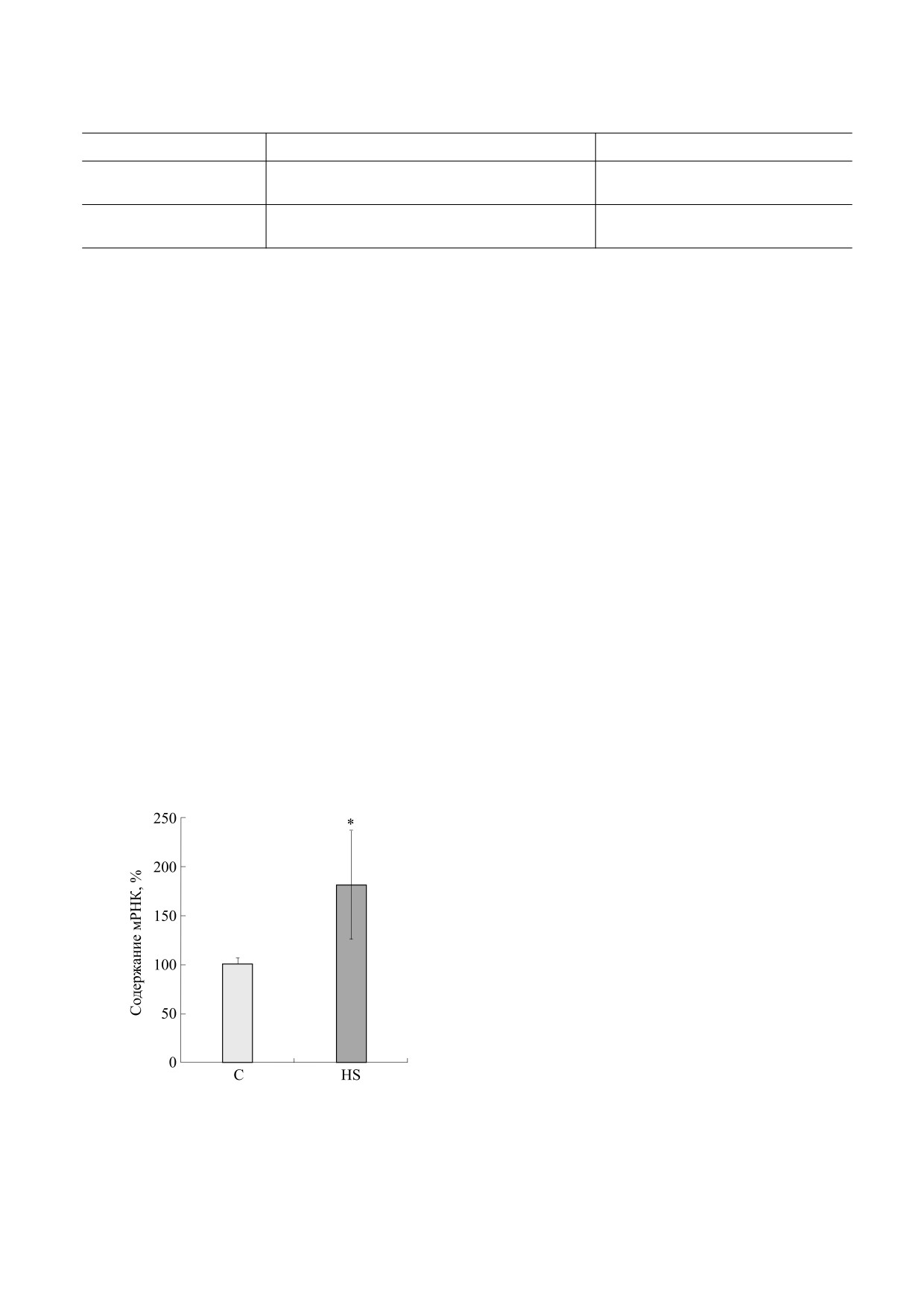
В результате проведенных экспериментов бы-
ло обнаружено достоверное увеличение (в 1,81 ра-
за, p ≤ 0,01) экспрессии гена титина в m. soleus
крыс группы HS (рис. 1).
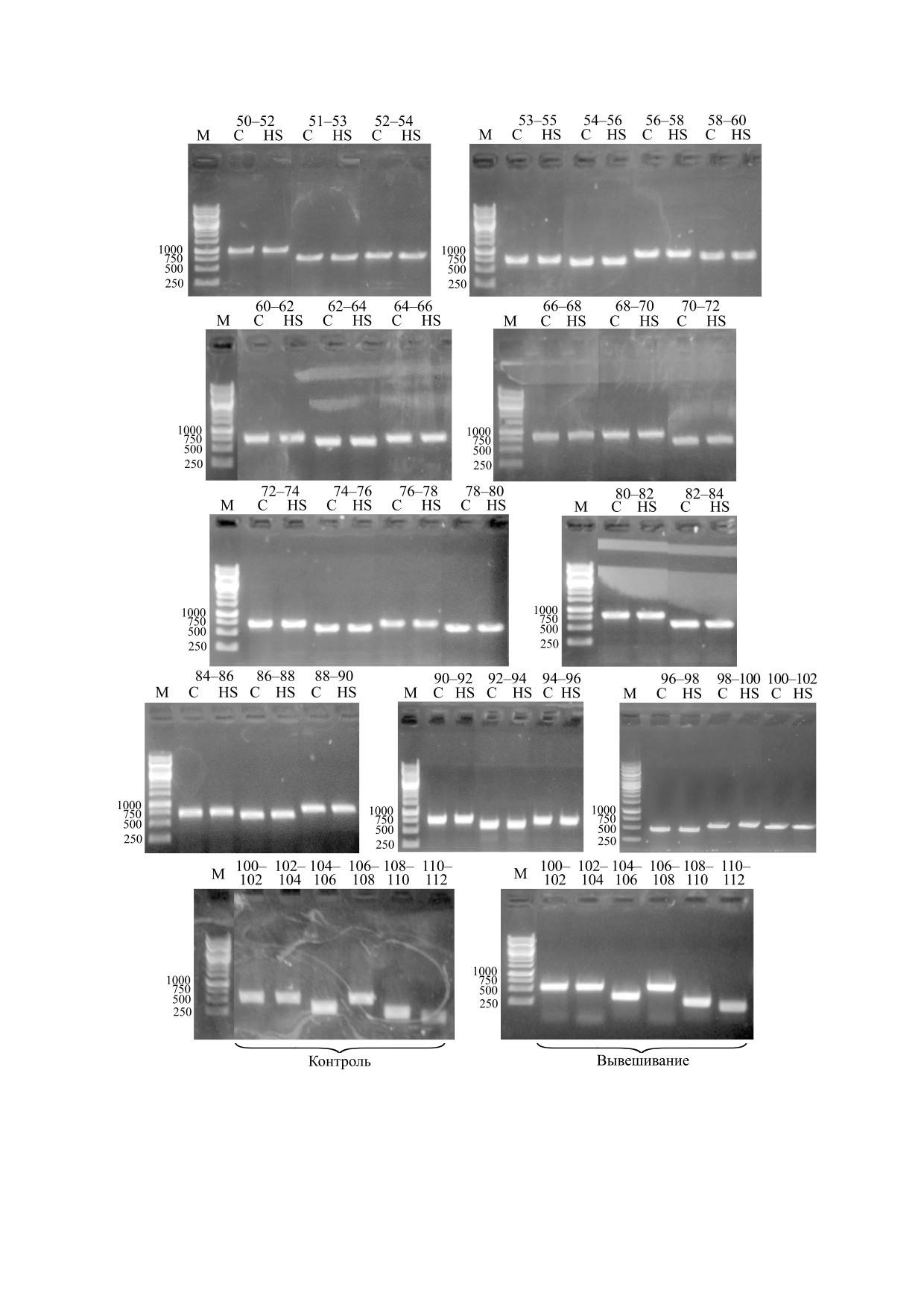
При этом, однако, не выявлено изменений в
Рис. 1. Экспрессия гена титина в m. soleus крыс: С -
контроль, HS - трехсуточное вывешивание; * p ≤ 0,01,
альтернативном сплайсинге пре-мРНК титина в
n = 6.
m. soleus обеих исследуемых групп крыс (рис. 2).
БИОФИЗИКА том 64
№ 5
2019
ТРЕХСУТОЧНАЯ ФУНКЦИОНАЛЬНАЯ РАЗГРУЗКА
857
Таблица 2. Пары праймеров для проведения ОТ-ПЦР
Праймер
Нуклеотидная последовательность
Праймер
Нуклеотидная последовательность
50F1
AGTGGTTCAAAGCTGGCAGAGAG
80F1
AGTTGCCAAGCAGGGAGAATC
51R1
CTGTTCAGGACAGGGGTGTT
80R1
GCATTCCAGTTGGATGGATT
52F1
CCTCCATTGCAAGCTGAAAG
82F1
AACTGGGAAGTCCAGAAGCA
52R1
AGTATGTGCCGCTGTCATCA
82R1
GGACAGTCAGGAGTGCCGAAC
53F1
AACTCTTGAGCCTGCACACA
84F1
AAGCTTAAGGACACGACTGCCAC
53R1
TGTGTGCAGGCTCAAGAGTT
84R1
AGTCAGAGTGCAGACGTTATCCG
54F1
CTGTTACCTTACAAGCTGCCGTG
86F1
CACGTACTCTGGCACACCAC
54R1
TCGTTTTCAGCTAGGCACGTGTA
86R1
TGGTGATGCCACAGTTAGGA
55R1
AAAATTTCAGCTGGGCAATG
88F1
CTCGGAACCTCTTCAGGTGT
56F1
GAGACATCGCTCCACTATTCACC
88R1
AGAGGTTCCGAGCCATTGAT
56R1
CCTTAAACCAGGAAACCCTCATG
90R1
CATTGCGAGCCTCACAGGTATAG
58F1
GAAAGGAGATGCCACTCAGCT
90F1
ATAGGCGGGTCTCCAGAAAT
58R1
CACTGCCAGCCTCGTTTTGA
92F1
CCAATGCTGGGAAATACACC
60F1
GAGGTACAGCTGCCTTCCAG
92R1
GCAGGTGTATTTCCCAGCAT
60R1
TTTCCCATCATGCTTGACTTC
94F1
ACAACACCTGTGCCCTAACC
62F1
AGCTGGAAGGCACAGACAGT
94R1
TAGGGCACAGGTGTTGTCTG
62R1
GTGCCTTCCAGCTGACTGAT
96F1
AATTCTGAGCAGCACTGTTGAGG
64F1
GGAGATACGGCATTTCTTTTGTG
96R1
TGCCAGAAGCGTTTTCAATGTAG
64R1
AGCTTCACAGACGTAAGTCCCAC
98F
GCACTGTTACGAACCCCATC
66F1
GAAAATGTCACCACCGTCTTG
98R
CAAAAACCACTGGCAATCCT
66R1
CACTCCTGACTCATTCTTGGC
100F
CTGCGAGATCACGTTCAAGA
68F1
CATTCCTCCATCGTTCACCAA
100R
CAGCAGAGCGTTGTTCTTGA
68R1
TCTCCTGCTACATTGGACACC
102F
CGCTGTGGATGGAAAACTCT
70R1
ATCTGCATGACTGCCACTGAG
102R
TGAGGCTCTGACACGAAGAA
70F1
TCAGTGGCAGTCATGCAGAT
104F
GCATGTATGGAATCACCGACT
72F1
GGTCAGAGAGAGCGAGAACG
104R
TTCAGCAGCTCAAATGCTTG
72R1
CGTTCTCGCTCTCTCTGACC
106F
AGCTGGAGCCCAGTAACAAA
74F1
TTCTCTGGACTCATCGGACA
106R
CCAGCTTCTCAGTCCCTTTG
74R1
CCACATTAGCAGCCACACAT
108F
CCATCGTCACCTGGTACAAA
76F1
TTGTCACCACGGAGAAAACA
108R
GGCATCGTCAAAGGACACTT
76R1
GGTCAGAATCTCCAGGATGC
110F
AAACTTGAGATGAAGCCTCCTG
78F1
GGCTTACCAGTTACCCTCAC
110R
CAGGAGGCTTCATCTCAAGTTT
78R1
CGTAGAAGATGCTGTGCCAAGTG
112R
TTCAATTCGTTTTGCTGGTG
Примечание. F - прямой праймер, R - обратный праймер.
В частности, результаты ПЦР-анализа показали
сии гена титина в m. soleus крысы после моделиру-
наличие всех экзонов (в диапазоне от 50-го до
емой в течение трех суток гравитационной
111-го), кодирующих самый вариабельный уча-
разгрузки. Известно, что атрофия, вызванная
сток скелетной N2A-изоформы титина в I-зоне
функциональной разгрузкой, сопровождается
саркомера.
снижением содержания интактных изоформ ти-
тина и увеличением содержания его протеолити-
ческих фрагментов [2-6,16]. Эти изменения явля-
ОБСУЖДЕНИЕ
ются следствием повышенного протеолиза
Увеличение экспрессии гена TTN обнаружено
титина кальпаинами
- кальций-зависимыми
ранее в сердечной и трех скелетных мышцах мы-
протеазами, активность которых возрастает в
шей после 30-суточного космического полета [6].
мышце уже в течение первых суток функциональ-
Наши результаты показали увеличение экспрес-
ной разгрузки [32]. Известны данные, что полу-
БИОФИЗИКА том 64
№ 5
2019
858
УЛАНОВА и др.
Рис. 2. Анализ альтернативного сплайсинга пре-мРНК титина: С - контроль, HS - трехсуточное вывешивание.
Обозначения: 50-52, 51-53 и т.д. - экзоны; М - маркер (п.о.).
БИОФИЗИКА том 64
№ 5
2019
ТРЕХСУТОЧНАЯ ФУНКЦИОНАЛЬНАЯ РАЗГРУЗКА
859
период жизни титина составляет трое суток [33].
БЛАГОДАРНОСТИ
Учитывая это, можно ожидать уменьшения со-
Работа выполнена с использованием оборудо-
держания титина на третьи сутки функциональ-
вания коллективного пользования ИТЭБ РАН и
ной разгрузки, однако ранее мы этого не обнару-
ГНЦ РФ - ИМБП РАН.
жили [17]. Вполне вероятно, что увеличение
экспрессии гена TTN (рис. 1) вносит компенса-
ФИНАНСИРОВАНИЕ РАБОТЫ
ционный вклад в поддержание содержания тити-
на в первые трое суток функциональной раз-
Работа выполнена при финансовой поддержке
грузки.
Российского научного фонда (грант
№18-15-
00062).
Изменений альтернативного сплайсинга пре-
мРНК титина в m. soleus вывешенных крыс не об-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
наружено. У млекопитающих в мышце soleus, со-
держащей преимущественно «медленную» изо-
Проведение опытов на животных было одоб-
форму I ТЦМ, транскрипты мРНК титина содер-
рено комиссией по биомедицинской этике ГНЦ
жат весь набор экзонов в диапазоне с 50-го по
РФ - ИМБП РАН (Москва).
111-й [23]. Трансляция таких транскриптов мРНК
сопровождается синтезом N2A-изоформы титина
КОНФЛИКТ ИНТЕРЕСОВ
с молекулярной массой 3,7 МДа. В мышце psoas,
Авторы заявляют об отсутствии конфликтов
содержащей «быстрые» изоформы ТЦМ, тран-
интересов.
скрипты мРНК не содержат экзоны в диапазоне с
53-го по 69-й [23]. Трансляция таких транскрип-
тов мРНК сопровождается синтезом N2A-изо-
СПИСОК ЛИТЕРАТУРЫ
формы титина с молекулярной массой ~3,4 МДа
1.
T. M. Mirzoev and B. S. Shenkman, Biochemistry
[23]. Показано, что в разгруженной мышце про-
(Moscow)
83
(11),
1299
(2018). DOI:
10.1134/
исходят изменения в изоформном составе ТЦМ,
S0006297918110020.
направленные на уменьшение доли «медленной»
2.
T. Toursel, L. Stevens, H. Granzier, and Y. Mounier, J.
изоформы I ТЦМ и увеличение доли «быстрых»
Appl. Physiol. (1985), 92 (4),
(2002).
1465. DOI:
изоформ этого белка [16,25,26]. При этом обнару-
10.1152/japplphysiol.00621.2001.
жено, что существенные изменения на уровне
3.
B. S. Shenkman, T. L. Nemirovskaya, IN Belozerova,
мРНК этих изоформ наблюдаются уже через 24 ч
et al., J. Gravit. Physiol. 9 (1), P139 (2002).
функциональной разгрузки, и еще больше увели-
4.
J. Udaka, S. Ohmori, T. Terui, et al., J. Gen. Physiol.
чиваются к третьим суткам [25,26]. Учитывая это,
131 (1), 33 (2008). DOI: 10.1085/jgp.200709888.
мы ожидали обнаружить в m. soleus вывешенных
5.
I. M. Vikhlyantsev and Z. A. Podlubnaya, Biophysics
крыс появление транскриптов мРНК, которые
53 (6), 592 (2008).
содержат не все экзоны в исследуемом нами диа-
6.
A. Ulanova, Y. Gritsyna, I. Vikhlyantsev, et al.,
пазоне (с 50-го по 111-й). Однако таковых измене-
Biomed. Res. Int.
2015,
104735
(2015). DOI:
ний обнаружено не было. В частности, ПЦР-ана-
10.1155/2015/104735.
лиз подтвердил наличие всех экзонов в исследуе-
7.
L. Tskhovrebova and J. Trinick, J. Mol. Biol. 265 (2),
мом диапазоне мРНК в m. soleus исследованных
100 (1997). DOI: 10.1006/jmbi.1996.0717.
групп крыс.
8.
D. O. Fürst, M. Osborn, R. Nave, and K. Weber, J. Cell
Biol. 106 (5), 563 (1988).
9.
A. Houmeida, J. Holt, L. Tskhovrebova, and J. Trinick,
ВЫВОДЫ
J. Cell Biol. 131 (6, Pt 1), 1471 (1995).
Впервые обнаружено, что трехсуточная функ-
10.
J. K. Freundt and W.A. Linke, J. Appl. Physiol. 126 (5),
циональная разгрузка сопровождается увеличе-
1474
(1985),
(2018). DOI:
10.1152/japplphysi-
нием экспрессии гена TTN. При этом не выяв-
ol.00865.2018.
лено изменений в альтернативном сплайсинге
11.
P. F. van der Ven, J. W. Bartsch, M. Gautel, et al., J.
пре-мРНК титина в диапазоне экзонов с 50-го по
Cell Sci. 113 (Pt 8), 1405 (2000).
111-й. Таким образом, в сравнении с общей экс-
12.
R. Horowits, E. S. Kempner, M. E. Bisher, and
прессией гена титина, альтернативный сплай-
R. J. Podolsky, Nature 323 (6084), 160 (1986). DOI:
синг его пре-мРНК является более консерватив-
10.1038/323160a0.
ным процессом. Вполне вероятно, что на более
13.
H. L. Granzier and S. Labeit, Exerc. Sport Sci. Rev. 34
поздних сроках разгрузки будут происходить из-
(2), 50 (2006).
менения не только экспрессии гена TTN, но и
14.
T. Voelkel and W. A. Linke, Pflugers Arch. 462 (1), 143
альтернативного сплайсинга пре-мРНК титина.
(2011). DOI: 10.1007/s00424-011-0938-1.
БИОФИЗИКА том 64
№ 5
2019
860
УЛАНОВА и др.
15. M. Gautel, Pflugers Arch. 462 (1), 119 (2011). DOI:
25. J. M. Giger, P. W. Bodell, M. Zeng, et al., J. Appl.
10.1007/s00424-011-0946-1.
Physiol. 107 (4), 1204 (2009). DOI: 10.1152/jap-
plphysiol.00344.2009.
16. B. S. Shenkman, Z. A. Podlubnaya, I. M. Vikhlyantsev,
et al., Biophysics 49 (5), 807 (2004).
26. N. A. Vilchinskaya, E. P. Mochalova, T. L. Nemirov-
skaya, et al., J. Physiol. 595 (23), 7123 (2017). DOI:
17. E. V. Ponomareva, V. V. Kravtsova, E. V. Kachaeva,
10.1113/JP275184.
et al., Biophysics 53 (6), 615 (2008). DOI: 10.1134/
27. V. E. Novikov and E. A. Ilyin, Aviat. Space Environ.
S0006350908060274.
Med. 52 (9), 551 (1981).
18. M. L. Bang, T. Centner, F. Fornoff, et al., Circ. Res. 89
28. R. K. Globus and E. Morey-Holton, J. Appl. Physiol.
(11), 1065 (2001).
120
(10),
1196
(2016). DOI:
10.1152/japplphysi-
19. W. Guo and M. Sun, Biophys. Rev. 10 (1), 15, (2018).
ol.00997.2015.
DOI: 10.1007/s12551-017-0267-5.
29. G. J. Viljoen, L. H. Nel, and J. R. Crowther, Molecular
20. Z. Chen, R. Maimaiti, C. Zhu, et al., J. Cell Biochem.
diagnostic PCR handbook (Springer, The Netherlands,
119 (12), 9986 (2018). DOI: 10.1002/jcb.27328.
2005).
30. K. J. Livak and T. D. Schmittgen, Methods 25, 402
21. S. Labeit, S. Lahmers, C. Burkart, et al., J. Mol. Biol.
(2001). DOI: 10.1006/meth.2001.1262.
362 (4), 664 (2006). DOI: 10.1016/j.jmb.2006.07.077.
31. S. Li, W. Guo, C. N. Dewey, and M. L. Greaser, Nucl.
22. A. Freiburg, K. Trombitas, W. Hell, et al., Circ. Res. 86
Acids Res.
41
(4),
2659
(2013).
DOI:
(11), 1114 (2000).
10.1093/nar/gks1362.
23. W. Guo, S. J. Bharmal, K. Esbona, and M. L. Greaser,
32. D. L. Enns, T. Raastad, I. Ugelstad, and A. N. Belcas-
J. Biomed. Biotechnol. 2010, 753675 (2010). DOI:
tro, Eur. J. Appl. Physiol. 100 (4), 445 (2007). DOI:
10.1155/2010/753675.
10.1007/s00421-007-0445-4.
24. C. Neagoe, C. A. Opitz, I. Makarenko, and W. A. Lin-
33. W. B. Isaacs, I. S. Kim, A. Struve, and A. B. Fulton, J.
ke, J. Muscle Res. Cell Motil. 24 (2-3), 175 (2003).
Cell Biol. 109 (5), 2189 (1989).
Three-Day Functional Unloading is Accompanied by an Increase in the TTN Gene
Expression in Rat Soleus Muscle without Changes in Alternative Splicing
from Exon 50 to Exon 111
A.D. Ulanova*, Yu.V. Gritsyna*, V.K. Zhalimov**, L.G. Bobyleva*, S.P. Belova***,
T.L. Nemirovskaya***, B.S. Shenkman***, and I.M. Vikhlyantsev* ****
*Institute of Theoretical and Experimental Biophysics, Russian Academy of Sciences,
Institutskaya ul. 3, Pushchino, Moscow Region, 142290 Russia
**Institute of Cell Biophysics, Russian Academy of Sciences, Institutskaya ul. 3, Pushchino, Moscow Region, 142290 Russia
***State Scientific Center - Institute of Biomedical Problems, Russian Academy of Sciences,
Khoroshevskoye Shosse 76a, Moscow, 123007 Russia
****Pushchino State Institute of Natural Sciences, Prosp. Nauki 3, Pushchino, Moscow Region, 142290 Russia
Changes in the expression of a titin gene and alternative splicing of titin pre-mRNA from exon 50 to exon 111
in rat soleus muscle were analyzed following three-day functional unloading (the HS group). Using real-time
RT-PCR, it was found that the expression level of the titin gene in rat soleus muscle from the HS group was
higher by 1.81 times (p ≤ 0.01, n = 6) than the control one (n = 7). It was shown that all studied exons of titin
mRNA are present in rats soleus muscle from the two groups. Our results demonstrate that three-day func-
tional unloading is accompanied by an increase in the expression of the titin gene in rat soleus without chang-
es in alternative splicing from exon 50 to exon 111.
Keywords: functional unloading, titin gene, alternative splicing, m. soleus
БИОФИЗИКА том 64
№ 5
2019