БИОФИЗИКА, 2019, том 64, № 5, с. 910-916
БИОФИЗИКА КЛЕТКИ
УДК 57.081.23
РАЗРАБОТКА И ПРОГРАММНАЯ РЕАЛИЗАЦИЯ АЛГОРИТМА
ОПРЕДЕЛЕНИЯ СРЕДНЕЙ ДЛИНЫ САРКОМЕРА КАРДИОМИОЦИТА
© 2019 г. Т.А. Мячина* **, К.А. Бутова* **, О.Н. Лукин* **
*Институт иммунологии и физиологии Уральского отделения РАН, 620049, Екатеринбург, ул. Первомайская, 106
**Уральский федеральный университет имени первого Президента России Б.Н. Ельцина,
620002, Екатеринбург, ул. Мира, 19
E-mail: myachina.93@mail.ru
Поступила в редакцию 01.07.2019 г.
После доработки 01.07.2019 г.
Принята к публикации 12.07.2019 г.
Представлен разработанный авторами настраиваемый алгоритм для определения средней длины
саркомера в изолированных кардиомиоцитах, активно сокращающихся в ненагруженных или на-
груженных условиях. Алгоритм основан на применении дискретного преобразования Фурье и
включает в себя дополнительную обработку исходного сигнала саркомерной исчерченности кар-
диомиоцита и полученного в результате дискретного преобразования Фурье амплитудно-частотно-
го спектра для более точного определения длины саркомера. Продемонстрирована эффективность
алгоритма для анализа данных саркомерной исчерченности изолированных клеток сердца.
Ключевые слова: кардиомиоцит, саркомер, сократительный ответ, дискретное преобразование Фурье.
DOI: 10.1134/S0006302919050119
черченности, связанная с ограничениями изме-
Изолированные сократительные клетки сер-
рительной аппаратуры и невозможностью пол-
дечной мышцы (кардиомиоциты) являются удоб-
ной пространственной фиксации измеряемого
ным экспериментальным объектом для изучения
профиля этой исчерченности из-за подвижности
механических характеристик сокращения, так
активно сокращающегося кардиомиоцита [1,9].
как позволяют избежать влияния соединитель-
При этом если в условиях эксперимента с задани-
ной ткани и эндотелия, значительно изменяю-
ем (не)контролируемой нагрузки на изолирован-
щих вязкоэластичные свойства миокарда [1-4].
ный миоцит (например, при использовании ме-
Кардиомиоциты могут быть задействованы в раз-
тода карбоновых волокон [3,4]) неточность опре-
личных исследованиях, в том числе при изучении
деления саркомерной исчерченности снижается
региональной и трансмуральной функциональ-
за счет механической фиксации самой клетки, то
ной неоднородности различных участков мио-
в условиях ненагруженных сокращений могут
карда [4-6]. При этом сократительный ответ са-
возникать заметные смещения кардиомиоцита из
мого кардиомиоцита, т.е. амплитуда его укороче-
поля зрения. Кроме того, для оценки сократи-
ния, может отличаться от сократительного
тельного ответа клетки и всех составляющих ее
поведения составляющих его саркомеров, как по
саркомеров необходима обширная автономная
причине внутренней неоднородности свойств
обработка массива данных с учетом наличия не-
саркомеров и наличия внутреннего укорочения
однородности среди отдельных саркомеров
[7], так и в силу ограничений, связанных с недо-
[2,4,10], что препятствует быстрой регистрации
статочной фиксацией измерительно-задающих
сокращений и снижает точность определения их
устройств к кардиомиоциту [4]. Поэтому динами-
амплитудных и временных характеристик.
ческое изменение длины саркомера, по сравне-
нию с длиной клетки или мышцы, является наи-
Большинство методов определения длины
более информативным и точным для оценки
саркомеров кардиомиоцита основано на исполь-
механических характеристик сократительной ак-
зовании быстрого или дискретного преобразова-
тивности миокарда в условиях in vitro [1,4,7-9].
ний Фурье [1,8-11]. Применимость этих алгорит-
Однако существует проблема адекватной реги-
мов зависит от размера области изображения
страции динамического сигнала саркомерной ис- клетки, выбранной для анализа [10]. На сего-
дняшний день может быть использовано изобра-
Сокращения: ДПФ - дискретное преобразование Фурье.
жение целого кардиомиоцита для регистрации
910
РАЗРАБОТКА И ПРОГРАММНАЯ РЕАЛИЗАЦИЯ АЛГОРИТМА
911
саркомерной динамики с активно сокращающей-
Регистрация данных. Измерение механической
ся клетки [1,8,10,11], но в большинстве случаев
функции изолированных кардиомиоцитов вы-
сохраняется необходимость выбора ограничен-
полняли при помощи системы лазерной сканиру-
ной области саркомерной исчерченности с целью
ющей конфокальной микроскопии LSM 710 (Carl
ускорения регистрации данных [9,12]. При этом
Zeiss, Германия) и программного обеспечения
существующие программные реализации лише-
для регистрации сигнала изображения клетки в
ны возможности расширенной пользовательской
проходящем свете ZEN 2010 (Carl Zeiss, Герма-
настройки алгоритма определения средней дли-
ния). Непосредственно перед измерением в зоне
ны саркомера, а также не предусматривают ис-
наилучшей саркомерной исчерченности клетки
пользования безотносительно к режиму нагруже-
выбирали узкую область сканирования высотой в
ния кардиомиоцита, т.е. как для ненагруженного,
два-три пикселя и длиной, соответствующей дли-
так и нагруженного сокращений.
не всего кардиомиоцита, включая карбоновые
волокна, в случае нагруженного сокращения
Для корректной оценки саркомерной динами-
(рис. 1а и 1б). Это позволяло осуществлять скани-
ки в кардиомиоците, активно сокращающемся в
рование изображения клетки в выбранной обла-
ненагруженном или нагруженном режиме, нами
сти со скоростью 300-500 кадров в секунду (~2-
был разработан настраиваемый алгоритм для
3 мс на кадр) и регистрировать изменение длины
определения средней длины саркомера в ограни-
саркомеров клетки с высокой временной дискре-
ченном регионе кардиомиоцита. Алгоритм осно-
тизацией. Результат сканирования такого изобра-
ван на методе дискретного преобразования Фурье
жения представлял собой профиль изменения
и реализован в рамках разработанного нами про-
интенсивности сигнала, который имел периоди-
граммного обеспечения EqapAll6.
ческий характер, связанный с саркомерной ис-
черченностью клетки (рис. 1в и 1г). Изображения
МЕТОДЫ ИССЛЕДОВАНИЯ
регистрировали с интервалом 2-3 мс, их последо-
вательность позволяла анализировать динамику
В экспериментах использованы здоровые кры-
саркомерной длины в активно сокращающемся
сы линии Вистар (самки и самцы возрастом два-
кардиомиоците.
три месяца).
Программное обеспечение. Интерфейс и моду-
Получение изолированных кардиомиоцитов.
ли программного обеспечения EqapAll6 были
Одиночные кардиомиоциты были получены с ис-
разработаны в среде объектно-ориентированного
пользованием стандартной методики ретроград-
программирования IDE Borland Delphi 6 (Borland
ной перфузии (по Лангендорфу) целого сердца
International Inc., США) с использованием языка
крысы модифицированным раствором Кребса-
программирования Object Pascal.
Хенселейта, содержащим коллагенлитический
фермент для разрушения соединительной ткани
[13]. Измерения сократительной активности кар-
РЕЗУЛЬТАТЫ
диомиоцитов были выполнены в растворе Тироде
Исходный сигнал. В качестве исходного (вход-
следующего состава (в мМ): NaCl - 140, KCl -
ного) сигнала для алгоритма выступает последо-
5,4, MgSO4, - 1, HEPES - 10, глюкоза - 11,1,
вательность изображений области сканирования
CaCl2 - 1,25.
кардиомиоцита, алгоритм анализирует отдельно
Метод карбоновых волокон. Реализация сокра-
каждое такое изображение, по профилю сарко-
щений кардиомиоцитов в ауксотоническом (на-
мерной исчерченности которого определяется
средняя длина саркомера. В случае использова-
груженном) режиме при последовательном уве-
ния карбоновых волокон получаемый профиль
личении степени диастолического растяжения
интенсивности включает два провала яркости,
клетки осуществлялась при помощи метода кар-
боновых волокон [3,4,14-16]. Были использова-
соответствующих положению волокон. Профиль
ны карбоновые волокна толщиной 7 мкм (Tsuku-
интенсивности сигнала представляет собой
функцию пространственного смещения от левой
ba Materials Information Laboratory Ltd., Япония),
границы изображения, поэтому основная частота
вклеенные в специальные стеклянные держатели.
сигнала определяется как расстояние между пи-
Держатели с карбоновыми волокнами монтиро-
вали на независимых прецизионных микромани-
ками периодической составляющей, для которой
пуляторах с использованием системы управления
рассчитывается среднее значение длины сарко-
мера.
микропозиционирующими устройствами MP285
(Sutter Instrument, США). Для задания растяже-
Начальные пользовательские настройки алго-
ния клетки и регистрации ее укорочения исполь-
ритма. На начальном этапе алгоритм определения
зовали два карбоновых волокна, которые механи-
среднего значения длины саркомера позволяет
чески фиксировали к концам кардиомиоцита за
пользователю вручную задать величину отступа
счет адгезии и сил электростатического взаимо-
от края миоцита (при регистрации ненагружен-
действия.
ного сокращения) или внутренних краев карбо-
БИОФИЗИКА том 64
№ 5
2019
912
МЯЧИНА и др.
Рис. 1. Регистрация профиля саркомерной исчерченности в изолированном кардиомиоците: (a) - изображение кар-
диомиоцита в ненагруженном режиме сокращения; (б) - пример профиля интенсивности изображения кардиомио-
цита при сокращении в ненагруженном режиме; (в) - изображение кардиомиоцита в нерастянутом и растянутом со-
стоянии при использовании карбоновых волокон; (г) - пример профиля интенсивности изображения кардиомиоцита
при сокращении в нагруженном режиме с использованием карбоновых волокон, области с минимальной интенсив-
ностью соответствуют положениям волокон. На панелях (а) и (в) показана область, для которой строится профиль ин-
тенсивности изображения кардиомиоцита. Данные на панелях (в) и (г) адаптированы и модифицированы из работы
[13].
новых волокон (при регистрации ауксотониче-
Наконец, до начала работы алгоритма пользо-
ского сокращения при разной степени растяже-
вателем вручную задаются следующие парамет-
ния клетки) и не включать в анализ часть сигнала,
ры: близость к пику спектрального распределе-
близко расположенного к указанным краям. Это
ния (Proximity) и максимальное число гармоник.
позволяет уменьшить неточность определения
Параметр Proximity задает, в пределах какой вели-
длины саркомера, если в исходную область ска-
чины можно отклоняться от пика спектра «про-
нирования включены участки изображения за
странственных» частот для определения диапазо-
пределами клетки. В случае использования кар-
на длин саркомеров, в котором будет вычислять-
боновых волокон, из анализа исключаются участ-
ся среднее значение длины саркомера (рис. 2а).
ки изображения за пределами карбоновых воло-
Например, при Proximity = 90% для спектра про-
кон (данные участки не подвергаются растяже-
странственных частот будет определяться окно
нию при увеличении длины клетки), а также
интенсивностей от 90% до 100% пика интенсив-
участки, близко расположенные к внутренним
ности, далее будут находиться соответствующие
краям карбоновых волокон (данные участки мо-
этому окну самое меньшее и самое большее зна-
чения длины саркомера, затем для этого диапазо-
гут иметь искажения саркомерной исчерченно-
сти из-за искривления поверхности клетки вбли-
на длин будет определяться средняя длина сарко-
мера. Причина использования такого параметра
зи карбоновых волокон).
состоит в том, что спектр частот может иметь не
Также вручную задаются минимальное и мак-
один острый пик, а форму с двумя близко распо-
симальное допустимое значение длины саркоме-
ложенными пиками, каждый из которых доста-
ра (например, 1,3 и 2,5 мкм, что соответствует фи-
точно достоверно описывает среднюю длину сар-
зиологическому диапазону длин саркомеров).
комера. Такая ситуация возможна, если в исход-
Это позволяет исключить из анализа гармоники
ном сигнале исчерченности имеются две
слишком высокой или низкой пространственной
«популяции» саркомеров с различными длинами
«частоты», которые могут перекрывать по интен-
(например, из-за внутренней неоднородности
сивности основную гармонику.
распределения длин саркомеров в кардиомиоци-
БИОФИЗИКА том 64
№ 5
2019
РАЗРАБОТКА И ПРОГРАММНАЯ РЕАЛИЗАЦИЯ АЛГОРИТМА
913
Рис. 2. Пояснение к начальным настройкам алгоритма определения средней длины саркомера: (а) - параметр Proxi-
mity задает окно относительно пика спектра пространственных частот для определения средней длины саркомеров
(затемненная область в примере на рисунке); (б) - максимальное число гармоник определяет корректность
определения основной гармоники, при недостаточном
числе гармоник спектр частот не содержит основную
гармонику.
те). В этом случае целесообразно определять не
ствует определенной длине саркомера), I - ин-
длину саркомера, соответствующую пику спектра
тенсивность гармоники.
гармоник, а некоторый диапазон длин для не-
Исключение низкочастотной компоненты. Не-
скольких квазипиковых значений. Чем ниже зна-
посредственно до применения к исходному сиг-
чение параметра Proximity, тем менее точно вы-
налу саркомерной исчерченности метода дис-
числяется средняя длина саркомера. Максималь-
кретного преобразования Фурье (ДПФ) пользо-
ное число гармоник, которые алгоритм будет
ватель может задать исключение низкочастотных
определять в периодическом сигнале, прямо вли-
колебаний этого сигнала, которые могут возник-
яет на возможность определения правильной ос-
нуть из-за неравномерной яркости изображения
новной гармоники, поскольку уменьшение этого
клетки или наличия посторонних структур в об-
числа приводит к сужению исследуемого спектра
ласти регистрации изображения (например, ядра
частот (рис. 2б). При задании слишком маленько-
или митохондрии). Для исключения низкоча-
го числа гармоник спектр будет ограничен диапа-
стотной составляющей применяется глубокая
зоном длин, не включающим основную гармони-
фильтрация исходного сигнала с управляемыми
ку, т.е. спектральный сигнал будет неинформати-
настройками фильтра, затем исходный сигнал
вен и средняя длина саркомера будет определена
корректируется с учетом базового уровня отфиль-
некорректно.
трованного сигнала (рис. 3а).
Как было указано выше, длина саркомеров в
исходном сигнале может быть распределена не-
Метод дискретного преобразования Фурье. Да-
равномерно. В связи с этим средняя длина сарко-
лее в алгоритме реализуется ДПФ для определе-
мера (SLMEAN) в заданном диапазоне длин по же-
ния спектральной характеристики сигнала про-
филя интенсивности, т.е. строится распределе-
ланию пользователя может определяться без уче-
ние интенсивности гармоники периодического
та или с учетом относительной амплитуды
интенсивности (веса) каждой пространственной
сигнала исчерченности от ее частоты. В данном
частоты; в последнем случае средняя длина сар-
случае сигнал имеет не временное, а простран-
ственное изменение (изменение яркости изобра-
комера вычисляется по формуле:
жения по горизонтали), в связи с этим периоды
N
составляющих его гармоник определяются в еди-
∑
SL
i
*
Ii
ницах длины. Откорректированный с учетом вы-
i
=1
шеописанных настроек сигнал исчерченности
SL
MEAN
=
,
N
является входным массивом для ДПФ, где опре-
SLi
∑
деляется частотный спектр, состоящий из масси-
i=1
вов реальных и мнимых чисел. При необходимо-
где N - число рассчитываемых гармоник, SL -
сти, к этим массивам может быть применена раз-
«пространственная» частота гармоники (соответ-
дельная фильтрация, после чего результирующие
БИОФИЗИКА том 64
№ 5
2019
914
МЯЧИНА и др.
Рис. 3. Примеры использования дополнительных настроек в алгоритме определения средней длины саркомера: (а) -
исключение низкочастотной компоненты исходного сигнала исчерченности; (б) - фильтрация амплитудно-
частотного спектра, полученного после применения ДПФ.
отфильтрованные массивы реальных и мнимых
нии длины саркомера в случаях ненагруженного
чисел для частотного спектра используются для
или нагруженного сокращения изолированного
повторной «сборки» этого спектра и вычисления
кардиомиоцита и реконструкции саркомерной
средней длины саркомера уже на основе этого
динамики, в том числе при задании диастоличе-
спектра. Полученный амплитудно-частотный
ского растяжения кардиомиоцита.
спектр также может быть подвергнут фильтрации
с пользовательскими настройками. Например, в
ЗАКЛЮЧЕНИЕ
случае если амплитудно-частотный спектр содер-
жит два близко расположенных пика, или имеет
В данном исследовании был разработан и про-
отклонения вблизи одного пика, применение
тестирован настраиваемый алгоритм для вычис-
этого фильтра может дать более точную оценку
ления средней длины саркомера, основанный на
средней длины саркомера (рис. 3б).
исходном паттерне поперечно-полосатой исчер-
ченности. Как и многие другие методы, описан-
Завершающим этапом является определение
ные в литературе [1,8,9,11], наш алгоритм базиру-
пикового значения амплитудно-частотного спек-
ется на применении дискретного преобразования
тра, и все последующие вычисления средней дли-
Фурье для определения основного простран-
ны саркомера (с учетом описанных ранее пара-
ственного периода в сигнале поперечной исчер-
метров) привязываются к этому пику. Именно из
ченности кардиомиоцита. Однако имеется прин-
амплитудно-частотного спектра выделяется ос-
ципиальное отличие в исходных данных, исполь-
новная гармоника или диапазон гармоник в соот-
зуемых для нашего алгоритма, поскольку эти
ветствии со значениями параметров Proximity,
данные получены при сканировании узкого
максимального числа гармоник, минимального и
длинного региона кардиомиоцита. В этом случае
максимального допустимого значения длины
невозможно реализовывать расширенные кор-
саркомера. Для этой гармоники вычисляется
ректировки, обязательные для данных, зареги-
длина саркомера. При использовании описанных
стрированных с целого кардиомиоцита (напри-
выше фильтров (исключение низкочастотного
мер, осуществлять оценку смещения миоцита
компонента, фильтрация массивов реальных и
относительно линии сканирования или неодно-
мнимых чисел частотного спектра, фильтрация
родности популяции саркомеров в различных об-
амплитудно-частотного спектра) вычисляются
ластях клетки). Это накладывает ограничения на
три различных значения средней длины саркоме-
применения представленного в данной работе ал-
ра, которые можно сопоставлять друг с другом и
горитма по сравнению с другими алгоритмами
оценивать степень искажения вычисления при
[1,8,10]. С другой стороны, задание узкого регио-
включении/выключении фильтра.
на сканирования изображения клетки позволило
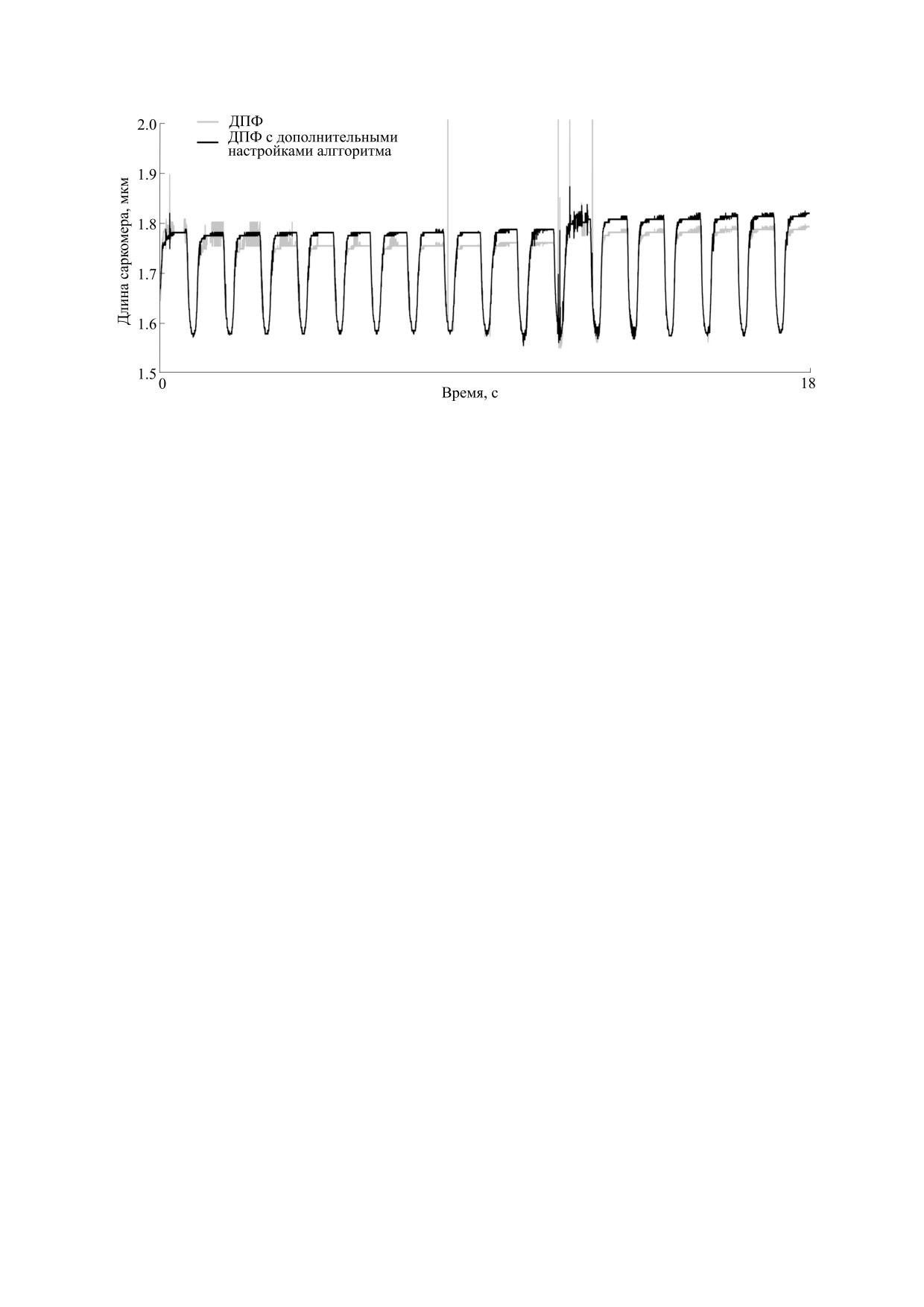
Пример работы алгоритма определения сред-
обеспечить временную дискретизацию, приемле-
ней длины саркомера показан на рис. 4. Алгоритм
мую для регистрации механического ответа кар-
может применяться при динамическом измене-
диомиоцита с высокой точностью.
БИОФИЗИКА том 64
№ 5
2019
РАЗРАБОТКА И ПРОГРАММНАЯ РЕАЛИЗАЦИЯ АЛГОРИТМА
915
Рис. 4. Репрезентативный пример работы алгоритма определения средней длины саркомера при использовании
дополнительных настроек (исключение низкочастотной компоненты, фильтрация амплитудно-частотного спектра).
Представленный в данной работе алгоритм
ней длины саркомера является адекватным для
был полностью реализован в самостоятельно раз-
оценки механической функции саркомера в ак-
работанном программном обеспечении EqapAll6.
тивно сокращающемся изолированном кардио-
Для этого есть ряд технических причин. Во-пер-
миоците при задании различных условий нагру-
вых, первоначально данные регистрировали в
жения или степени диастолического растяжения.
коммерческой программе ZEN2011 (Carl Zeiss,
Germany), которая не предоставляет возможно-
ФИНАНСИРОВАНИЕ РАБОТЫ
сти применения ДПФ для определения сарко-
мерной динамики. Во-вторых, по сравнению с
Работа выполнена в рамках госзадания ИИФ
другими коммерчески доступными программны-
УрО РАН (регистрационный номер НИОКТР
ми обеспечениями, специализирующихся на ана-
№АААА-А18-118020590031-8), при финансовой
лизе данных саркомерной исчерченности (на-
поддержке Российского фонда фундаментальных
пример, IonWizard 6.0, IonOptix Ltd.), наше про-
исследований (грант №18-04-00572-а) и поста-
граммное обеспечение позволяет настраивать
новления Правительства РФ № 211 от 16.03.2013 г.
ДПФ-алгоритм поиска основной гармоники в
сигнале саркомерной исчерченности и сохранять
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
исходные данные профилей исчерченности, а не
только полученные средние значения длины сар-
Все манипуляции с подопытными животными
комера, что дает возможность реализации расши-
проводились в соответствии с международным
ренного анализа динамики саркомеров [1,8,10].
правилам обращения с лабораторными животны-
ми (Директива 2010/63/ЕU) и были одобрены ко-
Также разработанный нами алгоритм преду-
миссией Института иммунологии и физиологии
сматривает возможность дополнительной филь-
УрО РАН по контролю за лабораторными живот-
трации данных, которая необходима в случае на-
ными.
рушения регистрируемого паттерна саркомерной
исчерченности вследствие неравномерного осве-
щения клетки или присутствия в кадре структур,
СПИСОК ЛИТЕРАТУРЫ
не являющихся саркомерами. Подобно ранее
1. P. Peterson, M. Kalda, and M. Vendelin, Am. J. Physi-
описанным в литературе методам [1,10], нами бы-
ol. Cell Physiol. 304 (6), C519 (2013).
ло реализовано установление основного «тренда»
2. A. J. Brady, Physiol. Rev. 71 (2), 413 (1991).
исходного сигнала (путем исключения низкоча-
3. G. Iribe, Biological, Physical and Technical Basics of Cell
стотной компоненты) непосредственно до запус-
Engineering (Springer, Singapore, 2018), pp. 193-207.
ка ДПФ. Кроме того, разработанный нами алго-
4. G. Iribe, M. Helmes, and P. Kohl, Am. J. Physiol.
ритм включает дополнительные фильтры с поль-
Heart Circ. Physiol. 292 (3), H1487 (2007).
зовательскими настройками, реализуемые уже
5. O. Cazorla, J. Y. Le Guennec, and E. White, J. Mol.
после ДПФ.
Cell. Cardiol. 32 (5), 735 (2000).
Таким образом, представленный в данной ра-
6. A. Khokhlova, G. Iribe, P. Konovalov, et al., J. Mol.
боте настраиваемый алгоритм определения сред-
Cell. Cardiol. 114, 276 (2018).
БИОФИЗИКА том 64
№ 5
2019
916
МЯЧИНА и др.
7. P. P. de Tombe and H. E. D. J. ter Keurs, J. Mol. Cell.
12. T. Myachina, A. Khokhlova, I. Antsygin, and
Cardiol. 91, 148 (2016).
O. Lookin, IOP Conf. Ser.: Mater. Sci. Eng. 350,
012011 (2018)
8. G. Bub, P. Camelliti, C. Bollensdorff, et al., Am. J.
Physiol. Heart Circ. Physiol. 298 (5), H1616 (2010).
13. O. N. Lookin and Y. L. Protsenko, Biophys. J. 63 (3),
441 (2018).
9. C. Pasqualin, F. Gannier, A. Yu, et al., Am. J. Physiol.
14. J. Y. Le Guennec, E. White, F. Gannier, et al., Exp.
Cell Physiol. 311 (2), C277 (2016).
Physiol. 76 (6), 975 (1991).
10. B. W. Infantolino, M. J. Ellis, and J. H. Challis, Anat.
15. S. Sugiura, S. Nishimura, S. Yasuda, et al., Nature Pro-
Rec. 293 (11), 1913 (2010).
tocols 1 (3), 1453 (2006).
11. M. Helmes, A. Najafi, B. M. Palmer, et al., Cardiovasc.
16. G. Iribe, T. Kaneko, Y. Yamaguchi, and K. Naruse,
Res. 111 (1), 66 (2016).
Prog. Biophys. Mol. Biol. 115 (2-3), 103 (2014).
The Development and Program Implementation of an Algorithm for Estimating a Mean
Sarcomere Length of a Cardiomyocyte
T.A. Myachina* **, K.A. Butova* **, and O.N. Lookin* **
*Institute of Immunology and Physiology, Ural Branch of the Russian Academy of Sciences,
ul. Pervomayskaya 106, Yekaterinburg, 620049 Russia
**Ural Federal University, ul. Mira 19, Yekaterinburg, 620002 Russia
This study describes our adjustable algorithm for determination of a mean sarcomere length in isolated car-
diomyocytes during active contraction under unloaded or loaded conditions. The algorithm is based on the
discrete Fourier transform and implements additional processing of an input signal for sarcomere striation in
cardiomyocyte and the amplitude-frequency spectral curve obtained after the discrete Fourier transform for
more precise determination of sarcomere length. The effectiveness of the algorithm to analyze sarcomere
striation patterns from isolated cardiac cells is shown.
Keywords: cardiomyocyte, sarcomere, contractile response, discrete Fourier transform
БИОФИЗИКА том 64
№ 5
2019