БИОФИЗИКА, 2020, том 65, № 1, с. 61-68
МОЛЕКУЛЯРНАЯ БИОФИЗИКА
УДК 57.07
ЭФФЕКТ ЗРИТЕЛЬНОЙ СТИМУЛЯЦИИ НА УРОВНИ ГАММА-
АМИНОМАСЛЯНОЙ КИСЛОТЫ И МАКРОМОЛЕКУЛ В ГОЛОВНОМ
МОЗГЕ ЧЕЛОВЕКА in vivo
© 2019 г. А.Н. Яковлев*, А. Манжурцев**, ***, П. Меньщиков *, М. Ублинский**, ***,
О. Божко***, Т. Ахадов***, Н. Семенова*, **, ***
*Федеральный исследовательский центр химической физики им. Н.Н. Семенова РАН, 119991, Москва, ул. Косыгина, 4
**Институт биохимической физики им. Н.М. Эмануэля РАН, 119334, Москва, ул. Косыгина, 4
***НИИ неотложной детской хирургии и травматологии, 119180, Российская Федерация,
Москва, ул. Большая Полянка, 22
E-mail: yakovlevalekcej@bk.ru
Поступила в редакцию 26.08.2019 г.
После доработки 19.09.2019 г.
Принята к публикации 19.09.2019 г.
GABA (гамма-аминомасляная кислота) - основной тормозной нейромедиатор в мозге - играет важную
роль в различных типах синаптической пластичности и патологии. В настоящей работе
для прижизненного исследования влияния видеостимуляции на уровень GABA в зрительной коре мозга
человека использован метод протонной магнитно-резонансной спектроскопии (1Н-МРС) и импульсная
последовательность MEGA-PRESS в двух модификациях: -GABAMEGA-PRESS и +GABAMEGA-PRESS.
Первая модификация позволяет получить 1Н-ЯМР-сигнал метиленовых протонов GABA с химическим
сдвигом δ = 3,01 м.д. (-GABA) без примеси сигнала макромолекул. Во второй модификации этот сигнал
(+GABA) является суперпозицией резонансов метиленовых протонов GABA и макромолекул. Измерено
также действие постоянной видеостимуляции на уровень N-ацетиласпартата и суммарный уровень глу-
тамата и глутамина. Постоянная видеостимуляция не изменяет содержание N-ацетиласпартата и глута-
мина в зрительной коре. По уменьшению интенсивности сигнала -GABA обнаружено статистически
значимое снижение уровня GABA, указывающее на инактивацию синтеза GABA. Выявлено отсутствие
статистически значимых изменений интенсивности резонанса +GABA, что может быть следствием воз-
действия видеостимуляции на макромолекулы. Для проверки этой гипотезы с помощью специально раз-
работанной импульсной последовательности инверсии-восстановления получен сигнал макромолекул
и показано, что его интенсивность нечувствительна к видеостимуляции, и отсутствие изменений ин-
тенсивности сигнала +GABA при активации следует отнести к маскирующему эффекту сигнала макро-
молекул.
Ключевые слова: 1H-МРС, GABA, MEGA-PRESS, макромолекулы, зрительная стимуляция.
DOI: 10.31857/S000630292001007Х
физико-химических процессов, обеспечивающих
Нарушения процессов, регулирующих кон-
перенос высвобождающихся из везикул нейроме-
центрацию основного тормозного нейромедиато-
диаторов в астроциты, преобразование нейроме-
ра γ-аминомаслянной кислоты (GABA), могут
диаторов в глутамин, транспорт глутамина в ней-
быть причиной развития церебральной патоло-
роны, где он превращается в глутамат [5]. В
гии (шизофрении [1], депрессивного расстрой-
ГАМКергических нейронах глутамат преобразу-
ства [2], тревожности [3], эпилепсии [4]).
ется глутаматдекарбоксилазой в GABA. Таким
Будучи основным тормозным нейромедиато-
образом, единственным предшественником
ром, GABA наряду с основным возбуждающим
GABA является глутамат, поэтому основной тор-
нейромедиатором глутаматом участвует в цикле
мозной нейромедиатор непосредственно связан с
возбуждающим. Концентрация GABA связана с
функцией коры: [GABA] коррелирует со скоро-
Сокращения: GABA - γ-аминомасляная кислота, МРС -
магнитно-резонансная спектроскопия, ИП - импульсная стью принятия решений [6], эффективность рас-
последовательность, NAA
- N-ацетиласпартат, tCr
-
познавания ориентации линий изображения пе-
фосфокреатин, Glx - сумма глутамина и глутамата, ЧСИ -
частотно-селективный импульс, SNR
- отношение
риферическим зрением тоже может быть соотне-
сигнал/шум.
сена с уровнем GABA
[7]. Эти данные
61
62
ЯКОВЛЕВ и др.
показывают, что прижизненные исследования
tCr. Оставшийся редактированный сигнал при 3
содержания GABA в различных зонах мозга зна-
м.д. является суперпозицией резонансов GABA,
чимы как для клинических целей, так и для полу-
макромолекул и гомокарнозина, поэтому в даль-
чения новой нейрохимической информации о
нейшем ИП с приведенным выше расположени-
механизмах биологических функций мозга.
ем ЧСИ мы будем называть +GABAMEGA-PRESS.
Единственным методом, позволяющим измерять
Вклад макромолекул в редактированный резо-
церебральные концентрации метаболитов in vivo,
нанс при использовании
+GABAMEGA-PRESS
является магнитно-резонансная спектроскопия
составляет 50%, а гомокарнозина - 16% [13]. ИП
(МРС). При помощи стандартных методик лока-
лизационной спектроскопии (импульсные по-
+GABAMEGA-PRESS наиболее часто применяет-
следовательности (ИП) PRESS [8] и STEAM [9]) в
ся для измерения уровня GABA, вероятно, в
мозге в зоне интереса можно определить концен-
предположении, что изменения интенсивности
трации ряда метаболитов, в том числе оценить со-
этого сигнала могут быть следствием изменения
держание глутамата, однако для измерения
исключительно GABA.
GABA такие спектры не пригодны: все резонансы
Из-за недостаточной селективности ЧСИ на
GABA (2CH2 - химический сдвиг δ = 3,01 м.д.;
δ⊕ = 1,9 м.д воздействует не только на триплет
3CH2 - δ = 1,89 м.д.; 4CH2 - δ = 2,28 м.д.) пере-
GABA, но и на сигнал метиленовых протонов
крыты интенсивными сигналами других метабо-
макромолекул при δ = 1,7 м.д., связанных J-взаимо-
литов, присутствующих в мозге в концентрациях,
действием с протонами макромолекул при δ = 3 м.д.
на порядок превышающих концентрацию
с константой J = 7,8 Гц. Это обуславливает присут-
GABA (в сером веществе мозга [GABA] ~2,8 мМ,
ствие в спектрах +GABAMEGA-PRESS наличие сиг-
в белом - ~0,3 мМ [10]). К мешающим определе-
нала макромолекул. Однако если в ⊖-серии ис-
нию GABA сигналам относятся сигналы N-аце-
пользовать ЧСИ на δ⊖ = 1.5 м.д., можно исключить
тиласпартата (NAA, 2CH3 - δ = 2,01 м.д.), креати-
сигнал макромолекул и получить сигнал GABA с
на и фосфокреатина (tCr, N(CH3) - δ = 3,03 м.д.),
минимальной примесью (гомокарнозин) [14]. Та-
кой вариант ИП MEGA-PRESS мы в дальнейшем
глутамина + глутамата (Glx, 4CH2 - δ = 2,4 м.д.)
будем называть -GABAMEGA-PRESS.
[11].
Для определения уровня GABA создана ИП
Таким образом, в зависимости от расположе-
MEGA-PRESS [12], которая позволяет из сигнала
ния селективных импульсов в
⊖-серии
при δ = 3,01 м.д. вычесть интенсивный синглет-
сигнал GABA может быть зарегистрирован либо с
ный сигнал tCr. В этой ИП используется эффект
макромолекулами (+GABA), либо без макромо-
спин-спинового (J)-взаимодействия и зависи-
лекул (-GABA).
мость фазы сигналов в мультиплете от времени
эха ТЕ (J-эволюция). В молекуле GABA протоны
Подавление макромолекул приводит к сниже-
соседних метиленовых групп при δ = 1,89 м.д. и
нию интенсивности сигнала на 3 м.д. и создает
δ = 3,01 м.д. связаны J-взаимодействием, в ре-
сложности при его обработке. Это объясняет ча-
зультате которого резонанс при 3,01 м.д расщеп-
стое использование ИП +GABAMEGA-PRESS для
ляется в триплет с константой J = 7,35 Гц. J-эво-
характеристики содержания GABA in vivo в локу-
люция вызывает расфазирование триплета, кото-
сах мозга человека. В состоянии покоя этим мето-
рый при ТЕ = 1/2J, превращается в триплет с
дом установлено, что вклад макромолекул в сум-
внешними компонентами в противофазе к цен-
марный сигнал одинаков в пределах ошибки в
тральной компоненте. Применение частотно-се-
разных корковых зонах (несмотря на разную ин-
лективного импульса (ЧСИ) на δ = 1,9 м.д рефо-
тенсивность сигнала +GABA или сигнала макро-
кусирует J-эволюцию сигналов триплета, и он
молекул) и сохраняется со временем [15]. Обнару-
оказывается сфазированным. Очевидно, что вли-
жено большее содержание макромолекул в сером
яние на J-эволюцию сигнала GABA не затрагива-
веществе, чем в белом [16]. Однако неясно, что
ет сигнал tCr, перекрывающий интересующий
происходит в случае патологии или при нейроак-
нас триплет. В ИП MEGA-PRESS, в отличиe от
тивации. В ответ на двигательную нагрузку [6],
стандартной ИП PRESS, проводится набор двух
активацию когнитивных функций [17], физиче-
серий спектров: В ⊕-серии в ИП PRESS добавля-
скую активность [18] зафиксированы изменения
ют ЧСИ на δ⊕ = 1,9 м.д. В ⊖-серии ЧСИ прила-
интенсивности сигнала
+GABA. Существуют
гается симметрично относительно сигнала
лишь четыре исследования GABA в зрительной
воды на δ⊖ = 7,6 м.д. и не затрагивает область
коре при видеостимуляции. В двух из них измеря-
сигналов церебральных метаболитов, поэтому
ли сигнал -GABA и обнаружили его снижение
спектр ⊖-серии представляет собой стандартный
[19,20]. При измерении сигнала +GABA отмеча-
PRESS-спектр. Вычитание спектров одной серии
ется как увеличение [21], так и уменьшение его
из спектров другой серии элиминирует сигнал
интенсивности [22].
БИОФИЗИКА том 65
№ 1
2020
ЭФФЕКТ ЗРИТЕЛЬНОЙ СТИМУЛЯЦИИ
63
здоровы и ознакомлены с процедурой исследо-
вания.
Зрительная стимуляция. Исследование состоя-
ло из двух блоков одинаковой длительности
(9 мин 36 с): «покоя» и «стимуляции». Для исклю-
чения незапланированной зрительной активации
во время «покоя» освещение в аппаратной томо-
графа было выключено заблаговременно, а испы-
туемого просили закрыть глаза. Затем испытуе-
мого просили открыть глаза, включали экран (си-
стема SensaVue) и через систему зеркал
предъявляли зрительную стимуляцию - шахмат-
ная доска, мерцающая с частотой 8 Гц. Внимание
испытуемого оценивали по появляющимся с раз-
ными промежутками времени красным крестам
на черном фоне (пять раз за две минуты), которые
испытуемый должен был считать и на каждое пя-
тое появление нажимать на кнопку, вызывавшую
звуковой сигнал в аппаратной томографа.
Получение диагностических изображений с
Рис. 1. Расположение спектроскопического вокселя.
помощью магнитно-резонансной томографии. Все
исследования выполнены на магнитно-резонанс-
Таким образом, участие основного тормозного
ном томографе Achieva dStream
3T (Philips,
Нидерланды) при помощи приемной катушки
нейромедиатора в регуляции процессов возбуж-
dStream HeadNeckSpine coil. Отсутствие очагово-
дения-торможения при нейроактивации изучено
го поражения и объемного процесса в головном
явно недостаточно. Имеющиеся данные получе-
мозга устанавливали по данным диагностической
ны с применением разных модификаций ИП
магнитно-резонансной томографии (Т1-, Т2-
MEGA-PRESS, что, возможно, приводит к неод-
нозначным результатам. Причиной может быть
взвешенные изображения, изображения с ослаб-
влияние нейроактивации не только на GABA, но
лением сигнала свободной жидкости и диффузи-
и на макромолекулы. Присутствие сигнала мак-
онно-взвешенные изображения). Спектроскопи-
ромолекул может маскировать эффекты измене-
ческий воксел располагали в зрительной коре
ния GABA [23]. Отсюда возникает необходимость
(рис. 1), которую визуализировали по данным
исследовать влияние видеостимуляции на интен-
функциональной магнитно-резонансной томо-
графии [24].
сивность сигналов +GABA и -GABA в 1Н-МР-
спектрах зрительной коры мозга человека, выде-
Протокол получения данных методом магнитно-
лить сигнал макромолекул, оценить влияние ней-
резонансной спектроскопии. В ИП -GABAMEGA-
роактивации на интенсивность сигнала макромо-
PRESS применяли следующие параметры: время
лекул и определить, маскирует ли этот сигнал
эха (TE) = 80 мс, задержка на релаксацию (TR) =
возможные изменения GABA.
2 с, число накоплений (NSA) = 288 (по 144 накоп-
ления на каждую серию), ЧСИ длительностью
В настоящей работе впервые исследовано вли-
20 мс применяли на δ⊕= 1,9 м.д. в ⊕-сериях и
яние видеостимуляции на концентрацию макро-
молекул. Получены значения уровня GABA и
δ⊖ = 1,5 м.д. в ⊖-сериях. В ИП +GABAMEGA-
суммарного уровня глутамина и глутамата в усло-
PRESS ТЕ = 68 мс, ЧСИ длительностью 14 мс рас-
виях постоянной видеостимуляции с использова-
полагали на δ⊕= 1.9 м.д. и δ⊖ = 7,5 м.д. J-модуля-
нием ИП -GABAMEGA-PRESS.
ция изменяет вид мультиплета GABA в
области 3 м.д. в разных сериях. Размер спектро-
скопического воксела для спектров макромоле-
МЕТОДЫ
кул и -GABA: 20 × 40 × 30 мм3, для +GABA -
25 × 45 × 35 мм3.
Объекты исследования. В исследовании при-
няли участие три группы добровольцев. Группа
Выделение сигнала макромолекул. Резонансы,
для исследования -GABA состояла из 16 человек
принадлежащие макромолекулам, имеют низкое
(средний возраст 29 ± 7 лет); измерение +GABA
время продольной релаксации Т1 [25]. Это дает
проводили в группе из 8 человек (средний возраст
возможность выделить сигнал макромолекул при
25 ± 3 лет); исследование макромолекул - в груп-
δ = 3 м.д., используя инвертирующий импульс с
пе из 16 человек (25 ± 7 лет). Все пациенты были
последующим восстановлением макроскопиче-
БИОФИЗИКА том 65
№ 1
2020
64
ЯКОВЛЕВ и др.
ской намагниченности [25]. Для исключения сиг-
налов GABA и гомокарнозина в области 3 м.д.
применяли дополнительный инвертирующий
импульс блочной формы шириной 50 Гц и време-
нем инверсии TI = 530 мс. Этот импульс исполь-
зовали как предварительный в обеих модифика-
циях ИП MEGA-PRESS - для получения сигнала
макромолекул в
+GABAMEGA-PRESS и для
оценки качества подавления сигналов метаболи-
тов в -GABAMEGA-PRESS. Оценку остаточного
сигнала в последнем случае проводили, сравни-
вая отношение сигнал/шум (SNR) в сигнале на
3 м.д. с макромолекулами и без них. В отношении
сигнал/шум оценка сигнала производится как
разница максимального и минимального значе-
ния в области δ = 2,8-3,5 м.д., а шум как стан-
дартное отклонение среднего значения интен-
сивности шумовых дорожек в области 11-12 м.д.
или 12-13 м.д после вычетания полиноминаль-
ной функции (n = 2) из спектра в каждой из этих
областей. [26].
Обработка спектральных данных. Все спектры
обрабатывали в программе GANNET [26] на базе
MATLAB. Программа автоматически подбирает
параметры для аппроксимации резонансов одной
гауссовой линией. Результатом обработки явля-
лись значения интенсивностей сигналов +GABA,
-GABA, макромолекул а также GLX, tCr (при об-
работке
⊖-серии в ИП -GABAMEGA-PRESS).
Погрешность аппроксимации резонансов оцени-
вали по выдаваемому программой параметру
CRLB [27]. Из дальнейшего анализа исключали
значения интенсивностей сигналов -GABA с по-
грешностью обработки > 20%, что уменьшило
размер выборки с 15 до 10 человек.
Для нормировки интенсивностей в качестве
референса использовали интенсивность tCr. Эф-
фект видеостимуляции на параметры +GABA/Cr,
Рис.
2. MEGA-PRESS спектр (усредненный по
-GABA/Cr, Glx/Cr и интенсивность сигнала
288 динамикам) с инвертирующим предварительным
макромолекул выявляли путем межгруппового
импульсом: (а) - с подавлением макромолекул, (б) -
сравнения данных, полученных в покое и в пери-
без подавления макромолекул. Как и ожидалось,
сигнал при δ = 3 м.д. отсутствует.
оде видеостимуляции. Наборы данных проверяли
на принадлежность к нормальному распределе-
нию с помощью критерия Шапиро-Уилка. До-
импульсом составляет 1,76, в то время как в спек-
стоверность эффекта оценивали по критерию
Вилкоксона для связанных выборок [28].
трах +GABAMEGA-PRESS с таким же импульсом
SNR = 7,42. Таким образом, наш метод обеспечи-
вает подавление сигнала -GABA в 4,2 раза.
РЕЗУЛЬТАТЫ
На рис. 2б представлен спектр +GABAMEGA-
Рис. 2а представляет спектр, полученный при
PRESS, полученный с использованием с инвер-
использовании ИП -GABAMEGA-PRESS с пред-
тирующего импульса, в котором сигнал на 3 м.д.
варительным добавлением инвертирующего им-
на 90% (см. обсуждение) соответствует сигналу
пульса. Спектр не содержит резонансных линий
макромолекул.
при δ = 3 м.д., сигнал подавлен. Для точной ин-
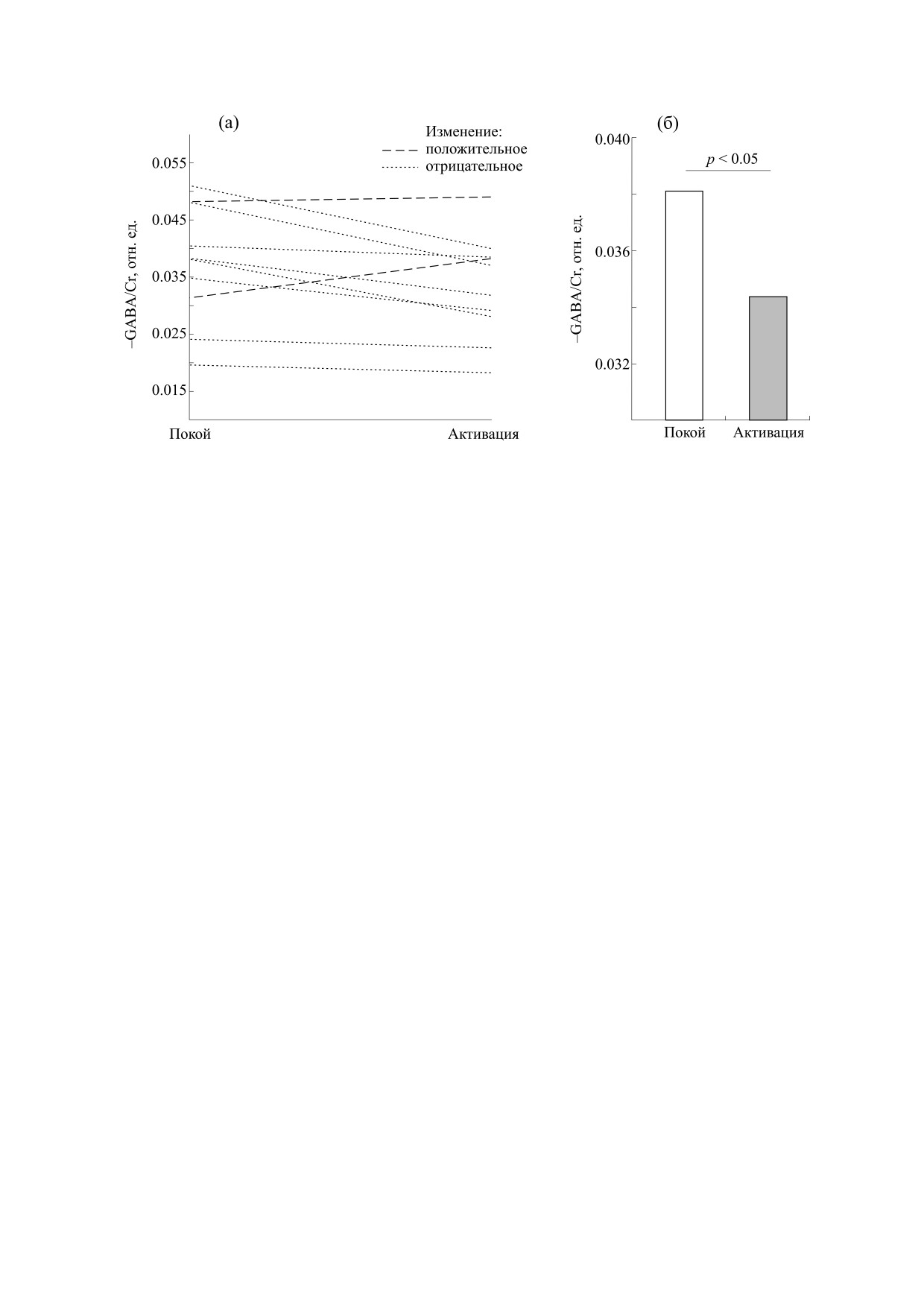
На рис. 3а изображены изменения интенсив-
терпретации результата проведена следующая ко-
ности сигнала -GABA, нормированного на креа-
личественная оценка. Среднее значение SNR в
тин, у каждого испытуемого. Видно, что в восьми
спектрах -GABAMEGA-PRESS с инвертирующим
случаях из десяти изменение отрицательное и
БИОФИЗИКА том 65
№ 1
2020
ЭФФЕКТ ЗРИТЕЛЬНОЙ СТИМУЛЯЦИИ
65
Рис. 3. (а) - Изменения относительной интенсивности сигнала -GABA в зрительной коре в состоянии покоя и
активации у каждого испытуемого; (б) - сравнение медиан -GABA/Cr в покое и при зрительной стимуляции.
лишь в двух случаях - положительное. При помо-
жение +GABA/Cr. Противоположный по знаку
щи рангового критерия Вилкоксона установлено,
эффект зрительного возбуждения, достоверный
что найденное изменение -GABA/Cr статистиче-
рост [+GABA], найден в работе [21]. Из приве-
ски достоверно. Из рис. 3б видно, что медиана из-
денных данных следует, что снижение концен-
менения -GABA/Cr составляет 12% (p < 0,05).
трации GABA наблюдается лишь в тех МРС-ис-
следованиях, в которых удается избавиться от
В результате зрительной стимуляции не выяв-
вклада макромолекул, тогда как измерения
лено статистически достоверного изменения
+GABA при зрительном возбуждении дают про-
отношений Glx/Cr, +GABA/Cr, а также макромо-
тиворечивые результаты [21,22].
лекул.
Мы не нашли изменения интенсивности сиг-
нала Glx под влиянием видеостимуляции. Одна-
ОБСУЖДЕНИЕ
ко измерения глутамата в поле 7Т, в котором раз-
решаются сигналы глутамата и глутамина, пока-
Снижение уровня GABA в ответ на зрительную
зали, что в ответ на зрительную стимуляцию
стимуляцию. Полученный нами результат соот-
содержание глутамата статистически значимо
ветствует данным измерений сигнала GABA на
увеличивается на 3%, если стимул предъявляют в
томографе с напряженностью постоянного маг-
течение 10 мин [27], и на 2% - при блочной пяти-
нитного поля В0 = 7 Тл; авторы показали, что
минутной стимуляции [30].
концентрация GABA в зрительной коре досто-
После высвобождения из везикул глутамат по-
верно снижается на 5% под влиянием десятими-
чти полностью попадает в астроциты, где из него
нутной видеостимуляции [19]. Из данных работы
синтезируется глутамин, превращающийся после
[19] следует, что даже увеличение спектрального
переноса в нейроны в глутамат [31]. Таким обра-
разрешения при В0 = 7 Тл не позволяет разделить
зом, взаимопревращение глутамата и глутамина в
сигналы GABA и макромолекул, и обнаруженный
цикле может быть причиной постоянства Glx. К
авторами эффект относится к сумме GABA и мак-
сожалению,
1Н-МРС-измерений глутамина
ромолекул, хотя вклад последних в этом случае
практически нет, за исключением исследования в
должен уменьшиться. Вероятно, по этой причине
поле 7Т с использованием зрительного возбужде-
эффект видеостимуляции меньше, чем обнару-
ния, под действием которого обнаружены проти-
женный нами по данным, полученным с помо-
вонаправленные и примерно одинаковые изме-
щью ИП -GABAMEGA-PRESS, когда сигнал мак-
нения (содержание глутамата увеличивается на
ромолекул элиминирован полностью. В отличие
0,22 ± 0,11 мМ, а содержание глутамина снижает-
от результатов, представленных в настоящей ра-
ся на 0,18 ± 0,11 мМ) [32]. Это означает, что изме-
боте, исследование влияния такой же стимуля-
нения суммарной концентрации глутамата и глу-
ции с помощью +GABAMEGA-PRESS МРС в поле
тамина под влиянием видеостимуляции не будут
3Т [22] обнаружило статистически значимое сни-
наблюдаться, что соответствует нашим данным.
БИОФИЗИКА том 65
№ 1
2020
66
ЯКОВЛЕВ и др.
Из постоянства концентрации Glx могут сле-
при текущем уровне погрешности. Согласно на-
довать два вывода. Либо при активации зритель-
шим измерениям, вклад макромолекул в суммар-
ной коры глутамат участвует исключительно в
ный сигнал при δ = 3 м.д. составляет ~63%, а
цикле нейромедиаторов и не включается в другие
GABA (и гомокарнозин) - соответственно 37%.
процессы (например, окисление, синтез GABA),
Расчет выполнен, исходя из сравнения средних
либо его концентрация пополняется за счет по-
значений интенсивностей сигналов макромоле-
вышения потребления глюкозы. В пользу первого
кул и -GABA. В нашей работе погрешность обра-
предположения свидетельствует обнаруженное
ботки сигнала +GABA составляет ~10% (ошибка
нами снижение GABA, которое может быть след-
обработки, полученная в Gannet), при этом вклад
ствием уменьшения расхода глутамата в синтезе
зафиксированного изменения GABA (12%) в сум-
GABA, а также следствием использования GABA
марный сигнал (+GABA) равен ~4%.
в качестве субстрата вместо глутамата в цикле
В то же время каждое третье измерение
Кребса.
-GABA превышает погрешность обработки в
Содержание N-ацетиласпартата в ответ на дли-
20% из-за низкого значения SNR. Суммируя вы-
тельную стимуляцию сохраняется. В нескольких
шесказанное, можно сделать вывод, что измере-
цитированных ранее работах, выполненных на
ние сигнала +GABA сопровождаются риском по-
томографах с В0 = 7 Тл, исследовали влияние зри-
тери эффекта. При этом следует иметь в виду низ-
кую интенсивность сигнала -GABA, что создает
тельной стимуляции на содержание метаболитов
серьезные трудности при его обработке, а это
в зрительной коре и, как и в нашем исследовании,
означает необходимость улучшения методики для
не зафиксировали статистически достоверного
изменения суммарного содержания NAA и N-аце-
повышения значения SNR.
тиласпартат-глутамата [21,30]. Однако измерение
Оценка качества подавления сигналов метабо-
суммарного содержания NAA и N-ацетиласпар-
литов. На результат измерения сигнала макромо-
тат-глутамата с помощью ИП MEGA-PRESS в
лекул может повлиять неполное подавление сиг-
поле 3Т показало, что такой же, как использован-
нала GABA и гомокарнозина. По остаточному
ный нами, способ видеостимуляции вызывает в
вкладу этих сигналов оценим возможность лож-
зрительной коре испытуемых снижение суммар-
ного постоянства сигнала макромолекул при ви-
ного содержания NAA и N-ацетиласпартат-глута-
деостимуляции. До подавления доля GABA в об-
мата на 60% и увеличение NAA на 18% [33]. Если
щем сигнале +GABA равнялась 37%; после по-
учесть, что вклад NAA и N-ацетиласпартат-глута-
давления она снизится до
12%, поскольку
мата в суммарный сигнал составляет 10% и ис-
интенсивность сигнала макромолекул не изме-
пользовать данные работы [33], то следует ожи-
нится, а интенсивность GABA снизится в 4,2 ра-
дать снижения интенсивности суммарного сиг-
за. Снижение остающейся части GABA на 12%
нала tNAA примерно на 10%. По нашим данным,
при видеостимуляции соответствует ~1,7% от из-
этот эффект зрительного возбуждения не наблю-
меренного сигнала макромолекул. Увеличение
дается. Возможно, расхождение данных с наши-
интенсивности сигналамакромолекул, превыша-
ми результатами объясняются разными способа-
ющее данное значение, могло бы быть обнаруже-
ми оценки концентраций: авторы работы [33] из-
но в настоящей работе в отсутствие других факто-
меряют не интегральные интенсивности
ров, влияющих на погрешность измерения. Та-
сигналов, а высоты их пиков.
ким образом, не более 2% от интенсивности
сигнала макромолекул могут объясняться нали-
Таким образом, наблюдаемое в нашем иссле-
чием остаточного сигнала -GABA в активиро-
довании постоянство tNAA в ответ на видеости-
ванной зрительной коре.
муляцию в основном совпадает с результатами
тех авторов, которые использовали стандартную
Наблюдаемые макромолекулы не участвуют в
оценку эффектов видеостимуляции на основе ин-
процессе нейротрансмиссии. Общую характери-
тегральной интенсивности сигнала tNAA.
стику резонансов макромолекул можно найти в
работе [34]. Достоверных биохимических данных
Маскировка эффекта более интенсивным сигна-
лом макромолекул. В настоящей работе мы пока-
о структуре макромолекул все еще нет. В 1Н-МР-
зали, что видеостимуляция влияет на интенсив-
спектрах мозга крысы in vivo сигналы этих соеди-
ность сигнала -GABA, тогда как интенсивность
нений отнесены согласно их химическим сдвигам
сигнала +GABA не изменилась. При этом интен-
к метильным и метиленовым протонам амино-
сивность сигнала макромолекул в ответ на стиму-
кислотных остатков цитозольных белков или
ляцию не изменяется. Отсюда можно сделать вы-
аполипопротеинов [34]. В частности, сигнал мак-
вод, что сигнал макромолекул маскирует измене-
ромолекул на 3 м.д. соответствует структурному
ния GABA. Причина в том, что сигнал
фрагменту с концевой группой -CH2NH2 лизина
макромолекул значительно превышает по интен-
(как и GABA). В настоящей работе в ответ на зри-
сивности сигнал GABA, это делает невозможным
тельную активацию изменение сигнала макромо-
зафиксировать небольшой эффект изменения
лекул на δ = 3 м.д. не выявлено. Это указывает на
БИОФИЗИКА том 65
№ 1
2020
ЭФФЕКТ ЗРИТЕЛЬНОЙ СТИМУЛЯЦИИ
67
отсутствие концентрационных, конформацион-
8. P. A. Bottomley, Ann. N.-Y. Acad. Sci. 508 (1), 333
ных и структурных изменений макромолекул, за-
(1987).
трагивающих фрагмент, резонирующий при δ = 3
9. J. Frahm, K. D. Merboldt, and W. Hänicke, J. Magn.
м.д., под влиянием процесса нейротрансмиссии.
Resonance 72 (3), 502 (1987).
10. P. K. Bhattacharyya, M. D. Phillips, L. A. Stone, and
M. J. Lowe, Magn. Reson. Imaging 29 (3), 374 (2011).
ВЫВОДЫ
11. V. Govindaraju, K. Young, and A. A. Maudsley, NMR
Из результатов настоящей работы можно сде-
Biomed. 13 (3), 129 (2000).
лать вывод, что макромолекулы со структурным
12. M. Mescher, H. Merkle, J. D. Kirsch, et al., NMR
фрагментом, резонирующим на 3 м.д., не вовле-
Biomed. 11 (6), 266 (1998).
чены в процесс нейротрансмиссии при обработке
зрительного стимула. Снижающийся в условиях
13. S. J. Kish, T. L. Perry, and S. Hansen, J. Neurochem.
нагрузки уровень GABA при постоянстве уровня
32 (6), 1629 (1979).
Glx свидетельствует о роли тормозного нейроме-
14. R. A. Edden, N. A. Puts, and P. B. Barker, Magn.
диатора в обеспечении баланса процессов воз-
Reson. Med.
68
(3),
657
(2012).
DOI:
буждения-торможения в зрительной коре при
10.1002/mrm.24391.
длительной видеостимуляции.
15. D. C. Shungu, X. Mao, R. Gonzales, et al., NMR
Biomed. 29 (7), 932 (2016).
16. M. Považan, G. Hangel, B. Strasser, et al., Neuroimage
ФИНАНСИРОВАНИЕ РАБОТЫ
121, 126 (2015).
Работа выполнена при финансовой поддержке
17.
L. Michels, E. Martin, P. Klaver, et al., PloS One 7 (4),
Российского научного фонда (грант № 18-13-
e31933 (2012).
00030).
18.
R. J. Maddock, G. A. Casazza, D. H. Fernandez, and
M. I. Maddock, J. Neurosci. 36 (8), 2449 (2016).
КОНФЛИКТ ИНТЕРЕСОВ
19.
R. Mekle, S. Kühn, H. Pfeiffer, et al., NMR Biomed.
Авторы заявляют об отсутствии конфликта
30 (2), e3672 (2017).
интересов.
20.
C. Chen, H. P. Sigurdsson, S. Pépés, et al., NeuroIm-
age 156, 207 (2017).
21.
P. Bednařík, I. Tkáč, F. Giove, et al., J. Cereb. Blood
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
Flow Metab. 35 (4), 601 (2015).
Все процедуры были одобрены этическим ко-
22.
K. Kurcyus, E. Annac, N. M. Hanning, et al., J. Neu-
митетом Научно-исследовательского института
rosci. 38 (46), 9967 (2018).
неотложной детской хирургии и травматологии
23.
П. Е. Меньщиков, Н. А. Семенова, Т. А. Ахадов и
Департамента здравоохранения города Москвы, а
др., Биофизика 62 (6), 1221 (2017).
также соответствовали Хельсинкской декларации
24.
А. В. Манжурцев, Н. А. Семенова, М. В. Ублин-
о проведении исследований с участием людей.
ский и др., Изв. РАН. Сер. хим., № 6, 1630 (2016).
Все участники исследования дали информи-
25.
R. A. De Graaf, In vivo NMR Spectroscopy: Principles
рованное согласие на участие в экспериментах.
and Techniques (John Wiley & Sons, 2019).
26.
СПИСОК ЛИТЕРАТУРЫ
27.
S. Cavassila, S. Deval, C. Huegen, et al., NMR
1. D. A. Lewis, D. W. Volk, and T. Hashimoto, Psycho-
Biomed. 14 (4), 278 (2001).
pharmacology 174 (1), 143 (2004).
28.
2. G. Sanacora, G. F. Mason, D. L. Rothman, et al.,
Ch10.wilcoxon.pdf.
Arch. Gen. Psychiatry 56 (11), 1043 (1999).
29.
S. Mangia, I. Tkác, R. Gruetter, et al., J. Cereb. Blood
3. A. W. Goddard, G. F. Mason, A. Almai, et al., Arch.
Flow Metab. 27 (5), 1055 (2007).
Gen. Psychiatry 58 (6), 556 (2001).
30.
B. Schaller, R. Mekle, L. Xin, et al., J. Neurosci. Res.
4. O. A. Petroff and D. L. Rothman, Mol. Neurobiol. 16
91 (8), 1076 (2013).
(1), 97 (1998).
31.
A. Schousboe and U. Sonnewald, Glutamate/-GABA-
5. L. K. Bak, A. Schousboe, and H. S. Waagepetersen, J.
glutamine Cycle (Springer International Pu, 2016).
Neurochem. 98 (3), 641 (2006).
32.
Y. Lin, M. C. Stephenson, L. Xin, et al., J. Cereb.
Blood Flow Metab. 32 (8), 1484 (2012).
6. A. Floyer-Lea, M. Wylezinska, T. Kincses, and P. M.
Matthews, J. Neurochem. 95 (3), 1639 (2006).
33.
R. C. G. Landim, R. A. Edden, B. Foerster, et al.,
Magn. Reson. Imaging 34 (3), 239 (2016).
7. R. A. E. Edden, S. D. Muthukumaraswamy,
T. C. A. Freeman, and K. D. Singh, J. Neurosci. 29
34.
K. L. Behar and T. Ogino, Magn. Reson. Medicine 30
(50), 15721 (2009).
(1), 38 (1993).
БИОФИЗИКА том 65
№ 1
2020
68
ЯКОВЛЕВ и др.
The Effect of Visual Stimulation on GABA and Macromolecules Levels
in Human Brain in vivo
A. Yakovlev*, A. Manzhurtsev**, ***, P. Menshchikov*, M. Ublinskiy**, ***, О. Bozhko***,
T. Akhadov***, and N. Semenova*, **, ***
*Semenov Institute of Chemical Physics, Russian Academy of Sciences, ul. Kosygina 4, Moscow, 119991 Russia
**Emanuel Institute of Biochemical Physics, Russian Academy of Sciences, ul. Kosygina 4, Moscow, 119334 Russia
***Clinical and Research Institute of Emergency Paediatric Surgery and Traumatology,
ul. Bolshaya Polyanka 22, Moscow, 119180 Russia
Gamma-aminobutyric acid (GABA) is the main inhibitory neurotransmitter in the brain and plays an im-
portant role in various types of synaptic plasticity and pathology. In present work, proton magnetic resonance
spectroscopy (1H-MRS) and two modified MEGA-PRESS pulse sequences such as -GABA MEGA-PRESS
and +GABA MEGA-PRESS were used to investigate the effect of visual stimulation on the GABA level in hu-
man visual cortex in vivo. With -GABA MEGA-PRESS it was possible to acquire 1H-NMR-signal of methy-
lene protons of GABA with the chemical shift δ = 3.01 ppm without signals from macromolecules. When
+GABA MEGA-PRESS was implemented, the GABA signal was a superposition of resonances of methylene
protons of GABA and macromolecules. The effect of constant visual stimulation on the level of N-acetylas-
partate and total level of glutamate and glutamine was also estimated. Constant visual stimulation has no ef-
fect on the levels of N-acetylaspartate and glutamine in the visual cortex. -GABA signal intensity decreased
with a statistically significant decrease in the level of GABA signal intensity leading to inactivation of GABA
synthesis. No statistically significant changes in the intensity of +GABA resonance were found, probably due
to the effect of visual stimulation on macromolecules. In order to test this hypothesis a signal from macro-
molecules was acquired using specifically-designed inversion-recovery pulse sequence. It was shown that the
intensity of this signal is unaffected by visual stimulation and the lack of changes in the intensity of +GABA
signal during stimulation has to be considered as the masking effect of macromolecular signal..
Keywords: 1H-MRS, GABA, MEGA-PRESS, macromolecules, visual stimulation
БИОФИЗИКА том 65
№ 1
2020