БИОФИЗИКА, 2020, том 65, № 1, с. 104-111
БИОФИЗИКА КЛЕТКИ
УДК 577.352, 576.32
ИНГИБИРОВАНИЕ ГАМКЕРГИЧЕСКОЙ ПЕРЕДАЧИ КАК МОДЕЛЬ
ГИПЕРАКТИВАЦИИ КЛЕТОК ПУРКИНЬЕ МОЗЖЕЧКА КРЫС
© 2020 г. Т.В. Карелина, Ю.Д. Степаненко, Д.А. Сибаров, П.А. Абушик, С.М. Антонов
Институт эволюционной физиологии и биохимии им. И.М. Сеченова РАН, 194223, Санкт-Петербург, пр. Тореза, 44
E-mail: polinaabushik@gmail.com
Поступила в редакцию 06.08.2019 г.
После доработки 14.10.2019 г.
Принята к публикации 18.11.2019 г.
Патологические изменения в мозжечке зачастую сопряжены с дисфункцией клеток Пуркинье, ко-
торая проявляется в чрезмерной спайковой активности. Вопреки классической модели нейродеге-
нерации, когда гиперактивацию нейронов вызывают долговременным действием высоких концен-
траций возбуждающих агонистов (глутамата или N-метил-D-аспартата), в наших экспериментах
увеличение частоты спайковой активности клеток Пуркинье было вызвано за счет ингибирования
тормозной передачи антагонистом ГАМКА-рецепторов габазином (гидробромидом 6-имино-3-(4-
метоксифенил)-1(6H)-пиразинобутановой кислоты). У взрослых и молодых животных габазин вы-
зывал достоверное увеличение частоты разрядов клеток Пуркинье через 25 и 45 мин действия соот-
ветственно. Таким образом, на взрослых животных данная модель работала более эффективно, чем
на молодых, что определяется онтогенетическими особенностями формирования коры мозжечка.
Более того, применение NS 309 (6,7-дихлор-1H-индол-2,3-дикетон-3-оксима), модулятора Са2+-
активируемых К+-каналов малой проводимости, в данной модели позволяло компенсировать вы-
званное антагонистом габазином увеличение частоты простых спайков в разряде клеток Пуркинье
до уровня контрольных значений у молодых и взрослых крыс.
Ключевые слова: клетки Пуркинье, мозжечок, SK-каналы, ГАМК, спайк.
DOI: 10.31857/S0006302920010135
ют активность КП [6]. Ранее при создании моде-
Мозжечок млекопитающих играет ключевую
ли гиперактивации КП, сопровождающей спин-
роль в моторной активности, осуществляя мони-
номозговую атаксию, было показано, что
торинг всех двигательных актов и снижая ошибку
активация рецепторов глутамата в срезах мозжеч-
между задуманным и совершенным действием
ка и in vivo, вопреки ожиданиям, приводила к ин-
[1]. На клетках Пуркинье (КП), которые являют-
гибированию постсинаптических токов КП и
ся ключевым морфофункциональным элементом
снижала частоту простых спайков. Важно, что ап-
коры мозжечка, происходит конвергенция всей
пликация антагониста ГАМКА-рецепторов
сенсорной информации, поступающей в мозже-
чок. Аксоны этих клеток формируют единствен-
SR 95 531 (гидробромида 6-имино-3-(4-метокси-
ный эфферентный путь из коры мозжечка к его
фенил)-1(6H)-пиразинобутановой кислоты) сни-
ядрам и вестибулярным центрам продолговатого
мала ингибирующее действие агониста рецептора
мозга [2, 3]. Спинномозговая атаксия является
глутамата N-метил-D-аспартата, тем самым уве-
нейродегенеративным заболеванием, которое
личивая спонтанную активность КП [7]. Более
проявляется в нарушении координации и точно-
того, на срезах мозга в гиппокампе [8], тканевых
сти движений и сопровождается изменением пат-
эксплантантах неокортекса [9], в первичной мо-
терна активности КП, а затем их дисфункцией и
торной и соматосенсорной коре in vivo [10] и ней-
гибелью [4, 5]. Известно, что за счет выброса
ронах коры в первичной культуре ткани [11] было
ГАМК (γ-аминомасляной кислоты) интернейро-
показано, что блокирование тормозной
ны молекулярного слоя коры мозжечка регулиру-
ГАМКергической передачи бикукуллином при-
водит к возникновению эпилептиформных волн
деполяризации нейронов, увеличению частоты
Сокращения: КП - клетки Пуркинье, ГАМК - γ-амино-
масляная кислота, SR 95 531 - 6-имино-3-(4-метоксифе- спонтанных возбуждающих постсинаптических
нил)-1(6H)-пиразинобутановой кислоты гидробромид
токов и генерации эпилептоподобной электриче-
(габазин), SK-каналы - Са2+-активируемые К+-каналы
малой проводимости, NS 309 - 6,7-дихлор-1H-индол-2,3-
ской активности. Следует отметить, что отмывка
дикетон-3-оксим.
бикукуллина не подавляла генерацию подобных
104
ИНГИБИРОВАНИЕ ГАМКЕРГИЧЕСКОЙ ПЕРЕДАЧИ
105
аномальных форм электрической активности
МАТЕРИАЛЫ И МЕТОДЫ
нейронной сети [11].
Исследование проводили c помощью метода
В развитии центральной нервной системы
внеклеточной регистрации активности КП моз-
ГАМК регулирует морфогенез, а именно мигра-
жечка на самцах крыс линии Вистар. При прове-
цию клеток и их синаптогенез, т. е. рост аксонов
дении экспериментов животные были разделены
к мишеням иннервации, образование синапсов, а
на две группы: молодые (всего 22 крысы возрас-
также и клеточную гибель. Показано, что только
том от 14 до 15 суток или P14-15 соответственно)
к 30-м суткам постнатального развития (P30)
и взрослые (всего 26 крыс возрастом от 3 до 6 ме-
формируется полноценная тормозная ГАМКер-
сяцев или P90-150 соответственно). Для нарко-
гическая синаптическая передача, присущая зре-
тизации животных использовали уретан, кото-
лой центральной нервной системе [12]. При ис-
рый вводили внутрибрюшинно из расчета
1200 мг/кг массы животного. При необходимости
следовании функциональной активности КП не-
в ходе эксперимента дозу увеличивали до
обходимо также учитывать и возрастные
1600 мг/кг. Регистрацию и идентификацию КП
особенности дифференцировки клеточных эле-
осуществляли по ранее описанной методике [20].
ментов и их связей в коре мозжечка. Например,
У наркотизированного животного удаляли
звездчатые клетки начинают образовывать си-
скальп, снимали мышечный слой и сверлили от-
напсы с дендритами КП в Р14, что совпадает с на-
верстие диаметром 2 мм в затылочной кости над
чалом расширения дендритного дерева КП [13].
червем мозжечка. Затем крысу закрепляли в сте-
Другие авторы указывают на то, что процесс си-
реотаксической установке. Для регистрации вне-
наптогенеза протекает в коре мозжечка вплоть до
клеточной активности КП использовали стек-
Р90 [14], а на Р21 в белом веществе мозжечка еще
лянные микроэлектроды из боросиликатного
присутствуют пролиферирующие КП [15]. Кроме
стекла (внешний диаметр 1.5 мм, внутренний -
этого, на третьей неделе постнатального развития
1.1 мм, Sutter Instrument, США), заполненные
продолжается формирование электрофизиологи-
раствором 2.5 М NaCl. Погружение микроэлек-
ческих свойств КП, так как в этот период появля-
трода в ткань мозжечка осуществляли с помощью
ются смешанные Na+-Ca2+-спайки и тримодаль-
автоматического манипулятора с шагом погруже-
ный паттерн разряда КП [16]. Также онтогенети-
ния 5 мкм на глубину до 5 мм. КП идентифициро-
ческие особенности присущи и экспрессии Ca2+-
вали по характерному для них паттерну активно-
сти: наличию простых и сложных спайков, а так-
активируемых K+-каналов малой проводимости
же тормозной паузы после сложного спайка
(SK-каналы), модуляция которых приводит к
перед серией простых. Сигнал от регистрируемой
снижению частоты простых спайков КП, тем са-
клетки усиливали с помощью дифференциально-
мым оказывая терапевтический эффект при лече-
го усилителя (АС/DC Differential Amplifier, model
нии спинномозговой атаксии [4, 5, 17, 18]. У ста-
3000, A-M Systems, Inc, США) и оцифровывали с
рых животных снижение частоты простых спай-
частотой дискретизации 10000 изм/с с помощью
ков КП за счет введения положительных
аналого-цифрового преобразовтеля L-791 (ЗАО
модуляторов SK-каналов происходит быстрее,
«Л-КАРД», Россия) в оригинальной программе
чем у взрослых животных, что определяется воз-
Bioactivity Recorder v. 5.3, разработанной Д.А. Си-
растными особенностями динамики внутрикле-
баровым
точного Са2+ и снижением экспрессии потенци-
для последующего анализа частоты простых
спайков в программе Clampfit 10.2 (Molecular De-
ал-зависимых Ca2+-каналов в молекулярном слое
vices Corp, США). Количество зарегистрирован-
мозжечка [19].
ных КП соответствует количеству использован-
Таким образом, ввиду несостоятельности ги-
ных животных, т. е. на одном животном была за-
перактивирующего действия агонистов рецепто-
регистрирована одна КП. Подачу действующих
ров глутамата на КП [7] и наличием отногенети-
веществ осуществляли согласно стандартной ме-
тодике [21] путем их аппликации микропипеткой
ческих особенностей развития коры мозжечка,
цель данной работы заключалась в создании мо-
на обнаженную (диаметром 2 мм) поверхность
мозжечка в области введения микроэлектрода.
дели гиперактивации КП и сравнении ее эффек-
тивности у разных возрастных групп животных. В
Сначала была проведена контрольная серия экс-
работе был проведен анализ изменения частоты
периментов, в которой осуществляли аппликацию
простых спайков КП при ингибировании тормоз-
физиологического раствора (0.9% NaCl) на обна-
ной передачи через ГАМКА-рецепторы у моло-
женную поверхность мозжечка в зоне введения
дых (P14-15) и взрослых (P90-150) крыс в модели
микроэлектрода. Затем в следующих двух сериях
in vivo. В данной модели активации КП проведен
экспериментов для ослабления тормозного влия-
анализ изменения частот образования спайков
ния интернейронов на КП и, как следствие, созда-
при модуляции SK-каналов.
ния условия для повышенного возбуждения КП
БИОФИЗИКА том 65
№ 1
2020
106
КАРЕЛИНА и др.
был использован избирательный антагонист
значения. Изменение среднего значения относи-
ГАМКА-рецепторов SR 95 531 (габазин, Tocris,
тельной частоты простых спайков по сравнению с
США). Для снятия гипервозбуждения КП исполь-
исходной частотой в отдельные периоды реги-
зовали позитивный модулятор SK- и IK-каналов -
страции в возрастной группе Р14 составило 7-
NS 309
(6,7-дихлор-1H-индол-2,3-дикетон-3-ок-
18%, а у взрослых животных - 7-13%. Таким об-
сим) (Tocris, США). При аппликации веществ и
разом, было продемонстрировано, что сама про-
физиологического раствора осуществляли визуаль-
цедура аппликации не влияла на картину разряда
ный контроль уровня жидкости в краниальном ок-
КП.
не и для поддержания его на одном уровне повтор-
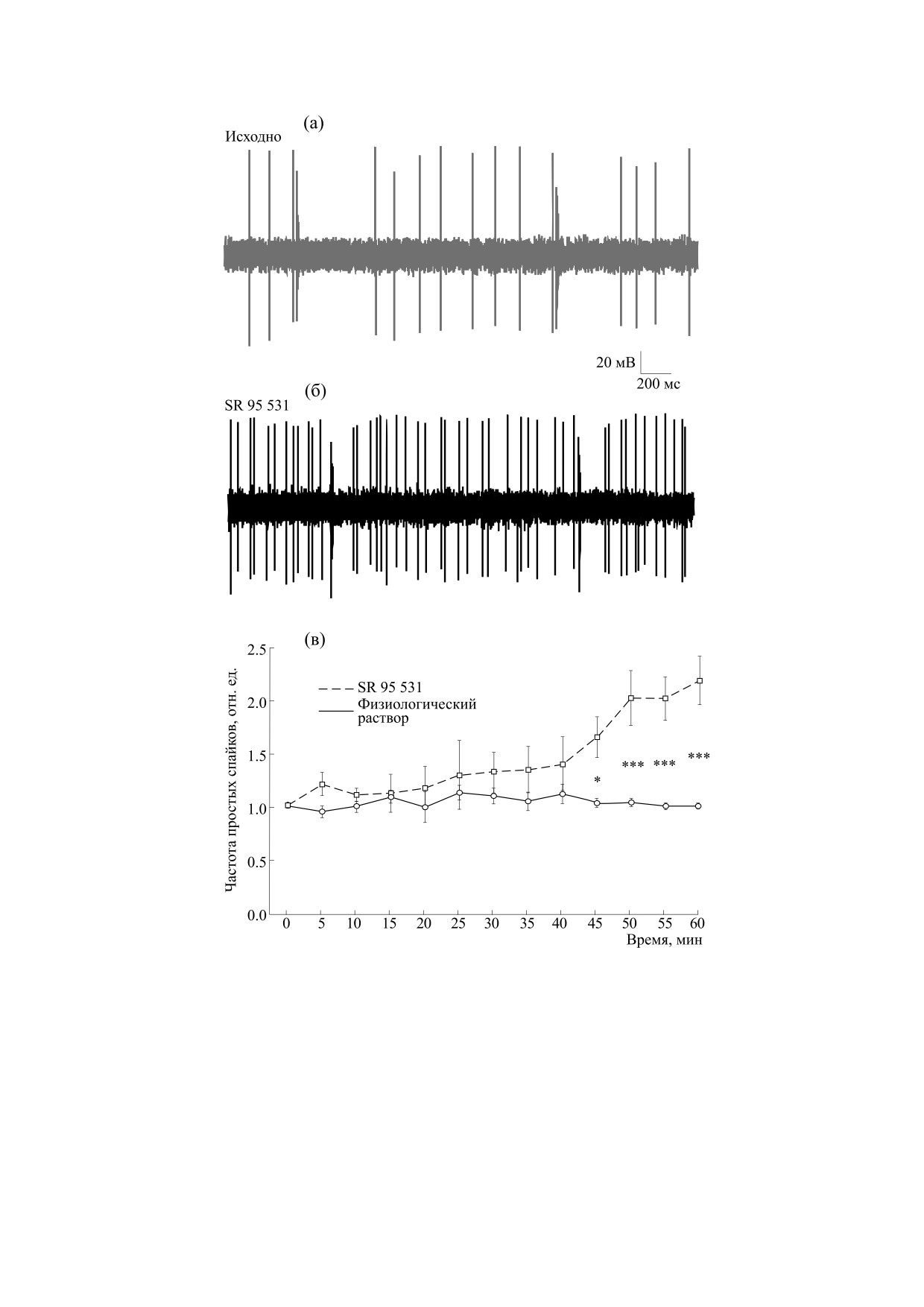
У крысят SR 95 531 вызывал постепенное уве-
но апплицировали раствор микропипеткой. За час
личение частоты простых спайков в разряде КП
регистрации общий объем апплицируемого веще-
(верхняя кривая на рис. 1в). Через 45 мин после
ства составлял 500 мкл и соответственно за 30 мин
аппликации были обнаружены достоверные от-
регистрации - 250 мкл. Действующие вещества ис-
личия от контроля в соответствующей временной
пользовали в концентрации
200 мкМ как для
точке (p < 0.05, n = 6, ANOVA, поправка Бонфер-
SR 95 531, так и для NS 309. Растворы действующих
рони), которые сохранялись до конца периода ре-
веществ готовили на 0.9%-м NaCl. Внеклеточную
гистрации. Наибольшее значение частота про-
регистрацию активности идентифицированной
стых спайков достигала через 60 мин от начала
КП осуществляли 30-секундными интервалами в
аппликации и была более чем в два раза больше
течение 15 мин до аппликации вещества (исходная
контрольного значения в это же время. У взрос-
частота) и 60 мин после аппликации, через каждые
лых животных, в отличие от молодых крыс, при
5 мин.
аппликации SR 95 531 достоверное увеличение
частоты простых спайков в разряде КП (рис. 2а,б)
В первой серии экспериментов после реги-
достигалось через 25 мин после начала апплика-
страции исходной активности КП (рис. 1а) было
ции (p < 0.05, n = 7, ANOVA, поправка Бонферро-
исследовано влияние SR 95 531 на паттерн разря-
ни), т. е. раньше, чем у молодых крыс. При этом
да КП мозжечка (рис. 1б). Во второй серии экспе-
максимальное увеличение происходило уже к
риментов помимо регистрации исходной актив-
30 мин регистрации, но в среднем лишь на 30%
ности КП и после 60 мин действия антагониста
(рис. 2в).
ГАМКА-рецепторов SR 95 531 осуществляли ап-
пликацию NS 309 и продолжали регистрацию ак-
Как видно из рис. 1 и 2, характер изменений
частоты простых спайков в разряде КП у живот-
тивности КП в течение еще 40-60 мин.
ных разных возрастных групп отличается друг от
При сравнении данных из контрольной серии
друга. Если у двухнедельных крысят наблюдался
экспериментов с экспериментальными для каж-
неуклонный рост частоты простых спайков на
дой клетки определяли среднее значение частоты
протяжении всего периода аппликации SR 95 931,
простых спайков в течение 30 с по всем указан-
то у взрослых животных, начиная с 25 мин от на-
ным временным отметкам. После этого высчиты-
чала аппликации, значение частоты простых
вали среднее значение по всем зарегистрирован-
спайков практически не менялось до конца пери-
ным клеткам и стандартную ошибку среднего для
ода аппликации. Таким образом, можно сделать
всех временных отметок. Затем высчитывали от-
вывод о том, что данный подход - использование
носительные частоты, приняв за единицу исход-
селективного антагониста ГАМКА-рецепторов
ную частоту спайков до аппликации. В дальней-
SR 95 931 как блокатора тормозной активности в
шем для выявления достоверности отличий меж-
нейронной сети [7] - применим для получения
ду контрольной серией экспериментов и сериями
модели гипервозбуждения КП мозжечка.
с аппликацией антагониста ГАМКА-рецепторов
Влияние положительного модулятора SK/IK-
SR 95 531 и активаторов SK-каналов NS 309 для
каналов на частоту простых спайков клеток Пурки-
каждой временной отметки использовали двух-
нье, вызванных действием антагониста ГАМКА-
факторный дисперсионный анализ (ANOVA) с
применением поправки Бонферрони.
рецепторов SR 95 531. Во второй серии экспери-
ментов мы проверяли гипотезу о том, что гипер-
активацию КП мозжечка, полученную при ослаб-
РЕЗУЛЬТАТЫ
лении тормозного влияния, оказываемого на нее
интернейронами молекулярного слоя, можно ни-
Влияние антагониста ГАМКА-рецепторов
велировать за счет активации Са2+-активируемых
SR 95 531 на частоту простых спайков клеток Пур-
калиевых каналов. Для этой цели мы применяли
кинье у молодых и взрослых крыс. В контрольной
серии экспериментов как у молодых животных,
агонист SK/IK-подтипов Са2+-активируемых ка-
так и у взрослых крыс при аппликации физиоло-
лиевых каналов NS 309. Как в группе молодых
гического раствора в течение 60 мин не было вы-
крыс (Р14-15), так и в группе взрослых животных
явлено достоверного изменения частоты простых
(Р90-150) нам удалось получить снижение часто-
спайков в разряде КП относительно исходного
ты простых спайков в паттерне разряда КП. Од-
БИОФИЗИКА том 65
№ 1
2020
ИНГИБИРОВАНИЕ ГАМКЕРГИЧЕСКОЙ ПЕРЕДАЧИ
107
Рис. 1. Активность клеток Пуркинье мозжечка молодых крыс (14-е-15-е сутки постнатального развития, P14-15) в
контроле и под влиянием антагониста ГАМКА-рецепторов SR 95 531. Представлены фрагменты записи активности
КП исходно, до аппликации SR 95 531 (200 мкМ) (а), и во время аппликации антагониста (б). (в) - Изменение средней
частоты простых спайков КП мозжечка в течение 1 ч при аппликации физиологического раствора (серая линия) и
антагониста SR 95 531 (черная пунктирная линия). Звездочками показано статистически достоверное отличие от
контрольных значений частоты в соответствующие моменты времени (ANOVA, поправка Бонферрони; * - p < 0.05,
*** - p < 0.001).
БИОФИЗИКА том 65
№ 1
2020
108
КАРЕЛИНА и др.
Рис. 2. Активность клеток Пуркинье мозжечка взрослых крыс (90-е-150-е сутки постнатального развития, P90-150)
исходно и под влиянием антагониста ГАМКА-рецепторов SR 95 531 (200 мкМ). Представлены фрагменты записи
активности КП исходно, до аппликации SR 95 531 (а), и во время аппликации антагониста (б). (в) - Изменение
средней частоты простых спайков КП мозжечка в течение 1 ч при аппликации физиологического раствора (серая
линия) и антагониста SR 95 531 (черная пунктирная линия). Звездочками показано статистически достоверное
отличие от контрольных значений частоты в соответствующие моменты времени (ANOVA, пост-тест Бонферрони; * -
p < 0.05, ** - p < 0.01, *** - p < 0.001).
нако было зафиксировано возрастное отличие
крыс достигала 6.0 ± 0.9 имп/с, а при действии
(рис. 3). В контрольных условиях к 60-й минуте
антагониста ГАМКА-рецепторов SR 95 531 в этой
измерений частота простых спайков у молодых
же временной точке наблюдалось достоверное
БИОФИЗИКА том 65
№ 1
2020
ИНГИБИРОВАНИЕ ГАМКЕРГИЧЕСКОЙ ПЕРЕДАЧИ
109
Рис. 3. Частота простых спайков (ПС) клеток Пуркинье исходно (0 мин), после 60 мин действия антагониста ГАМКА-
рецепторов SR 95 531 (200 мкМ) и последующей аппликации положительного модулятора SK-каналов NS 309 (200 мкМ)
в течение 15-20 мин и 25-30 мин. Представлена средняя частота простых спайков КП мозжечка: (а) - молодых крыс
(14-е-15-е сутки постнатального развития, P14-15); (б) - взрослых крыс (90-е-150-е сутки постнатального развития,
P90-150). Звездочками и линиями показано статистически достоверное отличие от исходных значений частоты или
частоты после воздействия (ANOVA, пост-тест Бонферрони; * - p < 0.05, ** - p < 0.01, *** - p < 0.001).
увеличение их частоты по сравнению с контролем
вой активности. Вопреки классической нейроде-
до 12.1 ± 2.3 имп/с (p < 0,001, n = 9, ANOVA, пост-
генеративной модели, когда гиперактивацию
тест Бонферрони, рис. 3а). При последующей ап-
нейронов вызывают долговременным действием
пликации агониста SK/IK-каналов NS 309 часто-
высоких концентраций возбуждающих агонистов
та простых спайков достоверно снижалась через
[23, 24], в наших экспериментах гиперактивация
15-20 мин и достигала 5.6 ± 0.9 имп/с, что досто-
КП была вызвана за счет ингибирования тормоз-
верно не отличалось от контрольных значений
ной передачи, опосредованной ГАМКА-рецепто-
(p > 0.5, n = 9, ANOVA, поправка Бонферрони). У
рами, что согласуется с данными о применении
взрослых (P90-150) крыс, в отличие от молодых
антагонистов и блокаторов ГАМКА-рецепторов
(P14-15) животных, уровень частоты простых
как инициаторов эпилептиформных волн депо-
спайков в контрольных условиях был существен-
ляризации нейронов и временному увеличению
но выше и достигал в среднем 29.1 ± 3.5 имп/с
частоты спонтанных возбуждающих постсинап-
(n = 10), тем не менее SR 95 531 вызывал досто-
тических токов [11]. Активность КП напрямую
верное по сравнению с контролем увеличение ча-
зависит от зрелости остальных клеточных эле-
стоты простых спайков до
41.1
± 2.5 имп/с
ментов коры мозжечка и формирования полно-
(р < 0,001, n = 10, ANOVA, пост-тест Бонферро-
ценной ГАМКергической передачи [12, 16]. У
ни). Важно, что, в отличие от молодых крыс, ап-
крысят процесс морфо- и гистогенеза коры моз-
пликация NS 309 приводила к достоверному сни-
жечка продолжается весь первый месяц постна-
жению частоты простых спайков до уровня кон-
тальной жизни [13, 25]. При этом происходят
троля лишь через 25-30 мин (p > 0.05, n = 10
функциональные изменения КП, а именно уве-
ANOVA, поправка Бонферрони). Таким образом,
личение активности за счет усиления ГАМКерги-
можно сделать вывод, что как у молодых жи-
ческого влияния тормозных интернейронов мо-
вотных, так и у взрослых активация SK/IK-под-
лекулярного слоя на КП в результате созревания
типов Ca2+-активируемых K+-каналов способна
их связей с КП [26]. В наших экспериментах у мо-
снять гиперактивацию КП мозжечка в условиях
лодых и взрослых крыс ингибирование ГАМКА-
снижения тормозного влияния, оказываемого на
КП интернейронами молекулярного слоя. При
рецепторов SR 95 531 достигало насыщающих
этом исходное значение частоты простых спай-
концентраций вблизи КП постепенно по мере
ков у крысят восстанавливалось раньше, чем у
диффузии, поэтому скорость наступления эф-
взрослых животных.
фекта могла зависеть от возрастных особенностей
созревания коры мозжечка. Так, у молодых крыс
увеличение спайковой активности наступало в
два раза медленнее, чем у взрослых крыс (рис. 1в
ОБСУЖДЕНИЕ
и 2в). Это, по-видимому, объясняется различия-
Нейродегенеративные изменения в мозжечке
ми в степени зрелости морфологических и функ-
зачастую сопряжены с дисфункцией КП [22], ко-
циональных свойств КП и незаконченным про-
торая может проявляться в чрезмерной спайко-
цессом формирования ГАМКергических синап-
БИОФИЗИКА том 65
№ 1
2020
110
КАРЕЛИНА и др.
сов в этом возрасте. Следует отметить, что частота
пользования животных при выполнении работы
простых спайков в контроле (исходно) у молодых
были соблюдены.
крыс ниже, чем у взрослых. Это соответствует
нормальному постнатальному физиологическо-
СПИСОК ЛИТЕРАТУРЫ
му развитию, когда в онтогенезе происходит уве-
личение спайковой активности [19], и может объ-
1.
M. Ito, Brain Res. 886 (1-2), 237 (2000).
ясняться уменьшением длительности рефрактер-
2.
J. M. Bower, Front. Cell. Neurosci. 4, pii:27 (2010).
ного периода, следующего за возникновением
3.
J. C. Eccles, M. Ito, and J. Szentagothai, The cerebel-
спайка [27]. Таким образом, примененная нами
lum as a neuronal machine (Springer-Verlag, Berlin,
модель гиперактивации КП через ингибирование
1967).
ГАМКергической передачи in vivo может эффек-
4.
V. G. Shakkottai, M. do Carmo Costa, J. M. Dell'Or-
тивно применяться при исследовании нейродеге-
co, et al., J. Neurosci. 31 (36), 13002 (2011).
неративных состояний в мозжечке крыс. Более
5.
J. T. Walter, K. Alvina, M. D. Womack, et al., Nat.
того, использование модулятора SK-каналов
Neurosci. 9 (3), 389 (2006).
NS 309 в данной модели позволило предотвра-
6.
W. Pan, W.-Y. Wu, Y.-H. Bing, et al., Pharmacology
тить развитие гиперактивации КП, тем самым
103, 82 (2019).
продемонстрировав терапевтический эффект -
7.
H. Liu, S.-N. Zhao, G.-Y. Zhao, et al., Brain Res.
снижение частоты простых спайков КП. Это со-
1560, 1 (2014).
гласуется с данными о восстановлении нарушен-
8.
H. S. Swatzwelder, W. W. Anderson., and W. A. Wilson,
ного паттерна активности КП и снижении симп-
Epilepsy Res. 2 (4), 239 (1988).
томов спиномозговой атаксии у мутантных жи-
9.
M. J. Gutnick, B. Wolfson, and F. Baldino, Exp. Brain
вотных [4, 5, 17, 18]. В работе, выполненной in vivo
Res. 76 (1), 131 (1989).
на взрослых и старых крысах, показано, что ча-
10.
M. Sáeza, M. Ketzefb, J. Alegre-Cortésa, et al., Neuro-
стота простых спайков КП при аппликации
science 381, 115 (2018).
NS 309 снижается быстрее у старых крыс, что
11.
Д. А. Сибаров, П. А. Абушик, А. Е. Большаков и
др., Биол. мембраны 31 (1), 33 (2014).
опосредовано онтогенетическим изменением
экспрессии потенциал-зависимых кальциевых
12.
C. Takayama and Y. Inoue, Anat. Sci. Int. 79, 124
(2004).
каналов P/Q-типа и исчезновением потенциал-
зависимых кальциевых каналов L-типа в молеку-
13.
J. Altman, J. Comp. Neurol. 145, 399 (1972b)
лярном слое мозжечка [19, 28].
14.
J. Takács and J. Hámori, J. Neurosci. Res. 38 (5), 515,
(1994).
Таким образом, результаты, представленные в
15.
H. Yamanaka, Y. Yanagawa, and K. Obata, Neurosci.
данной работе, показывают различие в скорости
Res. 50, 1 (2004).
снижения активности КП: у молодых крыс сни-
16.
B. E. McKay and R. W. Turner, J. Physiol. 567 (3), 829
жение частоты простых спайков происходило
(2005).
быстрее, чем у взрослых, что может быть опосре-
17.
M. Gymnopoulos, L. A. Cingolani, P. Pedarzani, et al.,
довано различиями в экспрессии потенциал-за-
J. Comp. Neurol. 522 (5), 1072 (2014).
висимых Ca2+ каналов и Ca2+-проницаемых ре-
18.
M. D. Womack and K. Khodakhah, J. Neurosci. 23 (7),
цепторов N-метил-D-аспартата [29]. Причины
2600 (2003).
возрастных отличий в эффекте модуляции SK-
19.
Т. В. Карелина, Ю. Д. Степаненко, П. А. Абушик
каналов требуют дальнейшего изучения, по-
и др., Acta Naturae 8, 4(31), 53 (2016).
скольку могут быть связаны с дефицитом функ-
20.
П. А. Егорова, Т. В. Карелина, О. Л. Власова и др.,
ций КП, связанных с особенностями онтогенети-
Журн. эвол. биохим. физиол. 50 (2), 102 (2014).
ческого развития.
21.
Z. Gao, B. Todorov, C. F. Barrett, et al., J. Neurosci.
32 (44), 15533 (2012).
22. M. Samson, D. O. Claassen, Neurodegener Dis. 17 (4-
ФИНАНСИРОВАНИЕ РАБОТЫ
5), 155 (2017).
23. E. V. Mironova, A. A. Evstratova, and S. M. Antonov,
Работа выполнена в рамках государственного
J. Neurosci. Methods 163, 1 (2007).
задания № АААА-А18-118012290427-7.
24. A. Lau, M. Tymianski, Pflugers Arch. 460 (2), 525
(2010).
КОНФЛИКТ ИНТЕРЕСОВ
25. M. B. Pisu, E. Roda, D. Avella, et al., Neuroscience
129, 655 (2004).
Авторы заявляют об отсутствии конфликта
26. M. Hausser and B. A. Clark, Neuron 19, 665 (1997).
интересов.
27. S. Guan, S. Ma, Y. Zhu, et al., Brain Res. 1097 (1), 59
(2006).
28. Y. H. Chung, C. M. Shin, M. J. Kim, et al., Brain Res.
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
903 (1-2), 247 (2001).
Все применимые международные, националь-
29. D.A. Sibarov, J. D. Stepanenko, I.V. Silantiev, et al., J.
ные и институциональные принципы ухода и ис-
Mol. Neurosci. 64 (2), 300 (2018).
БИОФИЗИКА том 65
№ 1
2020
ИНГИБИРОВАНИЕ ГАМКЕРГИЧЕСКОЙ ПЕРЕДАЧИ
111
Inhibition of GABAergic Transmission as a Model of Purkinje Cell Hyperactivation
in the Rat Cerebellum
T.V. Karelina, J.D. Stepanenko, D.A. Sibarov, P.A. Abushik, and S.M. Antonov
Sechenov Institute of Evolutionary Physiology and Biochemistry, Russian Academy of Sciences,
prosp. Toreza 44, St. Petersburg, 194223 Russia
Pathological changes in the cerebellum are commonly associated with Purkinje cell dysfunction manifesting
with excessive spike activity. Despite a classical model of neurodegeneration according to which neuronal hy-
peractivity is induced by long-lasting effects of high concentrations of excitatory agonists (glutamate or
N-methyl-D-aspartate), in our experiments the frequency of Purkinje cell spike activity increased with sup-
pression of inhibitory transmission by gabazine (6-imino-3-(4-methoxyphenyl)-1(6H)-pyridazinebutanoic
acid hydrobromide), a GABAA receptor antagonist. Gabazine induced a significant increase in the frequency
of Purkinje cell spike output after 25 min and 45 min in adult rats and young rats, respectively. Thus, this
model was considered to be more appropriate for adult animals than for young animals; it can be explained
by ontogenetic aspects of the cerebellar cortex maturation. Moreover, the use of NS 309 (6,7-dichloro-1H-
indole-2,3-dione-3-oxime), a modulator of small conductance calcium-activated potassium channels, in
this model made it possible to compensate for an increase in the frequency of Purkinje cell simple spike out-
put induced by gabazine, that decreased Purkinje cells spiking frequency to control values typical for young
and adult animals.
Keywords: Purkinje cells, cerebellum, SK-channels, GABA, spike
БИОФИЗИКА том 65
№ 1
2020