БИОФИЗИКА, 2019, том 64, № 5, с. 974-981
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 576.5
РИТМОИНОТРОПНЫЕ ОТНОШЕНИЯ В ПАПИЛЛЯРНОЙ МЫШЦЕ
СЕРДЦА СУСЛИКА КАК ОТРАЖЕНИЕ ИЗМЕНЕНИЙ
ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ ЖИВОТНЫХ В ТЕЧЕНИЕ
ГОДОВОГО ЦИКЛА
© 2019 г. О.В. Накипова, А.С. Аверин, Л.С. Косарский, Д.А. Игнатьев
Институт биофизики клетки Российской академии наук - обособленное подразделение ФИЦ «Пущинский научный
центр биологических исследований РАН»,
142290, Пущино Московской области, ул. Институтская, 3
Поступила в редакцию 28.06.2019 г.
После доработки 28.06.2019 г.
Принята к публикации 08.07.2019 г.
Проведено исследование особенностей зависимости силы сокрашения (изометрический режим,
30°С) от частоты стимуляции (0,1-4,0 Гц) в папиллярных мышцах правого желудочка сердца сусли-
ка Spermophilus undulatus в группах животных летнего (июнь-июль) и осеннего (октябрь-ноябрь)
периодов, а также периода гибернации, включающих животных, активных между баутами спячки;
входящих в спячку (Tсердца ~ 30°С); спящих и пробуждающихся (Tсердца ~ 30°С). Показано, что вне
зависимости от функционального состояния животных, характер зависимости силы сокращения от
частоты стимуляции (ритмоинотропия) в папиллярных мышцах сердца суслика в основных прояв-
лениях относится к отрицательному типу: сила сокращения на высоких частотах (от 1,0 до 4,0 Гц)
всегда меньше ее величины на низких частотах (в области 0,1 Гц). При этом в каждой группе живот-
ных можно выделить два типа ритмоинотропии: с относительно слабой (тип 1) и с выраженной (тип
2) частотной зависимостью. В обоих типах ритмоинотропии может присутствовать слабая положи-
тельная компонента (обусловленная ростом силы сокращения): у типа 1 - в области частот стиму-
ляции от 0,2 до 0,5 Гц (она появляется только у животных периода гибернации - активных между
баутами и спящих или входящих в состояние спячки, т.е. стремящихся к снижению активности), а
у типа 2 - в области высоких частот стимуляции (свыше 1,0 Гц), с наибольшей выраженностью у
животных периодов возобновления функциональной активности - пробуждающихся в сезон спяч-
ки и у весенних при окончательном пробуждении. Полученные данные указывают на то, что про-
цесс формирования определенного типа ритмоинотропии в сердце зимоспящих сусликов предше-
ствует изменениям функционального состояния животных в течение годового цикла.
Ключевые слова: сердце, папиллярные мышцы, сократимость, ритмоинотропные отношения, гиберни-
рующие животные, суслик.
DOI: 10.1134/S0006302919050193
трансформируется при изменениях температуры
Частота сердцебиений является жизненно-
и в условиях патологий [8,10-14]. В сердце боль-
важным модулятором функции сердца и относит-
ся к разновидности врожденных механизмов,
шинства видов млекопитающих в области частот
впервые описанных H.P. Bowditch в 1871 г. [1], ин-
стимуляции от 0,5 до 4,0 Гц в норме зависимость
терес к изучению которых до сих пор не потерял
«частота - сила» (ЧС) положительная: увеличе-
свою актуальность. Зависимость силы сокраще-
ние частоты стимуляции приводит к значитель-
ния от частоты стимуляции (ритмоинотропные
ному увеличению силы сокращений. При патоло-
отношения) представляет собой важный меха-
гии зависимость ЧС приобретает на начальных
низм в сердце животных и человека, обеспечива-
стадиях двухфазную форму: слабо положитель-
ющий его способность увеличивать силу сокра-
ную в области низких частот стимуляции от 0,5 до
щения и сердечный выброс [2-9]. Известно, что
1,5 Гц, переходящую в отрицательную при более
характер ритмоинотропии зависит от вида и воз-
высоких частотах (до 4,0 Гц). По мере углубления
раста животных, типа миокардиальной ткани,
процесса, в частности в условиях сердечной недо-
статочности, зависимость ЧС становится полно-
Сокращения: зависимость ЧС - зависимость «частота -
сила».
стью отрицательной [8,9,13,14-16]. В сердце крыс
974
РИТМОИНОТРОПНЫЕ ОТНОШЕНИЯ В ПАПИЛЛЯРНОЙ МЫШЦЕ СЕРДЦА СУСЛИКА
975
и мышей зависимость ЧС также имеет двухфаз-
ный характер [25], чрезвычайно сходный с выше-
ный характер, однако с принципиально иной ча-
описанным для условий патологии у высших мле-
стотной зависимостью: выраженной отрицатель-
копитающих и человека [4,14-16,26-29]. Это и
ный в области низких частот стимуляции (до
определило наш интерес к более детальному изу-
1,0 Гц) и слабо положительный - в области высо-
чению особенностей ритмоинотропных отноше-
ких частот (1,0-4,0 Гц) [1,2,17,18]. При изучении
ний в сердце типичного представителя гетеро-
механизмов патологических изменений в мио-
термных млекопитающих - якутского суслика
карде особый интерес исследователей привлека-
S. undulatus. Гибернационный период зимоспя-
ют животные, сердце которых имеет отличный от
щих, длящийся с середины октября по март, не
положительного характер ритмоинотропии, в си-
является непрерывным процессом, он состоит из
лу чего крысы и мыши оказались наиболее часто
многократно повторяющихся баутов спячки, раз-
используемыми в сравнительных исследованиях.
деляемых короткими периодами активности (от
12 до 14 ч). Баут спячки включает в себя процесс
Интерес к изучению особенностей ритмоино-
вхождения в спячку, собственно спячку и про-
тропии у зимоспящих животных был иницииро-
цесс пробуждения, сопровождающийся гигант-
ван в середине 1980-х годов работами [19,20], ав-
ским изменением температуры тела (от
1°С
торы которых впервые исследовали характер за-
до 36°С) и частоты сердечных сокращений (от 3-
висимости ЧС в сердце бурундука (Tamias
5 ударов в минуту до 360-420 ударов в минуту)
sibiricus) и обнаружили, что в диапазоне частоты
[30-39]. В настоящей работе были исследованы
стимуляции 0,5-2,0 Гц у активных животных лет-
шесть групп сусликов - активные животные лет-
него периода она имеет положительную направ-
него, осеннего и зимнего периодов, спящие жи-
ленность, а у спящих становится отрицательной,
вотные, а также животные, входящие в состояние
подобно патологическому миокарду. Этот факт
спячки и пробуждающиеся в период гибернации.
впервые указал на возможность использования
Вход в спячку был исследован на его начальном
сердца зимоспящих животных в качестве модели
этапе, а пробуждение - на завершающем, но и в
для изучения механизмов, лежащих в основе
том и в другом случае при температуре сердца
формирования отрицательного типа ритмоино-
вблизи 30°С. Таким образом в настоящей работе,
тропии. Попытки ряда исследователей повторить
наряду с исследованиями стационарных состоя-
опыты на зимоспящих животных, предпринятые
ний летней активности и зимнего оцепенения,
в 1980-х годах, дали крайне неоднозначные ре-
было уделено особое внимание переходным со-
зультаты [21,22]. Тем не менее к началу наших ис-
стояниям вхождения животных в спячку и выхода
следований сохранилось представление о том,
из нее.
что зависимость ЧС сердца зимоспящих живот-
ных в активном летом состоянии подобна обыч-
ным млекопитающим и имеет положительную
МАТЕРИАЛЫ И МЕТОДЫ
направленность, а в состоянии зимнего оцепене-
ния - становится отрицательной.
Исследования проводили на папиллярных
мышцах правого желудочка сердца якутских сус-
Уже первые исследования, проведенные нами
ликов Spermophilus undulatus в шести группах жи-
на сусликах Spermophilus undulatus, внесли суще-
вотных: летнего (июнь-июль, n = 19) и осеннего
ственную коррекцию в эти первоначальные пред-
(октябрь-ноябрь, n = 16) периодов, а также пери-
ставления. Мы не обнаружили в чистом виде вы-
ода гибернации, включающего активных между
раженного положительного типа ритмоинотро-
баутами спячки (через 12 ч после пробуждения с
пии в сердце суслика в диапазоне частот
частотой сердечных сокращений 150-200 уд/мин,
стимуляции от 0,1 до 1,0 Гц, оказавшейся отрица-
Tтела
~
36°C, n
= 12); входящих в спячку
тельной не только у спящих, но и у активных жи-
(Tсердца 30°С, n = 7); спящих (на шестые сутки
вотных как летнего, так и зимнего периодов [23-
спячки с частотой сердечных сокращений 3-
25]. Тем не менее было обнаружено принципи-
5 уд/мин, Tтела ~ 4-6°С, n = 8); искусственно про-
альное иное различие в проявлении ритмоино-
бужденных (Tсердца ~30°С, n = 7).
тропии сердца сусликов сезона гибернации и лет-
него активного периода. Оно заключалось в том,
Активных животных предварительно наркоти-
что у сусликов, находившихся в состоянии глубо-
зировали эфиром. Выделенное сердце помещали
кого оцепенения, как и у этих же животных, бодр-
в раствор Тироде (20°С) следующего состава (в
ствующих в короткий период пробуждения меж-
мМ): Na+ - 150; K+ - 4,0; Ca2+ - 1,8; Mg2+ - 1,0;
ду баутами спячки, отмечалось появление поло-
HCO2- - 13,0; HPO4- - 1,8; Cl- - 148,4, глюкоза -
жительной компоненты в области низких частот
стимуляции (от 0,1 до 0,5 Гц) при сохранении ее
11,0, рН 7,4. Раствор аэрировали газовой смесью
отрицательной направленности в диапазоне до
О2 (96%) + СО2 (4%). Выделение папиллярных
1,0 Гц, что придавало зависимости ЧС двухфаз-
мышц, стимуляцию и измерение амплитуды со-
БИОФИЗИКА том 64
№ 5
2019
976
НАКИПОВА и др.
кращения в изометрическом режиме проводили
пии значительно отличаются по величине спада
по ранее описанной методике [23] при температу-
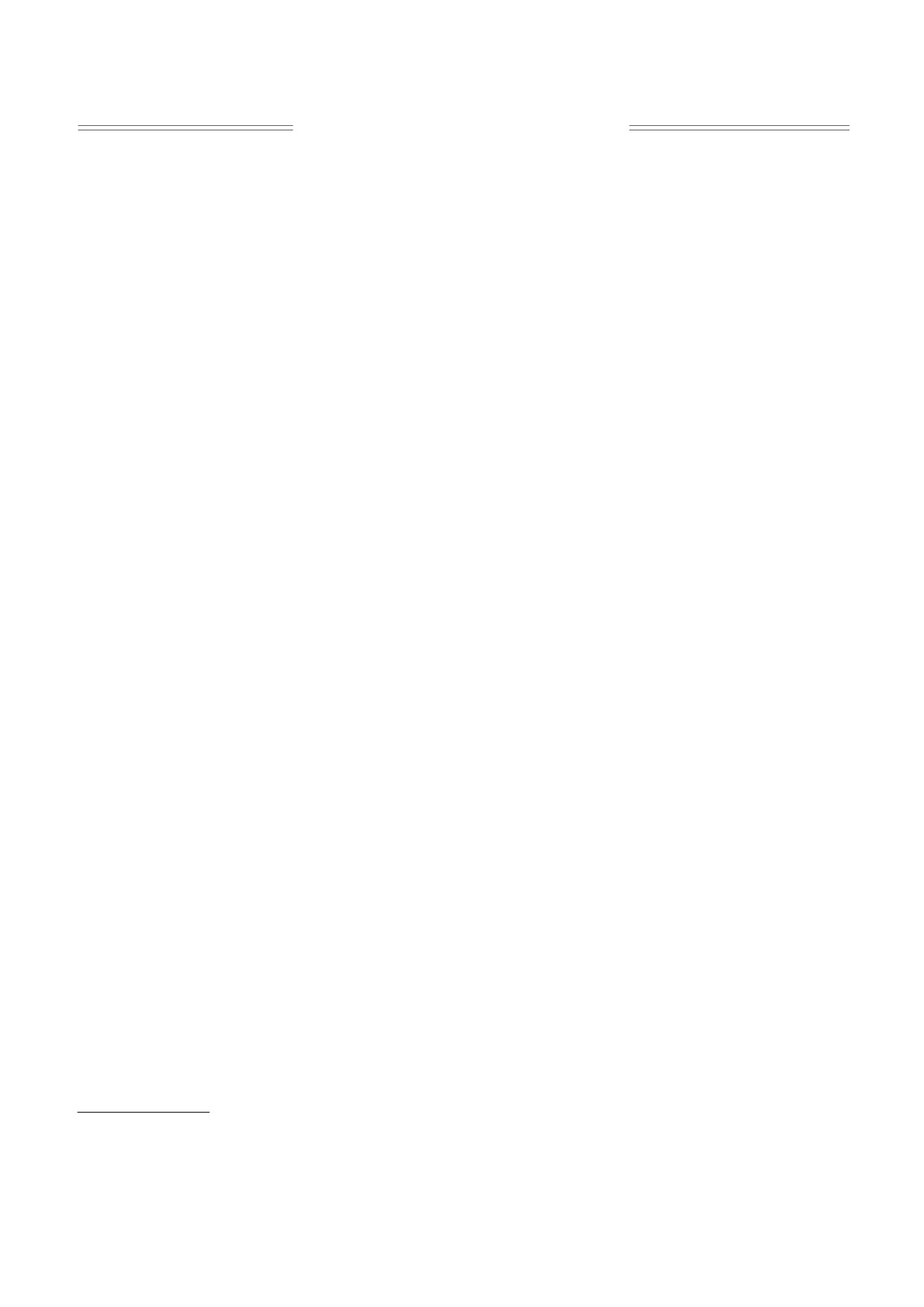
(ΔF) силы сокращения при увеличении частоты
ре перфузирующего раствора 30 ± 1°С. Диаметр
стимуляции от 0,1 до 0,5 Гц. Зависимость ЧС с ме-
папиллярных мышц варьировал от 0,6 до 1,0 мм, а
нее выраженным наклоном кривой (ΔF ≤ 40%)
длина - от 1,0 до 3,0 мм. Для исследования их ме-
обозначена нами как тип 1; зависимость с более
ханической активности использовали стандарт-
выраженной крутизной спада (ΔF ≥ 50%) - как
ную автоматизированную установку на основе
тип 2.
персонального компьютера и плат АЦП-ЦАП
Другим отличительным признаком выделен-
(L-Сard 154 и L-Card E14-440). Механическую
ных типов ритмоинотропии является наличие
активность мышц регистрировали с помощью
слабой положительной компоненты (обуслов-
механотрона 6Х-2М. В начале каждого опыта
ленной ростом силы сокращения) в узкой обла-
препарат стимулировали прямоугольными им-
сти частот стимуляции, местоположение которой
пульсами частотой 0,3 Гц в течение 1 ч.
меняется в зависимости от сезона активности и
Стационарную зависимость «частота-сила» в
функционального состояния животных.
изометрическом режиме регистрировали в интер-
У сусликов с типом 1 ритмоинотропии в осен-
вале частот стимуляции от 0,01 до 4,0 Гц. Силу со-
ний период отмечается появление положитель-
кращения F папиллярной мышцы определяли
ной компоненты в области низких частот стиму-
как отношение силы изометрического сокраще-
ляции (от 0,2 до 0,5 Гц), которая сохраняется в те-
ния к поперечному сечению несокращающейся
чение всего периода гибернации (с октября по
мышцы. При построении зависимости ЧС вели-
март). Для сусликов с типом 2 ритмоинотропии
чину стационарной амплитуды для каждой из ча-
характерно наличие положительной компоненты
стот стимуляции в исследуемом диапазоне выра-
в области высоких частот стимуляции (свыше
жали в процентах по отношению к ее величине на
1,0 Гц), наиболее выраженной в период весеннего
частоте стимуляции 0,1 Гц, принимаемой за
пробуждения. В папиллярных мышцах сердца
100%.
спящих сусликов при 30°С во всех случаях зареги-
Достоверность полученных результатов оце-
стрирован первый тип ритмоинотропии со сла-
нивали с помощью пакета программ Statistica 6.0
бой положительной компонентой в области низ-
(по уровню значимости p < 0,05). Данные пред-
ких частот стимуляции (выделено пунктиром на
ставлены как средние значения ± стандартная
рис. 1), сходный с первым типом ритмоинотро-
ошибка среднего.
пии в папиллярных мышцах активных сусликов.
Другим отличительным признаком выделен-
ных типов ритмоинотропии является наличие у
РЕЗУЛЬТАТЫ
второго типа слабой положительной компоненты
Известно, что физиологический диапазон ча-
в области частот стимуляции свыше 0,5 Гц. По-
стот для сердца суслика варьирует в широких пре-
добный тип ритмоинотропии практически нико-
делах - от 3-5 уд/мин (в спячке, при ректальной
гда не регистрируется у спящих и активных между
температуре 1-4°С) до 360-420 уд/мин (в процес-
баутами сусликов (рис. 1в).
се пробуждения в сезон спячки) [30,31,40-43]. На
Обращает на себя внимание то, что в группах
рис. 1 в сравнительном плане представлены ре-
летних и осенних сусликов количество животных
зультаты исследований зависимости силы сокра-
с разными типами ритмоинотропии представле-
щения от частоты стимуляции (в диапазоне, охва-
но примерно поровну. А в группах сусликов пери-
тывающем физиологический - от 0,1 до 4,0 Гц),
ода гибернации преобладает первый тип, незави-
проведенных на папиллярных мышцах сердца
симо от функционального состояния животных
сусликов летнего (июнь - начало июли), осенне-
(как у активных между баутами, так и у спящих)
го (конец сентября - начало ноября) и зимнего
(рис. 1в). В папиллярных мышцах сердца спящих
(декабрь - март) периодов. Отметим, что вне за-
сусликов при 30°С во всех случаях зарегистриро-
висимости от функционального состояния жи-
ван первый тип ритмоинотропии со слабой поло-
вотных характер зависимости ЧС в папиллярной
жительной компонентой в области низких частот
мышце сердца суслика в основных проявлениях
стимуляции (выделено «пунктиром» на рис. 1в),
относится к отрицательному типу - сила сокра-
сходный с первым типом ритмоинотропии в па-
щения на высоких частотах (от 1,0 до 4,0 Гц) все-
пиллярных мышцах активных между баутами
гда меньше ее величины на низких частотах (в об-
сусликов в период глубокой спячки.
ласти 0,1 Гц). При этом в каждой функциональ-
ной группе животных можно выделить два типа
Второй тип ритмоинотропии начинает появ-
зависимости ЧС, имеющих ряд отличительных
ляться у межбаутных активных животных только
особенностей. Как следует из графиков, пред-
в конце февраля - начале марта, т.е. к окончанию
ставленных на рис. 1, разные типы ритмоинотро-
сезона гибернации (рис. 1в). В этот же период он
БИОФИЗИКА том 64
№ 5
2019
РИТМОИНОТРОПНЫЕ ОТНОШЕНИЯ В ПАПИЛЛЯРНОЙ МЫШЦЕ СЕРДЦА СУСЛИКА
977
Рис. 1. Статистические кривые двух типов зависимости силы сокращения от частоты стимуляции в папиллярных
мышцах сердца сусликов: (а) - активных летних (n = 19); (б) - активных осенних (n = 16); (в) - животных в период
гибернации, активных между баутами спячки (n = 12) и спящих (пунктирная линия, n = 8) (30°С, 1,8 мМ Са2+). По оси
ординат - сила стационарного изометрического сокращения в % по отношению к частоте стимуляции на 0,1 Гц, при-
нимаемой за 100%; по оси абсцисс - частота стимуляции, Гц. Данные представлены как средние значения ± ошибка
среднего (* - достоверное отличие от значения при частоте стимуляции 0,1 Гц, P < 0,05).
может появляться и у спящих животных (данные
рис. 1в). В папиллярных мышцах пробуждаю-
не приводятся).
щихся животных (рис. 2б) зависимость ЧС имеет
двухфазный характер, сходный с описанным вы-
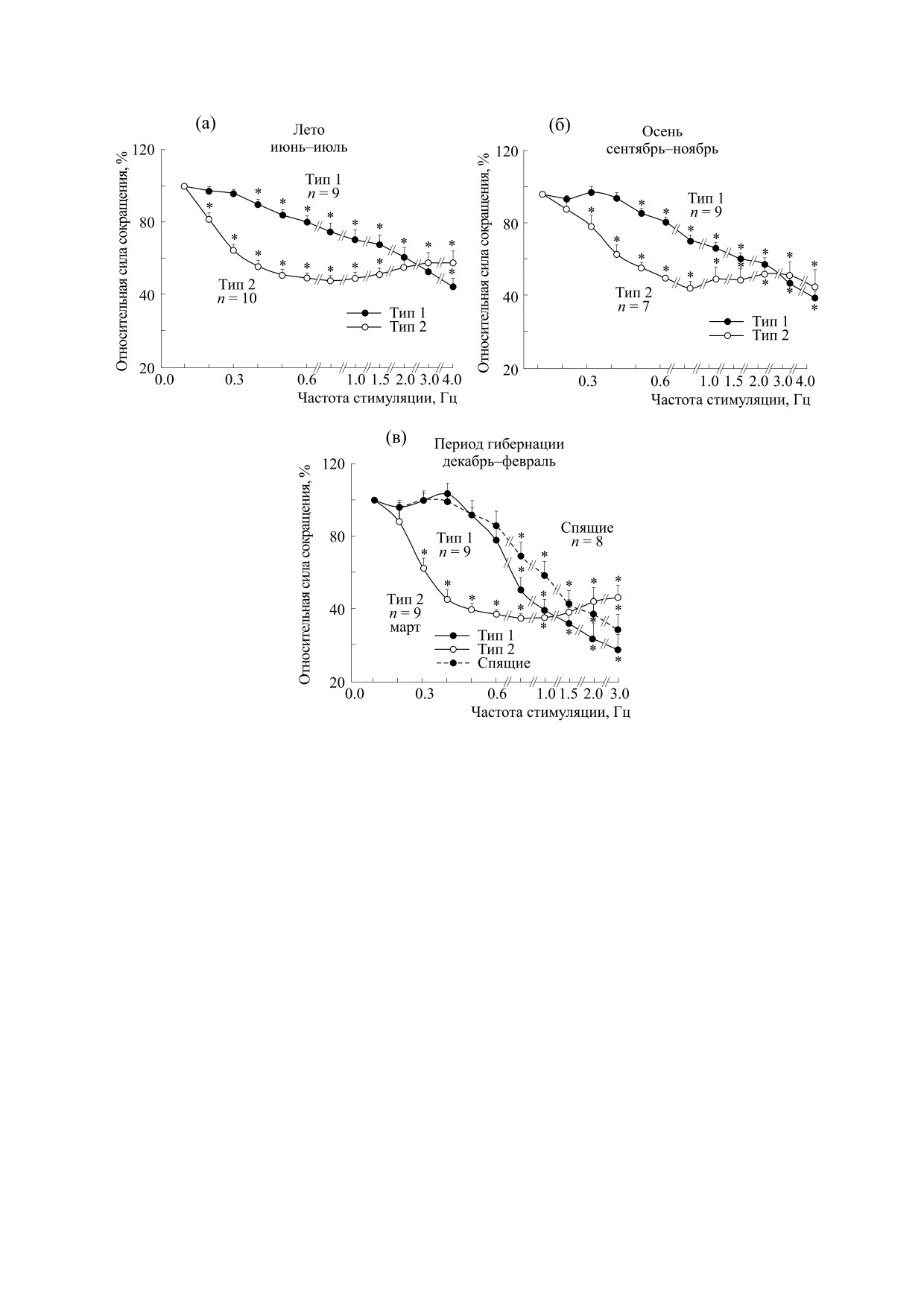
На рис. 2 представлены типы зависимости ЧС,
ше для папиллярных мышц со вторым типом рит-
зарегистрированные в папиллярных мышцах жи-
моинотропии (рис. 1).
вотных на начальных стадиях вхождения в спячку
(рис. 2а) и на поздних стадиях пробуждения
(рис. 2б) при температуре сердца, в том и в другом
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
случае близкой к 30°С. Можно видеть, что у сус-
ликов, входящих в состояние спячки, зависи-
На момент начала наших исследований счита-
мость ЧС имеет отрицательную направленность
лось, что зависимость ЧС в сердце зимоспящих
со слабой положительной компонентой в области
животных, как у большинства млекопитающих в
частот стимуляции от 0,1 до 0,4 Гц (рис. 2а), что
норме, имеет положительную направленность и
позволяет отнести ее к первому типу, свойствен-
только в состоянии спячки становится отрица-
ному животным периода гибернации (см.
тельной [19,20]. По результатам наших ранее
БИОФИЗИКА том 64
№ 5
2019
978
НАКИПОВА и др.
Рис. 2. Зависимости силы сокращения от частоты стимуляции в папиллярных мышцах сердца сусликов: (а) -
входящих в состояние спячки (n = 6), (б) - пробуждающихся (n = 6) при температуре сердца, близкой в обоих случаях
к 30°С. Данные представлены, как средние значения ± стандартная ошибка среднего (* - достоверное отличие от
значения при частоте стимуляции 0,1 Гц, P < 0,05).
опубликованных работ, выполненных в диапазо-
Первый тип ритмоинотропии имеет преимуще-
не частот стимуляции от 0,1 до 1,0 Гц, зависи-
ственно отрицательную направленность во всей
мость ЧС в папиллярных мышцах сердца сусли-
области исследованных частот, а второй тип -
ков, независимо от их функционального состоя-
двухфазную: резко-отрицательную в области низ-
ния, имеет отрицательную направленность [23-
ких частот стимуляции и положительную в области
25], что согласуется с результатами недавно опуб-
высоких частот (свыше 1,0 Гц). По внешним про-
ликованной работы [44], выполненной на изоли-
явлениям обнаруженный нами в сердце суслика
рованном сердце тринадцатиполосного суслика
первый тип ритмоинотропии (с менее выраженной
(Ictidomys tridecemlineatus), перфузируемом по
частотной зависимостью) сходен с особенностями
Лангендорфу. Авторы данной работы зарегистри-
ритмонотропии миокарда обычных млекопитаю-
ровали в сердце как летних активных, так и зим-
щих и человека при сердечной недостаточности
них спящих животных (при 37°С) снижение со-
[8,9,13-16]. Сердце суслика с двухфазным типом
кратимости в ответ на увеличение частоты стиму-
ритмоинотропии (тип 2) обнаруживает сходство с
ляции от 4 до 10 Гц.
особенностями сердца крыс и мышей [1,2,17,18].
Принципиально новым результатом нашей
Основной вопрос, который возникает при
работы является впервые обнаруженное нами в
анализе полученных результатов, касается при-
сердце активных сусликов разных сезонных пе-
чин наличия двух типов ритмоинотропии у жи-
риодов наличие двух типов ритмоинотропии с от-
вотных летнего и осеннего периодов. В отноше-
носительно слабой и выраженной частотной за-
нии осеннего периода ситуация представляется
висимостью в диапазоне от 0,1 до 1,0 Гц [25]. Ис-
достаточно понятной. Известно, что подготовка к
следование более широкого диапазона частот (от
спячке у зимоспящих начинается задолго до на-
0,1 до 4 Гц) в настоящей работе подтвердило на-
ступления периода гибернации. В течение октяб-
личие двух типов ритмоинотропии в группах лет-
ря и до середины ноября суслики входят в состо-
них и осенних животных и выявило во втором ти-
яние так называемой «пробной спячки» (перио-
пе ритмоинотропии ранее не обнаруженное нами
дически впадают в оцепенение на двое-трое
наличие положительной компоненты в диапазо-
суток, температура их тела при этом снижается
не частоты стимуляции от 1 до 4 Гц. При более
незначительно) [31,33,37-39]. Считается, что та-
высоких частотах, свыше 4 Гц, зависимость ста-
кие кратковременные погружения в спячку сти-
новилась отрицательной, как и в работе [44].
мулируют перестройку кальциевого гомеостаза
Таким образом, новые данные, подтверждая ос-
миокардиальных клеток, играющего ключевую
новные выводы наших ранее выполненных ра-
роль в адаптационных изменениях сердца гибер-
бот, вносят существенные дополнения в пред-
нирующих животных [29,45-50] и в процессах
ставления о феноменологических особеннотях
трасформации характера ритмоинотропии в
ритмоинотропных явлений в сердце зимоспящих
условиях патологий у высших млекопитающих и
животных.
человека [6,9,14-16]. Можно предположить, что
БИОФИЗИКА том 64
№ 5
2019
РИТМОИНОТРОПНЫЕ ОТНОШЕНИЯ В ПАПИЛЛЯРНОЙ МЫШЦЕ СЕРДЦА СУСЛИКА
979
этот процесс является одной из причин наличия
Суммируя данные проведенных нами исследо-
двух типов ритмоинотропии у животных осенне-
ваний, можно заключить, что появление слабо-
го периода и появления положительной компо-
выраженной положительной компоненты зави-
ненты зависимости ЧС в области низких частот
симости в области низких частот стимуляции яв-
стимуляции, наиболее физиологичных для пери-
ляется отличительным признаком животных
ода зимней спячки.
периода гибернации и входящих в состояние
спячки (стремящихся к снижению активности).
Наличие двух типов римоинотропии (рис. 1а)
Напротив, наличие положительной компо-
в группе летних сусликов, находящихся вне ги-
ненты в области высоких частот стимуляции яв-
бернационного периода с его сложными функци-
ляется отличительным признаком переходных
ональными перестройками, на первый взгляд, ка-
периодов, сопровождающихся повышением
жется странным. Однако, по нашим наблюдени-
функциональной активности животных. Поэто-
ям, в жаркое лето часть сусликов в условиях
му в наиболее ярком виде особенности, свой-
вивария способна входить в сонное состояние, с
ственные второму типу ритмоинотропии, прояв-
понижением температуры до 25-26°С. Легкая
ляются у активных животных по мере приближе-
провокация пробуждает животных, инициируя
ния сезона весеннего пробуждения, самого
повышение температуры тела в течение
30-
сложного в жизни зимоспящих животных.
35 мин до 36-37°С [31,40-43]. Не исключено, что
первый тип ритмоинотропии у летних животных
В плане установления возможных механиз-
(рис. 1а), напоминающий таковой у животных
мов, лежащих в основе формирования различных
периода гибернации (рис. 1в), может быть свой-
типов римоинотропии, обнаруженных нами в
ственен сусликам, находящимся в состоянии лет-
сердце суслика, наиболее актуальным представ-
ней спячки или входящим в нее. С подобной воз-
ляется изучение роли в этом процессе различных
можностью согласуются и литературные данные,
Са2+-транспортирующих систем, соотношение
указывающие на то, что ряд гетеротермных жи-
активности которых меняется в процессе измене-
вотных [51,52], в том числе и суслики [53], при
ния функционального состояния зимоспящих
летней жаре в сухие сезоны действительно спо-
[45-50,55] и вносит определяющий вклад в фор-
собны впадать в летнюю спячку (эстивацию).
мирование разных типов ритмоинотропии в нор-
ме и их трансформацию при патологиях у обыч-
Следует отметить, что при внешнем сходстве
ных млекопитающих и человека
[1,2,6-9,14-
первого типа ритмоинотропии сусликов летнего
16,19-22,55]. Выяснение механизмов физиологи-
и гибернационного периода у последних суще-
ческой трансформации характера ритмоинотро-
ствует одна важная особенность - наличие фазы
пии в течение годового цикла животных гибер-
повышения силы сокращений (положительной
нантов может иметь важное значение для
компоненты зависимости) в узкой полосе частот
понимания механизмов патологической транс-
от 0,1 до 0,4 Гц (рис. 1б,в). Таким образом, эта
формации ритмоинотропных отношений в усло-
особенность наблюдается только у сусликов,
виях сердечной недостаточности и для разработ-
спячка которых проходит при отрицательной
ки эффективных путей преодоления этой опас-
температуре внешней среды и проявляется толь-
ной патологии.
ко поздней осенью и зимой; в спячке летних жи-
вотных это характерное явление не отмечается.
ФИНАНСИРОВАНИЕ РАБОТЫ
Напротив, наличие положительной компо-
ненты в области высоких частот стимуляции, ха-
Работа выполнена при финансовой поддержке
рактерное для второго типа ритмоинотропии, яв-
Российского фонда фундаментальных исследова-
ляется, по-видимому, отличительным признаком
ний (в рамках научного проекта № 18-04-00764).
животных в переходных состояниях, сопровож-
дающихся повышением их функциональной ак-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
тивности после периодов относительного покоя.
Поэтому в наиболее ярком виде особенности,
Исследования проводили в соответствии с
свойственные данному типу ритмоинотропии,
требованиями Европейской конвенции по защи-
проявляются у кратковременно пробуждающих-
те животных (86/609/EEC, 1986).
ся (на завершающей стадии) сусликов в период
гибернации (рис. 2б) или готовящихся к весенне-
СПИСОК ЛИТЕРАТУРЫ
му пробуждению животных (данные для месяца
марта на рис. 1в). Создается впечатление, что
1. H. P. Bowditch, Ber. Sachs Ges. Wiss. 23, 652 (1871).
формирование определенного типа ритмоино-
2. J. Koch-Weser, J. R. Blinks, Pharmacol. Rew. J. 15, 601
тропии в сердце зимоспящих предшествует изме-
(1963).
нениям функционального состояния животных.
3. M. Endoh, Eur. J. Pharmacol. 500, 73 (2004).
БИОФИЗИКА том 64
№ 5
2019
980
НАКИПОВА и др.
4. J. Palomeque, M. G. Vila Petroff, and A. Mattiazzi,
31.
Н. И. Калабухов, Спячка млекопитающих (Наука,
Heart Lung Circ. 13 (4), 410 (2004).
М., 1985).
5. P. M. L. Janssen, Am. J. Physiol. Heart Circ. Physiol.
32.
С. Г. Колаева, Вестн. РАН 63 (12), 1076 (1993).
J. 299, 1741 (2010).
33.
Н. Г. Соломонов, Эколого-физиологические харак-
6. M. M. Monasky and P. M. L. Janssen, J. Comp. Physi-
теристики природных гипометаболических состоя-
ol. B 179, 469 (2009).
ний (Пущино, 1992).
7. Y. Xu, M. M. Monasky, N. Hiranandani, et al., J. Appl.
34.
M. T. Andrews, J. Exp. Biol. 25, 222 (2019).
Physiol. 21, 20 (2011).
35.
M. T. Andrews, K. P. Russeth, and L. R. Drewes, Am.
8. J. H. Chung, B. D. Canan, B. A. Whitson, et al., Physi-
J. Phys. Reg., Integr. Comp. Physiol. 296, 383 (2009).
ol. Rep. J. 6 (20), 13898 (2018).
36.
C. W. Wu, K. K. Biggar, B. E. Luu, et al., Physiol. Ge-
9. E. Bovo, S. R. Mazurek, and A. V. Zima, Pflugers
nom. 48 (6), 388 (2016)
Arch. 470 (6), 959 (2018).
37.
R. L. Russell, P. H. O'Neill, L. E. Epperson, and
10. B. C. Мархасин, В. Я. Изаков и В. И. Шумаков,
S. L. Martin, J. Comp. Physiol. B 180 (8), 1165 (2010).
Физиологические основы нарушения сократительной
функции миокарда (Наука, СПб. 1994).
38.
M. J. Sheriff, C. T. Williams, et.al., J. Comp. Physiol. B
182 (6), 841 (2012).
11. О. Э. Соловьева, В. С. Мархасин и др., Биофизика
44 (2), 337 (1999).
39.
M. Wiersma, T. M. A. Beuren, E. L. de Vrij, et al.,
Comp. Biochem. Physiol. B. Biochem. Mol. Biol. 223,
12. С. А. Афанасьев, Д. С. Кондратьева, Л. П. Цапко и
23 (2018).
др., Вестн. аритмологии 55, 56 (2009).
40.
Д. А. Игнатьев, Г. С. Сухова и В. П. Сухов, Журн.
13. F. Somura, I. Hideo, M. Iwase, et al., Circulation 104
общ. биологии 62 (1), 66 (2001)
(6), 658 (2001).
41.
Д. А. Игнатьев, В. В. Воробьев и Г. С. Сухова,
14. G. Hasenfuss, H. Reinecke, R. Studer, et al., Cardiol.
Нейрохимия 15 (3), 240(1998).
91 (2), 17 (1996).
42.
Д. А. Игнатьев, Г. С. Сухова и В. П. Сухов, Журн.
15. B. Pieske, L. S. Maier, D. M. Bers, et al., Circ. Res. 85,
эволюц. биохимии и физиологии 28 (5), 582 (1992).
38 (1999).
43.
А. И. Ануфриев, Экологические механизмы темпе-
16. J. Palomeque, M. V. Petroff, L. Sapia, et al., J. Card.
ратурных адаптаций млекопитающих и зимующих
Fail. 13 (2), 143 (2007).
птиц Якутии (СО РАН, Новосибирск, 2013).
17. V. J. Schouten and H. E. ter Keurs. J. Mol. Cell Cardiol.
44. F. I. Heinis, K. L. Vermillion, M. T. Andrews, et al.,
23 (9), 1039 (1991).
Am. J. Physiol. Regul. Integr. Comp. Physiol. 309 (4),
18. B. D. Stuyvers, A. D. McCulloch, J. Guo, et al., J.
368 (2015)
Physiol. 544 (Pt 3), 817 (2002).
45.
A. E. Alekseev, N. I. Markevich, A. F. Korystova, et al.,
19. N. Kondo and S. Shibata, Science 225, 641 (1984).
Biophys. J. 70, 786 (1996).
20. N. Kondo, Circ. Res. 59, 221 (1986).
46.
Yu. M. Kokoz, A. S. Grichenko, A. F. Korystova, et al.,
21. J. S. Charnock, W. F. Dryden, and R. J. Marshall, Br.
Membr. Cell. Biol. 14 (2), 277 (2000).
J. Pharmacol. 78 (1), 151 (1983).
47.
S. Q. Wang, E. G. Lakatta, H. Cheng, et al., J. Exp. Bi-
22. Z. Q. Zhou, W. F. Dryden, and L. С. H. Wang, J.
ol. 205, 2957 (2002).
Therm. Biol.. 12 (2), 167 (1987).
48.
A. Yatani, S. J. Kim, R. K. Kudej, et al., Am. J. Physiol.
23. О. В. Накипова, Л. А. Андреева, Н. А. Чумаева
Heart. Circ. Physiol. 286 (6), 2219 (2004).
и др., Биофизика 47 (4). 735 (2002).
49.
K. M. Dibb, C. L. Hagarty, A. S. Loudon, et al., Am. J.
24. О. В. Накипова, Л. А. Андреева, Н. А. Чумаева
Physiol. Regul. Integr. Comp. Physiol. 288 (3), 607
и др., Докл. РАН 407 (4), 547 (2006).
(2005).
25. O. V. Nakipova, N. M. Zakharova, L. A. Andreeva,
50.
X. C. Li, L.Wei, G. Q. Zhang, et al., PLoS One 6 (9),
et al., Cryobiology 55, 173 (2007).
e24787 (2011).
26. H. Reuter, C. Zobel, K. Brixius, et al., Basic Res. Car-
51.
T. Ruf, F. Geiser, Biol. Rev. Camb. Philos. Soc. 90 (3),
diol. 94 (3), 159 (1999).
891 (2015).
27. D. M. Bers, Circ. Res. 87, 275 (2000).
52.
F.Hoelzl, C. Bieber, J. S. Cornils et al., J. Comp. Physi-
ol. B 185 (8), 931 (2015).
28. L. S. Maier, D. M. Bers, B. Pieske, J. Mol. Cell Cardi-
ol. 32, 2249 (2000).
53.
W. T. Shaw, Ecology 6 (1), 75 (1925).
29. B. W. Johansson, Cardiovasc. Res. 31, 826 (1996).
54.
O. V. Nakipova, A. S. Averin, E. V. Evdokimovskii,
et al., PLoS One 12, e0177469 (2017).
30. C. P. Lyman, J. S. Willis, A. Malan, et al., Hibernation
and torpor in mammals and birds (Acad. Press, N.Y.-L.,
55.
A. Krishna, M. Valderrábano, P. T. Palade, et al., The-
1982).
or. Biol. Med. Model. 10, 54 (2013).
БИОФИЗИКА том 64
№ 5
2019
РИТМОИНОТРОПНЫЕ ОТНОШЕНИЯ В ПАПИЛЛЯРНОЙ МЫШЦЕ СЕРДЦА СУСЛИКА
981
Force-Frequency Relationship in the Ground Squirrel Papillary Heart Muscles Precedes
Changes in the Functional State of Animals During an Annual Cycle
O.V. Nakipova, A.S. Averin, L.S. Kosarsky, and D.A. Ignatiev
Institute of Cell Biophysics, Russian Academy of Sciences, Institutskaya ul. 3, Pushchino, Moscow Region, 142290 Russia
This work focuses on the study of peculiarities of the dependence of contractile force (isometric mode, 30°C)
on stimulation frequency (0.1-4.0 Hz) in the right ventricular papillary muscles isolated from the hearts of
ground squirrels Spermophilus undulatus using groups of hibernating animals that are active in summer
(June-July) and autumn (October-November) periods as well as hibernating animals during the hibernation
season, including interbout euthermic period; entrance into torpor (Theart ~ 30°C); torpor and interbout
arousal (Theart ~ 30°C). It is shown that regardless of the functional state of animals, the force-frequency de-
pendence (rhythm-inotropic effect) in the ground squirrel papillary heart muscles is mostly negative: con-
tractile force at high frequencies (from 1.0 to 4.0 Hz) is always less than that at low frequencies (around
0.1 Hz). At the same time, in each group of animals, 2 types of rhythm-inotropic effect are distinguished: the
first effect has a rather weak (type 1) frequency dependence, the second a pronounced one (type 2). Both
types of rhythm-inotropic effect may have a weak positive component (due to an increase in contractile
force): in case of type 1 effect, it appears in the low frequency stimulation range (from 0.2 to 0.5 Hz) (only
in animals during the hibernation season: interbout aroused and torpid or entered in a state of torpor—in
which their metabolism slows), and type 2 effect is observed in the high frequency stimulation range (more
than 1.0 Hz), mostly obvious in animals during the resumption of functional activities through interbout
arousals, and in spring during exit from torpor. The data obtained indicate that the process of formation of a
certain type of rhythm-inotropic effect in the heart of hibernating animals occurs before changes in the func-
tional state of animals during an annual cycle.
Keywords: heart, papillary muscles, contractility, force-frequency relationship, hibernating animals, ground squirrel
БИОФИЗИКА том 64
№ 5
2019