БИОФИЗИКА, 2020, том 65, № 1, с. 138-151
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 595.76,51.76,532.22
НЕПРЕРЫВНАЯ МОДЕЛЬ ДЛЯ ОСЦИЛЛИРУЮЩЕЙ ВСПЫШКИ
ЧИСЛЕННОСТИ ЧЕШУЕКРЫЛОГО ФИТОФАГА Malacosoma disstria
(Lepidoptera, Lasiocampidae)
© 2020 г. А.Ю. Пеpеваpюxа
Санкт-Петербургский институт информатики и автоматизации PАН,
199178, Санкт-Петербург, 14-я линия, 39
E-mail: temp_elf@mail.ru
Поступила в редакцию 27.05.2019 г.
После доработки 17.11.2019 г.
Принята к публикации 22.11.2019 г.
Вспышки численности отдельных популяций - важные во многих аспектах и многообразные с
точки зрения теории динамических систем явления. Наиболее стремительно развиваются вспышки
насекомых, имеющие длительные последствия для лесного хозяйства. Подобные события
относятся к экстремальным несбалансированным переходным процессам. Механизмы запуска и
фазы затухания различаются у разных таксономических групп вредителей. Отличны по
продолжительности и фазе наступления повторной активности вспышки у псиллид и у бабочек,
поражающих лиственные или хвойные насаждения в одном регионе. Их особенности динамики
актуальная задача для вычислительного моделирования. Для математического описания помимо
порогового варианта запуска вспышки интересно модифицировать непрерывные вычислительные
модели для анализа колебательной динамики. В статье рассмотрено моделирование специфичного
сценария вспышечной активности спонтанно затухающего осциллирующего характера в
непрерывной модели с запаздывающей регуляцией и нелинейным противодействием co стороны
биотического окружения. Описанный феноменологическим уравнением сценарий из серии
разновеликих максимумов и итоговым затуханием пиков у балансового равновесия реализуется для
бабочки-вредителя коконопряда Malacosoma disstria, поражающей лиственные массивы в Северной
Америке и вызывающей масштабную дефолиацию леса. Новый сценарий качественно отличается
от модели порогового развития и завершения вспышек у псиллид в Австралии.
Ключевые слова: экстремальная динамика экосистем, вспышки насекомых, затухающие флуктуации
численности, модели колебательной динамики популяций, уравнения с запаздыванием, бифуркации,
коконопряд Malacosoma disstria, дефолиация леса.
DOI: 10.31857/S0006302920010160
Вспышки численности одна из хрестоматий-
Случаи стремительного размножения необхо-
ных проблем для экологии биосистем и матема-
димо изучать, классифицировать и по возможно-
тической биологии. Актуальность изучения
сти прогнозировать подобные ситуации. Вспыш-
вспышек как группы экстремальных явлений
ки наиболее характерны для насекомых-фитофа-
гов. Наблюдаются случаи взрывного роста
только возрастает в современных условиях «пере-
численности не только у инвазивных, но в том
мешивания» фаунистических комплексов. Глоба-
числе у ряда автохтонных видов. Подобные ло-
лизация сопровождается все более частыми слу-
кальные явления с фазой бурного роста у популя-
чаями запуска агрессивных инвазионных процес-
ций, издавна включенных в состав сообществ,
сов [1] после нежелательных вселений и быстрого
под пристальным математическим взглядом вы-
размножения чужеродных видов. Иногда вселе-
глядят несхожими нелинейными процессами с
нец адаптируется многие годы до перехода к ак-
несколькими разнородными типами трансфор-
тивности (как интродуцент краб Paralithodes
маций фазового портрета.
camtschaticus в Баренцевом море [2], моллюск
Продолжаются исследования причин вспы-
Dreissena polymorpha в озерах США). Катастрофи-
шек насекомых, эффекторах/ингибиторах вне-
ческая для лесной растительности вспышка вре-
запной активности вредителей. Дискуссия ведет-
дителя может стартовать немедленно, как было в
ся о превалировании биотических [4] либо внеш-
2014 г. с проникшей самшитовой огневкой Cуdali-
них факторов, например циклах солнечной
ma perspectalis в Имеретинской долине [3].
активности и периодических климатических воз-
138
НЕПРЕРЫВНАЯ МОДЕЛЬ ДЛЯ ОСЦИЛЛИРУЮЩЕЙ ВСПЫШКИ
139
действиях на ключевые регуляторные механиз-
описания конкретных изменений и переходов
мы. В литературе систематически описан ряд
между режимами существования биосистем.
частных сценариев, как, например, в работе [5]
Интересным примером для интерпретации
для бабочки Dendrolimus sibiricus в засушливой
свойств новой математической модели является
Внутренней Монголии. По-видимому, главен-
агрессивное поведение вредителей в лесах Север-
ствующей единой причины для явлений не суще-
ной Америки [8], например кольчатого коконо-
ствует. Мы обратим вниманием именно на вари-
пряда Malacosoma disstria. Выбор в качестве эко-
ативность подобных экстремальных явлений да-
логического примера для построения модели
же у одного вида, но в разных ареалах. В
мотивирован наличием опубликованных долго-
некоторых случаях удается отметить пороговый
временных систематизированных наблюдений
путь развития и окончания краткой вспышки. В
лесного хозяйства в провинциях Квебек и Онта-
других интерпретировать поведение в рамках би-
рио за проявлениями активности вредителей. Од-
фуркационных изменений у осциллирующей ди-
на из актуальных задач для математического ис-
намики. Разграничение с точки зрения теории
толкования резких изменений у биологического
динамических систем типов наблюдаемых пере-
процесса состоит в обоснованном выборе подхо-
ходов между фазами процесса, связанных с уско-
дящей методики из конечного набора возможных
ренным размножением, может помочь система-
динамических систем.
тике экстремальных экологических явлений.
Для каждой разновидности нетривиальной
популяционной динамики целесообразно подби-
Участие автора в проекте совместно с группой
рать и настраивать оригинальный математиче-
энтомологов из Всероссийского научно-исследо-
ский аппарат, добиваясь принципиального каче-
вательского института защиты растений по ана-
ственного правдоподобия изменений. Рассуж-
лизу характеристик динамики смежных поколе-
дать о преимуществах какого-то подхода для
ний локальной популяции кукурузного мотылька
всего популяционного моделирования нет смыс-
Ostrinia nubilalis [6] привело к выводу, что иногда
ла, процессы в биосистемах имеют разные вре-
мы наблюдаем не некоторое выделенное событие
менные параметры. В работе [9] авторы предлага-
с типичными признаками вспышки. Проявляют-
ли представлять вспышки массового размноже-
ся иррегулярные переходы между длительными
ния насекомых как фазовые переходы второго
периодами депрессии популяции мотылька и ее
рода, а в работе [10] строили модель со спектраль-
высокой репродуктивной активности. В частном
ным анализом временных рядов. Важно пони-
случае оба состояния популяции отражаются ко-
мать, что дискретные, гибридные или непрерыв-
лебательной динамикой, но с разной амплитудой
ные динамические системы обладают отличаю-
внутри популяционного цикла.
щимися асимптотическими множествами
Вспышки численности эпизодичны как осо-
траектории, набором их трансформаций (бифур-
бые переходные режимы функционирования
каций) и свойств у границ областей притяжения
экосистем, и при моделировании развития ситуа-
аттракторов. Для интересующей задачи выберем
ции необходимо включать варианты их заверше-
непрерывную запись феноменологической моде-
ния. Известная модель в работе [7] - уравнение
ли - форму дифференциальных уравнений с от-
для вспышек насекомых - не покажет сценария
клоняющимся аргументом, отличающуюся регу-
спонтанного завершения явления, там требуется
ляционным и адаптационным запаздыванием.
внешний импульс для переходов между устойчи-
выми состояниями, таких как занос насекомых
ВАРИАТИВНОСТЬ ПОРОГОВОГО
ветром из других ареалов. Фазы экстремальных
РАЗВИТИЯ ВСПЫШКИ У ПСИЛЛИД
состояний численности для автохтонных и для
инвазивных видов отличаются нюансами дина-
Рассмотрим наглядно вариативность экстре-
мики ситуаций. В настоящей работе мы обсудим
мальных изменений численности. В предыдущей
модель специфической формы вспышечной ак-
работе [11] мы предложили метод вычислитель-
тивности у насекомых - осцилляционного харак-
ного моделирования порогового сценария еди-
тера. Автохтонная популяция продуцирует дли-
ничной локальной вспышки на основе анализа
тельную серию пиков. В таком режиме популя-
регуляторов размножения монофага Cardiaspina
ция все время между пиками существует при
albitextura из семейства Psyllidae. Развитие фаз ти-
достаточно высокой численности. Рассмотрим
пичной вспышки автохтонного вида псиллид в
моделирование явления серии разновеликих пи-
вечнозеленом эвкалиптовом лесу на восточном
ков как процесса при сложной регуляции с после-
побережье Австралии было описано в работе [12].
действием. Ранее мы использовали для математи-
Объяснимо, когда чужеродная дальневосточная
ческого описания пороговой формы вспышки в
бабочка, оснащенная химзащитой токсинами ге-
вечнозеленом лесу итерационную гибридную си-
молимфы гусениц от потенциальных местных
стемы, предложив специальный триггерный
врагов, способна достигать уровня численности,
функционал для описания ее завершения. Каж-
уничтожающей самшитовый лес. Однако вспыш-
дый формализм имеет свои возможности для
ки случаются у очень уязвимых мелких поливоль-
БИОФИЗИКА том 65
№ 1
2020
140
ПЕPЕВАPЮXА
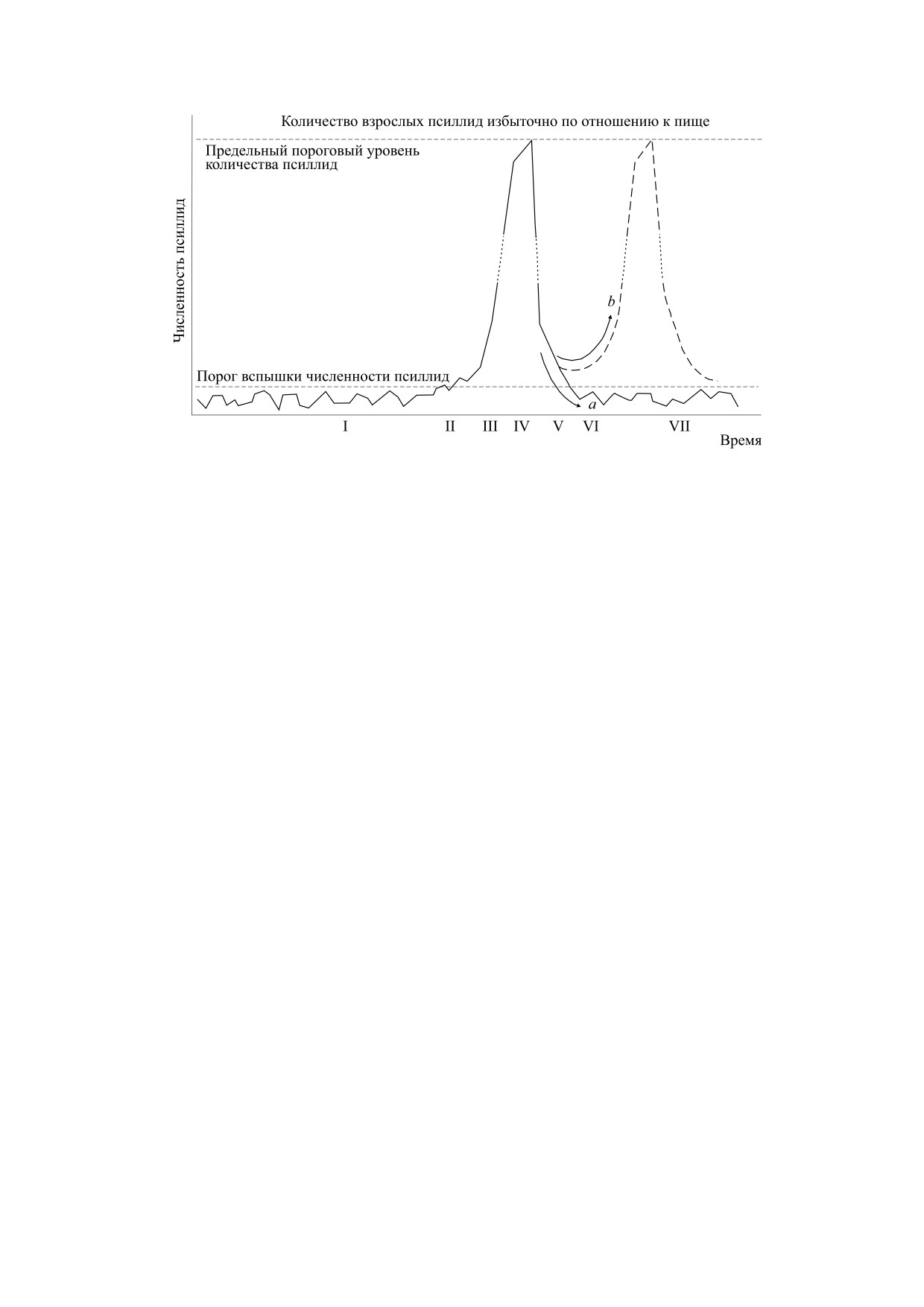
Рис. 1. Пороговое развитие вспышки эвкалиптовых псиллид и смена фаз (согласно работе [11]).
тинных фитофагов отряда Hemiptera - кокцид и
ресурсов - пригодных деревьев. Тут обычно
псиллид. По числу продуцируемых поколений за
службы учета фиксируют вспышку по масштабам
сезон активности насекомые (в своих локальных
дефолиации лесных массивов. Дефолиация при
ареалах) разделяются на поли-, би- и моноволь-
поражении растений полужесткокрылыми с ро-
тинных, дающих единственное поколение в году.
товым аппаратом колюще-сосущего типа насту-
пает в большей мере из-за развития вторичных
Ситуация взрывообразного популяционного
грибковых инфекций, развивающихся в их лип-
процесса у этих полужесткокрылых попала в ка-
ких выделениях, значит без необходимости пол-
честве иллюстративного примера в известную
ного уничтожения биомассы растительности не-
книгу Ю. Одума [13] без многих необходимых по-
посредственно вредителем для завершения явле-
яснений, как хрестоматийный образец развития
ния. В фазе V резко ухудшаются условия для
вспышек, который можно обобщить на другие
откладки яиц и выживаемости нимф. Процесс
примеры. В интерпретации у данного автора от-
локализован. После столь же резкого сокращения
сутствовала немаловажная на самом деле деталь.
численности до минимума в фазе VI малочислен-
В исходном описании [12] был предусмотрен ре-
ная популяция псиллид переходит в длительный
же встречавшийся сценарий b на графике популя-
режим хаотических флуктуаций. Самое интерес-
ционного процесса псиллид (см. рис. 1). Отмеча-
ное в сценарии - механизм образования порога
лось, что в некоторых специфических случаях по-
численности. Обычно размножение этих фитофа-
сле затухания вдруг наблюдался относительно
гов контролируется двумя-тремя видами мелких
скорый повторный пик численности. Вариант
паразитических ос из семейства Encyrtidae [14].
развития b связывался с замедленным перемеще-
нием псиллид в районах с менее плотным произ-
Предложен подход в форме гибридной дина-
растанием пригодных для их питания эвкалип-
мической системы. Идея строится на формализа-
тов. Степень поражающего растительность воз-
ции роли этапности в жизни исходного поколе-
действия от размножения этих насекомых
ния насекомых - убывающей численности на
обусловлена вторичными инфекциями.
дробном интервале его существования t ∈ [0,T].
Изменения уравнения непрерывной системы в
В концептуальном сценарии, представленном
выделенные условиями моменты времени были
на рис. 1, в фазе флуктуаций I при малой средней
соотнесены с переходами между тремя стадиями
численности, которую мы математически интер-
развития онтогенеза Psyllidae: яйца, нимфы, има-
претировали апериодическим режимом хаотиза-
го. Для каждой из стадий учтено, что отличаются
ции, использован другой хаотический переход-
факторы смертности, как зависящей от плотно-
ный режим, чем в известном сценарии появления
сти скопления псиллид, так и независимой от те-
хаоса Фейгенбаума. После непредсказуемого
кущей численности.
преодоления порога процесс переходит в эруп-
тивную фазу III. Взрывообразный рост популя-
Использующая непрерывно-дискретное вре-
ции в фазе III останавливается на пике IV в мо-
мя модель сформирована из дифференциального
мент исчерпания необходимых для размножения
уравнения убыли начальной численности N(0)
БИОФИЗИКА том 65
№ 1
2020
НЕПРЕРЫВНАЯ МОДЕЛЬ ДЛЯ ОСЦИЛЛИРУЮЩЕЙ ВСПЫШКИ
141
поколения с переопределяемой правой частью,
определения момента остановки расчетов и пере-
что происходит предикативно на трех последова-
хода к следующей стадии непосредственно из
тельных временных субинтервалах. Алгоритми-
времени для первой стадии или из расчета вспо-
чески реализуется модель в форме гибридного ав-
могательного показателя развития для второй
томата. Набор условий включен в модель для
стадии:
⎧−(α
1
wt)N t)+Ψ[N(0)]β)N t),
t
<ρ
dN
⎪
=
⎨-
2
N
(α ρΘ
N(ρ)] /
w(ρ)
+β)N t),
t
>ρ,
wt)
<
w
2
(1)
dt
⎩-α
3
wt)N t)N(t
-ζ),
t
<
T,
где ρ - фиксированная длительность первой ста-
Аспект вариативности на стадии завершения
дии, когда плотность яиц становится важным ре-
вспышки не был анализирован нами в предыду-
гулирующим фактором выживаемости, именно
щей публикации. В работе [11] мы показали сце-
плотность скопления доступных жертв усиливает
нарий с одним продолжительным пиком. Рас-
реакцию паразитов. При t > ρ расчет уравнения
смотрим модификацию сценария с двумя пиками
переходит на нимф, основную вредящую стадию
для псиллид, чтобы показать гибкость предло-
насекомых, где достигнутый показатель развития
женного метода с использованием дискретной
w(ρ) уменьшает темп убыли. Расчет длительности
составляющей траектории.
стадии продолжается с использованием вычисле-
ния вспомогательной величины w(t) до достиже-
Вариативность можно отразить без принципи-
ния переходного уровня w2.
альной модификации в предикативной структуре
Гибридная система (1) используется для вы-
и чрезвычайно трудоемкой перенастройки пара-
числения итогового N(T). Используя гибридную
метров (1), которая использует функции триггер-
вычислительную структуру, мы получили слож-
ного действия: Ψ,Θ. Можно вносить коррективы
ную зависимость N(T) = ϕ(λS) с двумя экстрему-
в эти функции, целенаправленно меняя их вклад
мами для дискретной составляющей траектории в
в расчет выживаемости на нужной стадии разви-
форме итераций, считая S = N(T): Sn+1 = ϕ(Sn),
тия. Функция Ψ для формы правой части
Nn+1(0) = λSn, где λ - средняя популяционная
на субинтервале t < ρ «ответственна» за учет эф-
плодовитость. Исследуем точки дискретной ите-
фекта Олли и существование критической низ-
рации S0,S1...Sn из расчета гибридных систем,
кой численности L при ухудшении эффективно-
связанных по начальным условиям на границах
сти воспроизводства при малочисленном состоя-
смежных непрерывных интервалов у дифферен-
нии «бутылочного горлышка», когда возникает
циальных уравнений. Итерации неунимодальной
трудность с нахождением пар для размножения.
функциональной зависимости демонстрируют
Мы можем говорить о данном эффекте для
спонтанное преодоление порогового равновесия
малой группы [15] применительно к эффективно-
S3* из переходного хаотического режима, так как
сти размножения у вредителя эвкалипта: для осо-
бей видов семейства Psyllidae партеногенез не ха-
max ϕ(λS) лишь немного превосходит ϕ(S3*).
рактерен, в отличие от других надсемейств He-
Длительность переходного хаоса до момента
miptera
- тлей Aphidoidea или червецов
ϕ(Sn) > ϕ(S3*) + ε зависит от выбора начальной
Coccoidea. Для большой численности N(0) функ-
точки S0.
ция единична и не влияет на результат расчетов:
Ψ (N(0)) = 1 + exp(-σ1N2(0)), limS→∞Ψ (N(0)) = 1, Ψ(0) = 2.
1a
Вторая функция-триггер Θ определяет порог исчерпания ресурсов для стадии нимф:
N(τ)
eс1
1
(N
Θ ρ))
=
1+
, lim
(N
Θ ρ))
=
1+
1б
с
1
N(τ)
N(τ)→∞
l
+c e
c
2
2
Действие Θ можем сдвигать в интервале вре-
численности после пика численности. Если мак-
мени относительно фиксированного момента пе-
симум не был затяжным, то вероятность повтор-
рехода ко второй стадии Θ(N(ρ - ζ)), усиливая
ной вспышки увеличится. На рис. 2 представлен
(или ослабляя в случае Θ(N(ρ + ζ))) ее роль и тем
альтернативный вычислительный эксперимент
самым меняя резкость перехода к фазе V падения
cо сценарием (1b) и с генерацией повторной
БИОФИЗИКА том 65
№ 1
2020
142
ПЕPЕВАPЮXА
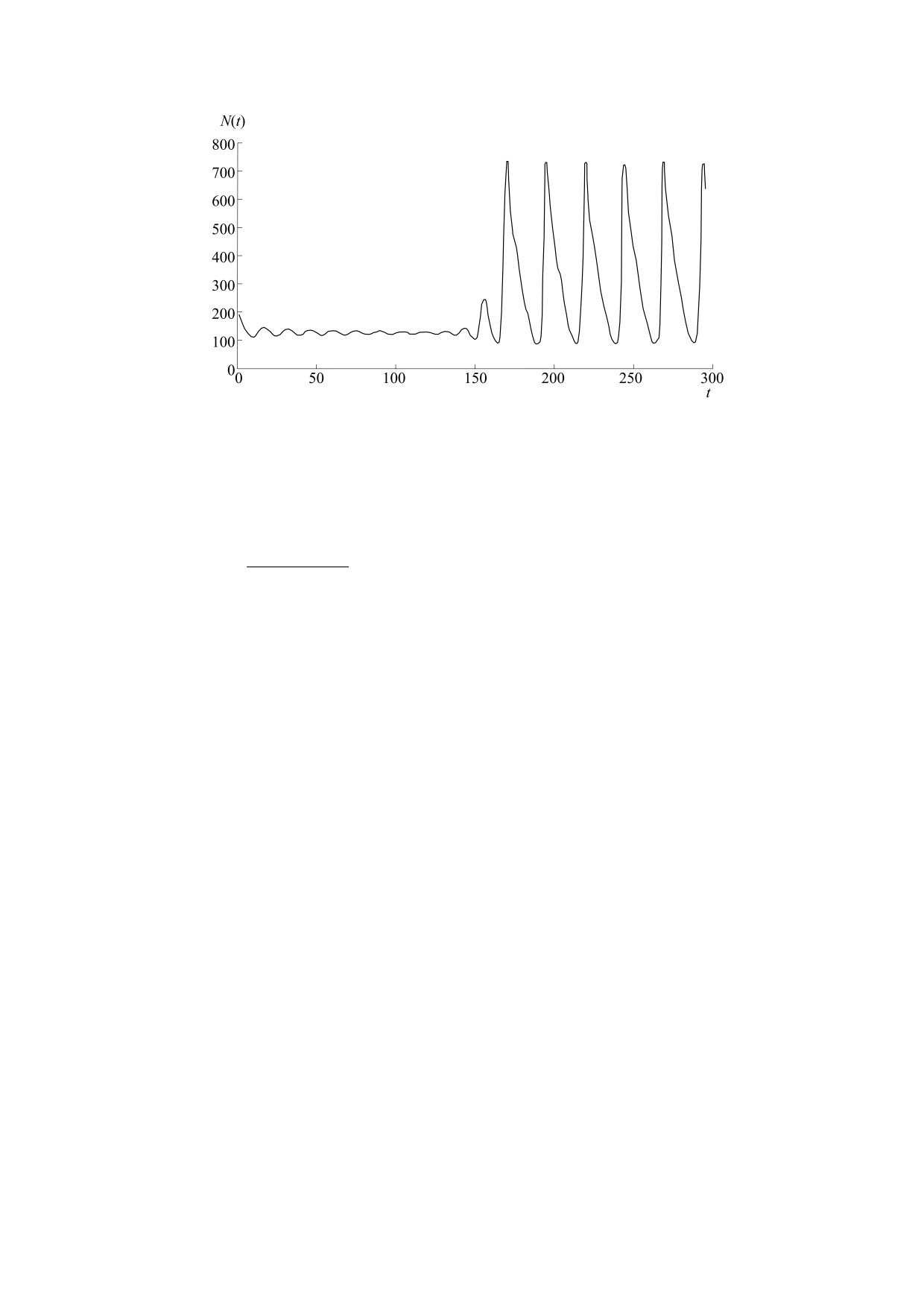
Рис. 2. Моделирование варианта повторной вспышки вредителя с краткими пиками.
вспышки через промежуток гибридного времени
шениями различных видов паразитических ос
и нерегулярных колебаний.
при выборе предпочтительного объекта для атак.
После эпизодического повышения численности
Модификация опишет вариант скорого по-
возрастает роль Coccidoctonus psyllae и других пе-
вторного преодоления порога. Обе вспышки про-
репончатокрылых ос подсемейства Encyrtinae
исходят в течение одного года, пик первой прихо-
[16]. Эти микроскопические осы - гиперпарази-
дится на август, второй - на ноябрь. Тут мы ис-
ты. Они активно начнут паразитировать на ли-
пользуем переходную хаотическую динамику,
чинках двух видов других размножившихся пара-
имитируя природную случайность. Переходный
зитических наездников из рода Psyllaephagus. В
хаотический режим из-за фрактальных границ
современной работе [17] подчеркивается завися-
областей притяжения воспроизводит естествен-
щая от плотности жертв активность Coccidoctonus
ную стохастичность среды для флуктуаций
psyllae. Так, у первичных паразитов, как основных
численности в полностью детерминированной
биологических врагов, уничтожающих яйца
модели. Для описания завершения вспышки бы-
псиллид, создается верхний порог достижимой
ла использована редуцирующая устойчивую ста-
численности.
ционарную точку обратная касательная бифурка-
ция. В рассмотренном примере бифуркация типа
В контексте проблемы с очаговой дефолиаци-
«backward tangent bifurcation» резко переводит по-
ей эвкалиптовых лесов вредоносным инвазион-
пуляцию в следующий интервал прохождения ха-
ным видом насекомых оказался именно гиперпа-
отических флуктуаций. Порог запуска вспышки
разит Coccidoctonus psyllae. После появления пара-
не может быть монотонно достижим из любого
зитической осы в Новой Зеландии в местных
состояния системы, все же вспышка численности
лесах участились вспышки фитофагов из большо-
с дефолиацией эвкалиптов эпизодическое явле-
го семейства Psyllidae [18]. Видовое разнообразие
ние, потому несвязные границы областей притя-
паразитических перепончатокрылых подотряда
жения двух точек-аттракторов в нашей базовой
Apocrita велико и их систематика еще не полно-
непрерывно-дискретной модели (1) удачно опи-
стью описана. В семействах Encyrtidae и Ichneu-
сали данный экологический аспект.
monidae встречаются сверхпаразиты даже третье-
го порядка. Осы находят личинки уже заражен-
Популяция в сценарии перед преодолением
ных другими осами фитофагов и поражают на
порога может не достигать на промежутке време-
ранних стадиях развития особи других перепон-
ни отчетливо наблюдаемых предпороговых зна-
чатокрылых паразитов. В Австралии встречается
чений, как это было в работе [11], и новый
гетерономный гиперпаразитоид рода Psyllaepha-
модельный сценарий менее доступен для прогно-
gus, у которого развитие из личинок самок и сам-
зирования признаков начала вспышки. Суще-
цов зависит от выбора хозяина: псиллиды или
ствование порога, отраженного в модели погра-
другого Psyllaephagus. Таксоны отряда Hymenop-
ничным и неустойчивым S3* (имеющим слева и
tera - одни из наименее изученных среди насе-
справа от себя только другие неустойчивые ста-
комых.
ционарные точки) положением равновесия
Нами разрабатывалась феноменологическая
ϕ(S3*) = S3*, объясняется сложными взаимоотно-
гибридная модель популяционного процесса, так
БИОФИЗИКА том 65
№ 1
2020
НЕПРЕРЫВНАЯ МОДЕЛЬ ДЛЯ ОСЦИЛЛИРУЮЩЕЙ ВСПЫШКИ
143
как вряд ли возможно описать традиционными
ним повторением. В работе [21] вспышки насеко-
методами математической биологии в системе
мых в Канаде рассмотрены как периодические
уравнений нетривиальное и недостаточно иссле-
возмущения лесных экосистем. Отметим, что в
дованное взаимодействие мелких видов субмил-
энтомологической литературе термин «циклич-
лиметровых ос-паразитов. Экодинамика тут
ность» используется гораздо либеральнее, чем в
принципиально отлична от системы «хищник-
математической, просто для указания повторяе-
жертва». Так, большая убыль личинок хозяев в за-
мости явлений. Если быть точным, то нужно го-
вершении вспышки ведет к гибели и личинок их
ворить о квазипериодичности (phase-forgetting
паразитов. В настоящее время для описания
quasi-cycle). После долгого состояния при сред-
вспышечных явлений осваиваются такие пер-
ней численности насекомых, не вызывающей по-
спективные методы искусственного интеллекта,
вреждения леса и вполне равновесного с биотиче-
как мультиагентные системы и нечеткие клеточ-
ским окружением, вдруг появляется серия очень
ные автоматы [19].
многочисленных несмежных поколений. В от-
дельных ситуациях в динамике численности
У представителей других отрядов насекомых и
бабочек авторы говорят о «пилообразных осцил-
в различных климатических зонах случаи массо-
ляциях» (sawtooth-like oscillations), подчеркивая
вого размножения развиваются с отличительны-
нерегулярность явления. В состоянии пиков чис-
ми особенностями. Итерационная модель в фор-
ленность много превышает среднемноголетнюю
ме xn+1 = f(xn) - Qxn для бивольтинных насеко-
за предшествующую декаду, в очаге гибнет лес и
мых столкнется с проблемой, что в бореальных
фиксируется локальная вспышка. Расстояния
условиях смежные поколения развиваются в раз-
между пиками пульсирующей активности север-
ных условиях. Одно поколение зимующее, другое
ных моновольтинных бабочек составляют не два
развивается в комфортных летних условиях, по-
месяца, как у эвкалиптовых псиллид в Австралии,
тому о единой зависимости ϕ(N(0)), N(0) > L го-
а годы.
ворить нельзя. Интервал онтогенеза t ∈ [0,T] у
двух смежных поколений бабочек, очевидно, раз-
Характерна длительная осциллятивная дина-
ный, у зимующего поколения почти в два раза
мика, но с разными характеристиками колеба-
больше. Зимуют лесные бабочки чаще на стадии
тельной активности, для опасных вредителей, ко-
яйца, которая у псиллид коротка.
торые не являются конкурирующими. Несин-
хронные иррегулярные колебания активности
отмечены в работе [22] и наблюдаются у бабочки-
ОСОБЕННОСТИ КАНАДСКОГО
листовертки Choristoneura fumiferana, поражаю-
ПУЛЬСИРУЮЩЕГО СЦЕНАРИЯ ВСПЫШКИ
щей хвойные леса в восточных провинциях Кана-
ды и севере США, и особенно у кольчатого коко-
Рассмотрим иные ситуации вспышек вредите-
нопряда Malacosoma disstria, вредящего листвен-
лей - двух видов бабочек в бореальных лесах и
ным деревьям в этих регионах.
покажем модель качественного описания подоб-
ных явлений. Регуляция эффективности воспро-
Вспышка оценивается по площадям поражен-
изводства насекомых фитофагов посредством ос
ного леса, учет которых в канадском лесном хо-
гиперпаразитов очень интересная для моделиро-
зяйства многие десятилетия ведется для каждой
вания, но специфическая ситуация. Такая колли-
провинции. Из сведений на графиках отчетности
зия у паразитов из подсемейств Encyrtidae част-
оказывается, что вспышки вредителей в регионах
ная и актуальна для лесов с относительно посто-
Северной Америки проходят различным образом.
янными сезонными условиями и богатым
На рис. 3 показаны графики масштабов пораже-
видовым разнообразием энтомофауны. Суще-
ния леса от активности коконопряда в провинци-
ствование хаотического режима со спонтанным
ях Онтарио и Квебек, где с математической точки
пороговым переходом к вспышке не может быть
зрения различен характер возникшей колебатель-
главенствующим сценарием механизма запуска
ной динамики [23].
вспышки там, где разнообразие насекомых не
Для Онтарио видим релаксационный цикл из
столь велико, а сезон вегетации существенно ко-
серии пиков возрастающей амплитуды, который
роче.
будет устанавливаться после общей формы би-
Вполне логично, что в бореальных лесах Се-
фуркации Андронова-Хопфа. Для проблем моде-
верной Америки действуют некоторые другие ме-
лирования более интересен сценарий вспышек в
ханизмы, вызывающие не менее масштабные
лесах Квебека с затухающими колебаниями, для
вспышки у таксономических групп отряда Lepi-
которого интересно предложить новую модель.
doptera [20], генерирующих единственное поко-
Учет и оценка состояния популяций насеко-
ление за сезон.
мого во время вспышек бабочек проводится кос-
В специфических и важных случаях вспышка
венным образом по площади пораженного леса,
численности насекомых выходит за рамки сцена-
покраснению и опадению хвои. Для обоснования
рия с одиночным пиком на графике динамики
феноменологической модели смены фаз пульси-
поражения лесов и даже не ограничивается од-
рующей вспышки достаточно сведений о пораже-
БИОФИЗИКА том 65
№ 1
2020
144
ПЕPЕВАPЮXА
Рис. 3. Динамика осцилляционной вспышки коконопряда Malacosoma disstria в Онтарио и Квебеке в масштабах гибели
леса (согласно работе [23]).
нии леса, но на не менее чем полувековом интер-
верной Америке. Выраженная пилообразная со-
вале времени. Вся вспышка с отдельно стоящими
ставляющая колебаний отмечена и для листо-
пиками колебания, которые демонстрируют
вертки и характерна для Восточной Канады, но
тренд затухания амплитуды, может растягиваться
не для лесов в США [26], там пульсирующая
на 30 лет. На рис. 4 показано развитие до 1991 г.
вспышка не начинается, как в провинции Нью-
запущенной в 1955 г. вспышки у листовертки в
Брансуик, а заканчивается наибольшим пиком,
провинции Нью-Брансуик согласно отчету о со-
но это уже другой сценарий, требующей своей
стоянии экосистем и тенденций в рамках выпол-
частной модели. Интересно, что для опасного ин-
нения Федеральным правительством Канады
вазивного вредителя Lymantria dispar в новом аре-
обязательств Конвенции о биологическом разно-
але штата Висконсин подходит пороговый сцена-
образии [24]. Публикуется отчетная документа-
рий вспышки, но в родном европейском - демп-
фированный осцилляционный [27].
Мы видим моментальный стремительный пе-
Столь интересное явление привлекало специ-
реход к вспышке без околопороговых промежу-
алистов в области математической биологии.
точных значений. Образуются три отчетливых от-
Предлагались модели вспышки с точки зрения
стоящих пика, переходящих в состояние, когда
формализма теории катастроф [28]. Описание
численность бабочки больше не оказывает столь
резкого перехода к состоянию вспышки от мини-
разрушительного (или обновительного) воздей-
мального уровня входит в перечень классических
ствия на экосистему. Воздействие исчисляется в
задач прикладного применения концепций тео-
миллионах гектаров. Максимальные значения
рии катастроф (для перехода использовали «Cusp
численности тут не являются самостоятельными
catastrophe» - катастрофу «сборки»). Методы тео-
явлениями вспышек, как в случае Cardiaspina albi-
рии катастроф развивались из результатов топо-
textura, но составляют часть долгого колебатель-
логии с основными представлениями «резиновой
ного процесса в локальной популяционной дина-
геометрии» о деформациях и проекциях искрив-
мике насекомых. Оба примера ситуаций вспыш-
лений трехмерных поверхностей. «Катастрофи-
ки затухают как волны и рябь после броска камня
ческое» описание в работе [29] носит наглядный,
в воду. О том, что представляет собой образный
но качественно геометрический характер, не учи-
«камень», вызывающий волны, есть несколько
тывающий череду фаз процесса и переходные ре-
гипотез [25]. Сценарий схож с динамикой реци-
жимы.
дивирующей инфекции, когда медленно включа-
ющийся адаптивный иммунный ответ не спосо-
бен подавить хронический очаг возбудителя.
ФОРМЫ ЗАПАЗДЫВАЮЩЕЙ РЕГУЛЯЦИИ
В МОДЕЛЯХ ПОПУЛЯЦИЙ
Качественный характер особенностей мест-
ной вспышки популяции не сохраняется по всему
Пилообразную вспышку необходимо рассмат-
огромному ареалу Choristoneura fumiferana в Се-
ривать как отдельный сценарий взрывообразного
БИОФИЗИКА том 65
№ 1
2020
НЕПРЕРЫВНАЯ МОДЕЛЬ ДЛЯ ОСЦИЛЛИРУЮЩЕЙ ВСПЫШКИ
145
развития популяционного процесса. С точки зре-
ния теории динамических систем ситуация не
может быть разновидностью рассмотренного ра-
нее порогового варианта модели запуска вспыш-
ки из временного хаотического режима. Методы
стационарных точек c возможными триггерными
воздействиями у дискретных итераций, развитые
в предыдущей работе и усовершенствованные на-
ми выше, подчиняются теоремам Шарковского и
Гукенхеймера, потому не очень применимы для
столь своеобразных перемен в лесах Канады, ко-
торые можно назвать самозатухающим релакса-
ционным осциллятором. Необходимо обосновать
другой вычислительный аппарат для описания и
сравнения вариантов развития сценариев, где ос-
новная сложность - необходимость демпфирую-
щихся бифуркационных изменений, соответ-
ствующих перемежающимся фазам пиковой ак-
тивности и депрессии у вредителей в нашей
Рис. 4. Динамика пилообразной вспышки с затуханием
задаче.
осцилляций y листовертки в Нью-Брансуик в масшта-
бах дефолиации леса (в млн га) из отчета ESTR [24].
Так как ситуация имеет существенно больший
временной масштаб развития, чем сжатая до се-
ми-девяти итераций фаза вспышки псиллид, ло-
ляется ключевым понятием моделей данного
гично воспользоваться для подобной задачи
типа.
представлениями о запаздывающем действии в
регуляции составляющих факторов популяцион-
Для совсем малых значений запаздывания τ и
ного процесса (и не только репродуктивного
небольших r поведение решения (2) идентично
цикла).
модели Ферхюльста. До значений rτ < π/2 реше-
ние продемонстрирует затухающие колебания с
Известно из ряда экспериментов, например,
. При переходе значения произведения
N(t) → K
из серии опытов Николсона с лабораторной по-
параметров через критическое rτ < π/2 происхо-
пуляцией мухи Lucilia cuprina при различном по-
дит возникновение бифуркации Андронова-
ступлении корма, что колебания с большой ам-
Хопфа, т. е. особая точка становится неустойчи-
плитудой могут возникать в случае изолирован-
вым фокусом и от нее отделяется замкнутая тра-
ной популяции насекомых в ограниченном
ектория - типичное проявление устойчивого
пространстве и при постоянных поддерживаемых
предельного цикла траектории [32]. Функцию-
условиях [30]. Помимо известных моделей попу-
предысторию на интервале [-τ, 0] тут и далее счи-
ляции (модель Ферхюльста, модель Гомпертца)
таем тождественной единице.
для монотонного роста численности к асимпто-
тическому состоянию K балансового равновесия
Бифуркацией в современном анализе динами-
со средой N(t) → K и с точкой перегиба макси-
ческих систем называют любое качественное из-
мальной скорости роста
N ''(t ) = 0
предложены
менение поведения - метаморфоз фазового порт-
модели для объяснения колебательной динамики
рета. Редукция устойчивой точки или возникно-
в одновидовом случае.
вение пары неустойчивых стационарных точек
это тоже типы бифуркаций, которые фиксируют-
В работе [31] была предложена известная мо-
ся в пространстве параметров модели. Не все би-
дификация уравнения Ферхюльста с включением
фуркации имеют истолкование в биологии по-
запаздывания τ для действия плотностной само-
пуляций. Появление орбитально устойчивой
регуляции численности:
циклической траектории у непрерывной динами-
ческой системы - наиболее часто встречающееся
dN
N(t
-τ)
=
rN t)
1−
,
(2)
и изученное бифуркационное изменение. Иссле-
(
)
dt
K
дованию модели (2) посвящена обширная мате-
матическая литература, но нас интересуют биоло-
где K отражает уровень равновесия со средой для
гические недостатки уравнения, от которых мы
популяции с репродуктивным потенциалом r.
оттолкнемся для построения новых модифи-
Введенное в (2) запаздывание τ является агреги-
каций.
рованной характеристикой регуляции, завися-
щей от предшествующих состояний среды, про-
Дальнейшее увеличение параметров при зна-
цесса исчерпания/восстановления ресурсов и ха-
чениях rτ > π/2 вызывает переход в режим колеба-
рактеристик онтогенеза. Именно балансовое
ний негармонической формы - релаксационных
состояние K как емкость экологической ниши яв-
циклов. Быстрое возрастание амплитуды колеба-
БИОФИЗИКА том 65
№ 1
2020
146
ПЕPЕВАPЮXА
Рис. 5. Возникновение релаксационного цикла в уравнении (2).
ний выраженной конической формы при увели-
из-за конкуренции между различными стадиями
чении временного промежутка между максиму-
у мух за лимитированные пищевые ресурсы.
мами и минимумы, стремящиеся к неотличимым
Уравнение (3) аналогично дискретной модели
от нуля значениям, приводят такой релаксацион-
пополнения
запасов
рыб
Рикера:
ный цикл (рис. 5) к затруднениям при обоснова-
Rn+1 = ϒRnexp(-bRn-i) - qRn, где показатель b от-
нии даже динамики насекомых. Минимумы ста-
ражает влияние переполнения нерестилищ при
новятся очень долгими и глубокими в ε-окрест-
высокой плотности, a q ∈ [0,1] - доля промысло-
ности нуля, пики - резкими и разреженными.
вого изъятия из нерестового запаса. Естественно,
Подобная популяция в реальности бореальных
что бифуркации у моделей-аналогов в итерацион-
лесов вымерла бы от случайных возмущений. Бо-
ной и в непрерывной форме различны [33]. Иссле-
лее оправдан для этой модели сценарий ошибки
дование поведения уравнения (3) оставляет не-
переполнения и завершение расчетов после чрез-
сколько открытых проблем для математики [34].
мерной вспышки - вариант гибели популяции
после исчерпания ресурсов среды.
МОДИФИКАЦИЯ УРАВНЕНИЯ
Для всех модификаций непрерывной модели
ХАТЧИНСОНА
Хатчинсона применительно к специфическим
Колебания колебаниям рознь в живых систе-
экологическим ситуациям актуален вопрос: что
мах и могут быть сложно устроены, например, из
отражает и как трактовать действие величины ем-
быстрых и медленных составляющих, как траек-
кости среды K во время такого нестационарного
тории-утки [35]. Флуктуации с большой амплиту-
режима, как вспышка численности? Имеет смысл
дой и высокими пиками для моделирования ди-
отказаться от априори заданной величины K для
намики общей численности популяций не долж-
случаев, когда нарушаются принципы плотност-
ны проходить окрестность нуля. Пусть действие
ной регуляции воспроизводства. Известно аль-
на регуляцию численности насекомых при коле-
тернативное дифференциальное уравнение для
баниях оказывает некоторая масштабируемая
возникновения автоколебаний большой ампли-
разность между потенциальной емкостью эколо-
туды, обходящееся без явного K-параметра емко-
гической ниши и состоянием популяции в мо-
сти и дважды включающее τ:
= |t - τ|. Идея с относительным положени-
мент φt
ем емкости ниши и влияющей разностью K от
dN = rN
(t
−τ)exp(−bN
(t
−τ))
−δN t),
(3)
N(t - τ) ранее использовалась в модели Гополса-
dt
ми-Куленовича [36]:
где присутствует экспоненциальная нелиней-
ность. Параметр b отражает давление негативных
dN
⎛
K -N(t
−τ)
⎞
=
rN t)
(4)
факторов, зависящих от плотности. Уравнение (3)
⎜
⎟
dt
⎝K + jN(t
−τ)⎠
известно под названием «Nicholson's blowflies dif-
ferential equation». Считается, что это уравнение
Насекомые вредители не просто занимают до-
лучше согласуется с данными экспериментов ав-
ступную экологическую нишу, но при вспышке
стралийского энтомолога Николсона. В лабора-
разрушают собственную среду, потому допустима
торных экспериментах он не наблюдал внешнего
терминальная интерпретация предельной чис-
давления на популяцию. Колебания возникали
ленности в модели - завершение существования.
БИОФИЗИКА том 65
№ 1
2020
НЕПРЕРЫВНАЯ МОДЕЛЬ ДЛЯ ОСЦИЛЛИРУЮЩЕЙ ВСПЫШКИ
147
Рис. 6. Появление незатухающих треугольных осцилляций в модели (5).
Для новой модификации мы неравномерно уве-
тянутого. Модели популяционной динамики для
личим степень нелинейности в числителе и зна-
неявного демпфирования вдруг возникших ос-
менателе и предложим уравнение, где дважды
цилляций не предлагались, рассматривались мо-
дели для определения внешнего управляющего
включено Nk(t - τ), C < K:
воздействия по вынужденному переводу колеба-
2
тельной системы в стационарный режим в [37].
dN
⎛
С -N
(t
−τ)
⎞
=
rN t)
(5)
⎜
3
⎟
Модификации моделей (3) и (4) обычно были на-
dt
⎝
K
+γN
(t
-τ)⎠
правлены на усложнение функций саморегуля-
Здесь мы переходим к представлениям об отно-
ции популяции, например, с введением периоди-
сительности для явной емкости экологической ни-
чески изменяющихся коэффициентов уравне-
ши, где K - только нижний возможный предел, ее
ний, однако независимое внешнее давление
инфинум. В новом уравнении (5) мы смогли пре-
может быть выражено сложнее, чем пропорцио-
одолеть недостаток слишком глубоких минимумов у
нальное изъятие c δ = const.
цикла. В вычислительных исследованиях для урав-
Для описания специфического развития по-
нения (5) после бифуркации Андронова-Хопфа при
пуляционного процесса более обосновано ис-
увеличении r получим реализацию несимметричных
пользование не степенной (как в уравнении (5)),
треугольных осцилляций большой амплитуды
а экспоненциальной нелинейности (как в уравне-
не вокруг неустойчивого равновесия как в уравне-
нии (3)) в плотностной регуляции, когда значе-
нии (2), но над равновесием (рис. 6).
ние понятия «емкость ниши» в условиях вспышек
Основной значимый результат новой модифика-
относительно. Предложим модель в предположе-
ции (уравнение (5)) - очередной стремительный не-
нии об относительно небольшой исходной груп-
контролируемый рост численности начинается от
пе особей вида N(0) > L с большим репродуктив-
значения, близкого к существовавшему перед би-
ным параметром, способной генерировать в от-
фуркацией равновесию, оптимального баланса со
дельных условиях пилообразную вспышку, где
средой. Колебания получаются частые и с одинако-
важным будет функционал внешнего биотиче-
вой амплитудой (впрочем, это можно компенсиро-
ского сопротивления. Убыль численности от био-
вать стохастическим возмущением). В реальной ди-
тических факторов хищничества и паразитизма в
намике локальной вспышки листовертки мы отме-
реальности представляет нетривиальную зависи-
чали другую характеристику - самозатухание.
мость, которая может включать пороговые значе-
ния эффективности подавления из-за усиления
реакции паразитов на массовую доступную жерт-
МОДЕЛИРОВАНИЕ ПРОИЗВОЛЬНО
ву [38]. Чем ближе численность разрушающего
ЗАТУХАЮЩЕЙ ВСПЫШКИ
среду вредителя к предельному порогу A, тем
Поведение модификации модели (4) не закры-
больше будет возрастать противодействие. Со-
вает проблемы для интерпретации осциллирую-
противление биологического окружения агрес-
щих экстремальных колебаний. Для пилообраз-
сивному размножению включается с запаздыва-
ной вспышки коконопряда в Канаде отмечено
нием и зависит от состояния биосистемы в про-
сокращение амплитуды у пиков колебаний. Для
шлом, причем это запаздывание будет отличным
любого процесса массового размножения актуа-
(большим) по значению, чем то, которое исполь-
лен переход в фазу его затухания, резкого или рас-
зуется в функции саморегуляции. Отложенная
БИОФИЗИКА том 65
№ 1
2020
148
ПЕPЕВАPЮXА
Рис. 7. Затухающая пилообразная вспышка в модели (6) при изменении параметра r: r1 = 5, r2 = 40, N(0) = 650,
τ = τ1 = 27, b = 5 · 10-2.7, A = 5900, q = 27, n = 2.
реакция у фактора противоборства вторжению
ние для резких популяционных явлений, но не
наглядна в аспекте запаздывающего антигенспе-
самое достоверное. Иногда трудно обосновать
цифического иммунного ответа на инфекцию.
резкое увеличение репродуктивного потенциала
r, если вид не инвазионный. Модель (6) отличает-
Используем вместо -δN(t) из уравнения (3)
ся тем, что может генерировать переход к непро-
функцию нелинейно-порогового сопротивления
должительному интервалу колебаний численно-
среды с запаздыванием в следующей форме
сти и обратно к равновесию без изменения ее па-
(n ≤ 2):
раметров. Для перехода к вспышке достаточно
dN
N(t
−
nτ)
резко вывести популяцию из устоявшегося рав-
=
rN(t
−τ)exp(−bN(t
−τ))
−
q
(6)
новесия, например, сократив численность на
dt
A-N(t
−τ
1
)
21%, как в имитационном сценарии, представ-
Модель (6) демонстрирует переход к медленно
ленном на рис. 9.
затухающим осцилляциям на уровне некоторого
Таким образом, мы получили реализацию сце-
не оказывающего давления на среду равновесия,
нария, где переход к масштабным, но затухаю-
форма колебаний зависит от N(0) и n. В вычисли-
щим колебаниям зависит не только от резких пе-
тельном эксперименте, представленном на рис. 7,
ремен популяционных характеристик, но и от
мы при резком изменении репродуктивного па-
быстрых перепадов состояния самой популяции.
раметра от r1 = 5 до r2 = 40 и τ = τ1, n =2 в уравне-
Если исходное значение N(0) приближается к ба-
нии (6), что может вызываться аномально благо-
лансовому равновесию, колебательную динамику
приятным климатическим фактором, вместо рас-
мы не обнаружим. Для вспышки нужна некото-
ходящихся колебаний в уравнениях (2) и (3) при
рая группа особей, где минимальны конкурент-
обычной бифуркации увидим затухающий релак-
ные факторы саморегуляции, но при этом она
сационный цикл над равновесием. Итоговое ба-
больше, чем порог L эффекта Олли. При добавле-
лансовое равновесие с r2 только сдвинется не-
нии линейной убыли, как в уравнении (3), наряду
сколько вверх.
с пороговой запаздывающей функцией мы полу-
чим возможность колебаний с неограниченно
Уменьшение коэффициента q демпфирует по-
возрастающей амплитудой и с отрицательными
следующие осцилляции, увеличение n удлиняет
значениями N. В модификации на основе уравне-
интервал времени между уменьшающимися пи-
ния (5) вместо прохождения минимальных значе-
ками. Интересно имитировать временное резкое
ний можно наблюдать полное исчезновение по-
увеличение r-параметра. В эксперименте, пред-
пуляции:
ставленном на рис. 8, значение параметра с r1 = 6
мы увеличили до r2 = 46 и через время, равное
2
dN
⎛
С -N
(t
−τ)
⎞
N(t
− τ
t = 100 единиц времени вычислительной среды,
=
rN t)
−
q
(7)
⎜
3
⎟
dt
K +γN
(t
−τ)
A-N(t
−τ)
вернули значения параметров в исходное состоя-
⎝
⎠
ние. Колебания не затухли моментально, но про-
Вариант (7) более сложен для согласования
должились, сходясь к прежнему равновесию.
расчетов. В предложенном непрерывном вариан-
Изменения параметров и следующие за ними
те описания пульсирующей вспышки все транс-
бифуркации - обычное математическое объясне-
формации поведения решения уравнения
(6)
БИОФИЗИКА том 65
№ 1
2020
НЕПРЕРЫВНАЯ МОДЕЛЬ ДЛЯ ОСЦИЛЛИРУЮЩЕЙ ВСПЫШКИ
149
Рис. 8. Временное увеличение параметра r в модели (6): r1 = 6, r2 = 46, N(0) = 650, τ = τ1 = 27, b = 5 · 10-2.7, A = 5900,
q = 37, n = 2.
происходят без явно заданных пороговых состоя-
инструментальной среды о недопустимых значе-
ний траектории и метаморфозов аттракторов. В
ниях. Интерпретируемые с точки зрения динами-
модели при изменении основных характеристик
ки численности насекомых переходные режимы
меняется форма переходного циклического ре-
поведения могут присутствовать в моделях мате-
матической биологии вне зависимости от воз-
жима, колебания получаются над по-прежнему
никновения бифуркаций и существования аль-
существующим притягивающим равновесием, а
тернативных аттракторов.
не вокруг потерявшей устойчивость особой точ-
ки, как было в уравнении (2). Как в любой нели-
нейной популяционной модели, в предложенных
ЗАКЛЮЧЕНИЕ
нами уравнениях (5), (6) или (7) необходимо раз-
граничивать биологически обоснованные пара-
В работе предложена непрерывная модель ло-
метрические диапазоны интересного для экоди-
кальной вспышки, развивающейся как серия за-
намики поведения траектории. Так, в имитаци-
тухающих пилообразных колебаний, которые ха-
онном эксперименте, представленном на рис. 8,
рактерны для кольчатого коконопряда в лесах Во-
при r > 48, τ > 28 существование популяции
стока Канады. Режим пилообразных колебаний
продолжить невозможно, также при N(0) > A,
переходный для динамической системы - «tran-
N(t) → ∞ вычисления закончатся сообщением
sient mode». Для любого переходного режима его
Рис. 9. Повторное возникновение осцилляций в модели (6) после индуцированного извне нарушения равновесия:
N1(0) = 640, N2(0) = 666, r = 48, равновесие K = 1067.
БИОФИЗИКА том 65
№ 1
2020
150
ПЕPЕВАPЮXА
длительность и выраженность прохождения зави-
цесса, потому полученные имитационные сцена-
сят помимо параметров от начальных условий. В
рии не исчерпывают всю возможную динамику.
предыдущей модели (1) переходный хаотический
Особенно интересны различия, если посмотреть
режим удачно имитировал действие комплекса
на хорошо документированные явления в экоси-
случайных внешних факторов на достижение по-
стемах через призму математической теории ди-
роговой численности псиллид. Развитие сцена-
намических систем. Например, инвазионный вид
рия как вспышки произойдет при наличии исход-
после стремительной вспышки может проходить
ной небольшой (но не околонулевой) группы
критическое состояние «бутылочного горлыш-
особей. Моновольтинная популяция в исходном
ка», с сохранением реликтовой популяции либо
состоянии, близком к существующему балансу со
полным исчезновением из нового ареала. Полу-
средой, согласно сценарию не имеет потенциала
ченный в уравнении переходный режим можно
для бурного роста. Рассмотрен в имитационных
рассматривать также для задачи анализа случая
вычислительных экспериментах как параметри-
специфического развития рецидивирующей ин-
чески зависимый сценарий запуска пульсирую-
фекции. Интересно дальнейшее расширение
шей вспышки, так и обходящийся без параметри-
уравнений для модельных исследований инвази-
ческих изменений. Достаточно временного, но
онных процессов других видов с независимым
аномального повышения репродуктивного пара-
противодействием, например динамика осцил-
метра r в уравнении (6) на три сезона, чтобы пи-
лирующей вспышки вредителей лесов в канад-
лообразные колебания сохранялись достаточно
ских провинциях Онтарио и Нью-Брансуик каче-
длительное время, как афтершоки после толчка
ственно схожа с изменениями численности при
землетрясения. Каждый пик в этом случае не са-
инвазии Mnemiopsis leidyi в Южном Каспии в
мостоятельная вспышка, но часть процесса. По-
2000-2010 гг. после появления вредоносного
том ситуация возвращается к равновесию, где на-
гребневика у берегов Ирана [41].
секомые уже не вызывают дефолиации леса.
Модель основана на учете проявления запаз-
ФИНАНСИРОВАНИЕ РАБОТЫ
дывания и при регуляции, связанной с зависящей
от конкуренции эффективностью воспроизвод-
Работа выполнена в рамках проекта Россий-
ства и при формировании ответа со стороны био-
ского фонда фундаментальных исследований
тического окружения. В результате после серии
(руководитель А.Ю. Переварюха) при частичном
колебаний динамика затухает и переходит в
финансировании за счет бюджетной темы
устойчивое равновесие при обычной для среды
СПИИРАН AAAA-A16-116051250009-8.
численности. В уравнении (5), модификации мо-
дели (2), после бифуркации Андронова-Хопфа
КОНФЛИКТ ИНТЕРЕСОВ
происходит появление устойчивых колебаний,
где пик начинается около неустойчивого равно-
Авторы заявляют об отсутствии конфликта
весия.
интересов.
Спровоцировать следующую серию колебаний
с дефолиацией в такой модели могут стремитель-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
ные изменения состояния насекомых, в пределах
четверти численности популяции. Вызвать изме-
Настоящая работа не содержит описания ка-
нения способны нарушения условий выживаемо-
ких-либо исследований с использованием людей
сти при диапаузе или миграция насекомых между
и животных в качестве объектов.
соседними областями, связанная, вероятно, с ло-
кальным исчерпанием ресурсов леса. Когда попу-
СПИСОК ЛИТЕРАТУРЫ
ляция насекомых становится еще меньше мини-
мального баланса, то это немедленно негативно
1. C. Lee and G. Gelembiuk, Evolutionary Applications 1
отражается на снижении численности паразитов,
(3), 427 (2008).
что нарушает на некоторое время механизмы ре-
2. M. Nilssen, Fisheries Research 82 (1), 319 (2006).
гуляции. Полученные модели можно использо-
3. А. Н. Демидова, Наука и жизнь, № 10, 58 (2017).
вать в составе систем уравнений явного межвидо-
4. N. Chapman, Ecological Monographs
9
(3),
261
вого противоборства, они вычислительно менее
(1939).
5. Q. Jin and L. Valsta, Inter. J. Forestry Res. 6 (3), 12
сложны, чем современные модели трофодинами-
(2015).
ки «хищник-жертва» [39], но обладают каче-
6. А. Н. Фролов и И. В. Грушевая, Вестн. защиты
ственным разнообразием поведения. Часто по-
растений 98 (4), 18 (2018).
вышенная плотность насекомых сопровождается
7. D. Ludwig, D. Jones, and S. Holling, J. Animal Ecol.
их физиологическими трансформациями и сме-
47 (1), 315 (1978).
ной экологической роли [40].
8. M. A. Rozendaal and R. K. Kobe, PLoS One 11 (11),
Вспышки автохтонных видов менее разнооб-
167 (2016).
разны по аспектам прохождения фаз, чем экстре-
9. А. С. Исаев, В. Г. Суховольский и Р. Г. Хлебопрос,
мальные варианты развития инвазионного про-
Лесоведение 2, 3 (2010).
БИОФИЗИКА том 65
№ 1
2020
НЕПРЕРЫВНАЯ МОДЕЛЬ ДЛЯ ОСЦИЛЛИРУЮЩЕЙ ВСПЫШКИ
151
10. В. Г. Суховольский, В. И. Пономарев и Г. И. Со-
26. R. Louis-Etienne, R. Brian, and J. Cooke, Ecography
колов, Журн. общ. биологии 76 (3), 179 (2015).
41 (9), 1556 (2018).
11. A. Y. Perevaryukha, Biophysics 61 (2), 334 (2016).
27. T. Hlásny and J. Trombik, J. Pest Sci. 89 (2), 413 (2016).
12. L. R. Clark, Austral. J. Zool. 12 (3), 362 (1964).
28. C. Loehle, Ecol. Modelling 49 (2), 125 (1989).
13. E. P. Odum, Ecology (Rinehart & Winston, N.Y.,
29. Z. S. Ma and E. J. Bechinski, Entomol. Res. 39 (3), 175
1963).
(2009).
14. V. A. Trjapitzin and M. G. Volkovitsh, Entomol. Rev.
30. D. R. Brillinger, J. Time Series Analysis 33 (5), 718
91 (5), 670 (2011).
(2012).
15. W. C. Allee and E. Bowen, J. Exp. Zool. 61 (2), 185
31. G. Hutchinson, Ann. N. Y. Acad. Sci. 50 (4), 221
(1932).
(1948).
16. A. A. Hall, Ecology and evolution of Cardiaspina psyllids,
32. Б. Хэссард, Н. Казаринов и И. Вэн, Теория и
their bacterial endosymbionts and parasitoid wasps (Doc-
приложения бифуркации рождения цикла (Мир, М.,
tor of Philosophy Thesis) (Western Sydney University,
1985).
2016).
33. Г. К. Каменев, Д. А. Саранча и В. О. Поляновский,
17. A. A. Hall, S. N. Johnson, and J. M. Cook, Insect Sci.
Биофизика 63 (4), 758 (2018).
26 (2), 351 (2019).
34. Т. Л. Сабатулина, Изв. вузов. Математика 47 (11),
18. J. Berry, Biosecurity New Zealand 68 (2), 18 (2006).
50 (2010).
19. H. Myers, Am. Sci. 81 (3), 240 (1993).
35. Е. В. Трошкина, Вестн. СамГУ. Естественнонаучн.
20. C. Bone and S. Dragicevic, Ecol. Modelling 192 (1-2),
сер. 9 (2) 215 (2013).
107 (2006).
36. K. Gopalsamy, M. Kulenovic, and G. Ladas, Applica-
21. B. Cooke, S. V. Neali, and J. Regniere, in Plant distur-
ble Analysis 31 (3), 225 (1988).
bance ecology: the process and the response (Elsevier,
37. Г. Е. Колосов и М. М. Шаров, Автоматика и
Burlington, 2007), p. 487-525.
телемеханика 53 (6), 146 (1992).
22. T. Royama, Ecol. Monographs 54 (4), 429 (1984).
38. J. Grieshop, W. Flinn, and R. Nechols, J. Insect Sci. 10
23. B. J. Cooke, F. Lorenzetti, and G. Roland, J. Entomol.
(1), 99 (2010).
Soc. Ont. 140, 3 (2009).
39. А. В. Епифанов и В. Г. Цибулин, Биофизика 61 (4),
24. ESTR Secretariat Atlantic Maritime Ecozone evidence for
823 (2016).
key findings summary (Canadian Biodiversity: Ecosystem
Status and Trends, 2010), Report № 3 (Canadian Coun-
40. В. Г. Суховольский, Биофизика 48 (2), 337 (2003).
cils of Resource Ministers, Ottawa, 2014).
41. A. Roohi and Z. Yasin, Marine Ecology 29 (4), 421
25. X. Zhang, Ecol. Evol. 4 (12), 2384 (2014).
(2008).
A Continuous Model for Oscillating Outbreak Population of the Phytophagous Moth,
Tent Caterpillar, Malacosoma disstria (Lepidoptera, Lasiocampidae)
A.Yu. Perevaryukha
St. Petersburg Institute for Informatics and Automation, Russian Academy of Sciences,
14-ya Linia 39, St. Petersburg, 199178 Russia
Outbreaks of individual species population are important phenomena in many aspects and not alike in terms
of a theory of multispecies community dynamics. Outbreaks of insect population develop more quickly with
long-lasting effects experienced by the forest industry. These events are considered extreme unbalanced and
transient processes. The mechanisms of the development and subsidence of insect outbreaks differ in differ-
ent taxonomic groups of pests. The duration and occurrence of repeated outbreaks of psyllids and forest
moths, affecting deciduous or coniferous forests in the same region, are different. Computational simulation
is needed for understanding the dynamics of insect outbreaks. For the mathematical description of the out-
breaks of forest tent caterpillar, in addition to the threshold version of the development of the insect outbreak,
it is interesting to modify the continuous computational models for the analysis of fluctuation dynamics. In
this paper, we simulate dynamics of spontaneously damped oscillations under specific scenario during pop-
ulation outbreak using a continuous model with delayed regulation and nonlinear counteraction from the bi-
otic environment. The scenario described by the new phenomenological equation, which consists of a series
of different-sized maxima and with a final attenuation of peaks near balance, is realized for the pest tent cat-
erpillar, Malacosoma disstria, which affects deciduous forests in North America leading to large-scale defoli-
ation. The new scenario is qualitatively different from our model of the threshold development and subsid-
ence of outbreaks of the psyllid, Cardiaspina albitextura, in Australia.
Keywords: extreme ecosystem dynamics, insect outbreaks, damped population fluctuations, models of oscillatory
dynamics of populations, equations with delay, bifurcations, forest tent caterpillar Malacosoma disstria, forest de-
foliation
БИОФИЗИКА том 65
№ 1
2020