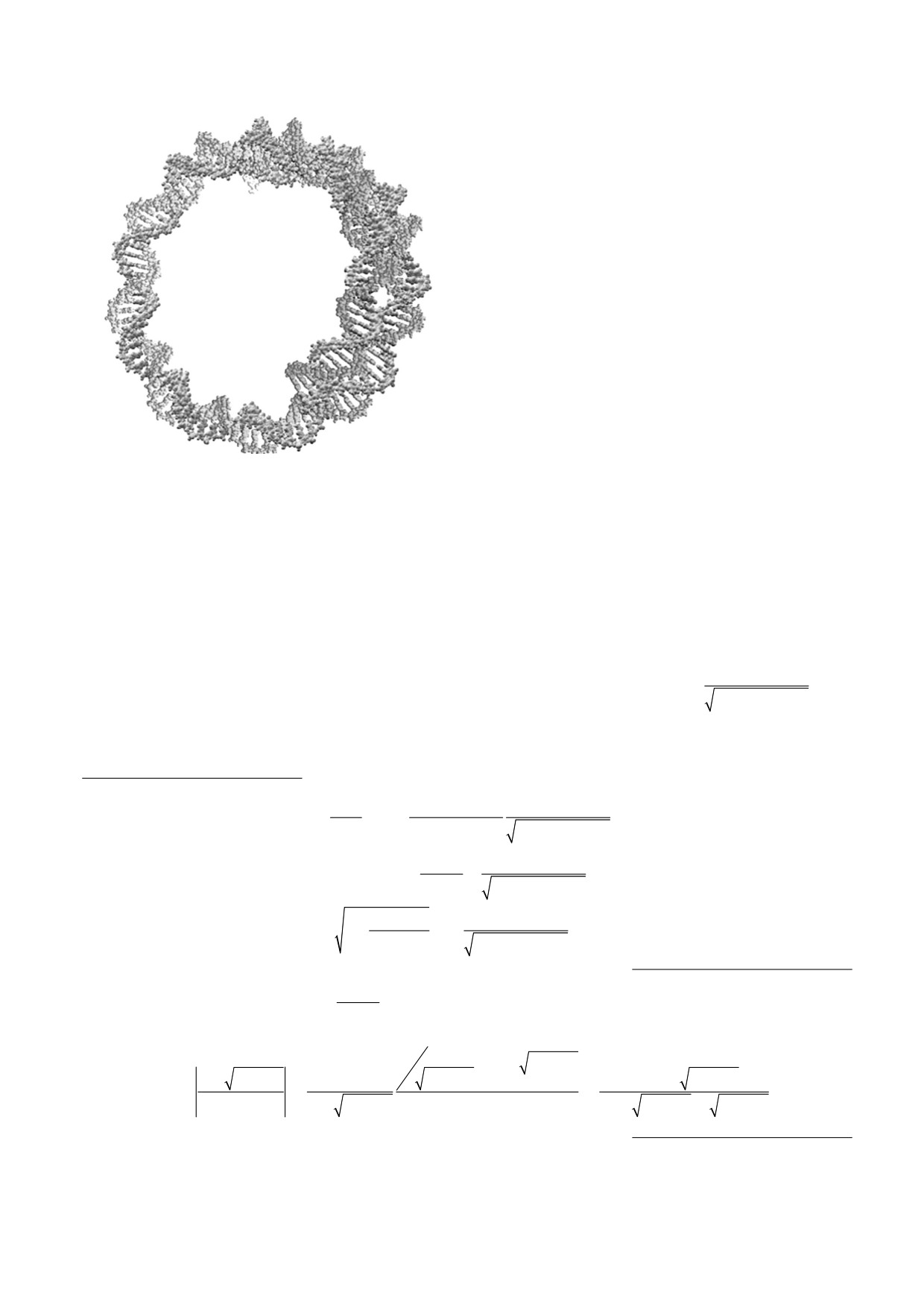БИОФИЗИКА, 2019, том 64, № 6, с. 1151-1162
БИОФИЗИКА КЛЕТКИ
УДК 576.311.347.3:536:66.087
МИТОХОНДРИАЛЬНЫЙ ТЕРМОДИНАМИЧЕСКИЙ
И ЭЛЕКТРОХИМИЧЕСКИЙ ЦИКЛЫ (ПРЯМОЙ И ОБРАТНЫЙ)
© 2019 г. А.С. Татевосян, А.В. Бунякин*
Кубанский государственный медицинский университет Минздрава России,
350063, Краснодар, ул. Митрофана Седина, 4
*Кубанский государственный университет, 350040, Краснодар, ул. Ставропольская, 149
E-mail: artur-krasnodar@bk.ru
Поступила в редакцию 04.04.2019 г.
После доработки 11.09.2019 г.
Принята к публикации 16.09.2019 г.
Проведен мета-анализ функциональных состояний митохондрии, к которым применена математи-
ческая модель, основанная на меняющейся направленности и скорости термодинамических и элек-
трохимических параметров (давление, объем, температура, энтропия, потенциал Гиббса, эксер-
гия…), сопряженных между собой. Основываясь на фундаментальном принципе супрамолекуляр-
ного функционального взаимодействия, создана математическая модель термодинамического и
электрохимического цикла митохондрии, в которой отражены термодинамические и электрохими-
ческие реакции, происходящие в системе «митохондреальный матрикс - внутренняя мембрана -
межмембранное пространство», сопровождающиеся накоплением электрохимического потенциа-
ла и разделением заряда (ионизацией). Рассмотрены четыре функциональных состояния митохон-
дрии, когда в толще ее внутренней мембраны изменяется скорость и направленность движения
электронов по дыхательной цепи, сопряженной с изменением теплового потенциала. Термодина-
мический и электрохимический цикл митохондрии представляет собой непрерывную последова-
тельность переходов между квазистатическими состояниями, при которых циклически изменяются
квазиравновесные состояния, являя собой типичную когерентную устойчивость неравновесных
процессов, о которых высказывался И. Пригожин. Основываясь на математической модели термо-
динамического поведения митохондрии и исходя из ограничений, накладываемых законами физи-
ческой и химической термодинамики, установлено, что наибольшей степени термодинамического
совершенства в процессе митохондриального дыхания соответствует состояние, при котором энер-
гия Гиббса и эксергия минимальны, а энтропия максимальна, что позволяет считать это функцио-
нальное состояние основным (базовым).
Ключевые слова: термо-динамический электро-химический цикл митохондрии, тепловой потенциал,
энергопреобразование АДФ в АТФ, кольцевой квантовый ток в замкнутой молекуле мтДНК.
DOI: 10.1134/S0006302919060152
Не меньший интерес представляет предложен-
Актуальность изучения процессов, протекаю-
ная Р. Вильямсом модель локального сопряжения
щих в митохондрии, очевидна, в первую очередь
дыхания и фосфорилирования [2]. Хотя сам автор
это касается энергопреобразования. Преимуще-
в своей схеме не указывает, в какую форму энер-
ственно рассматриваются процессы запасания
гии трансформируется энергия окислительной
энергии в виде молекул АТФ путем окисления
реакции, мы полагаем возможным по-новому
кислородом продуктов клеточного обмена ве-
взлянуть на старую версию под углом зрения био-
ществ, поступающих из цитоплазмы, с последую-
физической термодинамики и отнестись к функ-
ционированию митохондрии как энергопреобра-
щим выведением ионов водорода (Н+) в межмем-
зующей системе, к которой может быть примене-
бранное пространство, создавая межмембранный
на термодинамическая аналогия, поскольку
электрохимический градиент - модель П. Мит-
убеждены в том, что именно тепло правомочно
челла [1].
считать изначальной энергетической формой,
которую митохондрия аккумулирует, опосредо-
Сокращения: F-I, F-II, F-III, F-IV - ряд функциональ-
ванно увеличивая скорость движения электронов
ных состояний (Functional states) митохондрии, ТД-цикл -
термодинамический цикл, ЭХ-цикл - электрохимический
в электрон-транспортной дыхательной цепи, пе-
цикл.
решедшей в состояние суперкомплекса, с после-
1151
1152
ТАТЕВОСЯН, БУНЯКИН
дующим преобразованием молекулы АДФ в АТФ
МЕТОДЫ ИССЛЕДОВАНИЯ
без выхода Н+ в межмембранное пространство.
Проведен мета-анализ множества описанных
Как полагают отечественные исследователи [3-
состояний митохондрии [7-9], из которых, с
6], митохондрия переключается на выработку
учетом иерархичности термодинамического-
АТФ по схеме Вильямса, преобразуясь в особое
электрохимического (ТД-ЭХ) цикла, выделяем
состояние суперкомплекса с тем, чтобы избежать
четыре функциональных: F-I, F-II, F-III, F-IV
разобщающего эффекта жирных кислот, при
(схема 1) [11]. Все представленные термодинами-
этом авторы, не углубляясь в тепловые эффекты,
ческие параметры (давление, объем, температура,
обходят явления
«теплопотери», традиционно
энтропия, потенциал Гиббса, эксергия) и элек-
считая их неминуемым «злом», снижающим ко-
трохимический потенциал отражают состояние
эффициент полезного действия биологической
внутренней мембраны митохондрии и ее матрик-
са. Среда матрикса, строго говоря, не может быть
системы, каковой является митохондрия.
охарактеризована ни одним из трех классических
Описаны различные состояния митохондрии
агрегатных состояний, так как характерный раз-
[7-9], при которых в зависимости от наличия или
мер внутриматричного пространства имеет уль-
отсутствия субстратов, кислорода, кальция и т.д. на
трамикрогетерогенную величину, сравнимую с
внутренней мембране поэтапно создается электро-
конгломератами молекул коллоидного раствора
химический градиент, использующийся в последу-
(порядка 10-100 нм), что исключает коррект-
ющем для превращения АДФ в АТФ. При этом с
ность устойчивого преобладания какого-либо од-
позиции термодинамики супрамолекулярного вза-
ного фазового состояния [12]. Матрикс митохон-
имодействия квазизамкнутых квазиравновесных
дрии - это специфическая агрегатная среда, в ко-
торой
находятся
кольцевидно-замкнутые,
систем, о которых упоминал Г.П. Гладышев [10],
исключительно важную роль играет функциональ-
вытянутые полимерные молекулы митохондри-
альной ДНК (мтДНК), взаимно зацепленные и
ное состояние митохондрии, именуемое по Чансу
сверхспирализованные, приближающиеся к со-
«дыхательный контроль», которое является термо-
стоянию «жидкого кристалла». Авторы убежде-
динамически самым совершенным, поскольку
ны, что одноименная заряженность и, как след-
именно в этом состоянии энергии Гиббса и эк-
ствие, электростатическое отталкивание мтДНК
сергия самые минимальные. Среди множества
придает матричной среде сходство с ионизиро-
функциональных состояний (Functional states)
ванным газом (плазмой).
митохондрии, состояние «дыхательного контро-
Между характерными функциональными со-
ля» допустимо считать основным (базовым), т. е.
стояниями ТД-ЭХ-цикла митохондрии осу-
F-I, при котором осуществляется прямое влия-
ществляются четыре перехода, которые обозна-
ние электрохимического градиента на скорость
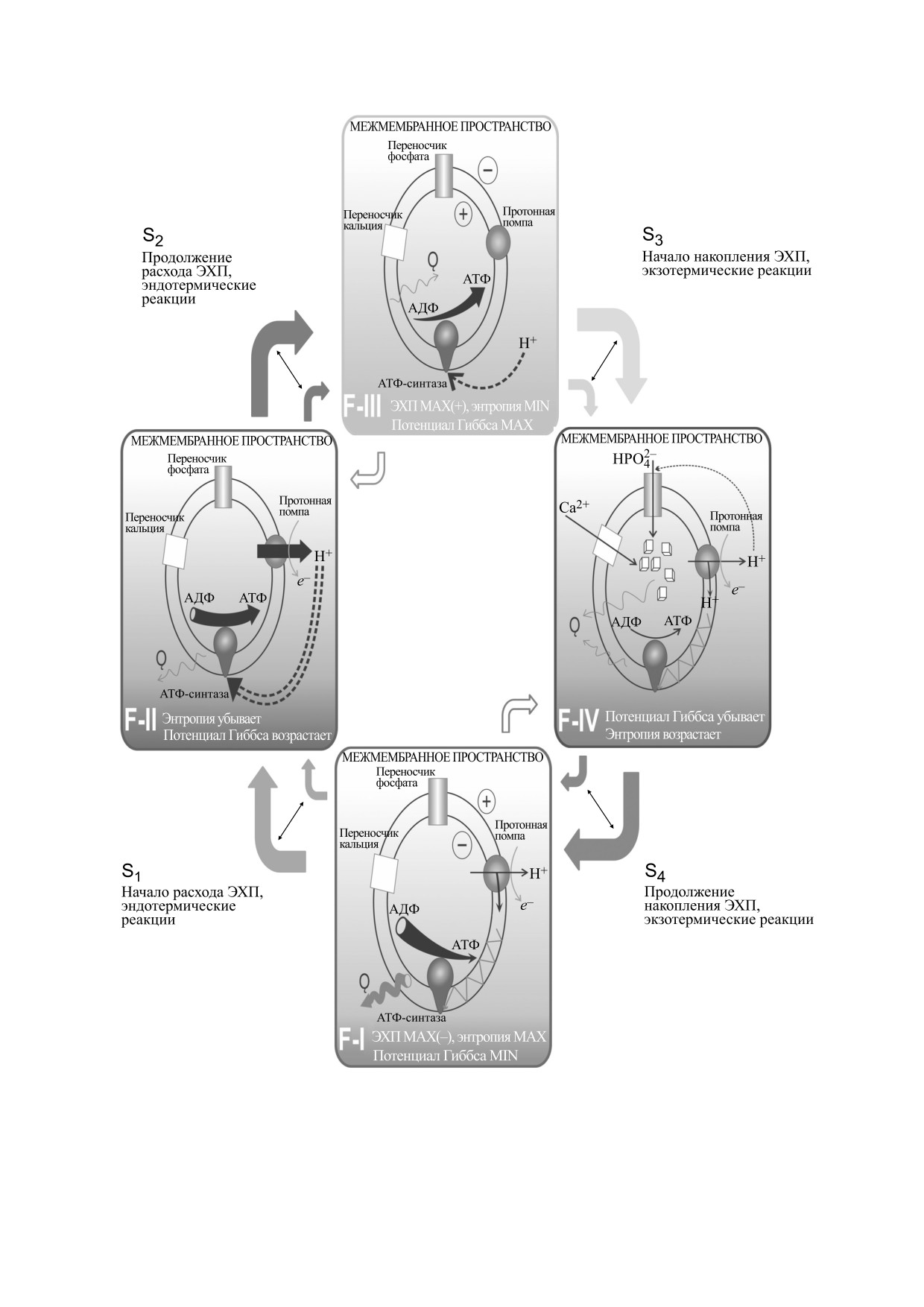
чены как S1, S2, S3 и S4. Переходы S2 и S4 обрати-
движения электронов по дыхательной цепи, уве-
личивая тепловой потенциал, который в свою
мые, при этом параметры процесса изменяются
противоположным образом (возрастание сменя-
очередь способен преобразовать АДФ в АТФ (мо-
ется убыванием и наоборот; на схеме 1 показаны
дель Вильямса). Поскольку при столь малых уль-
стрелками без заливки).
трамикрогетерогенных величинах вести речь о
показателях температуры не корректно, предла-
Далее выделены основные термодинамиче-
гаем в первую очередь учитывать направленность
ские и электрохимические процессы, происходя-
щие на каждом из последовательных переходов
теплового потенциала, и, во-вторых, используя
аналогию с газом, для плазмоподобной среды
(от одного состояния к соседнему - по часовой
стрелке на схеме 1). С ориентировкой на то, что
внутри матрикса, под тепловым потенциалом
представлено на схеме 1, а также на то, что про-
подразумевать произведение давления на объем:
цесс кристаллообразования гидрофосфата каль-
Q = PV.
ция в матриксе (S3) идет с выделением тепла,
Целью работы было создание, с учетом схем
можно утверждать, что при этом процессе повы-
окислительного фосфорилирования, предложен-
шается степень упорядоченности - энтропия
ных Митчеллом и Вильямсом, математической
убывает, энергия Гиббса нарастает, т. е. можно
модели термодинамического (ТД) и электрохи-
заключить, что обратный процесс (S1 - поглоще-
мического (ЭХ) цикла митохондрии, в которой
ние тепла) является своего рода резервом охла-
основную роль играют направленность и скорость
ждения (кондиционирования) матрикса мито-
ТД- и ЭХ-параметров (давление, объем, темпера-
хондрии.
тура, энтропия, потенциал Гиббса, эксергия),
объясняющие энергетические трансформации
ПРЯМОЙ ЦИКЛ МИТОХОНДРИИ
при ее функциональной деятельности с потенци-
альной возможностью обратимости переходов на
При описании прямого ТД-ЭХ-цикла мито-
двух участках цикла.
хондрии приняты следующие термины и опреде-
БИОФИЗИКА том 64
№ 6
2019
МИТОХОНДРИАЛЬНЫЙ ТЕРМОДИНАМИЧЕСКИЙ И ЭЛЕКТРОХИМИЧЕСКИЙ ЦИКЛЫ
1153
Схема. 1. Функциональные (F) состояния и фазовые (S) переходы на термодинамическом/электрохимическом цикле
митохондрии.
БИОФИЗИКА том 64
№ 6
2019
1154
ТАТЕВОСЯН, БУНЯКИН
следнее означает, что состояние митохондрии
(как сложной ТД-ЭХ-системы) не характеризу-
ется однозначно термодинамическими парамет-
рами (P - давление, V - объем).
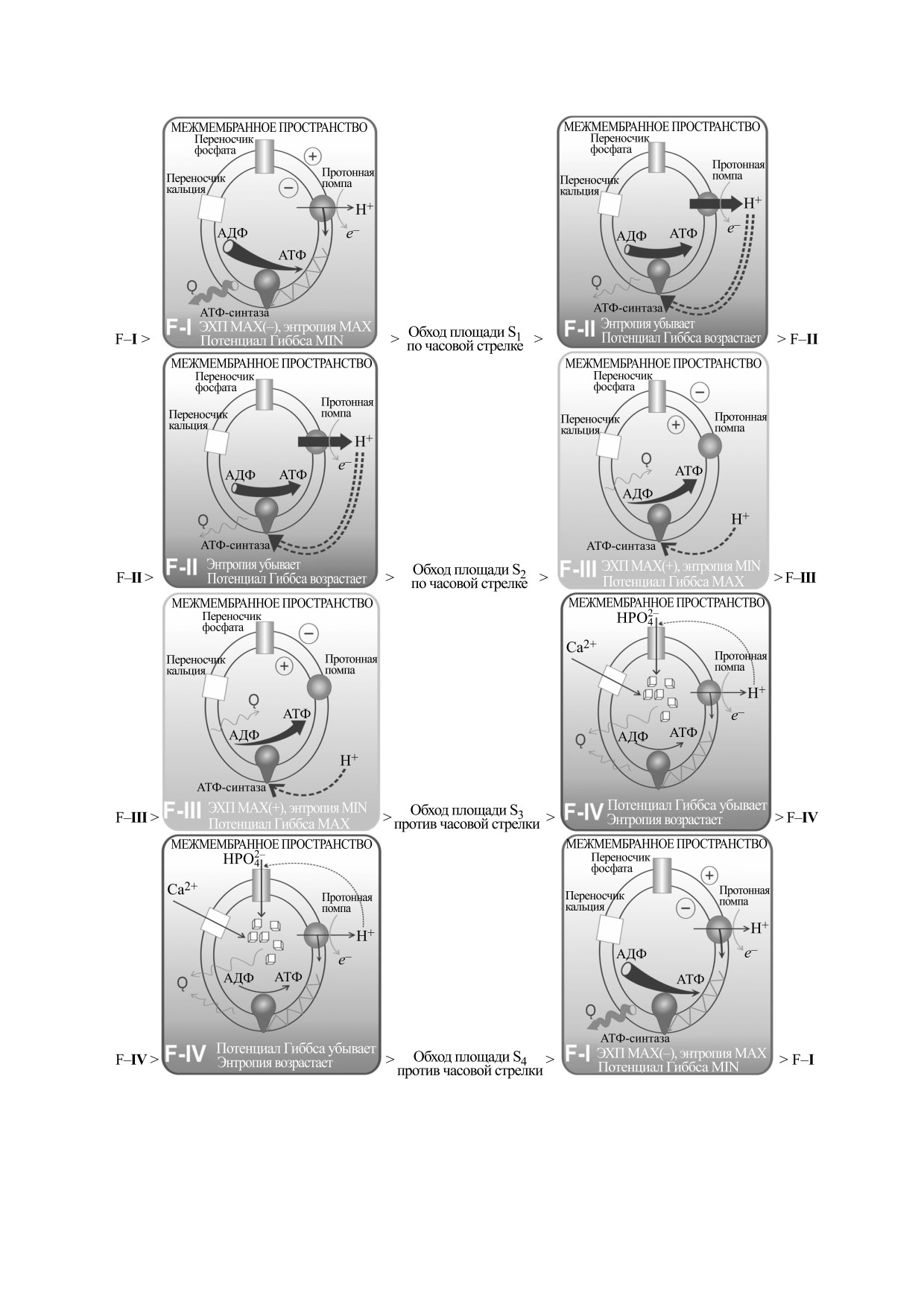
Топология цикла допускает отбрасывание об-
ластей, ограничивающих площади S1 и S3, т. е. за-
мыкание цикла может производиться обходом
лишь областей, ограничивающих площади S2 и S4
(при сохранении направления движения), это
важно с точки зрения перехода митохондрии на
Рис.
1. Термодинамический и электрохимический
малый цикл (об этом также будет сказано ниже).
циклы митохондрий на плоскости объема V и давления
P внутри матрикса митохондрии.
Биоэлектрохимическая и термодинамическая
характеристика состояний:
- F-I - максимум нагрева и заряда матрикса
ления: «нагрев и охлаждение», а также накопле-
митохондрии, работа протонной помпы, АТФ-
ние и расходование электрохимического потен-
синтазы останавливаются, кальций в недостатке,
циала
- «заряд и разряд»
- считаются для
кислород в избытке;
матрикса митохондрии и внутренней поверхно-
- F-II - начало разряда и охлаждения матрик-
сти внутренней мембраны, т. е. относятся к внут-
са митохондрии, работа протонной помпы, за-
риматричной среде.
пуск АТФ-синтаз, кальций в недостатке, кисло-
Накопление электрохимического потенциала
род в избытке;
(заряд Δφ) понимается как повышение концен-
- F-III - максимум разряда и охлаждения
трации ионов водорода Н+ в межмембранном
матрикса митохондрии, остановка протонной
пространстве, соответственно разряд - это сни-
помпы, работа АТФ-синтаз, кальций и кислород
жение там же концентрации ионов водорода Н+
в недостатке;
или накопление каких-либо отрицательно заря-
- F-IV - начало нагрева и заряда матрикса
женных ионов (гидроксильных групп ОН¯) в мат-
митохондрии, запуск протонной помпы, оста-
риксе.
новка АТФ-синтаз, кальций и кислород в из-
В соответствии с принципом систематизации
бытке.
нагрев и заряд матрикса митохондрии противо-
Примечательно то, что запуск и остановка
стоят (в смысле положения переходов в цикле на
протонной помпы синхронно сопровождаются
схеме 1) его охлаждению и разряду. Большие
нагревом с расширением и охлаждением со сжа-
стрелки соответствуют большей скорости реак-
тием матрикса митохондрии соответственно. Ха-
ций, малые - меньшей, а тонкие стрелки между
рактеристики переходов между состояниями в
ними обозначают возможное переключение ско-
соответствии с топологией цикла (номер перехо-
рости перехода. На основании представленной
да соответствует номеру состояния, из которого
схемы построен топологический цикл митохон-
он осуществляется - рис. 1) и соответствие
дрии на плоскости объема и давления, при этом
между участками цикла и переходами приведены
необходимо учесть энергетические преобразова-
на схеме 2.
ния, происходящие на каждом из переходов.
Пояснения требует то, что состояния F-II и
Характерные (функциональные) состояния
F-IV на плоскости «P, V» находятся в одной
F-I и F-III на плоскости объема и давления «P,
точке - это результат проекции цикла на эту
V» (рис. 1) соответствуют точкам самопересече-
плоскость, т. е. того, что только эти два термоди-
ния цикла, лежащим на границах областей с пло-
намических параметра не характеризуют состоя-
щадями S1 и S3 (не контактируют между собой).
ние митохондрии как сложной ТД-ЭХ-системы.
Переходы между состояниями (обозначены так
Необходимо добавить как минимум еще один па-
же, как и площади внутри областей) осуществля-
раметр - это величина электрохимического по-
ются в направлениях, указанных стрелками, на-
тенциала Δφ, тогда цикл станет трехмерным, со-
чинаются в точках функциональных состояний с
стояния F-II и F-IV будут находиться одно над
соответствующими номерами и заканчиваются в
другим, а на плоскости «P, V» их можно отличить
точках со следующими по порядку номерами.
по участкам цикла, которые приводят в точку их
Точка самопересечения, в которой контакти-
проекции (состояния меняются в порядке их ну-
руют области с площадями S2 и S4, соответствует
мерации при движении по трехмерному циклу -
рис. 2).
состоянию F-II, когда прибытие в нее (при дви-
жении по циклу) происходит из состояния F-I, и
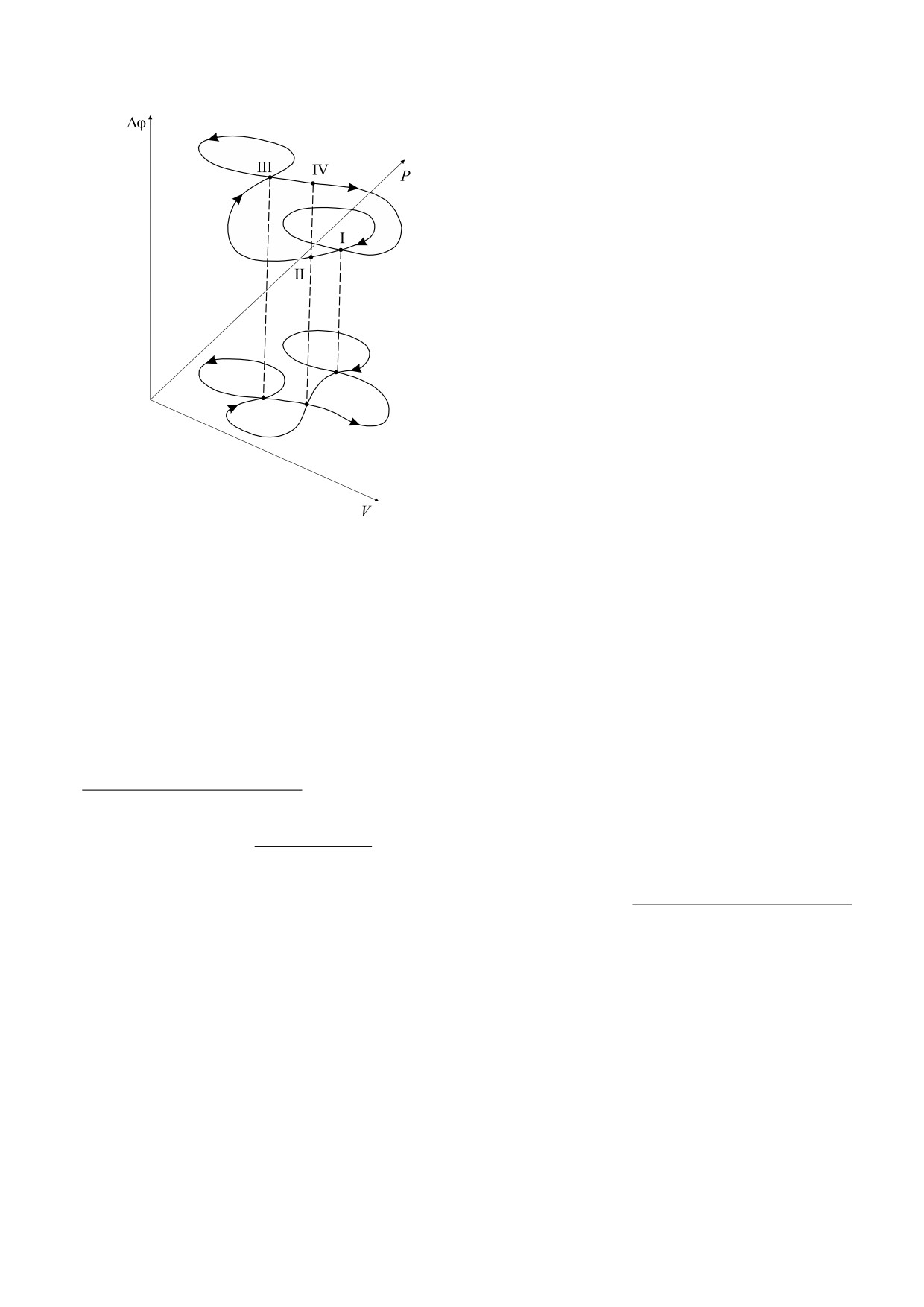
Такая картина трехмерного цикла все же явля-
эта точка соответствует состоянию F-IV, когда
ется идеализированной хотя бы потому, что «ве-
прибытие происходит из состояния F-III. По-
роятность» самопересечения (выхода митохон-
БИОФИЗИКА том 64
№ 6
2019
МИТОХОНДРИАЛЬНЫЙ ТЕРМОДИНАМИЧЕСКИЙ И ЭЛЕКТРОХИМИЧЕСКИЙ ЦИКЛЫ
1155
Схема 2. Характеристика фазовых переходов (S) между функциональными состояниями (F) согласно топологии и
соответствие между участками цикла и переходами.
БИОФИЗИКА том 64
№ 6
2019
1156
ТАТЕВОСЯН, БУНЯКИН
сти «P, V») представляет собой замкнутую кривую
с тремя самопересечениями, а в пространстве «P,
V, Δφ» - с двумя самопересечениями (рис. 2, нуль
Δφ условный).
Площадь внутри замкнутой кривой (на плос-
кости «P, V») вычисляется криволинейным инте-
гралом в двух вариантах (обход контура произво-
дится по часовой стрелке):
S = ∫ PdV = -∫VdP = A
Этот же интеграл имеет смысл работы, совер-
шаемой средой с давлением «P» при изменении
объема «V».
По аналогии с идеальным газом удельное по
массе тепло, отдаваемое термодинамической си-
стемой, вычисляется следующим интегралом [13]
(поглощаемое тепло условно берется с обратным
знаком, или же обход контура производится про-
тив часовой стрелки):
μ
Q
=
(C
P
PdV +C
V
VdP),
∫
R
Рис.
2. Трехмерный цикл митохондрии (разница
где CV и CP - это теплоемкости газа при постоян-
между состояниями F-II и F-IV).
стве объема или давления соответственно; для иде-
ального газа известно тождество CP - CV = R/μ,
дрии на те же параметры) нулевая, т. е. реальный
представляющее собой термодинамический ана-
цикл «расслаивается» и в этих точках, но накла-
лог закона сохранения энергии - «первое начало
дывающиеся при проекции (в точки I и III) «вет-
термодинамики» в терминах теплоемкостей;
R ≈ 8,31 Дж/(град·моль) - универсальная газовая
ви» цикла близко расположены (пояснение со-
постоянная; μ - молярная масса плазмоподобной
стоит в интерпретации малого цикла митохон-
среды внутри матрикса митохондрии.
дрии).
Согласно теории термодинамических потен-
Исходя из вышесказанного, выдвигается ги-
циалов, после деления подынтегрального выра-
потеза (принцип систематизации), состоящая в
жения для тепла Q на Q = PV, т. е. - на тепловой
том, что ТД-ЭХ-цикл митохондрии (на плоско-
потенциал (аналог температуры):
μ
C
PdV +C
VdP
μ
μ
⎛C
⎞
P
V
dV
dP
P
dV dP
=
C
+C
=
C
+
∫
∫ (
P
V
)
V
∫
⎜
⎟
R
PV
R
V
P
R ⎝C
V P
V
⎠
под интегралом появится полный дифференциал
по термодинамическому циклу в идеале должна
от энтропии (для идеального газа она выражается
иметь нулевое изменение, а в реальности - воз-
следующим образом):
растает вследствие необратимых энергетических
потерь - непостоянства теплоемкостей (второе
C
V
P
начало термодинамики).
P
S = S
0
+
ln
+
ln
C
V
V
0
P
0
При электрохимических реакциях аналогом
механической работы является изменение потен-
Поскольку энтропия - это функция, являю-
циала Гиббса G. Для идеального газа он аналоги-
щаяся наиболее часто упоминаемым термодина-
чен потенциальной энергии, при этом величина
мическим потенциалом, она качественно похожа
ΔH = CPΔT представляет собой изменение эн-
на тепло (связана монотонной зависимостью),
но, в отличие от тепла, при прохождении системы тальпии
БИОФИЗИКА том 64
№ 6
2019
МИТОХОНДРИАЛЬНЫЙ ТЕРМОДИНАМИЧЕСКИЙ И ЭЛЕКТРОХИМИЧЕСКИЙ ЦИКЛЫ
1157
μ
Размер митохондриального генома меньше 1%
ΔG =ΔH −ΔQ =C
ΔT
−
(C
PΔV +C
VΔP)
=
P
P
V
всей клеточной ДНК и составляет 16,6 Кб (около
R
1000 нуклеотидов) - порядок длины кольцевой
μ
μ
=
C
(PΔV +VΔP)
−
(C
PΔV +C
VΔP)
=
молекулы ДНК митохондрии (мтДНК) 0,1 мкм и
P
P
V
R
R
более - обозначается как L ≈ 10-7 м.
μ
=
(C
-C
)V
ΔP PΔV
= -ΔA
На прямом термодинамическом цикле внача-
P
V
R
ле (в состоянии F-I) тепловая энергия трансфор-
По этой аналогии полагается, что на частях
мирует электрон-транспортную дыхательную
цикла, ограничивающих области с площадями S1
цепь в суперкомплекс согласно с моделью Ви-
и S2, за счет изменения объема и давления при
льямса. При этом Н+, не покидая пределов внут-
ренней мембраны митохондрии и возвращаясь в
электрохимических реакциях на внутренней
матрикс через АТФ-синтазный комплекс, преоб-
мембране митохондрии, тепло преобразуется в
разует АДФ в АТФ. Это другой вариант - альтер-
механическую работу (изменение объема V) и
биохимическую работу (преобразование АДФ в
нативный выведению ионов Н+ через протонные
АТФ) с накоплением электрического потенциала
помпы в межмембранное пространство с после-
как промежуточной стадией этого процесса, а на
дующим их возвращением в матрикс митохон-
частях цикла, ограничивающих площади S3 и S4,
дрии через АТФ-синтазы - та же функциональ-
митохондрия поглощает тепло (из внешнего ис-
ность, направленная на выработку АТФ, но с раз-
точника).
ной скоростью. Поскольку в обоих случаях
возможно преобразование тепла в АТФ, возника-
Используя принцип, состоящий в том, что те
ет вопрос: всегда ли обратное преобразование
части цикла, которые обходятся по часовой
должно сопровождаться расщеплением АТФ
стрелке, относятся к (физиологически функцио-
внутри самой митохондрии, когда «ей надо со-
нальной) работе, а части, обходящиеся против ча-
греться»?
совой стрелки, относятся к поглощаемой энергии
Как известно, независимо от места расщепле-
(в основном в форме тепла), можно написать
ния АТФ приблизительно 50% энергии преобра-
формулу для коэффициента полезного действия
зуется в тепло. Если происходит эта трансформа-
митохондрии
ция, то с термодинамической точки зрения полу-
A S
+
S
R
S
+
S
чается, что митохондрия в состоянии поглощать
1
2
1
2
η=
=
⋅
=
тепло, вырабатывая АТФ, и при функциональной
(−Q)
S
3
+
S
4
(
P
C C )μ
V
S
3
+
S
4
необходимости часть АТФ (внутри нее же) рас-
В отличие от обычного термодинамического
щепляется с выделением тепла (теплопродукция,
коэффициента полезного действия, работа здесь
внутреннее энергопотребление митохондрии).
делится не на подводимое к системе тепло, а на
Оставшаяся часть АТФ, проходя через внешнюю
разность подводимого и отводимого тепла (-Q),
мембрану митохондрии в цитоплазму, использу-
при таком определении коэффициент полезного
ется клеткой для обеспечения биохимических ре-
действия идеальной тепловой машины Карно [14]
акций. Это практически все процессы, протекаю-
оказывается равным единице (100%) - это соот-
щие в многоклеточном организме, кроме тех, ко-
носится с современными исследованиями [15], в
торые происходят внутри самой митохондрии.
которых авторы указывают, что тепло, отдавае-
Косвенно это подтверждается тем, что хладно-
мое митохондрией вовне, нельзя считать поте-
кровные существа могут впадать в анабиоз, похо-
рянным с учетом теплопродукционной функции
же, что в этом случае митохондрии работают «са-
митохондрии.
ми на себя», а все остальные процессы жизне-
Обратный цикл митохондрии. Представленный
обеспечения в организме
«замирают» до
выше материал дает только «однонаправленное»
наступления подходящих внешних условий.
представление об энергетической жизни мито-
Кроме того, имеются эффекты, происходящие
хондрии в системе внутриклеточной среды, т. е.
в живой природе, которые в еще большей степени
основные энергетические переходы осуществля-
парадоксальны в смысле
«перепроизводства»
ются по циклу «механическая энергия + тепло -
АТФ митохондриями и недопотребления этой
электрохимический потенциал - АТФ из АДФ -
«энергетической валюты» во внешней по отно-
механическая энергия + тепло» с описанными
шению к митохондрии среде, например, это эф-
выше промежуточными этапами. Однако термо-
фект возникновения мышечной дрожи у всех теп-
динамический подход приводит к естественному
локровных существ! Мышечная дрожь - это ме-
вопросу о возможности обращения этого цикла
ханическая работа, возникающая, когда живое
подобно тому, как это происходит в технике. Для
существо мерзнет, и на нее непременно расходу-
подхода к ответу на этот вопрос необходимо до-
ется АТФ. Возникает вопрос: «Зачем тратить
полнить картину жизнедеятельности мтДНК но-
энергетическую «валюту» на «бесполезную» рабо-
выми (важными, на наш взгляд) функциями и
ту по сокращению миоцитов, если ее можно не-
учесть определенные факты.
посредственно преобразовать в тепло?» Или все
БИОФИЗИКА том 64
№ 6
2019
1158
ТАТЕВОСЯН, БУНЯКИН
ся, и ТД-ЭХ-цикл митохондрии сводится к обхо-
ду площадей S4 и S2 (малый цикл митохондрии).
Возможно ли обращение малого цикла, и какими
физическими процессами должны сопровож-
даться энергетические переходы на обращенном
малом цикле митохондрии?
В приводимых ниже рассуждениях дается
предположение о принципиальной возможности
преобразования механической энергии (внешне-
го воздействия со стороны мышц, проявляюще-
гося в виде колебаний давления и объема - дро-
жи) внутри мышечных клеток и соответственно
внутри матрикса митохондрии. В основу предпо-
лагаемого эффекта положен квантовый ток в по-
лимерных замкнутых молекулах мтДНК (рис. 3).
Для обоснования возможности преобразова-
ния механической энергии в тепло внутри мат-
рикса митохондрии сахарофосфатный остов цепи
мтДНК рассматривается в качестве нанопрово-
дов, а сами полимерные молекулы кольцевидной
Рис. 3. Кольцевая мтДНК (показан процесс репродук-
(замкнутой) мтДНК - как цилиндры малого ра-
ции). Диаметр кольца по одной спирали РНК - поряд-
диуса δ и значительно превосходящей длины
ка 2 нм, средний диаметр одной РНК - порядка 0,2 нм.
(δ << L) «нанопроводы», внутри которых содер-
Длина спирали РНК - порядка 100 нм = 0,1 мкм. Дли-
жится электронный газ с плотностью заряда ρ(x),
на одного витка спирали - порядка 10 нм.
равномерно распределенный по сечению, x - ко-
ордината вдоль нанопровода).
Напряженность электрического поля E(x0)
же эта актин-миозиновая работа не совсем беспо-
лезна и может идти, например, на согревание ми-
(осевая составляющая), создаваемая этим рас-
тохондрий без расходования АТФ, что выглядит
пределенным зарядом на оси нанопровода в точ-
невероятно. Но тогда возникает следующий во-
ке с координатой x0, вычисляется следующим ин-
прос: «Существует ли внутри митохондрии меха-
тегралом (в нем ε0 - электростатическая постоян-
низм выработки тепла без расщепления АТФ»?
x − x
0
Исходя из представленной термодинамиче-
ная в стандарте СИ,
cosα =-
,
угол
2
2
ской аналогии для плазмоподобной внутримат-
(
x − x
)
+δ
0
ричной среды матрикса митохондрии, данный
α - между осью нанопровода и направлением из
вопрос сводится к тому, что петли (на рис. 1),
точки x0 на его оси в точку с координатой x, рас-
ограничивающие площади S1 и S3, отбрасывают-
положенную на расстоянии δ от оси):
L
δ
ρ(x)
1
x - x
0
E(x
0
)
=-
dx
2πδdδ=
∫
∫
2
2
2
2
4πε
(x - x
)
+δ
0
−L
0
0
(x - x
0
)
+δ
L
⎛
⎞
1
1
1
=-
(
x - x ρ x)⎜
-
⎟dx
=
∫
0
⎜
2
2
⎟
2
ε
x - x
0
−L
⎝
0
(x − x
0
)
+δ
⎠
L
2
⎛
⎞
(
x - x ρ x)
0
1
δ
=-
⎜
1+
-1
⎟
dx
∫
2
2
2
2
ε
(x - x
)
0
−L
⎝
0
⎠
(
x - x
0
)
+δ
δ
Рассматривается асимптотика
→
0,
затем интегрируется по частям, с использованием следу-
x - x
0
ющего тождества:
2
y
2
2
−δ-
y
+δ
2
2
2
2
2
2
2
d
δ+
y
+δ
y
y
+δ
−δ
−δ
y
+δ
ln
=
=
2
2
2
2
2
2
2
dy
y
y
δ+
y
+δ
δ+
y
+δ
y y
+δ
(
)
БИОФИЗИКА том 64
№ 6
2019
МИТОХОНДРИАЛЬНЫЙ ТЕРМОДИНАМИЧЕСКИЙ И ЭЛЕКТРОХИМИЧЕСКИЙ ЦИКЛЫ
1159
Таким образом, напряженность выражается следующим образом:
x=L
2
L
2
2
ρ(x)dx
δ+
(x - x
)
+δ
δ
ρ(x)δ
0
E(x
0
)
=-
=
ln
−
∫
2
2
4ε
4ε
x - x
0
−L
(x − x
) (x − x
)
+δ
0
0
0
0
x L
L
2
2
δ
δ+
(x - x
0
)
+δ
−
ρ′(x)ln
dx
∫
4ε
x - x
0
-L
0
Первым слагаемым можно пренебречь при
тегрирование лишь по β-окрестности особой точ-
δ << L, а оставшееся выражение имеет интегри-
ки при условии дифференцируемости функции ρ
руемую особенность при x → x0. Выделение ее (ин-
в ней) дает:
x +0
β
2
x
0
+β
x
−
x
x
−
x
x
−
x
ρ′( x )δ
0
0
ρ′( x )δ
0
0
0
E(x
)
≈
ln
dx
=
ln
d
=
0
∫
∫
4
ε
2δ
2ε
2
δ
2δ
0
x
0
-
β
0
x
0
−
β
x=x
0
+β
2
ρ′( x0)δ
x
−
x
⎛
x
-
x
⎞
0
0
=
⎜ln
-
1
⎟
⎜
⎟
2ε
2δ
2δ
0
⎝
⎠
x=x
−β
0
Добавление третьего условия β = 2δ (к сделан-
eρ
кулы мтДНК) является
a
=δ
, как видно, эта
δ
ным ранее двум асимптотикам
→
0
и
m
ε
e
0
x
-
x
0
величина пропорциональна квадратному корню из
δ << L), основанного на том, что на осевую со-
величины электронной плотности.
ставляющую напряженности электрического по-
Оценка порядка величины a, если принять ра-
ля существенно не влияет распределение заряда,
отстоящее на расстояние от данного сечения,
диус нанопровода равным δ = 10-10 м, электрон-
большее диаметра нанопровода, дает выражение:
ную плотность ρ = 108 Кл/м3 (порядка внутри-
2
ρ'(x
0
)δ
атомной), ε0 ≈ 8,854304 · 10-12 (Кл2 · с2)/(кг · м3),
E(x
)
≈
0
ε
0
дает значение a ≈ 1,41 · 105
м/с. Это составляет ве-
личину порядка процента от скорости внутри-
Хотя приведенные выкладки и являются при-
атомного движения электронов и, учитывая, что
ближенными в смысле сделанных допущений, но
движение зарядов по нанопроводу нестационар-
полученное выражение естественно - напряжен-
ное (типа одномерных волн сжатия-разряжения
ность пропорциональна градиенту плотности за-
в обычном газе), скорость его может иметь такой
ряда (с обратным знаком). Теперь можно запи-
же порядок.
сать уравнения движения электронного газа (ти-
па уравнений Эйлера для обычного газа), считая
Итак, в предположении, что степень полимери-
уже плотность заряда ρ(x,t) нестационарной:
зации (количество однотипных звеньев в цепочке,
имея в виду мтДНК) может достигать значений по-
2
2
∂V
∂V
ρ
ρδ
∂ρ
a
∂ρ
+V
=
E
=
-
=-
,
рядка 100 и более, длина ее порядка L ≈ 10-7 м, по-
∂t
∂x
ρ
ρ ε ∂x
ρ∂x
m
m
0
лучается следующее: волны колебания электрон-
ной плотности могут иметь частоту порядка ν =
где ρm - массовая плотность заряженных элемен-
a/L ≈ 1,41 · 1012 Гц и менее. Этой частоте соответ-
тов полимерной молекулы ДНК (отнесенная к
Дж
ствует энергия квантов E = ħν ≈ 1,48 · 10-22
ρ
e
длине вдоль нанопровода)
=
, где e
≈
(около 10-3 эВ излучается в миллиметровом диапа-
ρ
m
m
e
зоне), где ħ ≈ 1,0544·10-34 Дж·с - постоянная
1,60202·10-19 Кл - элементарный заряд, me ≈
Планка. Это близко к нижней границе инфракрас-
9,1083·10-31 кг
- масса электрона, отсюда
ного излучения (мягкое тепло), т. е. такая среда
2
2
2
δ
ρ
2
δ
e
(матрикс митохондрии, заполненный замкнутыми
=
a
=
ρ
полимерными молекулами мтДНК) принципиаль-
ερ
ε
m
0
m
0
e
но может быть рабочим органом при непосред-
Таким образом, аналогом скорости звука в элек-
ственном преобразовании механической энергии в
тронном газе (внутри замкнутой полимерной моле-
тепло (колебание молекул в основном - попереч-
БИОФИЗИКА том 64
№ 6
2019
1160
ТАТЕВОСЯН, БУНЯКИН
Рис. 4. График изменения потенциала Гиббса G в термодинамическом/электрохимическом цикле митохондрии.
ные, как следствие - деформация замкнутых по-
ется максимизация времени пребывания системы
лимерных цепочек мтДНК и возбуждение кванто-
внутри области допустимых значений регулируе-
вого кольцевого тока в них).
мых переменных (первичных и вторичных по-
Таким образом, митохондрия осуществляет
требностей), мы убеждены в том, что с учетом
клеточное дыхание, периодически увеличивая
ограничений, накладываемых законами иерархи-
объем и уменьшая давление или наоборот (как и
ческой термодинамики, наибольшая степень тер-
легочная альвеола). Не вызывает сомнения то,
модинамического совершенства процесса мито-
что увеличение и уменьшение в объеме заложено
хондреального дыхания осуществляется в состоя-
в ее мтДНК, но не только в качестве обеспечения
нии F-I, которому соответствуют максимальная
выработки АТФ. Для самой митохондрии с пози-
энтропия и минимальная энергия Гибса (G), ха-
ции термодинамической целесообразности пре-
рактеризующие максимальное термодинамиче-
образование энергии в АТФ в состоянии F-II не
ское равновесие митохондрии с окружающей
является главенствующей функцией, которую ей
приписывают большинство исследователей [16].
средой (цитоплазмой). Такой подход согласуется
Существуют модели, использующие термодина-
с законами супрамолекулярной термодинамики,
мический подход для описания процесса жизне-
изучающей квазизамкнутые квазиравновесные
деятельности митохондрии, как например, в ра-
системы, которая утверждает, что в процессе он-
боте [17], где автор указывает на термодинамиче-
тогенеза (а также филогенеза и эволюции в це-
скую основу процессов, протекающих в матриксе
лом) каждая иерархическая структура биомира
митохондрии, однако не говорит о характере их
стремится к состоянию с минимальной удельной
цикличности.
энергией Гиббса [10,19,20].
На рис.
4 отражена динамика изменения
ЗАКЛЮЧЕНИЕ
потенциала Гиббса во время ТД-ЭХ-цикла. Из
Между характерными физиологическими со-
рисунка видно, что в состоянии F-I, несмотря на
стояниями ТД-ЭХ цикла митохондрии выделены
то что энтропия максимальная, система находит-
четыре функциональных состояния: F-I, F-II,
ся в наиболее устойчивом положении поскольку
F-III, F-IV, между которыми осуществляются
энергия Гиббса и эксергия минимальны (< 0), а
четыре перехода: S1, S2, S3 и S4. ТД-ЭХ-цикл ми-
при F-III энтропия минимальная, а сама система
тохондрии представляет собой непрерывную по-
наименее устойчива, поскольку энергия Гиббса и
следовательность переходов между квазистатиче-
эксергия максимальны (> 0), т. е., по мере удале-
скими состояниями, при которых циклически
ния от более устойчивого квазиравновесного со-
изменяются квазиравновесные состояния, являя
стояния (F-I) к менее устойчивому квазиравно-
собой типичную когерентную устойчивость не-
весному состоянию (F-III) диссипативная струк-
равновесных процессов, о которых упоминал
тура самой митохондрии движется от
И. Пригожин [18].
универсального функционального состояния (F-
Исходя из того, что фундаментальным прин-
I) к уникальному (F-III) в направлении богатства
ципом функциональной целесообразности явля-
и разнообразия (эволюционный скачок!), что на-
БИОФИЗИКА том 64
№ 6
2019
МИТОХОНДРИАЛЬНЫЙ ТЕРМОДИНАМИЧЕСКИЙ И ЭЛЕКТРОХИМИЧЕСКИЙ ЦИКЛЫ
1161
ходится в полном соответствии с высказывания-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
ми И. Пригожина (21).
Настоящая работа не содержит описания ка-
ких-либо исследований с использованием людей
Полагаем, что в состоянии обратного функци-
и животных в качестве объектов.
онального цикла F-I ↔ F-IV митохондрия может
повторно поглощать, выделяющееся тепло, вновь
создавая суперкомплекс электрон-транспортной
СПИСОК ЛИТЕРАТУРЫ
цепи с локальным сопряжением дыхания и фосфо-
1.
P. Mitchell, Nature 191, 144 (1961).
рилирования (модель Вильямса), т. е. митохон-
2.
R. J. Williams, J. Theor. Biol. 1, 1 (1961).
дрия в функциональном цикле F-I ↔ F-IV, спо-
3.
И. М. Солодовникова, В. И. Юрков, А. А. Тонь-
собна регулировать теплопродукцию и управлять
шин и Л. С. Ягужинский, Биофизика 49 (1), 47
ею, преобразуя тепло в АТФ.
(2004).
С одной стороны, может показаться, что в та-
4.
С. В. Нестеров, Ю. А. Скоробогатова и Л. С. Ягужин-
ком случае, к примеру, сокращающиеся миоциты
ский, Биофизика 59 (6), 1113 (2014).
не только теряют часть энергии АТФ в виде тепла,
5.
Ю. А. Скоробогатова, Дисс
магистра (МФТИ,
но и сами митохондрии сопряженно расщепляют
М., 2014).
часть «лишней» АТФ, чтобы самим «согреться».
6.
В. И. Юрков, Автореферат дисс
канд. биол. наук
Однако такой подход представляется нерацио-
(МГУ, М., 2008).
нальным (неэкономичным), следовательно,
7.
B. Chance, G. Williams, W. Holmes, and J. Higgins, J.
должна существовать модель, при которой нагрев
Biol. Chem. 217 (1), 439 (1955).
матрикса митохондрии производится за счет дру-
8.
A. P. Halestrap, P. T. Quinlan, D. E. Whipp, and
гого эффекта, не связанного с расщеплением
A. E. Armston, Biochem. J. 236, 779 (1986).
АТФ.
9.
V. P. Skulachev, J. Bioenerg. Biomembr. 31 (5), 431
(1999).
Исходя из термодинамической аналогии с теп-
ловой машиной, в которой тепло из источника
10.
Г. П. Гладышев, Супрамолекулярная термодинами-
ка - ключ к осознанию явления жизни, 2-е изд. (Ин-
частично преобразуется в механическую работу, а
ститут компьютерных исследований, М.- Ижевск,
частично рассеивается в окружающую среду, сле-
2003).
дует рассматривать то, что на обратном термоди-
11.
С. Н. Алексеенко, А. С. Татевосян и А. В. Бунякин,
намическом цикле охлаждения тепло перемеща-
Кубанский науч. мед. вестн. 25 (5), 123 (2018). DOI:
ется, при этом расходуется меньшая механиче-
10.25207/1608-6228-2018-25-5-123-135.
ская работа, чем само перемещаемое тепло. Этот
12.
В. И. Слесарев, Химия. Основы химии живого (Химиз-
освоенный в технике принцип предполагает на-
дат, Санкт-Петербург, 2015).
личие источника и «утилизатора» тепла. В каче-
13.
Ю. Б. Румер и М. Ш. Рывкин, Термодинамика, ста-
стве последнего нами рассматривается окружаю-
тистическая физика и кинетика, 2-е изд., испр. и
щая цитоплазма и межклеточное пространство,
доп. (Изд-во Носиб. ун-та, Новосибирск, 2000).
но поскольку природа рациональна и экономич-
14.
S. Carnot, Réflexions sur la puissance motrice du feu et sur
на, то в теплокровных организмах должны суще-
les machines propres à développer cette puissance (Gauthi-
ствовать механизмы сохранения тепла по кумуля-
er-Villars, Imprimeur-Libraire, Paris, 1878).
тивному типу, одним из которых может являться
15.
J.-S. Kang, Biophys. Rep. 4 (2), 63
(2018). DOI:
сокращение миоцитов (дрожь). Таким образом
10.1007/s41048-018-0054-2.
колебание давления в цитоплазме, включая плаз-
16.
I. P. Krasinskaya, V. N. Marshansky, S. F. Dragunova,
моподобную среду матрикса митохондрии и, как
and L. S. Yaguzhinsky, FEBS Lett. 167 (1), 176 (1984).
следствие, колебание закольцованных молекул
17.
C. F. Matta and L. Massa, Biochemistry 54 (34), 5376
мтДНК (с уже большей частотой), допускает воз-
(2015). DOI: 10.1021/acs.biochem.5b00834.
никновение в них кольцевого квантового тока,
который закономерно сопровождается теплопро-
18.
И. Пригожин и Г. Николис, Самоорганизация в не-
равновесных системах (Мир, М., 1979).
дукцией, поддерживающей гомеостаз.
19.
А. Мюнстер, Химическая термодинамика (УРСС, М.,
2002).
КОНФЛИКТ ИНТЕРЕСОВ
20.
А. В. Панов, Функциональная биоэнергетика (Create
Space, 2016). 29356.
Авторы заявляют об отсутствии конфликта
21.
И. Пригожин и И. Стенгерс, Порядок из хаоса
интересов.
(Прогресс, М., 1986).
БИОФИЗИКА том 64
№ 6
2019
1162
ТАТЕВОСЯН, БУНЯКИН
Mitochondrial Thermodynamic and Electrochemical Cycles
(in the Direct and Reverse Direction)
A.S. Tatevosyan* and A.V. Bunyakin**
*Kuban State Medical University, Ministry of Health of the Russian Federation,
ul. Mitrofana Sedina 4, Krasnodar, 350063 Russia
**Kuban State University, ul Stavropolskaya 149, Krasnodar, 350040 Russia
A meta-analysis of the functional states of mitochondria was carried out with mathematical modelling. The
mathematical model is based on the change of the direction and speed of thermodynamic and electrochem-
ical parameters (pressure, volume, temperature, entropy, Gibbs potential, exergy ...), between which there are
links. Using the fundamental principle of supramolecular functional interaction, a mathematical model of
the thermodynamic and electrochemical cycle of mitochondria was deduced. This model describes thermo-
dynamic and electrochemical reactions that occur in the “mitochondrial matrix - inner membrane - inter-
membrane space” system and are accompanied by accumulation of electrochemical potential, charge sepa-
ration (ionization). We studied four functional states of mitochondria when the velocity and direction of the
movement of electrons along the respiratory chain changed in the thickness of its inner membrane due to
f luctuations in heat flowing. The thermodynamic and electrochemical cycle of mitochondria is a continuous
succession of transitions between quasi-static states, in which quasi-equilibrium states recurrently change
which are considered as a typical coherent stability criterion for non-equilibrium processes, as stated by
I. Prigogine. The mathematical model of the thermodynamic behavior of mitochondria and the limitations
imposed by laws of physical and chemical thermodynamics provide the basis for a statement that the highest
degree of thermodynamic perfection in the process of mitochondrial respiration corresponds to a state in
which Gibbs energy and exergy are minimal and entropy maximum, that enables us to use this functional state
as the main (basic) one.
Keywords: thermodynamic and electrochemical cycle of mitochondria, thermal potential, energy conversion of
ADP to ATP, ring quantum current in a closed mtDNA molecule
БИОФИЗИКА том 64
№ 6
2019