БИОФИЗИКА, 2020, том 65, № 1, с. 112-124
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 58.035
ВЛИЯНИЕ СПЕКТРАЛЬНЫХ ОСОБЕННОСТЕЙ СВЕТОВОЙ СРЕДЫ
НА ПОГЛОЩЕНИЕ СВЕТА ЛИСТЬЯМИ САЛАТА
И ЕГО НЕТТО-ПРОДУКТИВНОСТЬ
© 2020 г. Т.Э. Кулешова*, И.Н. Черноусов**, О.Р. Удалова**, Л.М. Аникина**,
Ю.В. Хомяков**, А.В. Александров**, И.С. Середин***, С.В. Феофанов***,
С.А. Щеглов****, Н.Р. Галль*, Г.Г. Панова**
*Физико-технический институт им. А.Ф. Иоффе РАН, 194021, Санкт-Петербург, Политехническая ул., 26
**Агрофизический научно-исследовательский институт, 195220, Санкт-Петербург, Гражданский просп., 14
***ООО «О2 Световые Системы», 196084, Санкт-Петербург, ул. Новорощинская, 4а
****Санкт-Петербургский национальный исследовательский университет информационных технологий,
механики и оптики, 197101, Санкт-Петербург, Кронверкский проспект, 49
Поступила в редакцию 29.11.2019 г.
После доработки 10.05.2019 г.
Принята к публикации 21.08.2019 г.
Исследована эффективность поглощения света с различным спектральным составом листьями
салата и выбору оптимальных условий световой среды для повышения продуктивности и качества
растительной продукции, выращиваемой в условиях искусственного освещения. Салат сорта
Тайфун выращивали методом тонкослойной панопоники в автоматизированной вегетационной
установке с использованием пяти вариантов различных источников света. Разработанный авторами
неповреждающий метод измерения спектров поглощения листьев in vivo позволил сделать
заключение о влиянии особенностей спектрального состава светового потока на оптические
характеристики, отражающие физиологическое состояние растений. С приростом биомассы листа
салата на 1 г в течение 10 суток в условиях максимальной продуктивности количество поглощенной
энергии фотонов увеличивалось на ~140 мкмоль · м-2 · с-1, значения индекса светопоглощения
ниже 70 мкмоль · м-2 · с-1 соответствовали образцам с минимальными результатами по ростовым
характеристикам. Схожесть спектральных характеристик освещения в области фотосинтетически
активной радиации ламп ДНаТ и светодиодных светильников, излучающих желтый свет, и
практически одинаковой доли фотосинтетически активных фотонов в синей, зеленой и красной
областях спектра привели к одинаковому приращению поглощения света в процессе развития
растений. Однако значительное отличие в продуктивности салата (~50%), а также росте, развитии и
биохимическом составе свидетельствует о лучшем влиянии ламп ДНаТ на свойства растительной
культуры. Полученные данные свидетельствуют, что спектр освещения с интенсивностью
~25 мкмоль · м-2 · с-1 в диапазоне 400-500 нм, ~150 мкмоль·м-2 · с-1 в 500-600 нм, ~150 мкмоль ·
· м-2 · с-1 в 600-700 нм приводит к высокой продуктивности салата.
Ключевые слова: интенсивная светокультура, спектры освещения, светодиодные светильники, оптика
листа, светопоглощение, нетто-продуктивность.
DOI: 10.31857/S0006302920010147
При выращивании растений в условиях защи-
ния возможностей использования света разного
щенного грунта или в тепличных комплексах с
спектрального состава в организации направлен-
недостатком или отсутствием естественного света
ного биосинтеза биологически ценных соедине-
ключевыми факторами являются правильно по-
ний различного назначения остаются малоизу-
добранные спектр, интенсивность и длитель-
ченными и весьма актуальными. При этом важ-
ность искусственно созданного освещения. В ли-
ным аспектом исследований является поиск и
тературе подчеркивается, что вопросы оптимиза-
обоснование возможности снижения затрат элек-
ции спектра и интенсивности фотосинтетически
троэнергии на единицу производимой продук-
активной радиации в продукционном процессе
ции, которые при реализации интенсивной све-
растений на разных этапах их вегетации, изуче-
токультуры составляют более 40% [1-3]. Пере-
112
ВЛИЯНИЕ СПЕКТРАЛЬНЫХ ОСОБЕННОСТЕЙ СВЕТОВОЙ СРЕДЫ
113
численные аспекты создания и регуляции
ственной оценки состояния сельскохозяйствен-
световой среды становятся реализуемыми при ис-
ных полей и посевов [14]. Существуют методы
пользовании светодиодных источников света.
оценки всхожести семян фотометрическим мето-
Перспективность их применения в интенсивной
дом: например, в работе [15] вместо измерения
светокультуре отмечена многими авторами [4-6].
электропроводности раствора с выдержанными в
Появление мощных сверхъярких и модульных
нем семенами определяли его оптическую плот-
светодиодов расширило технические возможно-
ность при 360 нм. Одним из наиболее распростра-
сти для увеличения продуктивности и эффектив-
ненных методов исследования работы пигмент-
ности выращивания тепличных культур [7-9].
ного аппарата и воздействия факторов внешней
Однако весь потенциал светодиодов можно ис-
среды на параметры фотосинтеза является анализ
пользовать, лишь понимая механизм влияния
параметров флуоресценции хлорофилла [16, 17].
светового потока и спектра излучения в области
В настоящее время активно разрабатываются
фотосинтетически активной радиации как на об-
методы диагностики поглощения света листьями
щую физиологию растений, так и на их отдель-
[18, 19], создаются спектральные библиотеки [20].
ные виды и сорта. Без этих знаний практическое
Для одновременной регистрации в одном экспе-
применение светильников на основе светодиодов
рименте коэффициентов отражения, пропуска-
не приводит к желаемым результатам, и, более то-
ния и поглощения излучения были предложены
го, исследователи столкнулись с проблемой сни-
методы на основе одной интегрирующей сферы,
жения продуктивности и качества выращиваемой
двух и полусферы [13, 14]. По характеру диффуз-
растительной продукции [10].
ного отражения и светоотражающей анизотро-
В основе выбора диапазонов и полос излуче-
пии возможно изучение структуры поверхности
ния большинства представленных на рынке све-
листьев, величина светопропускания связана с
тодиодных фитосветильников лежит представле-
прозрачностью листьев, а по величине поглощен-
ние о важности двух полос - в синей (450 нм) и
ной световой энергии можно судить об особенно-
красной областях (660 нм) спектра [11], соответ-
стях усвоения излучения в зависимости от длины
ствующих максимумам поглощения основных
волны.
фотосинтетических пигментов растений. Отсут-
Для анализа спектров поглощения света расте-
ствие в светильниках других полос спектра в диа-
ниями in vivo необходимо обеспечить возмож-
пазоне длин волн фотосинтетически активной
ность экспрессных многодневных измерений
радиации - 380-720 нм или неоптимальное их
спектров отражения и пропускания света у живых
соотношение в так называемых светильниках
листьев, неоднородных по длине и ширине, на
«полного спектра» является причиной возникно-
одном растении или их популяции на протяже-
вения вышеуказанной проблемы с реализацией
нии роста и развития без причинения вреда расте-
продукционного потенциала растений. Для со-
нию, что является технически сложной задачей -
здания светодиодных светильников, обеспечива-
должны быть реализованы следующие парамет-
ющих адекватные условия освещения растений,
ры: 1) одновременная регистрация в одном экс-
необходимо дальнейшее исследование физиоло-
перименте отражения и пропускания излучения;
гических эффектов узкополосного освещения с
2) эффективный захват диффузно-отраженного
учетом энергетической и регуляторной роли раз-
или проходящего света; 3) фиксация и позицио-
личных спектральных составляющих освещения,
нирование тестируемого образца; 4) варьирова-
а также видоспецифичности реакций растений на
ние спектра и интенсивности излучения источ-
изменение спектрального состава света.
ника света в физиологически значимых для рас-
тений диапазонах.
Как известно, чувствительным показателем
физиологического состояния растений, характе-
Таким образом, цель данной работы заключа-
ризующим их потребности, особенно в отноше-
лась в разработке, апробации методики и экспе-
нии фотосинтетически активного потока фото-
риментальной установки, позволяющей измерять
нов, являются спектральные характеристики
спектры поглощения листьев in vivo, и исследова-
прошедшего через листья света, его интенсивно-
нии влияния спектральных характеристик излу-
сти и максимумов на соответствующих длинах
чения на поглощение света листовой поверхно-
волн [12, 13]. Изучение оптических характеристик
стью растения, их рост, нетто-продуктивность и
растений может стать основой для выбора источ-
качество формируемой растительной продукции.
ников освещения с наиболее эффективным спек-
тром излучения для светокультуры.
МАТЕРИАЛЫ И МЕТОДЫ
Спектральные методы исследования жизнеде-
ятельности растений широко распространены и
Исследования проводили в вегетационном
применяются. Спектрофотометрирование почв и
опыте в регулируемых условиях на базе биополи-
растительных покровов является одним из эф-
гона Агрофизического НИИ (Санкт-Петербург).
фективных методов изучения свойств и количе-
Объектом изучения служили растения салата
БИОФИЗИКА том 65
№ 1
2020
114
КУЛЕШОВА и др.
(Lactūca sātiva L.) сорта Тайфун, так как парамет-
ру воздуха в пределах 20-22°С днем и 18-20°С -
ры его листьев напрямую связаны с нетто-про-
ночью.
дуктивностью и характеризуют выход продукции.
Для получения спектров поглощения листьев
Растения выращивали методом тонкослойной
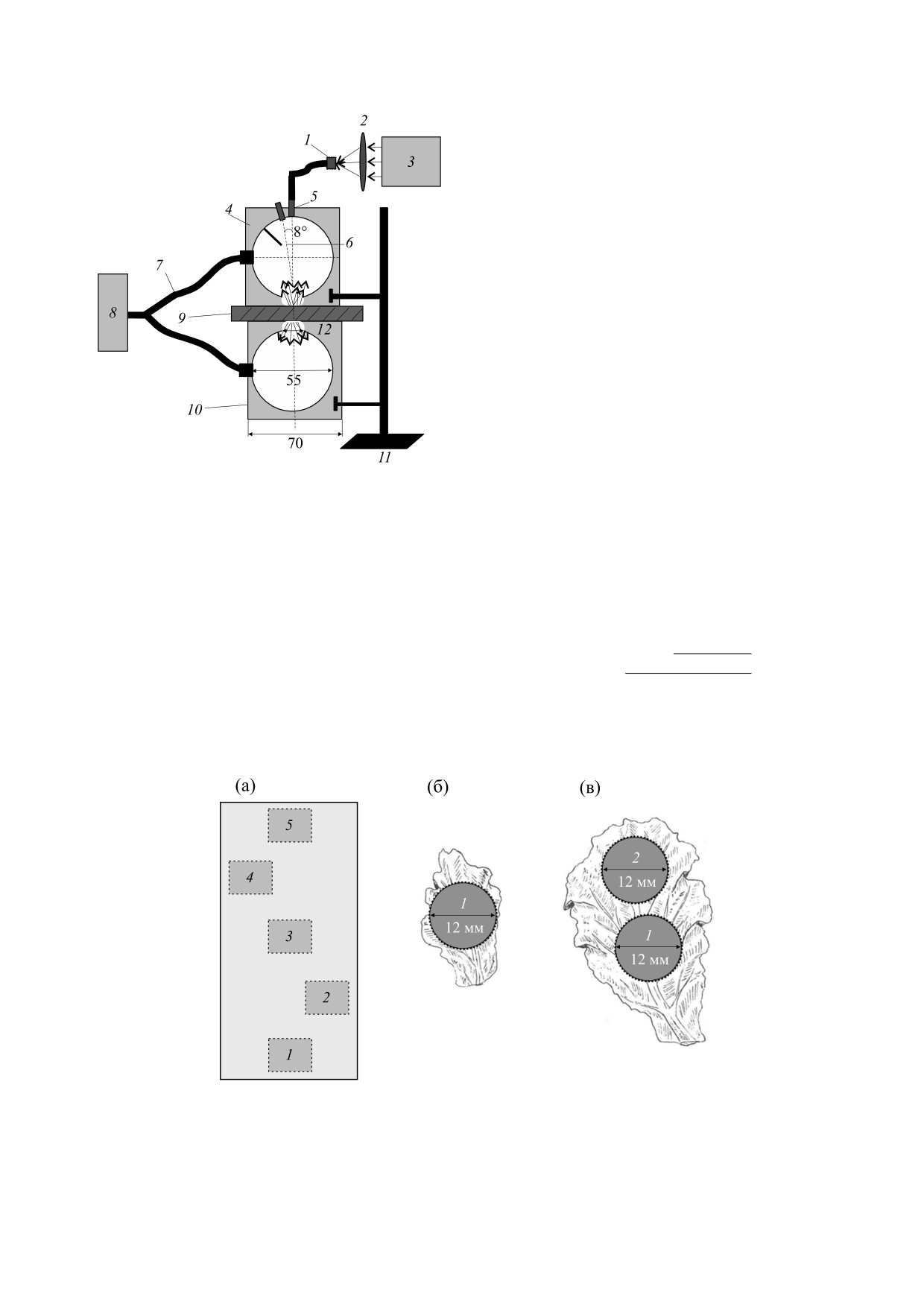
растений in vivo была разработана и собрана экс-
панопоники в автоматизированной вегетацион-
периментальная установка [25] (рис. 1). За основу
ной установке площадью 6 м2, при полностью ис-
была взята система измерения оптических
кусственном освещении [21, 22]. В качестве ис-
свойств тканей с помощью двух интегрирующих
точника минерального питания, подаваемого в
сфер [26] - исследуемый образец помещается
растильни, использовали раствор Кнопа [23].
между сферами, одна из которых (ИС1) регистри-
рует отраженный свет, а другая (ИС2) - прошед-
В исследовании оценивали влияние световой
ший.
среды различного спектрального состава на по-
глощение света листьями салата и его нетто-про-
Интегрирующие сферы с внутренним диамет-
дуктивность. Установка для выращивания была
ром 50 мм сконструированы в СПбПУ и выпол-
условна разделена светонепроницаемыми пере-
нены на 3D-принтере из АБС-пластика. Окраска
городками на пять световых зон, источниками
внутренней поверхности сфер произведена в со-
света в которых служили лампы ДНаТ-400 и све-
ответствии с приложением Д ГОСТ Р 55702-2013,
тодиодные светильники с различным спектраль-
оптические свойства оттестированы по сравне-
ным составом излучения, обозначенные далее
нию с эталонной поверхностью сферы EVER-
следующим образом: 1) S - дуговая натриевая
FINE (EVERFINE Corporation, Китай). Спек-
трубчатая лампа, 2) Y - светодиодный аналог
тральный коэффициент отражения краски в ви-
ДНаТ-400 (ООО «О2 Световые системы» и Агро-
димом диапазоне составил 85 ± 5%. В качестве
физический НИИ, Санкт-Петербург), излучаю-
источника излучения света в схеме использовали
щий желтый свет с максимумами излучения на
специализированный светодиодный светильник
длинах волн 450, 500 и 595 нм и соотношением
разработки российской инжиниринговой компа-
интенсивностей этих пиков 1 : 3 : 13, 3) P - аналог
нии «О2 Световые системы» (Санкт-Петербург) с
используемых на данный момент в тепличных
регулируемыми каналами излучателей (светодио-
комплексах светодиодных светильников (ООО
дов), рассчитанными под фотосинтетически ак-
«О2 Световые системы», Санкт-Петербург), из-
тивный диапазон и позволяющий работать в фи-
лучающий розовый свет и имеющий максимумы
зиологически важных для растения областях
излучения на длинах волн 450 и 665 нм с соотно-
спектра с максимумами в диапазонах длин волн
шением интенсивностей 1 : 2, 4) W - светодиод-
650-670 нм, 720-740 нм, 440-460 нм, 415-435 нм,
ный светильник с промышленным белым светом
а также белый спектр с коррелированной цвето-
с соотношением максимумов 1 : 1 на длинах волн
вой температурой 4500 К. Интенсивность излуче-
450 и 600 нм, 5) B - промышленный светодиод-
ния каждого канала управляется драйвером, что
ный светильник с белым светом и с соотношени-
позволяет формировать необходимый спектр и
ем максимумов 1 : 3 на длинах волн 450 и 600 нм.
световой поток при воздействии на объект иссле-
Фактически Y отличается от светильников W и В
дования. Световой поток от светильника собира-
большей интенсивностью излучения в зеленой и
ется с помощью плоско-выпуклой линзы и через
красной частях спектра, а светильник P имеет
коллиматор 74-UV (Ocean Optics, США) с расхо-
смещенный максимум в красной области с 600 до
димостью пучка менее 2° заводится в интегриру-
665 нм.
ющую сферу ИС1, таким образом, освещается ис-
следуемая область образца диаметром 12 мм. Де-
В табл. 1 приведены измеренные с помощью
тектором служит оптический оптоволоконный
интегрирующей сферы и ПЗС-спектрометра
ПЗС-спектрометр CCS200 (Thorlabs, США) с
CCS200 (Thorlabs, США) и усредненные по всей
диапазоном регистрации спектра 200-1000 нм и
поверхности выращивания спектры излучения
шагом длины волны <2 нм, соединенный с инте-
использованных светильников. На основе пред-
грирующими сферами с помощью оптоволокна
ложенного в работе [24] алгоритма и полученных
BFL200HS02 (Thorlabs, США) диаметром 200 мкм.
экспериментально спектров излучения рассчита-
Исследуемый образец помещается между двумя
на плотность фотосинтетически активной радиа-
сферами, одна из которых (ИС1) регистрирует от-
ции во всем видимом диапазоне и в условно раз-
раженный от объекта свет, другая (ИС2) - про-
деленных на синюю (400-500 нм), зеленую (500-
шедший сквозь него свет. Коэффициент погло-
600 нм) и красную (600-700 нм) части спектра.
щения листа А(λ) рассчитывается по формуле
Для освещения растений величину облучен-
ности от различных источников света установи-
А(λ) = 1 - R(λ) - T(λ),
(1)
ли равную 70-75 Вт/м2 фотосинтетически ак-
где R(λ) - коэффициент отражения образца, рав-
тивной радиации, продолжительность светового
ный отношению отраженного потока излучения
периода - 14 ч в сутки. Поддерживали относи-
к потоку, упавшему на тело, T(λ) - коэффициент
тельную влажность воздуха 65-70% иемперату-
пропускания образца, равный отношению пото-
БИОФИЗИКА том 65
№ 1
2020
ВЛИЯНИЕ СПЕКТРАЛЬНЫХ ОСОБЕННОСТЕЙ СВЕТОВОЙ СРЕДЫ
115
Таблица 1. Величина плотности фотосинтетически активной радиации и скорость усвоения листьями салата
света различного спектрального состава
Спектральный
Спектры поглощения
состав
Поглощение 400-700 нм
света в диапазоне 400-
PPFD, мкмоль · м-2 · с-1
излучения
21-й день → 31-й день
700 нм на 21-й и 31-й
Источник
источников
дни развития растений
света
света
в диапазоне
400-500
500-600
600-700
400-500
500-600
600-700
400-700 нм
нм
нм
нм
нм, %
нм, %
нм, %
S,
341 ± 24 (+72 в ИК)
42 ± 5% → 57 ± 3%
лампа
143 → 194 мкмоль · м-2 · с-1
ДНаТ
29
185
127
19 → 23
10 → 14
13 → 20
Y,
41 ± 6% → 56 ± 9%
желтый
140 → 191 мкмоль · м-2 · с-1
342 ± 24
23
156
163
19 → 22
9 → 14
13 → 20
P,
39 ± 4% → 51 ± 11%
розовый
129 →1 70 мкмоль · м-2 · с-1
332 ± 23
61
123
148
18 → 21
9 → 11
12 → 18
W,
58 ± 13% → 62 ± 8%
белый
189 → 201 мкмоль · м-2 · с-1
325 ± 23
69
150
106
23 → 24
14 → 16
20 → 21
B,
50 ± 1% → 62 ± 3%
белый
167 → 207 мкмоль · м-2 · с-1
334 ± 23
39
162
133
22 → 24
10 → 16
18 → 22
Примечание. PPFD - плотность фотосинтетически активной радиации (photosynthetic photon flux density).
БИОФИЗИКА том 65
№ 1
2020
116
КУЛЕШОВА и др.
мых объектов произведена по всей зоне роста.
Спектры поглощения регистрировали для обла-
сти площадью 115 мм2 для центральной части ли-
ста на 21-й день развития (рис. 2б), для централь-
ной части и верхушки на 31-й день развития
(рис. 2в).
Для оценки влияния параметров освещения на
поглощение света листьями салата мы предложи-
ли использовать первую оценочную версию вели-
чины, потенциально описывающей приращение
поглощения света на прирост биомассы и обозна-
ченный нами в дальнейшем как индекс светопо-
глощения I. За первоначальное поглощение при-
нято среднее значение между поглощением света
21-дневным листом салата, имеющего общую
площадь, сравнимую с окном интегрирующей
сферы, и центральной частью 31-дневного листа,
предположительно не отличающегося по своей
структуре от той же области более раннего перио-
да развития. Разница между поглощением «но-
вой» и «старой» частей листа ΔA нормируется на
Рис. 1. Схема экспериментальной установки для из-
значение накопленной за период между измере-
мерения спектров пропускания и отражения листьев
ниями биомассы Δm. Таким образом, на данном
растений, включающей в себя: 1 - косинусный кор-
этапе индекс светопоглощения определяет вели-
ректор, 2 - собирающую линзу, 3 - источник света,
4 - интегрирующую сферу ИС1, 5 - коллиматор, 6 -
чину поглощения фотосинтетически активного
шторку, 7 - оптоволокно, 8 - спектрометр, 9 - обра-
потока фотонов, необходимого для прироста 1 г
зец, 10 - интегрирующую сферу ИС2, 11 - штатив.
биомассы за исследуемый диапазон времени ро-
ста при заданных условиях освещения, и рассчи-
тывается по следующей формуле:
ка излучения, прошедшего через среду, к потоку
A
+
A
излучения, упавшему на ее поверхность.
31D
21D
A
−
31U
ΔA
2
I
=
=
,
(2)
Зоны отбора проб из среды роста салата схема-
Δm
m
31
−
m
21
тично показаны на рис. 2а, для каждого варианта
освещения и концентрации раствора взяты по
где I - индекс светопоглощения (мкмоль · м-2 · с-1),
пять тестируемых объектов, выборка тестируе-
A31U - интегральное поглощение света в диапазо-
Рис. 2. Зоны (1-5) отбора проб из среды роста салата (а); условная область измерения поглощения света на листовой
пластине салата - в центральной части листа на 21-й день развития (б) и в центральной части и верхушке листа на
31-й день развития (в).
БИОФИЗИКА том 65
№ 1
2020
ВЛИЯНИЕ СПЕКТРАЛЬНЫХ ОСОБЕННОСТЕЙ СВЕТОВОЙ СРЕДЫ
117
Таблица 2. Влияние источников света различного спектрального состава на рост и продуктивность растений
салата в регулируемых условиях биополигона Агрофизического НИИ
Сухое вещество
Сухое вещество
Масса надземной
Высота растений
Площадь листа
листьев
корней
части
Источники
света*
% от
% от
% от
% от
% от
см
см2
%
%
г/м2
контроля
контроля
контроля
контроля
контроля
S
20.6 ± 1.0
100
97.2
100
3.7
100
7.4
100
5990
100
Y
18.0 ± 0.7
87
70.7
73**
4.2
114
6.3
85**
3350
56**
P
17.1 ± 0.6
83**
74.3
76**
3.9
105
6.6
89
3090
52**
W
13.3 ± 0.4
65**
41.2
42**
4.8
130**
6.3
85**
1920
32**
B
12.9 ± 0.5
63**
30.6
32**
5.4
146**
6.7
91
2250
38**
Примечание. * - Источники света: S -лампы ДНаТ-400; Y - светодиодный аналог ДНаТ; P - аналог светодиодных
фитосветильников с максимумами излучения на длинах волн 450 и 665 нм; W - светодиодный светильник с промышленным
белым светом; B - светодиодный светильник с промышленным белым светом. ** - Значение достоверно отличается от
контрольного на 5%-м уровне значимости.
не
400-700 нм верхушкой
31-дневного листа
тивных документов и по общепринятым методи-
кам [27-29].
(мкмоль · м-2 · с-1), A31D - интегральное погло-
щение света центральной частью 31-дневного ли-
Дисперсионный и регрессионный анализы, а
ста (мкмоль · м-2 · с-1), A21D - интегральное по-
также другая статистическая обработка обобщае-
мых данных выполнены с помощью программно-
глощение света центральной частью 21-дневного
го обеспечения MS Excel 2010 и Statistica 8. В тек-
листа (мкмоль · м-2 · с-1), m31 - масса исследуе-
сте и таблицах приведены средние арифметиче-
мого листа 31-дневного салата (г), m21 - масса ис-
ские значения параметров и их доверительные
следуемого листа 21-дневного салата (г).
интервалы при 95%-м уровне вероятности по
t-критерию.
Реакция овощных культур на моделируемые
условия световой и корнеобитаемой среды оце-
нивалась по показателям роста, нетто-продук-
РЕЗУЛЬТАТЫ
тивности и биохимическому составу получаемой
растительной продукции. На протяжении вегета-
Результаты анализа данных по росту, разви-
ционных периодов также проводили фенологи-
тию, продуктивности растений салата и качеству
ческие наблюдения. Уборку салата проводили на
его растительной продукции свидетельствуют,
31-е сутки от высева семян. При уборке учитыва-
что в большинстве своем они имели достоверно
ли массу растений, длину, ширину максимально-
более высокие значения у растений, выращенных
го развитого листа, высоту растений, число ли-
под лампами ДНаТ по сравнению с таковыми под
стьев, определяли содержание нитратов и других
светодиодными светильниками при одинаковых
биохимических показателей.
облученности и условий микроклимата (табл. 2).
Биохимический состав растительной продук-
В связи с этим значения показателей роста, про-
ции, характеризующий ее качество и безопас-
дуктивности растений салата, качества его расти-
ность, определяли в аккредитованной на техни-
тельной продукции под лампами ДНаТ были
ческую компетентность и независимость Испы-
выбраны как референтные. Так, по показателям
тательной лаборатории Агрофизического НИИ
роста, в частности, по высоте растения, сформи-
Россельхозакадемии (Санкт-Петербург) в соот-
рованные под лампами ДНаТ, превышали в виде
ветствии с требованиями современных норма-
выраженной тенденции или достоверно (на 13-
БИОФИЗИКА том 65
№ 1
2020
118
КУЛЕШОВА и др.
Таблица 3. Биохимический состав растительной продукции салата в зависимости от спектра облучения растений
под различными источниками света в регулируемых условиях биополигона Агрофизического НИИ
Источники света*
Содержание
S
Y
P
W
B
Витамин С, мг/100 г.н.в.
16.61
11.99**
12.65**
8.10**
11.09**
Сумма сахаров, % а.с.в.
19.31
13.61**
11.31**
12.89**
16.04
Нитраты, мг/кг н.в.
1560
2154**
1920**
3043**
1423
Азот, % а.с.в.
3.51
3.64
4.11**
4.28**
3.72
Фосфор, % а.с.в.
0.58
0.72**
0.76**
0.57
0.61
Калий, % а.с.в.
7.7
8.18
9.25**
11.20**
8.36
Кальций, % а.с.в.
2.29
1.95**
2.35
2.02
2.31
Магний, % а.с.в.
0.604
0.567
0.625
0.468**
0.587
Свинец, мг/кг а.с.в.
<0.10
<0.10
<0.10
<0.10
<0.10
Кадмий, мг/кг а.с.в.
<0.010
<0.010
<0.010
<0.010
<0.010
Примечание. * - Источники света: S -лампы ДНаТ-400; Y - светодиодный аналог ДНаТ; P - аналог светодиодных
фитосветильников с максимумами излучения на длинах волн 450 и 665 нм; W - светодиодный светильник с промышленным
белым светом; B - светодиодный светильник с промышленным белым светом. ** - Значение достоверно отличается от
контрольного на 5%-м уровне значимости.
37%) таковые под светодиодными светильника-
Данные оценки биохимического состава рас-
ми; по площади листьев - на 27-68%; по массе
тений свидетельствуют о более низком содержа-
надземной части на квадратном метре или про-
нии витамина С, сахаров в листьях салата, выра-
дуктивности - на 44-62%. Следует отметить, что
щенного под светодиодными светильниками по
сравнению салатом, выращенным под лампами
существенно более низкие значения оценивае-
ДНаТ-400 (табл. 3). При этом в листьях, за исклю-
мых показателей были у растений под промыш-
чением варианта с промышленными светильни-
ленными светильниками белого света W и B. Рас-
ками белого света, отмечается более высокое со-
тения под светодиодными светильниками с жел-
держание нитратов, даже превышающее значе-
тым (Y) и розовым (P) светом имели несколько
ния ПДК или близкое к ее граничным значениям
лучшие значения показателей роста и продуктив-
в остальных вариантах с светодиодными светиль-
ности, чем под светодиодными светильниками W
никами. Следует отметить также несколько более
и В, но также значимо уступали таковым под лам-
высокое содержание макроэлементов - общего
пами ДНаТ. Судя по изменению процентного со-
азота, фосфора, калия - в надземной части салата
держания сухого вещества, в корнях растений под
при падении показателей роста и продуктивности
светодиодными источниками оно преимуще-
под светодиодными светильниками, по сравне-
ственно несколько ниже, а в листьях - выше, чем
нию с растениями под лампами ДНаТ-400.
под лампами ДНаТ-400. С учетом более низких
Выявленные различия в показателях роста,
показателей роста и продуктивности выявленные
развития, продуктивности и в биохимическом со-
закономерности позволяют предположить, что
ставе растений салата, сформированных под лам-
растения в менее благоприятных условиях свето-
пами ДНаТ-400 и под светодиодными светильни-
вой среды под тестируемыми светодиодными све-
ками с различным спектральным составом, обу-
тильниками пытаются адаптироваться к ним по-
словлены особенностями его влияния на
средством синтеза и накопления в листьях раз-
процессы поглощения и усвоения света листьями
личных соединений защитного плана, как это
растений. В ходе вегетационного периода сала-
отмечается в литературе для растений, выращива-
тов, освещаемых различным спектром излуче-
емых под источниками света с высокой интен-
ния, на 21-й и 31-й дни развития, были сняты
сивностью излучения в сине-фиолетовой области
спектры поглощения света центральной частью и
спектра [30].
верхушкой листовой поверхности (табл. 1). Рас-
БИОФИЗИКА том 65
№ 1
2020
ВЛИЯНИЕ СПЕКТРАЛЬНЫХ ОСОБЕННОСТЕЙ СВЕТОВОЙ СРЕДЫ
119
Рис. 3. Отношения коэффициентов максимального поглощения, соответствующих разным длинам волн - A440/А667
(а) и A460/А620 (б) для листовой поверхности 21-дневного (темные столбики) и верхушки листа 31-дневного салата
(светлые столбики), освещаемого лампой ДНаТ S и светодиодными светильниками с различными спектрами
освещения Y, P, W и B.
считано количество поглощенного листьями фо-
ционально соотношениям для центральной части
тосинтетически активного потока фотонов, рас-
21-дневного и 31-дневного листьев.
тущее с увеличением возраста растения, спектр
Получено различие в форме спектральных
поглощения условно разделен на синий (400-500
кривых для образцов, освещаемых ДНАТ (S),
нм), зеленый (500-600 нм) и красный (600-700
желтым (Y), розовым (P) и двумя вариантами бе-
нм) диапазоны, для которых также рассчитан
лого (W, B) источниками света. В области 470-
процент поглощенного светового потока на раз-
640 нм различия в интенсивностях достигают
ных стадиях развития.
15%, хотя при этом величины коэффициентов по-
Измеренные значения суммарного поглоще-
глощения на основных максимумах длин волн
ния в диапазоне длин волн от 400 до 700 нм варьи-
440 и 667 нм в большинстве случаев совпадают.
руют от 42 до 62 %, причем наибольшая доля по-
Также важно отметить разницу между спектрами
глощения приходится на синюю область от 400 до
поглощения 21-дневных и 31-дневных растений:
500 нм и составляет 18-24%, для красной (600-
в первом случае на спектрах видно наличие более
700 нм) области характерны значения от 12 до
четко выраженных максимумов на длинах волн
22% и в зеленой (500-600 нм) области листья об-
480, 500, 590 и 620 нм.
ладают наименьшим поглощением - 9-16%. При
Наибольшее различие между вариантами вы-
освещении лампами ДНаТ, желтым и розовым
ражено в соотношении максимальных интенсив-
светом интегральное поглощение света листьями
ностей поглощения, соответствующих максиму-
схоже на 21-й и на 31-й день развития в централь-
мам поглощения хлорофилла а (440 нм и 667 нм)
ной части листьев, но увеличивается в области их
и хлорофилла b (460 нм и 620 нм). В качестве ха-
верхушки. Этот экспериментальный результат
рактеристики, определяющей форму полученных
коррелирует с представлением о нарастании био-
спектральных кривых, мы использовали отноше-
массы листа - на 31-й день «старая» часть листа
ние максимумов коэффициентов поглощения:
поглощает так же, как и раньше (на 21-й день), а
A440/А667 и A460/А620, соответствующие длинам
новая - верхушка поглощает больше энергии
волн 440 нм к 667 нм и 460 нм к 620 нм (рис. 3).
квантов для ускорения развития. Однако такая
динамика не наблюдается при освещении белым
Соотношение поглощения света с длинами
светом - значения поглощения практически для
волн 440 и 667 нм A440/А667, потенциально свя-
всех случаев идентичны. Это объясняется замед-
занных с количеством хлорофилла а, с увеличе-
ленным ростом и отставанием в развитии листьев
нием периода развития растения уменьшается и
салата в этом варианте облучения. Также заметна
на 31-й день для всех вариантов отношение этих
разница в соотношении поглощения по областям
максимумов становится равным единице. Преоб-
спектра. На 31-й день верхушка листа почти срав-
ладающее на более ранних этапах роста поглоще-
нивает поглощение света в синей и красной обла-
ние света синей части спектра для вариантов cо
стях - разница составляет <3%, при этом погло-
светильниками S, Y, P выравнивается с поглоще-
щение в зеленой части спектра остается пропор-
нием красного диапазона в конце вегетационного
БИОФИЗИКА том 65
№ 1
2020
120
КУЛЕШОВА и др.
спектров освещения на спектры поглощения ли-
стьев растений в процессе их развития, что позво-
ляет предложить использовать анализ спектра по-
глощения листа растения как один из вариантов
фитомониторинга. Разработанный и использо-
ванный в работе метод спектрометрического ана-
лиза с помощью системы из двух интегрирующих
сфер хорошо применим и достаточно удобен для
измерения спектров поглощенной энергии ли-
стьями салата в ходе их жизнедеятельности и дает
возможность получать информацию о поглоще-
нии излучения от источника освещения и связан-
ных с этим фотосинтетических реакциях.
Взятый в качестве референтного спектр осве-
щения дуговой натриевой трубчатой лампы дей-
ствительно показал лучшие результаты по пока-
зателям нетто-продуктивности салата и поглоще-
Рис. 4. Индекс светопоглощения - приращение по-
нию света листьями растений. В связи с тем, что
глощения света на прирост биомассы для салата,
для описания эффективности поглощения света
освещаемого источниками света с различным спек-
листьями на определенной длине волны нет об-
тральным составом: S - лампами ДНаТ-400, Y - све-
тодиодным аналогом ДНаТ, P - аналогом используе-
щепринятой количественной характеристики,
мых в тепличных комплексах светодиодных светиль-
мы использовали значения, полученные для об-
ников, W - светодиодным светильником с белым
разцов, освещаемых лампами ДНаТ, как кон-
светом и соотношением максимумов 1:1, B - светоди-
трольные. Самые приближенные к ним результа-
одным светильником с белым светом и с соотноше-
нием максимумов 1 : 3.
ты наблюдали при использовании светодиодных
светильников Y, разработанных ООО «О2 Свето-
вые системы» совместно с Агрофизическим НИИ
периода. Различия в соотношении A460/А620, по-
и ориентированных по своим спектральным
тенциально характеризующих содержание хлоро-
свойствам на лампы ДНаТ.
филла b, менее выражено для всех вариантов, од-
Данные по показателям роста, продуктивно-
нако заметна тенденция к увеличению его значе-
сти и биохимическому составу салата позволили
ния с ростом растения для источников света S, Y,
сделать следующие заключения о влиянии свето-
P и уменьшение для светильников W, B.
вой среды на растения. Максимальные высота,
площадь листьев, продуктивность растений, а
Для того чтобы охарактеризовать, каким обра-
также более высокие качественные характери-
зом меняется поглощение с ростом растения, на-
стики растительной продукции были зафиксиро-
ми был введен индекс светопоглощения (форму-
ваны при освещении растений лампами ДНаТ
ла (2)), рассчитанные значения которого для те-
при одинаковых облученности растений и усло-
стируемых вариантов приведены на рис.
4.
вий микроклимата. Под светодиодными светиль-
Интересно отметить совпадение значений индек-
никами Y и P высота растений была снижена на
са при освещении салата натриевой лампой и ее
20%, а под светильниками W и B - отличалась по-
светодиодным аналогом. Данные условия были
чти на 40%. Более сильные отличия заметны в
наиболее эффективными и привели к быстрому
размере площади листьев, где разница между па-
приросту биомассы и лучшему развитию расте-
раметрами растений, освещаемых светильника-
ния. Из этого можно сделать вывод, что с приро-
ми S и светодиодными источниками W и B, до-
стом биомассы листа салата на 1 г в течение 10 су-
стигает почти 70%. Это говорит о замедленном
ток оптимальным является увеличение количе-
развитии салата при его облучении белым светом,
ства поглощенной энергии фотонов на
излучаемым промышленными светильниками W
~140 мкмоль · м-2 · с-1.
и B. В результате такое снижение показателей ро-
ста и развития растений салата под влиянием те-
стируемых светодиодных источников света зако-
ОБСУЖДЕНИЕ
номерно приводит к снижению значений продук-
Для понимания механизма поглощения фото-
тивности на 44-62%. При этом растения под
нов листьями растений и выявления роли длины
светодиодными светильниками с желтым (Y) и
волны световой энергии был проведен экспери-
розовым (P) светом имели несколько лучшие зна-
мент, который заключался в облучении растений
чения показателей роста и продуктивности, чем
салата светом с различным спектральным соста-
под светодиодными светильниками W и В, но
вом и регистрации поглощенного светового пото-
значимо уступали таковым под лампами ДНаТ.
ка. В ходе исследования была выявлено влияние
Более низкое содержание сухого вещества в кор-
БИОФИЗИКА том 65
№ 1
2020
ВЛИЯНИЕ СПЕКТРАЛЬНЫХ ОСОБЕННОСТЕЙ СВЕТОВОЙ СРЕДЫ
121
нях и более высокое его накопление в листьях
Через 10 дней разница в поглощении в области
растений, сформированных под тестируемыми
600-700 нм для всех вариантов не превышает 4%.
светодиодными источниками света Y, P, W и B,
В большинстве случаев действие этого диапазона
при существенно более низкой массе надземной
связывают с морфологическими изменениями и
части салата, предположительно свидетельствует
стимуляцией роста растений, однако в проведен-
об образовании и накоплении в них веществ за-
ных нами экспериментах отмечено отставание в
щитной природы. Образование этих веществ свя-
развитии для образцов, потреблявших большее
зано с расходованием энергии и ресурсов и отри-
количество «красных» фотонов на ранних этапах.
цательным образом отражается на продуктивно-
В диапазоне 500-600 нм для верхушки 31-дневно-
сти и качестве урожая растительной продукции.
го листа салата характерно большее поглощение
Косвенным подтверждением этого факта являет-
света в вариантах освещения светодиодными све-
ся увеличенное поглощение листьев в области
тильниками W и B. Известно, что зеленый свет
470-640 нм для растений, освещаемых белым
необходим оптически более плотным листьям,
светом. Возможно, это связано с повышенным
так как обладает высокой проникающей способ-
содержанием пигментов, выполняющих защит-
ностью. Благодаря этому излучение данного диа-
ные функции и синтезирующихся в стрессовых
пазона достигает нижних ярусов и восполняет на-
условиях - каротиноидов с максимумом погло-
ходящимся там листьям недостающую для проте-
щения в районе 480 нм и антоцианов, поглощаю-
кания фотосинтеза энергию. Также в образцах
щих в области 500-520 нм.
салата, облучаемых светодиодными светильника-
О неоптимальности спектрального состава те-
ми W и B с белым светом, содержание сухого ве-
стируемых светодиодных светильников по срав-
щества больше, чем в других образцах, что гово-
нению с лампами ДНаТ свидетельствуют и дан-
рит о более плотной структуре листа. Максималь-
ные оценки биохимического состава растений, а
ная доля поглощения в области 400-500 нм
именно: отмечается снижение содержания вита-
свойственна для растений, освещаемых белым
мина С и содержания сахаров в листьях салата
светом и показавших наименьшее показатели по
(табл. 3). При этом в них, за исключением вари-
продуктивности. На первый взгляд, данный эф-
анта с промышленными светильниками белого
фект повышенного поглощения в синем диапазо-
света, отмечается более высокое содержание нит-
не спектра напрямую не коррелирует с более ин-
ратов, даже превышающее значения ПДК или
тенсивным излучением источника света в обла-
близкое к ее граничным значениям в остальных
сти 400-500 нм. Большее поглощение листьями
вариантах с светодиодными светильниками. Не-
растений в этой части спектра, с одной стороны,
сколько более высокое содержание макроэлемен-
может быть связано с протеканием гравиотропи-
тов - общего азота, фосфора, калия - в надзем-
ческих реакций, а с другой - с работой фотосин-
ной части салата при падении показателей роста и
тетического аппарата. Высота салата, выращен-
продуктивности под светодиодными светильни-
ного под лампой S, светильниками Y и P достига-
ками, по сравнению с таковым под лампами
ет 20 см, тогда как для образцов, освещаемых
ДНаТ-400, позволяет предположить более низ-
светильниками W и B и поглощающих меньшее
кую интенсивность физиологических процессов
количество света в диапазоне 400-500 нм от инте-
фотосинтеза, роста и развития, где данные соеди-
грального, высота составляет 10-13 см.
нения используются и трансформируются, а так-
Рассмотрим классическое представление ор-
же о расходовании пластичных веществ растений
ганизации светособирающего комплекса пиг-
салата в процессах его адаптации к неблагоприят-
ным световым условиям среды обитания.
ментов как антенны, представив упрощенную
модель антенного комплекса и условно разделив
Данные по поглощению света листьями салата
в зависимости от спектра их освещения (табл. 1)
входящие в нее пигменты на поглощающие в си-
можно охарактеризовать следующим образом.
ней (400-500 нм), зеленой (500-600 нм) и крас-
Поглощение в синей (400-500 нм), зеленой (500-
ной (600-700 нм) областях спектра (рис. 5). Для
600 нм) и красной (600-700 нм) областях спектра
возбуждения реакционных центров P680 и P700,
варьирует в пределах 40-49%, 19-25%, 30-39% от
осуществляющих превращение энергии света в
интегрального соответственно на 21-й день раз-
химическую, необходимо поглощение фотонов с
вития и 37-42%, 22-28%, 35-36% - на 31-й день.
энергиями 1.82 и 1.77 эВ. При поглощении свето-
Хотя излучение от источников света во всех вари-
собирающими пигментами квантов больших
антах в синем диапазоне (400-500 нм) меньше,
энергий (например, в синем диапазоне 2.48-
чем в зеленом и красном, поглощение фотонов
3.09 эВ) электроны переходят на орбитали с более
энергией 2,5-3,1 эВ листьями выше других и до-
стигает почти половины от суммарного. Для
высоким энергетическим уровнем, и перенос
21-дневного салата отмечено большее поглоще-
энергии на нижние уровни (первый синглентный
ние излучения в красной части спектра (на 6-8%)
уровень реакционного центра и переход в основ-
при освещении обоими вариантами белого света.
ное состояние) возможен только с потерей энер-
БИОФИЗИКА том 65
№ 1
2020
122
КУЛЕШОВА и др.
жить, что поглощение одинакового количество
квантов разных энергий ведет к стимуляции и
развитию пигментного аппарата, что в свою оче-
редь повышает фотосинтез и скорость развития
растений.
Важно отметить, что непотраченная на фото-
синтез энергия используется растениями на осу-
ществление других процессов. Об этом свиде-
тельствует различие в поглощении светочувстви-
тельными пигментами в растворе от поглощения
листьями in vivo, наиболее заметное в зеленой об-
ласти спектра.
На основании проведенных комплексных ис-
следований установлено наибольшее положи-
Рис. 5. Схема передвижения и превращения энергии
тельное влияние ламп ДНаТ на рост, продуктив-
по антенному комплексу от светособирающих пиг-
ность и качество растений салата сорта Тайфун
ментов к реакционному центру с переходами между
при выращивании его в регулируемых условиях
возбужденными состояниями пигментов после по-
глощения синего, зеленого и красного света.
интенсивной светокультуры, что связано с более
эффективным поглощением и усвоением света
листьями растений по сравнению с наблюдаемым
гией - выделением в виде света или тепла. Пере-
под тестируемыми светодиодными источниками
нос энергии от одной молекулы пигмента к дру-
света с различным спектральным составом. Сре-
гой или к реакционному центру осуществляется с
ди последних наиболее близкое к референтному
большой скоростью (~10-10-10-12 с), но только
влияние на растение отмечается под светодиод-
между близлежащими молекулами. Несомненно,
ными светильниками - аналогами ДНаТ, наи-
такая организация значительно увеличивает эф-
меньшее - под светильниками с белым светом W
фективность усвоения света за счет обеспечения
и В. Светодиодные светильники по степени поло-
пигмент-белковыми комплексами захвата фото-
жительного влияния на растения их спектрально-
нов разных длин волн и транспорта энергии воз-
го состава света расположились в ряду: аналог
буждения в реакционные центры. Однако, во-
ДНаТ ≥ аналог тепличных светильников с макси-
первых, «лишняя» энергия, испускаемая в виде
мумами излучения на длинах волн 450 и 665 нм >
флуоресценции или теплоты, не используется на
белый светодиодный светильник с соотношени-
фотосинтез. Во-вторых, в сооответствии с полу-
ем максимумов излучения на длинах волн 450 нм
ченными экспериментальными данными, при
к 600 нм 1 : 3 > белый светодиодный светильник с
избыточном интегральном поглощении (167 и
соотношением максимумов излучения на длинах
волн 450 нм к 600 нм 1 : 1.
188 мкмоль · м-2 · с-1 для вариантов со светильни-
ками W и B) наблюдается замедленное развитие,
С приростом биомассы листа салата на 1 г в те-
что, возможно, связано с тем, что пигменты, воз-
чение 10 суток оптимальным является увеличе-
бужденные коротковолновым светом не успева-
ние количества поглощенной энергии фотонов
ют передавать энергию на пигменты, которые
на ~140 мкмоль · м-2 · с-1. Такой вывод был сде-
также перешли в возбужденное состояние, но при
лан исходя из того, что это значение характерно
поглощении фотонов с большей длиной волны.
для референтного варианта, взятого за эталон и
Возникает конкуренция за перенос энергии на
показавшего наилучшие результаты практически
фотоактивный пигмент.
по всем показателям. Такое же значение прира-
Рассчитанный коэффициент А440/А667, опре-
щения поглощения было получено для варианта
освещения светодиодным светильником Y, ори-
деляющий «сглаженность» спектра поглощения -
ентированным по спектральным характеристи-
чем он ближе к единице, тем меньше разность
кам в области фотосинтетически активной ради-
между доминирующими максимумами, подтвер-
ации на ДНаТ. Необходимо отметить, что значе-
ждает, что преобладание поглощения в синей об-
ние индекса светопоглощения ниже 70 мкмоль ·
ласти приводит к более развитой миграции энер-
гии по фотосинтетической пигментной матрице.
· м-2 · с-1 соответствует образцам с минимальны-
Так как с увеличением этого коэффициента рас-
ми результатами по продуктивности и ростовым
тет продуктивность растений, можно предполо-
характеристикам.
БИОФИЗИКА том 65
№ 1
2020
ВЛИЯНИЕ СПЕКТРАЛЬНЫХ ОСОБЕННОСТЕЙ СВЕТОВОЙ СРЕДЫ
123
Таким образом, полученные данные свиде-
10.
О. В. Аверчева, Ю. А. Беркович, А. Н. Ерохин и др.,
тельствуют о том, что спектр освещения, излуча-
Физиология растений 56, 17 (2002).
ющий ~25 мкмоль · м-2 · с-1 в диапазоне 400-
11.
Anderson Jr, W. Grant, and L. S. Capen, U.S. Patent
№ 6921182. Washington, DC: U.S. Patent and Trade-
500 нм, ~150 мкмоль · м-2 · с-1 в диапазоне 500-
mark Office (2005).
600 нм, ~150 мкмоль · м-2 · с-1 в диапазоне 600-
12.
А. А. Шахов, В. С. Хазанов и С. А. Станко, Ботан.
700 нм приводит к высокой продуктивности сала-
журн. 46 (2), 222 (1961).
та. Схожесть спектральных характеристик осве-
13.
А. Б. Брандт и С. В. Тагеева, Оптические парамет-
щения в области фотосинтетически активной ра-
ры растительных организмов (Наука, М., 1967).
диации ламп ДНаТ и светодиодных светильников
14.
И. С. Лискер, в кн. Физические методы и средства
Y, излучающих желтый свет, и практически оди-
получения информации в агромониторинге (АФИ, Л.,
наковой доли фотосинтетиечски активных фото-
1987), сс. 3-21.
нов в синей, зеленой и красной областях спектра
15.
М. В. Архипов, В. Н. Савин, Е. В. Канаш и др.,
привела к одинаковому приращению поглоще-
в кн. Биофизика растений и фитомониторинг
ния света (~140 мкмоль · м-2 · с-1) в процессе раз-
(АФИ, Л., 1990), сс. 186-208.
вития растений. Однако значительное отличие в
16.
В. С. Лысенко, Т. В. Вардуни, В. Г. Сойер и
продуктивности салата (~50%), а также росте,
В. П. Краснов, Фундаментальные исследования 1
развитии и биохимическом составе свидетель-
(4), 112 (2013).
ствует о лучшем влиянии ламп ДНаТ на свойства
17.
G. H. Krause and E. Weis, Annu. Rev. Plant Biol. 42
растительной культуры. Возможно, это связано с
(1), 313 (1991).
наличием излучения в ИК-диапазоне, исследова-
18.
M. Mõttus, A. Hovi, and M. Rautiainen, Appl. Optics
ние влияния которого является предметом изуче-
56 (3), 563 (2017).
ния в дальнейших экспериментах.
19.
Е. М. Басарыгина, О. Г. Лицингер и Т. А. Путило-
ва, АПК России 24 (5), 1141 (2017).
КОНФЛИКТ ИНТЕРЕСОВ
20.
A. Hovi, P. Raitio, and M. Rautiainen, Silva Fenn. 51,
Авторы заявляют об отсутствии конфликта
1 (2017).
интересов.
21.
Ю. И. Желтов и Г. Г. Панова, Патент РФ на полез-
ную модель №108705, Бюл. Федеральной службы
по интеллектуальной собственности, патентам и
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
товарным знакам, № 27 (2011).
Настоящая работа не содержит описания ка-
22.
G. G. Panova, I. N. Chernousov, O. R. Udalova, et al.,
ких-либо исследований с использованием людей
Reports of the Academy of Agricultural Sciences
и животных в качестве объектов.
[Doklady RASHN] 4, 17 (2015).
23.
В. А. Чесноков, Е. Н. Базырина, Т. М. Бушуева,
Выращивание растений без почвы (Изд-во ЛГУ, Л.,
СПИСОК ЛИТЕРАТУРЫ
1960).
1. S. Kurihara, T. Ishida, M. Suzuki, and A. Maruyama,
24.
Т. Э. Кулешова, М. Н. Блашенков, Д. О. Кулешов
Focusing on Modern Food Industry 3, 1 (2014).
и Н. Р. Галль, Научное приборостроение 26 (3), 35
2. O. Hitoshi, H. Tatsuya, К. Kouji, and N. Yoshifumi,
(2016).
Environ. Control Biol. 53 (2), 93 (2015).
25.
T. E. Kuleshova, I. S. Seredin, S. A. Cheglov, et al., J.
3. J. E. Park and K. Nakamura, Environ. Control Biol. 53
Phys.: Conf. Ser. 1135, 012013 (2018).
(2), 89 (2015).
26.
J. W. Pickering, S. A. Prahl, N. Van Wieringen, et al.,
4. N. Yeh and J. P. Chung, Renewable and Sustainable
Appl. Optics 32 (4), 399 (1993).
Energy Reviews 13 (8), 2175 (2009).
27.
В. А. Тутельян и Е. Н. Беляев, СанПиН 2.3.2.1078-
5. Y. Qi-chang, J. Agricultural Sci. Technol. 6, 42 (2008).
01 (2001).
6. С. А. Ракутько и А. Е. Пацуков, Світлотехніка та
28.
А. И. Ермаков, В. В. Арасимович и Н. П. Ярош,
електроенергетика 2, 18 (2013).
Методы биохимического исследования растений
7. А. В. Аладов, Е. Д. Васильева, А. Л. Закгейм и др.,
(Агропромиздат (Ленинградское отд-ние), Л.,
Светотехника 3, 8 (2010).
1987).
8. G. Tamulaitis, P. Duchovskis, Z. Bliznikas, et al., In
29.
И. М. Скурихина и В. А. Тутельяна, Руководство по
Abstr. Book of 4th Int. Conf. on Solid State Lighting (Int.
методам анализа качества и безопасности пищевых
Soc. for Optics and Photonics, 2004), pp. 165-173.
продуктов (Брандес - Медицина, М., 1998).
9. D. T. Nhut, T. Takamura, H. Watanabe, et al., Plant
30.
Е. В. Вязов и Н. В. Шалыго, Докл. Нац. Академии
Cell, Tissue and Organ Culture 73, 43 (2003).
наук Беларуси 59 (2), 87 (2015).
БИОФИЗИКА том 65
№ 1
2020
124
КУЛЕШОВА и др.
The Influence of Spectral Properties of Lighting Environments on Light Absorption
by Lettuce Leaves and Net Photosynthesis of Lettuce
T.E. Kuleshova*, I.N. Chernousov**, O.R. Udalova**, L.M. Anikina**, Yu.V. Khomyakov**,
A.V. Aleksandrov**, I.S. Seredin***, S.V. Feofanov***, S.A. Shcheglov****,
N.R. Gall*, and G.G. Panova**
*Ioffe Physical-Technical Institute, Polytekhnicheskaya ul. 26, St. Petersburg, 194021 Russia
**Agrophysical Research Institute, Grazhdanskiy prosp. 14, St. Petersburg, 195220 Russia
***LLC “O2 Lighting Systems”, ul. Novoroshchinskaya 4, St. Petersburg, 196084 Russia
****Saint Petersburg National Research University of Information Technologies, Mechanics and Optics,
Kronverkskiy prosp. 49, St. Petersburg, 197101 Russia
In the current study, we investigated the effect of light with different spectral composition on lettuce leaf light
absorption and found optimal light conditions to enhance the yield and quality of plant species grown under ar-
tificial light. “Typhoon” lettuce exposed to five different light intensities was grown by the method of small-vol-
ume panoponics in an automated greenhouse. The method developed by researchers of this study was employed
to estimate in vivo light absorption in lettuce leaves. The use of this method does not cause an injury to the plant.
Based on data obtained, we conclude that spectral properties of lighting environments can influence leaf optical
traits reflecting the physiological status of plants. When the plant productivity was maximum, the biomass of let-
tuce increased by 1 g for 10 days with the percentage photon of light absorbed up to ~140 μmol · m-2 · s-1, the
plant had worse growth characteristics under light intensity of below 70 μmol · m-2 · s-1. Because light spectral
characteristics of high-pressure sodium arc lamp and LED producing yellow light were similar in the range of
photosynthetic active radiation and photosynthetic active photon flux density was almost equal in the blue,
green, and red spectral regions, plants had similar light absorption during the growth cycle. At the same time,
there is significant increase in yield of lettuce (~50%), as well as better plant growth, development and biochem-
ical composition under HPS lamps. Differences observed under different artificial light showed that HPS lamp
produced a light spectrum which was more suitable for photosynthesis. The data obtained indicate that the light-
ing spectrum emitting ~25 μmol · m- 2 · s-1 in the range of 400-500 nm, ~150 μmol · m-2 · s-1 in 500-600 nm,
~150 μmol · m-2 · s-1 at 600-700 nm leads to high lettuce productivity.
Keywords: intensive photoculture, lighting spectra, LED, leaf optical properties, light absorption, net photosyn-
thesis
БИОФИЗИКА том 65
№ 1
2020