БИОФИЗИКА, 2019, том 64, № 6, с. 1045-1051
МОЛЕКУЛЯРНАЯ БИОФИЗИКА
УДК 577.355
СИНГЛЕТ-ТРИПЛЕТНОЕ ДЕЛЕНИЕ ВОЗБУЖДЕНИЯ КАРОТИНОИДОВ
СВЕТОСОБИРАЮЩИХ КОМПЛЕКСОВ Thermochromatium tepidum
© 2019 г. А.А. Грязнов, И.Б. Кленина, З.К. Махнева, А.А. Москаленко, И.И. Проскуряков
Институт фундаментальных проблем биологии РАН - обособленное подразделение ФИЦ ПНЦБИ РАН,
142290, Пущино Московской области, ул. Институтская, 2
E-mail: pros@issp.serpukhov.su
Поступила в редакцию 10.07.2019 г.
После доработки 10.07.2019 г.
Принята к публикации 22.08.2019 г.
Освещение пурпурных фототрофных бактерий в полосе поглощения каротиноидов светособираю-
щих комплексов часто приводит к низкой эффективности использования энергии поглощенного
света. Известно, что это связано с синглет-триплетным делением возбуждения каротиноидов. В ра-
боте исследован механизм данного процесса на примере фототрофной бактерии Thermochromatium
tepidum. С использованием методов ЭПР высокого временного разрешения и модуляции выхода
флуоресценции магнитным полем показано, что развиваемое в ряде работ представление о внутри-
молекулярном делении возбуждения не находит своего экспериментального подтверждения. Полу-
чены свидетельства межмолекулярного характера деления возбуждения с участием двух молекул ка-
ротиноидов светособирающих пигмент-белковых комплексов LH1-RC и LH2. Обсуждаются пре-
имущества межмолекулярного деления возбуждения для использования в фотопреобразователях
солнечной энергии.
Ключевые cлова: cветоcобиpающие комплексы, каpотиноиды, cинглет-тpиплетное деление возбужде-
ния, электронный парамагнитный pезонанc, эффекты магнитного поля, фототpофные бактеpии.
10.1134/S0006302919060012
в процессе бактериального фотосинтеза выпол-
Первичные процессы преобразования солнеч-
няют две главные функции [3]. Первая - погло-
ной энергии в химическую при фотосинтезе про-
щение света в области оптической пpозpачноcти
исходят в специализированных пигмент-белко-
бактеpиоxлоpофиллов и передача энергии элек-
вых комплексах - реакционных центрах [1]. Для
тронного возбуждения на БXл. Вторая - тушение
повышения скорости фотосинтеза реакционные
тpиплетныx cоcтояний БХл [4], т. е. защита от ге-
центры в фотоcинтетичеcкиx мембpанаx окpуже-
нерации синглетного киcлоpода, возникающего в
ны пигмент-белковыми комплекcами дpугого ти-
результате взаимодействия триплетного БХл с
па, так называемыми антенными или cве-
триплетным киcлоpодом воздуха [5,6].
тоcобиpающими комплекcами. В cоcтав светосо-
бирающих комплексов пурпурных фототрофных
Известно, что реакции фотопереноса электрона
бактерий входят пигмент-белковый комплекс пе-
в реакционных центрах идут с высокой скоростью
риферической антенны LH2 и комплекс LH1-RC.
и с квантовой эффективностью, близкой к едини-
Оба типа комплексов содержат пигментные мо-
це. Процессы переноса энергии между пигмента-
лекулы - бактериохлорофиллы (БХл) и кароти-
ми также оптимизированы. Каротиноиды светосо-
ноиды (Кар), поглощающие свет. При этом ком-
бирающих комплексов LH1 и LH2 за времена в пи-
плекс LH1-RC состоит из прицентровой антенны
ко- и субпикосекундном диапазоне передают
LH1 и реакционного центра, осуществляющего
энергию молекулам БХл. Несмотря на высокую
указанное выше преобразование энергии. Ком-
скорость, эффективность переноса энергии
плексы LH2 не содержат реакционных центров и
1Кар* → БХл у некоторых пурпурных бактерий не
обеспечивают поглощение света и передачу элек-
превышает 25-30% [7]. При этом регистрируется
тронного возбуждения на LH1 [2]. Каротиноиды
образование триплетных состояний каротиноидов.
Прямое заселение 3Каp путем интеркомбинацион-
Сокращения: БХл - бактериохлорофилл, Кар - каротино-
ид, ЭПР - электронный парамагнитный резонанс, РНП -
ной конверсии имеет низкую вероятность, по-
расщепление в нулевом поле.
скольку время жизни синглетно-возбужденныx со-
1045
1046
ГРЯЗНОВ и др.
стояний каротиноидов, составляющее в растворах
низме деления возбуждения. В целях проверки
обычно 1-10 пс [8,9], слишком мало для ее проте-
данного результата мы провели исследование
кания. Синглетное возбуждение каротиноидов в
LH1-RC- и LH2-комплексов Th. tepidum метода-
cоcтаве cветоcобиpающиx комплексов - еще более
ми ЭПР-спектроскопии и модуляции квантового
короткоживущее из-за переноса энергии на БХл.
выхода триплетных состояний магнитным полем
и пришли к выводу, что синглет-триплетное де-
Для объяснения факта образования 3Каp было сде-
ление возбуждения у этой бактерии происходит
лано предположение, что триплетные cоcтояния
по межмолекулярному механизму с участием двух
каротиноидов LH2 возникают из cинглетно-воз-
молекул каротиноидов. Предварительное сооб-
бужденныx в результате деления на два триплета
щение о данной работе было опубликовано в ра-
[10,11]: 1Кар* → 3Кар + 3Кар. Этот разрешенный по
боте [17].
спину процесс может развиваться очень быстро, в
пико- и фемтосекундном диапазоне времен.
Явление синглет-триплетного деления воз-
МАТЕРИАЛЫ И МЕТОДЫ
буждения было открыто в 1965 г. (см. обзоры
Светособирающие комплексы LH1-RC и LH2
[12,13]) в органических молекулярных кристал-
выделяли из клеток Thermochromatium tepidum ме-
лах. Интерес к этому явлению резко возрос после
тодом электрофореза, как описано в работе [18], с
2004 г., когда была выдвинута идея, что его
переводом препаратов в 10 мМ трис-НСl-буфер,
использование может существенно повысить ко-
рН 7,5. В измерениях флуоресценции использо-
эффициент полезного действия фотопреобразо-
вали препараты с исходной оптической плотно-
вателей солнечной энергии [14]. Начиная с ука-
стью 2 ед. при 470 нм. Для приготовления образ-
занного времени, синглет-триплетное деление
цов к препаратам добавляли 66% глицерина и по-
возбуждения активно исследуется с целью выяс-
сле тщательного перемешивания помещали в
нения механизмов этого процесса и поиска осу-
кварцевые капилляры диаметром 3 мм. В резуль-
ществляющих его материалов. Таким образом,
тате максимальная оптическая плотность образ-
изучение данного явления в каротиноидах свето-
цов для измерения флуоресценции составляла
собирающих антенн важно как для понимания
0,2 ед. при 470 нм, обеспечивая их однородное
первичных процессов фотосинтеза, так и для ре-
возбуждение. В измерениях ЭПР использовали
шения прикладных задач повышения эффектив-
аналогично приготовленные препараты с оптиче-
ности преобразователей энергии света.
ской плотностью в полосе поглощения каротино-
Первая прямая регистрация синглет-триплет-
идов, достаточно высокой для практически пол-
ного деления возбуждения каротиноидов LH1
ного поглощения возбуждающего света. Ряд из-
пурпурной бактерии Rhodospirillum rubrum была
мерений ЭПР был проведен при температуре
выполнена в работе [10] методом пикосекундной
260 К. В этом случае образцы помещали в одно-
спектроскопии. Авторами было высказано пред-
миллиметровые капилляры без добавки глицери-
положение, что деление возбуждения протекает
на. В препараты LH1-RC комплексов для оптиче-
внутримолекулярно, с локализацией двух трипле-
ских измерений добавляли 10 мкМ феррициани-
тов на половинах одной молекулы каротиноида.
да для поддержания окисленного состояния
Было показано, что деление возбуждения проис-
первичного акцептора QA и подавления рекомби-
ходит за время менее 5 пс, что объясняет его
национной флуоресценции реакционных цен-
конкуренцию с переносом энергии на БХл. Три-
тров.
плетные состояния БХл в этом процессе не обра-
Оптические и основные ЭПР-измерения про-
зуются. Позднее методом электронного парамаг-
водили при температуре 175 К, которую поддер-
нитного резонанса (ЭПР) высокого временного
живали с помощью криостатов ESR900 (Oxford
разрешения нами было показано, что в комплек-
Instruments, Англия). Для оптических измерений
сах LH2 Allochromatium minutissimum регистриру-
криостат помещали в зазор электромагнита, спо-
ются спектры, соответствующие локализации
собного создавать поле до 480 мТл. Образцы осве-
триплетных состояний на отдельных молекулах
щали вспышками света с частотой повторения
каротиноидов [15], что свидетельствует в пользу
15 Гц, энергией около 1 мДж и длительностью
межмолекулярного механизма деления возбужде-
6 нс от лазерной системы Solar Laser Systems
ния.
(«СОЛАР ЛС», Минск, Беларусь) на длине волны
В недавней работе [16] методом оптического
470 нм. Вспышку флуоресценции после прохож-
флеш-фотолиза было проведено количественное
дения фильтра КС-19 и интерференционного
сравнение выцветания полосы поглощения каро-
фильтра
890 нм регистрировали с помощью
тиноидов за счет образования триплетного состо-
ФЭУ-83 и записывали на осциллографе
яния и появления полосы 3Кар в комплексах
HDO4022 (Teledyne LeCroy, США). Значение для
LH1-RC и LH2 Thermochromatium tepidum и Rhodo-
каждой точки на рис. 2 (см. ниже) получали по-
bacter sphaeroides. На основании этого анализа
парным вычитанием десяти измерений при
был сделан вывод о внутримолекулярном меха-
включенном поле и десяти измерений при вы-
БИОФИЗИКА том 64
№ 6
2019
СИНГЛЕТ-ТРИПЛЕТНОЕ ДЕЛЕНИЕ ВОЗБУЖДЕНИЯ КАРОТИНОИДОВ
1047
ключенном поле, накапливая в одном измерении
Каротиноидный состав светособирающих комплексов
1000 импульсов флуоресценции. Полученные
Thermochromatium tepidum по данным высокоэффек-
тивной жидкостной хроматографии
разностные значения усредняли и делили на
среднюю амплитуду флуоресценции в нулевом
Каротиноиды
LH1-RC
LH2
поле. Измерения ЭПР проводили на спектромет-
ре высокого временного разрешения, как описа-
Дидегидрородопин
5
29,7
но в работе [18]. Для возбуждения образцов ис-
пользовали лазерную систему, описанную выше.
Родопин
-
43,6
При регистрации сигналов триплетных состоя-
ний каротиноидов применяли метод стробоско-
Спириллоксантин
71,6
9,5
пического интегрирования в интервале 150 нс -
1,5 мкс после возбуждающей вспышки при не-
Ангидрородовибрин
6,4
11,1
прерывной развертке магнитного поля. Сигналы
сохраняли в памяти компьютера после их преоб-
Неидентифицированный
13,2
-
разования аналого-цифровым преобразователем
глюкозил-каротиноид
Е14-140-М («L-Card», Россия). В работе пред-
ставлены результаты накопления сигнала после
Остальные
3,8
6,1
суммирования 300 измеренных спектров. Для ка-
либровки магнитного поля использовали ЯМР-
измеритель магнитной индукции Ш1-1 (СССР).
РНП отвечают триплетному состоянию преобла-
Количественный анализ каротиноидов прово-
дающего
каротиноида
спириллоксантина
дили хроматографически, как описано в работе
(n = 13). В случае LH2 спектр хорошо интерпре-
[19].
тируется в предположении вклада каротиноидов
с n = 11, 12 и 13, что также достаточно хорошо со-
РЕЗУЛЬТАТЫ
гласуется с анализом каротиноидного состава.
Важно отметить, что эти параметры однозначно
Результаты анализа каротиноидного состава
указывают на локализацию триплетных состоя-
приведены в таблице. Комплексы LH1-RC содер-
ний на отдельных молекулах каротиноидов, что
жат в качестве основного каротиноида (почти
согласуется с выводами работы [17]. Поскольку
72 мол. %) спириллоксантин (число сопряжен-
данные в работе [16] были получены при комнат-
ных двойных связей n = 13). Основные каротино-
иды LH2 - родопин (n = 11), 44 мол. % и дидегид-
рородопин (n = 12), 30 мол. %. Присутствует так-
же спириллоксантин (n = 13). Количественные
отличия от данных, приведенных в работе [16],
связаны, по-видимому, с разными условиями вы-
ращивания культуры клеток. Спектры поглоще-
ния комплексов совпадают с опубликованными в
работе [16].
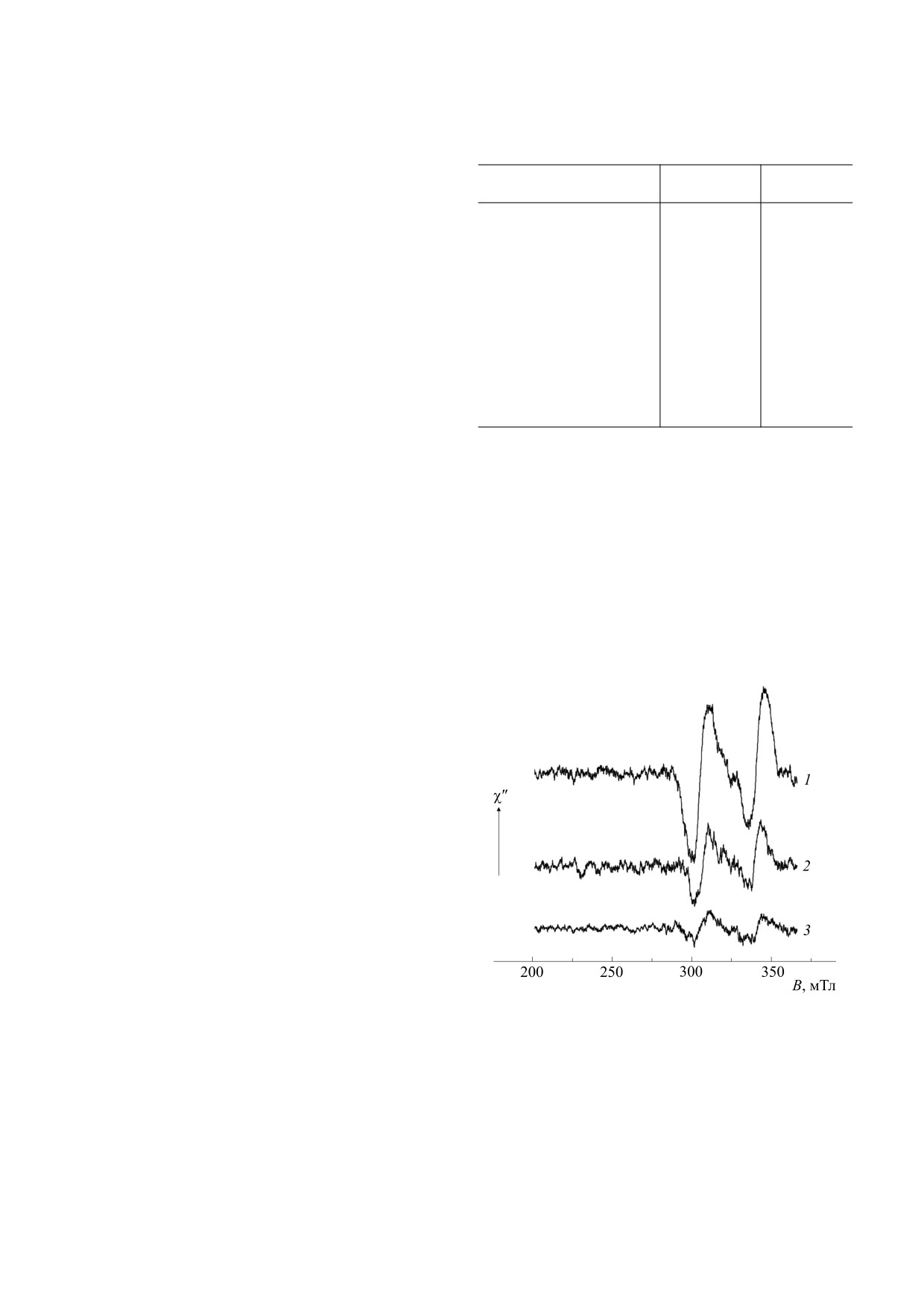
На рис. 1 представлены спектры ЭПР высоко-
го временного разрешения светособирающих
комплексов LH1-RC и LH2 Thermochromatium
tepidum. Ранее спектры этих препаратов были
приведены в диапазоне магнитных полей 280-
365 мТл [17]. Чтобы убедиться в отсутствии до-
полнительных сигналов в изучаемых образцах,
развертку сканируемого поля увеличили с 85 мТл
до 165 мТл. Спектры получены во временном ин-
тервале 150 нс - 1,5 мкс после возбуждающей
вспышки, также более широком, чем в работе
[17]. Представленные спектры соответствуют
триплетным состояниям каротиноидов [15,18] и
Рис. 1. Спектры ЭПР комплексов LH2 (1) и LH1-RC (2)
описываются следующими константами расщеп-
фототрофной бактерии Thermochromatium tepidum,
полученные при Т = 175 К и возбуждении в полосу
ления в нулевом поле (РНП) │D│ = 0,0225 см-1,
поглощения каротиноидов (λвозб = 470 нм) в интервале
│E│ = 0,0022 см-1 (LH1-RC) и │D│ = 0,0250 см-1,
от 150 нс до 1,5 мкс после возбуждающей вспышки
света. Спектр (3) получен на таком же препарате LH2
│E│ = 0,0028 см-1 (LH2). Величина РНП зависит
при 260 К без добавления глицерина. Вертикальная
от длины системы сопряженных связей кароти-
стрелка указывает направление, соответствующее
ноида [20]. Наблюдаемые в LH1-RC параметры
поглощению СВЧ.
БИОФИЗИКА том 64
№ 6
2019
1048
ГРЯЗНОВ и др.
исключительно в синтетических соединениях, в
которых две молекулы одной или разной приро-
ды соединены ковалентным мостиком (см., на-
пример, работу [21]). В работе [22] изучены ли-
нейные молекулы дифенилдицианполиена с чис-
лом сопряженных двойных связей n от 3 до 11,
структурно напоминающие каротиноиды. Время
жизни продукта деления возбуждения в соедине-
нии с n = 7 составляет всего 40 пс. Предположе-
ние о внутримолекулярном механизме деления
возбуждения каротиноидов светособирающих
комплексов пурпурных бактерий было впервые
выдвинуто в работе [10]. Такое предположение
было основано на том, что в светособирающих
комплексах расстояние между каротиноидами
(13-14 Å, [23,24]) слишком велико для реализа-
ции межмолекулярного деления возбуждения,
требующего перекрывания электронных оболо-
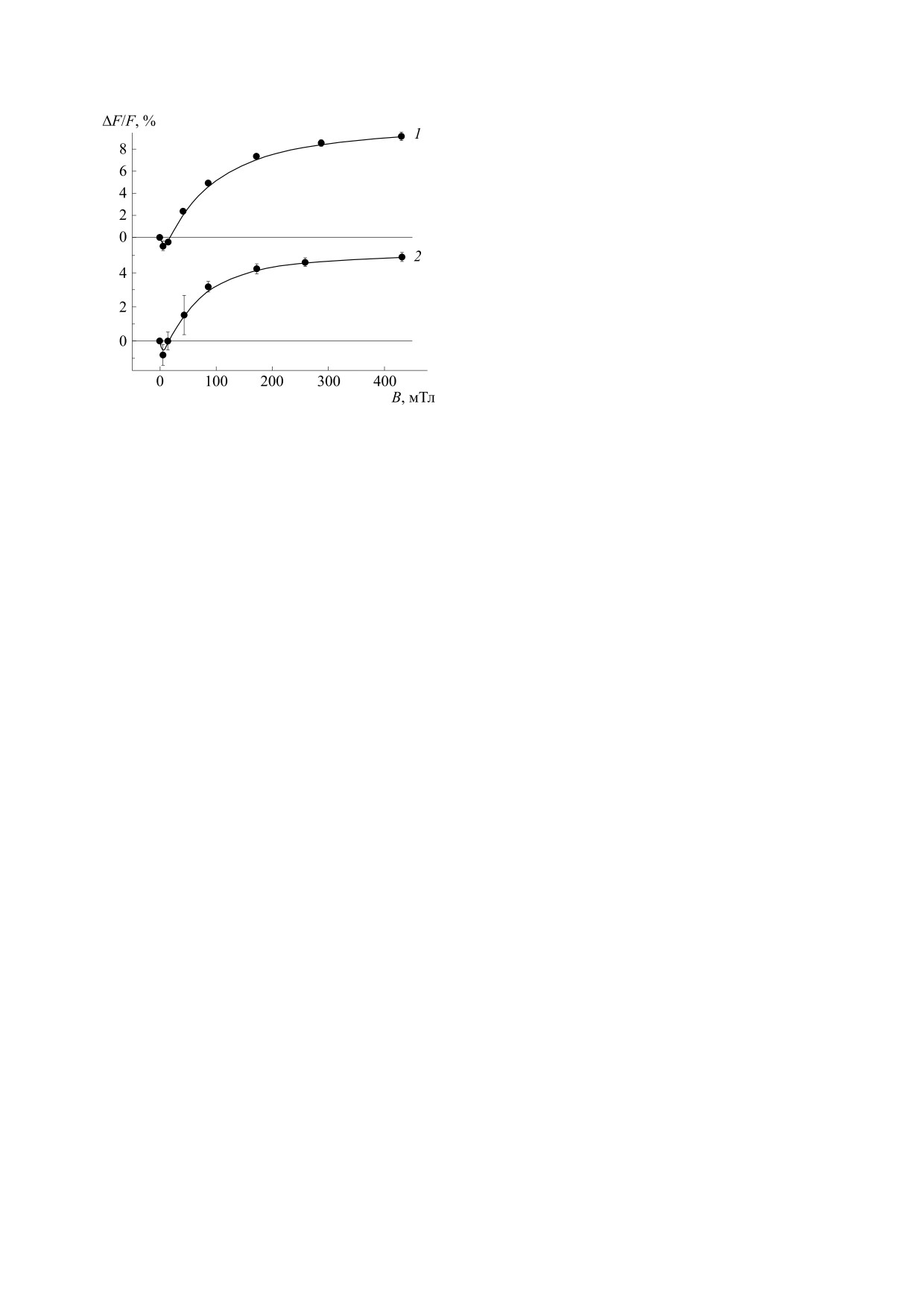
Рис.
2. Относительные изменения флуоресценции
чек молекул, участвующих в процессе. Выводы о
комплексов LH2
(1) и LH1-RC
(2), вызванные
внутримолекулярном механизме деления возбуж-
наложением постоянного магнитного поля. Т = 175 К,
λизм = 885 нм. Поглощение образцов было выровнено
дения были подкреплены теоретическим рас-
при 470 нм.
смотрением процесса на примере длинных и бес-
конечных полиненасыщенных линейных моле-
кул [25]. В работе [16] была предпринята попытка
ной температуре, мы также провели измерения
аккуратно определить число триплетных молекул
при повышенной температуре. На рис.
1
каротиноидов, образующихся при возбуждении
(спектр 3) приведен спектр комплексов LH2, по-
молекулы этого пигмента. Был сделан вывод, что
лученный при Т = 260 К. Значительное уменьше-
в комплексах LH1-RC реализуется внутримоле-
ние его амплитуды по сравнению с рис.
1
кулярный механизм, а в комплексах LH2 наблю-
(спектр 1) объясняется менее благоприятными
условиями измерения (меньшее количество об-
дается примесь иных каналов заселения 3Кар.
разца, плохое оптическое качество препарата без
Напомним, что в наших предыдущих работах
добавки глицерина).
по исследованию деления возбуждения в светосо-
На рис. 2 представлена зависимость относи-
бирающих комплексах LH2 пурпурной серной
тельных изменений флуоресценции при наложе-
бактерии Allochromatium minutissimum, проведен-
нии магнитного поля от величины магнитной ин-
ных методом ЭПР высокого временного разре-
дукции. Следует отметить качественно схожий
шения, получены данные, однозначно указываю-
характер приведенных полевых зависимостей для
щие на локализацию состояний 3Кар на отдель-
LH2- (рис. 2, кривая 1) и LH1-RC-комплексов
ных молекулах каротиноидов
[15,18]. В этих
(рис. 2, кривая 2), а также наличие отрицательных
работах признаков внутримолекулярного деле-
значений эффекта в низких магнитных полях.
ния возбуждения и триплетов, локализованных
Обе кривые в высоких полях выходят на насыще-
на половинах молекулы каротиноида, обнаруже-
ние, что особенно очевидно в случае комплексов
но не было. Такие триплеты имели бы значитель-
LH1-RC (рис. 2, кривая 2). Значения максималь-
но бόльшие константы РНП, поскольку величина
ного эффекта магнитного поля составляют при-
близительно 5% для LH1-RC и около 9% для LH2.
этих констант линейно зависит от 1/n, где n -
Эти величины превышают полученные в работе
число сопряженных двойных связей в молекуле
[17], что связано с бόльшим значением магнит-
каротиноида [20]. В работе [26] была продемон-
ных полей и более корректными условиями изме-
стрирована роль молекул бактериохлорофилла с
рения (низкая оптическая плотность образцов)
максимумом поглощения при 850 нм в делении
при получении данных, представленных на
возбуждения каротиноидов LH2 комплексов Alс.
рис. 2.
minutissimum. Был сделан вывод, что молекулы
БХл, находящиеся между молекулами каротино-
идов, участвуют в этом процессе по суперобмен-
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
ному механизму, создавая «мостик» между каро-
Необходимо отметить, что внутримолекуляр-
тиноидами. Это наблюдение снимает проблему
ное деление возбуждения с образованием доста-
большого расстояния между молекулами кароти-
точно долгоживущих триплетных состояний
ноидов, поскольку БХл удален от обеих соседних
встречается относительно редко, причем почти
молекул каротиноидов лишь на ~6 Å.
БИОФИЗИКА том 64
№ 6
2019
СИНГЛЕТ-ТРИПЛЕТНОЕ ДЕЛЕНИЕ ВОЗБУЖДЕНИЯ КАРОТИНОИДОВ
1049
В работах [15,18] измерения ЭПР были прове-
образующихся в результате синглет-триплетного
дены в интервалах времен 1,5-2,5 мкс и 0,2-
деления возбуждения, наложением магнитного
0,4 мкс после возбуждающей вспышки света и
поля, и регистрируя изменения флуоресценции
только на комплексах LH2 Alc. minutissimum и
бактериохлорофилла комплексов, которая воз-
Rps. palustris. В работе [17] были изучены ком-
никает после переноса энергии синглетного воз-
плексы LH1-RC и LH2 Th. tepidum в условиях экс-
буждения каротиноида на БХл. Магнитное поле
перимента, максимально приближенных к усло-
оказывает влияние на синглет-триплетное деле-
ние возбуждения, изменяя волновые функции
виям работы [16], где оптические измерения 3Кар
промежуточного двутриплетного комплекса и та-
проводили при задержке 150 нс после вспышки.
ким образом изменяя число его спиновых под-
Однако и в этих условиях нами были зарегистри-
уровней, содержащих примесь синглетного со-
рованы только спектры ЭПР 3Кар, локализован-
стояния [27,28]. При этом зависимость от магнит-
ных на отдельных молекулах каротиноидов. В
ного поля выходит на насыщение при переходе к
случае внутримолекулярного механизма деления
высоким полям, когда внешнее поле становится
возбуждения, т. е. при локализации триплетов на
значительно сильнее спин-спиновых взаимодей-
половинах молекул каротиноидов, должны реги-
ствий триплетных состояний. При наличии силь-
стрироваться спектры ЭПР, соответствующие со-
ных спин-спиновых взаимодействий внутри три-
пряженной системе связей с n = 5, 6 или 7. Со-
плетных молекул (РНП, существенно превышаю-
гласно работе [20], параметры РНП │D│ при этом
щее наложенное магнитное поле) или между
двумя триплетами магнитный эффект отсутствует
составляли бы 0,081, 0,065 и 0,0535 см-1 соответ-
в полях, более слабых, чем указанные взаимодей-
ственно. Регистрация подобных триплетов с ис-
ствия [28]. В единицах магнитного поля параметр
пользованием ЭПР трехсантиметрового диапазо-
на возможна. В данной работе мы увеличили раз-
РНП D 3Кар составляет приблизительно 25 мТл.
вертку поля спектрометра ЭПР для поиска таких
Приведенные на рис. 2 зависимости выхода флу-
триплетных состояний и провели измерения в бо-
оресценции БХл от магнитного поля соответству-
лее широком временном интервале (150 нс - 1,5
ют такому значению параметра РНП, поскольку
они выходит на плато в полях ~250 мТл (10 ∙ D),
мкс). При указанных параметрах D сигналы 3Кар
удовлетворяющих условию сильных полей для
ожидаются в диапазонах полей 265-381 мТл (D =
молекул 3Кар. В целом форма этих кривых каче-
0,0535 см-1), 253-394 мТл (D = 0,065 см-1) и 235-
ственно согласуется с ожидаемой теоретически
411 мТл (D = 0,081 см-1). Из рис. 1 видно, что ис-
для межмолекулярного деления возбуждения
комые триплетные состояния отсутствуют и реги-
между двумя одинаково ориентированными мо-
стрируются только спектры ЭПР с параметрами
лекулами одной химической природы (так назы-
РНП, соответствующими триплетам мономеров
ваемое гомогенное деление возбуждения) [28].
каротиноидов. Спектр 3Кар был также измерен
Напомним, что соседние молекулы каротинои-
дов светособирающих комплексов в самом деле
при повышенной температуре 260 К (рис. 1,
ориентированы практически одинаково, по
спектр 3) для проверки предположения, что засе-
крайней мере, в изученных структурах светособи-
ление по мономолекулярному механизму требует
рающих комплексов [3]. Оценим ожидаемую ве-
термоактивации. Как видно из рисунка, и в этом
личину эффекта магнитного поля на флуоресцен-
случае регистрируются только спектры ЭПР, со-
цию БХл в предположении межмолекулярного
ответствующие локализации триплетов на от-
деления возбуждения. Возбужденное состояние
дельных молекулах каротиноидов.
1
Кар* (синглетный уровень S2) дезактивирует по
Оценка квантового выхода состояния 3Кар из
трем каналам: перенос возбуждения на БХл
спектров ЭПР значительно усложнена из-за не-
(ФЕТ ~ 30% в LH2 Th. tepidum [29]), синглет-три-
равновесной спиновой поляризации сигналов.
плетное деление возбуждения (ФST ~ 30% в хро-
Существует вероятность, что при измерениях
матофорах R. rubrum [30], 25% в LH1 R. rubrum
ЭПР наблюдается лишь небольшая доля 3Кар, по
[10]). Оставшаяся часть возбуждения растрачива-
каким-то причинам локализованных на отдель-
ется в тепло (ФH). Для примерной оценки при-
ных молекулах, в то время как основное количе-
мем, что ФЕТ = ФST = 30%. Оценку удобнее про-
ство триплетов, возникших по внутримолекуляр-
водить, используя не квантовые выходы, а доли
ному механизму деления, не регистрируется трех-
молекул N, участвующих в упомянутых процес-
сантиметровым спектрометром. Такая ситуация
сах. Тогда, учитывая возникновение двух три-
может возникнуть при локализации двух трипле-
плетных молекул в процессе деления возбужде-
тов на одной молекуле каротиноида и достаточно
ния, можно записать:
сильных (превышающих энергию кванта сверх-
высокой частоты спектрометра) спин-спиновых
NET + NST + NH = 1 и
взаимодействиях. Мы проверили это предполо-
жение, изменяя выход триплетов каротиноидов,
NET = 0,3; NST = 0,15; NH = 0,55.
БИОФИЗИКА том 64
№ 6
2019
1050
ГРЯЗНОВ и др.
Выход флуоресценции БХл пропорционален
роятность Т-Т-аннигиляции. При межмолеку-
лярном делении вероятность потерь энергии по
NET. Упрощенная теория влияния магнитного поля
предсказывает, что в сильных полях вероятность
этому каналу понижается.
синглет-триплетного деления возбуждения умень-
В заключение отметим, что биологический
шается на 1/3 [27,28]. Таким образом, доля молекул
смысл потерь энергии при делении возбуждения
каротиноидов остается непонятным. Возможно,
3Кар в магнитном поле упадет до 10%, а 5% перерас-
этот процесс является частью механизма защиты
пределится между двумя оставшимися каналами,
бактерий от света высокой интенсивности.
увеличив выход флуоресценции до 31,7%. Отсюда
эффект магнитного поля на флуоресценцию БХл
составит ~ 5,7%, что близко к значению, получен-
ФИНАНСИРОВАНИЕ РАБОТЫ
ному для LH1-комплексов (4,9 ± 0,3%, рис. 2, кри-
вая 2). Максимальное значение магнитного эффек-
Работа по выделению пигментов Th. tepidum и
та на флуоресценцию БХл в случае LH2 комплексов
пигментному анализу (см. раздел «Материалы и
составляет 9,1 ± 0,4% (рис. 2, кривая 1). Близкое
методы») выполнена при частичной финансовой
значение можно получить при небольших измене-
поддержке Российского фонда фундаментальных
ниях распределения по каналам дезактивации
исследований (проект 17-04-00929_а); остальные
результаты получены в рамках Государственного
1Кар*. Так, при NET = 0,28, NST = 0,2, NH = 0,52 по-
задания №АААА-А17-117030110140-5.
лучаем ΔF/F ~ 8%. Следует отметить, что состояние
3Кар с небольшой вероятностью возникает также в
процессе переноса энергии и интеркомбинацион-
КОНФЛИКТ ИНТЕРЕСОВ
ной конверсии 1Кар* → 1БХл* ~ 3БХл → 3Кар, не за-
Авторы заявляют об отсутствии конфликта
висящем от магнитного поля. Это в небольшой сте-
интересов.
пени уменьшает наблюдаемый эффект поля, так
что полученные выше значения являются оценка-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
ми сверху. В целом можно сделать вывод, что ам-
плитуда эффекта с учетом принятых упрощений до-
Настоящая работа не содержит описания ка-
статочно хорошо согласуется с экспериментально
ких-либо исследований с использованием людей
наблюдаемыми величинами. Из этого следует, что
и животных в качестве объектов.
по крайней мере значительная доля актов деления
возбуждения происходит в отсутствие сильных
СПИСОК ЛИТЕРАТУРЫ
спин-спиновых взаимодействий, т. е. по межмоле-
кулярному механизму.
1. B. Ke, Photosynthesis. Photobiochemistry and Photobio-
physics (Kluwer Acad. Publ., Dordrecht, 2001).
H. Hashimoto, Y. Sugai, C. Uragami, et al., J. Photo-
2.
ВЫВОДЫ
chem. Photobiol. C 25, 46 (2015).
3. R. J. Cogdell, A. T. Gardiner, A. W. Roszak, et al.,
Таким образом, полученные нами результаты
Photosynth. Res. 81, 207 (2004).
противоречат представленным в работе [16] выво-
4. R. J. Cogdell and H. A. Frank, Biochim. Biophys. Acta
дам о внутримолекулярном механизме синглет-
895, 63 (1987).
триплетного деления возбуждения каротиноидов
5. А. А. Кpаcновcкий, Биофизика 22 (5), 927 (1977).
светособирающих комплексов Th. tepidum. При-
6. A. A. Krasnovsky, Photochem. Photobiol. 29,
29
чины такого расхождения сложно проанализиро-
(1979).
вать, поскольку в работе [16] не приведено доста-
7. H. A. Frank and R. J. Cogdell, in Carotenoids in photo-
точно подробного описания условий эксперимен-
synthesis (Springer-Science, 1993), ch. 8, pp. 252-326.
та для проведения подобного анализа. Вывод о
8. T. Polivka and V. Sundstrom, Chem. Rev. 104, 2021
межмолекулярном механизме деления возбужде-
(2004).
ния подтверждается также нашими предыдущими
9. D. Niedzwiedzki, J. F. Koscielecki, H. Cong, et al., J.
работами [15,18,26]. Межмолекулярный механизм
Phys. Chem B. 111, 5984 (2007).
имеет важное преимущество перед мономолеку-
10. C. C. Gradinaru, J. T. M. Kennis, E. Papagiannakis,
лярным с точки зрения использования процесса
et al., Proc. Natl. Acad. Sci. USA 98 (5), 2364 (2001).
деления возбуждения при разработке фотопреоб-
11. E. Papagiannakis, J. T. M. Kennis, I. H. M. van Stok-
разователей солнечной энергии. Деление возбуж-
kum, et al., Proc. Natl. Acad Sci. USA 99, 6017 (2002).
дения протекает наиболее эффективно при при-
12. M. B. Smith and J. Michl, Chem. Rev. 110, 6891
мерном равенстве ES и 2ET, где ES - энергия син-
(2010).
глетно-возбужденного уровня, а ET - энергия
13. M. B. Smith and J. Michl, Annu. Rev. Phys. Chem. 64,
триплетного уровня молекул, участвующих в про-
361 (2013).
цессе [13]. При этом локализованные на одной
14. M. C. Hanna and A. J. Nozik, J. Appl. Phys. 100 (7),
молекуле триплеты будут иметь повышенную ве-
074510-1 (2006).
БИОФИЗИКА том 64
№ 6
2019
СИНГЛЕТ-ТРИПЛЕТНОЕ ДЕЛЕНИЕ ВОЗБУЖДЕНИЯ КАРОТИНОИДОВ
1051
15. И. Б. Кленина, З. К. Махнева, А. А. Москаленко
23. G. McDermott, S. M. Prince, A. A. Freer, et al., Na-
и др., Докл. РАН 441 (6), 833 (2011).
ture 374, 517 (1995).
16. J. Yu, L.-M. Fu, L.-J. Yu, et al., J. Am. Chem. Soc.
24. V. Cherezov, J. Clogston, M. Z. Papiz, et al., J. Mol. Bi-
139, 15984 (2017).
ol. 357, 1605 (2006).
17. И. Б. Кленина, А. А. Грязнов, З. К. Махнева и др.,
25. P. Tavan and K. Shulten, Phys. Rev. B 36, 4337 (1987).
Докл. РАН 485 (4), 511 (2019).
26. И. Б. Кленина, З. К. Махнева, А. А. Москаленко
18. И. Б. Кленина, З. К. Махнева, А. А. Москаленко и
и др., Биохимия 79 (3), 310 (2014)
др., Биофизика 58 (1), 54 (2013).
27. R. E. Merrifield, Pure Appl. Chem. 27 (3), 481 (1971).
19. A. Ashikhmin, Z. Makhneva, and A. Moskalenko,
Photosynth. Res. 119, 291(2014).
28. C. E. Swenberg and N. E. Geacintov, in Organic mole-
cular photophysics, Ed. by J. B. Birks (London. John
20. A. Angerhofer, F. Bornhauser, A. Gall, et al., Chem.
Wiley & Sons Ltd., 1973), pp. 489-564.
Phys. 194, 259 (1995).
21. K. Bhattacharyya, D. Dey, and A. Datta, J. Phys.
29. D. M. Niedzwiedzki, M. Kobayashi, and R. E. Blan-
Chem. C 123, 4749 (2019)
kenship, Photosynth. Res. 107, 177 (2011).
22. M. Tuan Trinh, Yu Zhong, Qishui Chen, et al., J. Phys.
30. H. Rademaker, A. J. Hoff, R. van Grondelle, et al.,
Chem. C 119, 1312 (2015).
Biochim. et Biophys. Acta 592, 240 (1980).
Singlet-Triplet Fission of Carotenoid Excitation in the Light-Harvesting Complexes
from Thermochromatium tepidum
A.A. Gryaznov, I.B. Klenina, Z.K. Makhneva, A.A. Moskalenko, and I.I. Proskuryakov
Institute of Basic Biological Problems, Russian Academy of Sciences,
ul. Institutskaya 2, Pushchino, Moscow Region, 142290 Russia
Excitation of purple phototrophic bacteria into the absorption band of carotenoids of the light-harvesting
complexes often results in low efficiency of light energy utilization. It is known that it is due to singlet-triplet
fission of carotenoid excitation. In the present work, the mechanism of this process is studied using the pho-
totrophic bacterium Thermochromatium tepidum. By means of time-resolved EPR spectroscopy and magnetic
field modulation of the fluorescence yield, it is demonstrated that the idea of intramolecular excitation fission
developed in several publications does not find its experimental confirmation. Evidence of intermolecular
character of excitation fission involving two carotenoid molecules of the light-harvesting pigment-protein
complexes LH1-RC and LH2 is obtained. The advantages of intermolecular excitation fission for application
in photovoltaic solar energy converters are discussed.
Keywords: light harvesting complexes, carotenoids, singlet-triplet fission of excitation, electron paramagnetic res-
onance, magnetic field effects, phototrophic bacteria
БИОФИЗИКА том 64
№ 6
2019