БИОФИЗИКА, 2019, том 64, № 5, с. 957-962
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 612.816
СТРУКТУРНО-ФУНКЦИОНАЛЬНЫЕ ХАРАКТЕРИСТИКИ КОНЦЕВЫХ
ПЛАСТИНОК КАМБАЛОВИДНОЙ МЫШЦЫ КРЫСЫ
ПРИ КРАТКОВРЕМЕННОМ НАРУШЕНИИ
ДВИГАТЕЛЬНОЙ АКТИВНОСТИ
© 2019 г. В.В. Кравцова, Е.А. Сабурова, И.И. Кривой
Cанкт-Петеpбуpгcкий гоcудаpcтвенный унивеpcитет, 199034, Cанкт-Петеpбуpг, Унивеpcитетcкая наб., 7/9
E-mail: iikrivoi@gmail.com
Поступила в редакцию 04.07.2019 г.
После доработки 04.07.2019 г.
Принята к публикации 12.07.2019 г.
Проведен анализ распределения никотиновых холинорецепторов, степени их фрагментации, пло-
щади концевых пластинок, а также мембранной локализации α2-изоформы Na,K-АТФазы в кам-
баловидной мышце крысы после функциональной разгрузки (метод антиортостатического выве-
шивания) в течение 1-3 суток. После разгрузки наблюдалось усиление фрагментации концевых
пластинок, сопровождающееся снижением площадей отдельных фрагментов; общая площадь кон-
цевых пластинок не изменялась. Плотность распределения никотиновых холинорецепторов также
не изменялась, однако наблюдалось достоверное снижение плотности распределения α2-изофор-
мы Na,K-АТФазы. Предположительно, выявленные изменения являются результатом адаптацион-
ных перестроек в скелетной мышце, которые развиваются не только при хронических нарушениях,
но уже на ранних этапах двигательной дисфункции.
Ключевые слова: моторная концевая пластинка, никотиновый xолиноpецептоp, изофоpмы Na,K-
АТФазы, двигательная активность.
DOI: 10.1134/S000630291905017Х
Ультраструктура нервно-мышечного соедине-
Моторная концевая пластинка нервно-мы-
ния и концевой пластинки существенно зависят
шечного синапса позвоночных представляет со-
от двигательной активности. Снижение двига-
бой высокоспециализированную область сарко-
тельной активности сопровождается многочис-
леммы, лежащую непосредственно под двига-
ленными нарушениями структуры концевой пла-
тельным нервным окончанием [1,2]. Мембрана
стинки, включая изменение ее площади и усиле-
концевой пластинки характеризуется исключи-
ние фрагментации в распределении нХР.
тельно высокой плотностью распределения ни-
Подобные нарушения наблюдаются после денер-
котиновых холинорецепторов (нХР), которые об-
вации [5] и продолжительной функциональной
разуют отдельные кластеры (фрагменты) [3]. В
разгрузки [6], при возрастных изменениях [3],
кластеризации нХР и стабилизации концевой
при миодистрофии [7-9] и других формах мы-
пластинки важную роль играют холестерин и ли-
шечной патологии [10].
пидные плотики, кроме того, нХР молекулярно и
функционально взаимодействуют с α2-изофор-
Дестабилизация концевой пластинки, сопро-
мой Na,K-АТФазы [4]. После освобождения из
вождающаяся нарушением структуры липидных
плотиков и снижением активности α2-изоформы
нервного окончания кванты ацетилхолина акти-
Na,K-АТФазы, наблюдалась уже в течение пер-
вируют нХР, что приводит к генерации потенци-
вых 6-12 ч снижения двигательной активности
ала концевой пластинки. Процесс трансформа-
камбаловидной мышцы крысы [11-13]. В дан-
ции этого локального потенциала в распростра-
ной работе нами впервые исследованы послед-
няющийся потенциал действия мышечного
ствия функциональной разгрузки задних конеч-
волокна является одним из ключевых этапов, от
ностей крысы в течение одних-трех суток. Про-
которого зависит не только надежность нервно-
веден анализ распределения нХР, степени их
мышечной передачи, но и ее пластичность [1,2].
фрагментации, площади концевых пластинок, а
Сокращения: нХР
- никотиновые холинорецепторы,
также мембранной локализации α2-изоформы
AMPK - АМФ-активируемая протеинкиназа.
Na,K-АТФазы.
957
958
КРАВЦОВА и др.
МЕТОДЫ ИССЛЕДОВАНИЯ
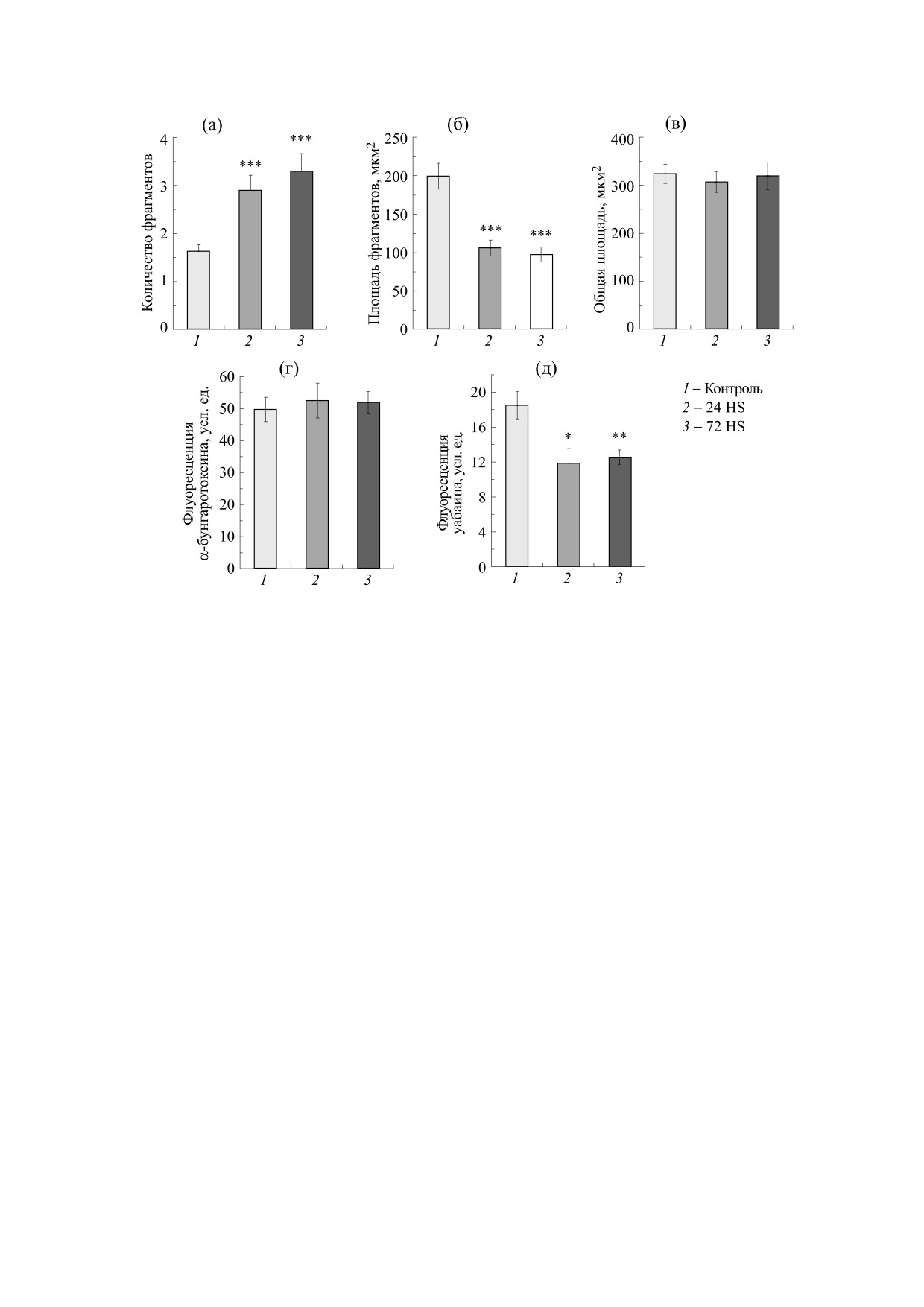
ментации (увеличение количества фрагментов)
концевых пластинок, сопровождающееся сниже-
Работа проведена на изолированных нервно-
нием площадей отдельных фрагментов. В резуль-
мышечных препаратах камбаловидных мышц
тате общая площадь концевых пластинок не
самцов крыс линии Wistar массой 190-210 г. Кры-
изменялась (рис. 2, а-в). Интенсивность флуо-
сы были разделены на две группы - контрольную
ресценции α-бунгаротоксина, т.е. плотность рас-
и группа крыс, которых подвергали антиортоста-
пределения нХР, также не изменялась после раз-
тическому вывешиванию задних конечностей
грузки (рис. 2г). Таким образом, наша модель
(hindlimb suspension, HS) в течение 24 или 72 ч
кратковременной разгрузки существенно отлича-
традиционным способом [14].
ется по этому показателю от моделей хрониче-
Сразу после выделения мышцу с отрезком
ской двигательной дисфункции - мышей линий
нерва помещали в экспериментальную камеру с
mdx (модель миодистрофии Дюшенна) и Bla/J
физиологическим раствором следующего состава
(одна из моделей дисферлинопатии), у которых
(в мМ): NaCl - 137; KCl - 5; CaCl2 - 2; MgCl2 - 2;
наблюдалось снижение плотности распределения
NaHCO3 - 24; NaH2PO4 - 1; глюкоза - 11; pH 7,4.
нХР [8,9]. В то же время через 24 и 72 ч разгрузки
Раствор аэрировали карбогеном (95% O2 + 5%
наблюдалось достоверное снижение относитель-
CO2). Для определения локализации нХР и α2-
ной интенсивности флуоресценции уабаина (α2-
изоформы Na,K-АТФазы в области моторной
изоформы Na,K-АТФазы) (рис. 2д), что может
концевой пластинки использовали соответствен-
отражать снижение плотности распределения
но α-бунгаротоксин (tetramethylrhodamine-α-
этого белка в мембране концевой пластинки.
bungarotoxin; Biotium, США) в концентрации
Аналогичный эффект был обнаружен у мышей
1 мкМ и меченный специфической флуоресцент-
Bla/J [9].
ной меткой BODIPY уабаин (Ouabain-Bodipy;
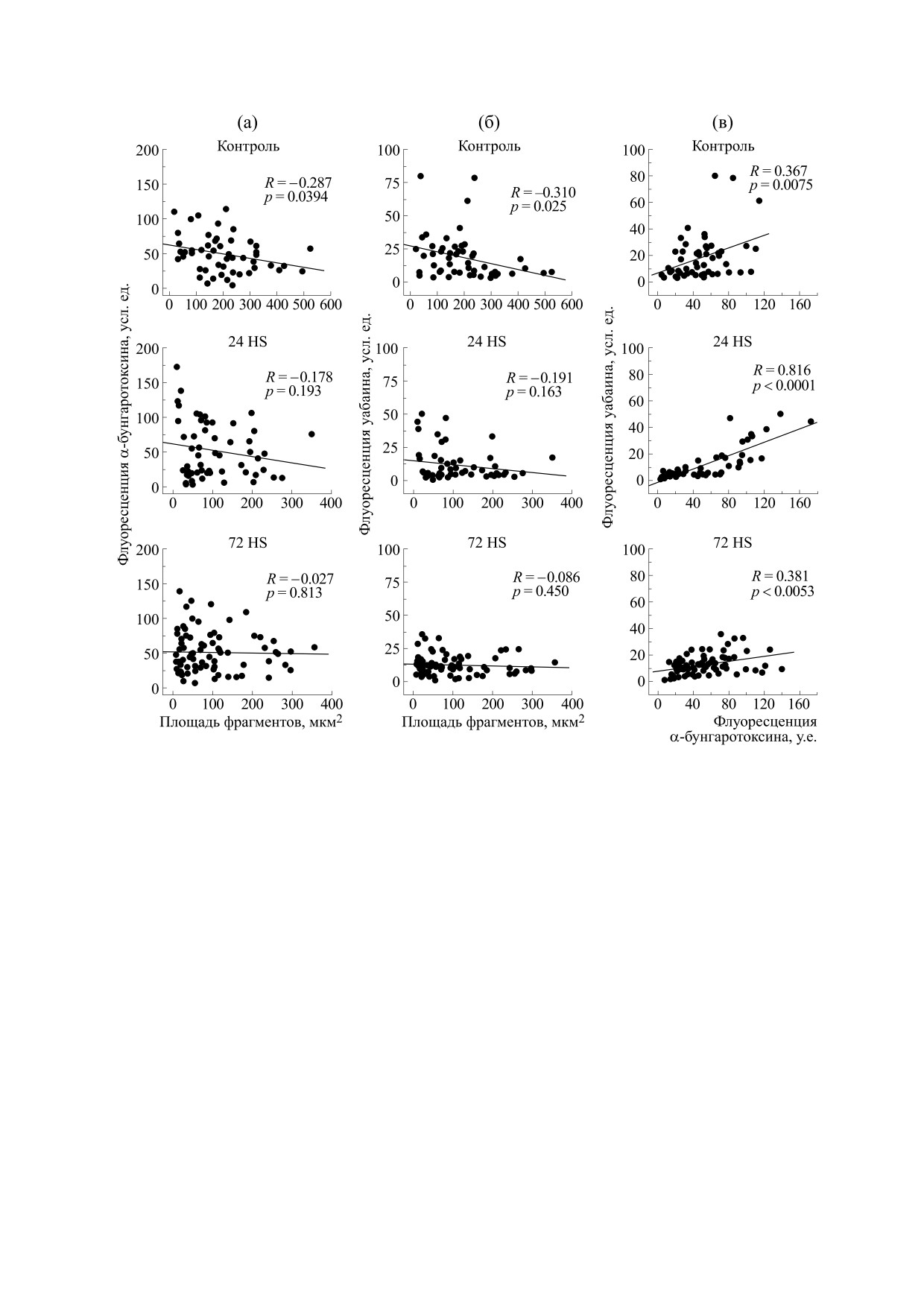
Необходимо отметить, что в контроле наблю-
Invitrogen, США) в концентрации 1 мкМ, как
далась достоверная отрицательная корреляция
описано ранее [15]. Опыты проводили при ком-
между интенсивностью флуоресценции α-бунга-
натной температуре (около 22°С). Для получения
ротоксина в отдельных фрагментах концевых
изображений применяли конфокальную систему
пластинок от площади этих фрагментов (рис. 3а).
Leica TCS SP5 с объективом 63× и числовой апер-
Аналогичная корреляция наблюдалась и для флу-
турой 1.30; обработку изображений проводили с
оресценции уабаина (рис. 3б). Причина суще-
помощью программы ImageJ.
ствования такой корреляционной зависимости
Использовали химреактивы производства Sig-
не ясна, но можно предположить, что эта корре-
ma (США). Статистическую обработку проводи-
ляция отражает соответствующую зависимость
ли с помощью программы Origin Pro 8 (ORIGIN
плотности распределения нХР и α2-изоформы
Software, США) и критериев Стьюдента и
Na,K-АТФазы от размера фрагментов. Через 24 ч
ANOVA. В тексте, таблице и на рисунках приведе-
разгрузки сохранялась лишь тенденция к такой
ны средние значения величин с их ошибками
корреляции, а через 72 ч разгрузки корреляция
(M ± S.E.).
полностью отсутствовала (рис. 3а и 3б). Меха-
низм этого явления неизвестен, можно лишь
предположить, что при двигательной разгрузке
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
происходит не только усиление фрагментации
В контроле были исследованы 32 концевые
концевой пластинки, но и перераспределение
пластинки (9 мышц от шести крыс); через 24 ч
нХР и α2-изоформы Na,K-АТФазы в мембране.
разгрузки - 19 концевых пластинок (7 мышц от
Поскольку локализация белков в плазматической
пяти крыс) и через 72 ч разгрузки - 24 концевые
мембране принципиально зависит от липидного
пластинки (4 мышцы от трех крыс).
окружения, важную роль в такой перестройке мо-
Примеры изображений распределения нХР и
жет играть нарушение структуры липидной
α2-изоформы Na,K-АТФазы в концевых пла-
структуры концевой пластинки, наблюдаемое
стинках в различных условиях показаны на рис. 1.
уже в первые сутки разгрузки [12,16].
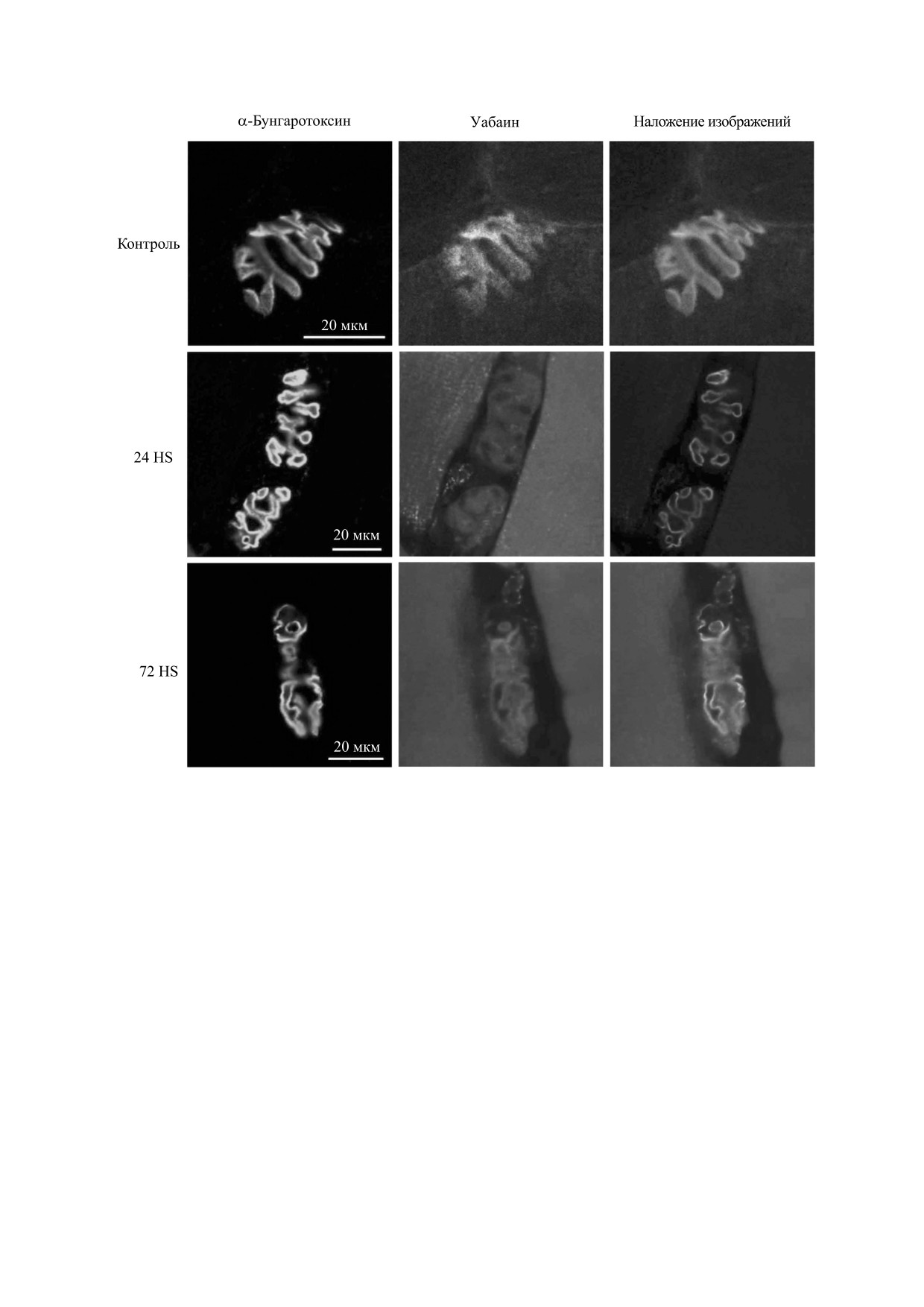
Сигналы от нХР, меченных α-бунгаротоксином,
Необходимо отметить, что мы наблюдали так-
и α2-изоформы Na,K-АТФазы, меченной уабаи-
же высокодостоверную положительную корреля-
ном, были сконцентрированы в области конце-
ционную зависимость между интенсивностью
вой пластинки и совпадали при наложении, что
флуоресценции уабаина и α-бунгаротоксина в
соответствует полученным ранее данным [15]. Та-
отдельных фрагментах (рис. 3в). Аналогичную за-
кая локализация сохранялась и после проведен-
висимость наблюдали и в концевых пластинках
ной разгрузки (рис. 1).
камбаловидных мышц мышей линий C57Bl/6 и
Хорошо известно, что нХР в пределах конце-
Bla/J (одна из моделей дисферлинопатии) [9]. Та-
вой пластинки распределены в виде отдельных
кая корреляционная зависимость сохранялась и
фрагментов [3]. Через 24 и 72 ч разгрузки наблю-
после 24 и 72 ч разгрузки (рис. 3в). Если, как мы
далось достоверное устойчивое усиление фраг-
полагаем, эта корреляция отражает молекулярное
БИОФИЗИКА том 64
№ 5
2019
СТРУКТУРНО-ФУНКЦИОНАЛЬНЫЕ ХАРАКТЕРИСТИКИ КОНЦЕВЫХ ПЛАСТИНОК
959
Рис. 1. Примеры изображений распределения нХР, меченных α-бунгаротоксином, и α2-изоформы Na,K-АТФазы,
меченной уабаином, в концевых пластинках камбаловидных мышц контрольной крысы и крыс через 24 ч и 72 ч
разгрузки.
взаимодействие между нХР и α2-изоформой
ния α2-изоформы Na,K-АТФазы наблюдаются
Na,K-АТФазы, можно предположить, что, не-
во всех случаях. При этом снижение плотности
смотря на возможное перераспределение нХР и
распределения нХР в мембране наблюдается
α2-изоформы в мембране, молекулярное взаимо-
только при хроническом нарушении двигатель-
действие этих белков сохраняется и после раз-
ной активности у мышей mdx и Bla/J [8,9]. Пред-
грузки.
положительно, наблюдаемые изменения являют-
ся результатом адаптационных перестроек в ске-
летной мышце, которые развиваются не только
ЗАКЛЮЧЕНИЕ
при хронических нарушениях, но уже на ранних
Несмотря на отличия в степени нарушений
этапах двигательной дисфункции. Молекуляр-
структурной организации концевых пластинок в
ный механизм такой адаптационной пластично-
мышцах с различной функциональной специали-
сти неизвестен. Одним из возможных регулято-
зацией (диафрагма и камбаловидная мышца мы-
ров является АМФ-активируемая протеинкиназа
шей mdx и Bla/J, как было показано в работах
(AMP-activated protein kinase, AMPK). В скелет-
[8,9]; камбаловидная мышца крысы, как показа-
ной мышце AMPK является ключевым регулято-
но в данной работе), функциональные наруше-
ром мышечного метаболизма, процессов тран-
БИОФИЗИКА том 64
№ 5
2019
960
КРАВЦОВА и др.
Рис. 2. Характеристики концевых пластинок камбаловидной мышцы крысы при двигательной разгрузке (HS - метод
антиортостатического вывешивания) в течение 24 и 72 ч: (а) - среднее количество фрагментов в индивидуальных
концевых пластинках, (б) - средняя площадь фрагментов, (в) - средняя общая площадь концевых пластинок, (г) -
относительная интенсивность флуоресценции (усл. ед.) α-бунгаротоксина (нХР), (д) - относительная интенсивность
флуоресценции (усл. ед.) уабаина (α2-изоформа Na,K-АТФазы); * - р < 0,05, ** - р < 0,01 и *** - р < 0,001 по
сравнению с контролем.
скрипции, аутофагии и многих других [17], при-
ной дисфункции, основанных на знании молеку-
чем уровень AMPK непосредственно связан с
лярных механизмов адаптационной пластично-
мышечной активностью [18]. В камбаловидной
сти скелетной мышцы.
мышце крысы снижение уровня фосфорилирова-
ния AMPK [13,18,19], сопровождающееся сниже-
БЛАГОДАРНОСТИ
нием электрогенной активности α2-изоформы
Na,K-АТФазы [11], наблюдается уже в первые
Авторы выражают благодарность Ресурсному
сутки двигательной разгрузки. Важно отметить,
центру «Развитие молекулярных и клеточных тех-
что в скелетной мышце AMPK влияет также и на
нологий» Санкт-Петербургского государствен-
активность Na,K-АТФазы [20,21]. AMРK можно
ного университета и лично Н.А. Костину за по-
рассматривать также в качестве важного фактора
мощь в проведении работ с конфокальной мик-
поддержания структурно-функциональной орга-
роскопией.
низации нервно-мышечного соединения. Так,
фармакологическая активация АМРК с помощью
ФИНАНСИРОВАНИЕ РАБОТЫ
препарата AICAR (5-aminoimidazole-4-carboxam-
ide ribonucleotide) препятствует структурным на-
Работа выполнена при финансовой поддержке
рушениям нервно-мышечного соединения и раз-
Российского научного фонда (грант № 18-15-
витию симптомов мышечной атрофии [19,22].
00043).
Превентивное введение AICAR способно также
стабилизировать структуру концевых пластинок
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
и предотвращать деполяризацию мембраны, вы-
зываемую двигательной разгрузкой [23]. Полу-
Все применимые международные, националь-
ченные нами новые факты могут быть полезными
ные и институциональные принципы ухода и ис-
с точки зрения поиска путей коррекции мышеч-
пользования животных при выполнении работы
БИОФИЗИКА том 64
№ 5
2019
СТРУКТУРНО-ФУНКЦИОНАЛЬНЫЕ ХАРАКТЕРИСТИКИ КОНЦЕВЫХ ПЛАСТИНОК
961
Рис. 3. Корреляционные зависимости интенсивности флуоресценции α-бунгаротоксина (усл. ед., нХР) и уабаина
(усл. ед., α2-изоформа Na,K-АТФазы) в отдельных фрагментах концевых пластинок камбаловидной мышцы крысы
при двигательной разгрузке (HS - метод антиортостатического вывешивания) в течение 24 и 72 ч. Представлены за-
висимости интенсивности флуоресценции α-бунгаротоксина (а) и уабаина (б) от площади фрагментов; взаимозави-
симость интенсивности флуоресценции уабаина и α-бунгаротоксина в отдельных фрагментах (в).
были соблюдены. Условия содержания животных
5. P. Yampolsky, P. G. Pacifici, and V. Witzemann, Eur. J.
и приемы работы с ними соответствовали нормам
Neurosci. 31, 646 (2010).
международного и российского законодатель-
6. M. R. Deschenes and M. H. Wilson, J. Neurobiol. 57,
ства.
246 (2003).
7. E. M. van der Pijl, M. van Putten, E. H. Niks, et al.,
СПИСОК ЛИТЕРАТУРЫ
Eur. J. Neurosci. 43, 1623 (2016).
1. S. J. Wood and C. R. Slater, Prog. Neurobiol. 64, 393
8. V. V. Kravtsova, N. A. Timonina, G. F. Zakir'yanova,
(2001).
et al., Neurochem. J.
12,
305
(2018). DOI:
10.1134/S1819712418040049.
2. C. R. Slater, Int. J. Mol. Sci. 18 (10), pii: E2183 (2017).
DOI: 10.3390/ijms18102183.
9. V. V. Kravtsova, E. V. Bouzinova, V. V. Machkov, et al.,
Bull. Exp. Biol. Med.
166,
593
(2019). DOI:
3. S. Willadt, M. Nash, and C. R. Slater, Sci. Rep. 6,
10.1007/s10517-019-04398-z.
24849 (2016). DOI: 10.1038/srep24849.
4. I. I. Krivoi and A. M. Petrov, Int. J. Mol. Sci. 20, 1046
10. D. J. Falk, A. G. Todd, S. Lee, et al., Hum. Mol. Gen-
(2019). DOI: 10.3390/ijms20051046.
et. 24, 625 (2015).
БИОФИЗИКА том 64
№ 5
2019
962
КРАВЦОВА и др.
11. V. V. Kravtsova, A. M. Petrov, V. V. Matchkov, et al., J.
18. N. A. Vilchinskaya, I. I. Krivoi, and B. S. Shenkman,
Gen. Physiol.
147,
175
(2016). DOI:
10.1085/
Int. J. Mol. Sci. 19, 3558 (2018). DOI: 10.3390/
jgp.201511494.
ijms19113558.
12. A. M. Petrov, V. V. Kravtsova, V. V. Matchkov, et al.,
19. N. A. Vilchinskaya, E. P. Mochalova, T. L. Nemi-
Am. J. Physiol. Cell Physiol. 312, C627 (2017). DOI:
rovskaya, et al., J. Physiol. 595, 7123 (2017). DOI:
10.1152/ajpcell.00365.2016.
10.1113/JP275184.
13. A. V. Chibalin, B. Benziane, G. F. Zakyrjanova, et al.,
J. Cell. Physiol. 233, 6329 (2018). DOI: 10.1002/
20. M. S. Ingwersen, M. Kristensen, H. Pilegaard, et al., J.
jcp.26594.
Membrane Biol. 242, 1 (2011). DOI: 10.1007/s00232-
011-9365-7.
14. E. Morey-Holton, R. K. Globus, A. Kaplansky, and G.
Durnova, Adv. Space Biol. Med. 10, 7 (2005).
21. B. Benziane, M. Bjornholm, S. Pirkmajer, et al., J.
15. J. A. Heiny, V. V. Kravtsova, F. Mandel, et al., J. Biol.
Biol. Chem.
287,
23451
(2012). DOI:
10.1074/
Chem.
285,
28614
(2010).
DOI:
10.1074/
jbc.M111.331926.
jbc.M110.150961.
22. C. Cerveró, N. Montull, O. Tarabal, et al., Neurother-
16. I. G. Bryndina, M. N. Shalagina, A. V. Sekunov, et al.,
apeutics 13, 198 (2016). DOI: 10.1007/s13311-015-
Neurosci. Lett.
664,
1
(2018). DOI:
10.1016/
0399-x.
j.neulet.2017.11.009.
17. D. G. Hardie, B. E. Schaffer, and A. Brunet, Trends
23. В. В. Кравцова, Н. А. Вильчинская, В. Л. Розломий
Cell. Biol.
26,
190
(2016).
DOI:
10.1016/
и др., Биохимия 84, 1335 (2019). DOI: 10.1134/
j.tcb.2015.10.013.
S0320972519090112.
Structural and Functional Characteristic Features of Rat Soleus Endplates
under Short-Term Disruption of Motor Activity
V.V. Kravtsova, E.A. Saburova, and I.I. Krivoi
St. Petersburg State University, Universitetskaya nab. 7/9, St. Petersburg, 199034 Russia
In this study, we analyzed distribution of nicotinic acetylcholine receptors, the degree of fragmentation of
endplates and their area, and membrane localization of the Na,K-ATPase α2 isoform in the rat soleus muscle
after functional unloading (the method of hindlimb suspension) for 1-3 days. After unloading, increased
endplate fragmentation, accompanied by a decrease in the area of individual fragments was observed; the total
area of the endplates was not changed. The distribution density of nicotinic acetylcholine receptors also re-
mained unaltered, however, a significant decrease in the distribution density of the Na,K-ATPase α2 isoform
was observed. Presumably, identified changes resulted from adaptive skeletal muscle remodeling, developing
not only under chronic disorders, but even in the early stages of motor dysfunction.
Keywords: motor endplate, nicotinic acetylcholine receptor, Na,K-ATPase isoforms, motor activity
БИОФИЗИКА том 64
№ 5
2019