БИОФИЗИКА, 2020, том 65, № 1, с. 198-201
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 573.7.017.6+519.711.3
МОДЕЛИРОВАНИЕ ПОЛНОЙ КРИВОЙ СМЕРТНОСТИ ЧЕЛОВЕКА:
РЕГУЛЯТОРНАЯ МОДЕЛЬ СТАРЕНИЯ
© 2020 г. В.И. Донцов, В.Н. Крутько
Институт системного анализа Федерального исследовательского центра «Информатика и управление» РАН,
117312, Москва, просп. 60-летия Октября, 9
E-mail: dontsovvi@mail.ru
Поступила в редакцию 04.05.2019 г.
После доработки 21.11.2019 г.
Принята к публикации 25.11.2019 г.
Моделирование процесса старения организма человека основывалось на связи общей жизнеспо-
собности организма с процессами роста и самообновления тканей, регулируемых предположитель-
но центрами вегетативного мозга. Наличие двух регуляторных центров, стимулирующих и ингиби-
рующих такой рост, и спонтанная гибель клеток таких центров с различающейся для каждого
скоростью позволяют моделировать этапы роста, остановки и снижения скорости роста-самооб-
новления тканей. Получаемая кривая соответствует реальной полной кривой интенсивности
смертности для популяций, что, как известно, наилучшим образом описывает процессы роста и
старения. Такое соответствие модели и кривой смертности, обычно описываемой формулой Гом-
перца только для средней части кривой, получено впервые. Модель соответствует регуляторной
теории старения и связывает процессы старения с процессами регуляции роста и самообновления
тканей
Ключевые слова: развитие, старение, смертность, модели старения, рост тканей.
DOI: 10.31857/S0006302920010226
Исследователями было предложено немало
Для сравнения модели с реальной кривой
моделей старения, однако до настоящего време-
смертности была выбрана оптимальная страна в
ни наилучшей и практически используемой яв-
период наиболее благоприятных исторических
ляется феноменологическая модель Б. Гомперца
условий, для минимизации влияния внешних
(B. Gompertz, 1825), описывающая возрастную
факторов на кривую смертности. Были рассчита-
смертность популяций [1]. Однако эта модель
ны величины интенсивности смертности. Дан-
описывает только смертность средних возрастов -
ные смертности для различных стран были взяты
монотонное экспоненциальное повышение
смертности - и не способна моделировать на-
отражающем динамику смертности для 40 стран
чальные и конечные формы реальной кривой
на протяжении двух с половиной веков. Исполь-
смертности человека и млекопитающих, а также
зовали величину интенсивности смертности, так
не имеет четкого биологического содержания -
как именно этот показатель рассматривается ге-
связи с конкретными физиологическими меха-
ронтологами как оптимальный для описания из-
низмами.
менения скорости старения с возрастом.
Целью данной работы является построение
модели старения, описывающей все этапы реаль-
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
ной кривой смертности человека и связывающей
Модель построена на представлениях о взаи-
старение и смертность с основными жизненными
мосвязи старения с процессами роста и развития
процессами - ростом и самообновлением тканей
организма.
организма и самообновлением его тканей как
главной силе жизнеспособности, противостоя-
щей старению [2-7], и объединяет теории стоха-
МЕТОДЫ ИССЛЕДОВАНИЯ
стических повреждений и регуляторные теории
Для моделирования использовали дифферен-
старения [2-4, 6-14].
циальные уравнения, которые моделировали на
В основу модели положены максимально про-
компьютере и отображали в виде графика, по-
стые допущения, имеющие ясный биологиче-
строенного в программе Excel.
ский смысл:
198
МОДЕЛИРОВАНИЕ ПОЛНОЙ КРИВОЙ СМЕРТНОСТИ
199
– рост и самообновление организма управля-
ется двумя типами взаимодействующих регуля-
торных клеток (стимулирующих и ингибирую-
щих, аналогично центральной регуляции гормо-
нов гипоталамусом и гипофизом) с различными
скоростями спонтанной гибели;
- жизнеспособность эквивалентна скорости
самообновления тканей за счет механизмов кле-
точного роста и деления;
- смертность рассматривается как обратная
величина жизнеспособности.
На основе данных допущений возможно объ-
яснить как регуляцию роста живой системы, так и
прекращение этого развития в нужный момент с
последующим спонтанным старением.
Учитывая упомянутые допущения, возрастная
динамика регуляторных клеток может описы-
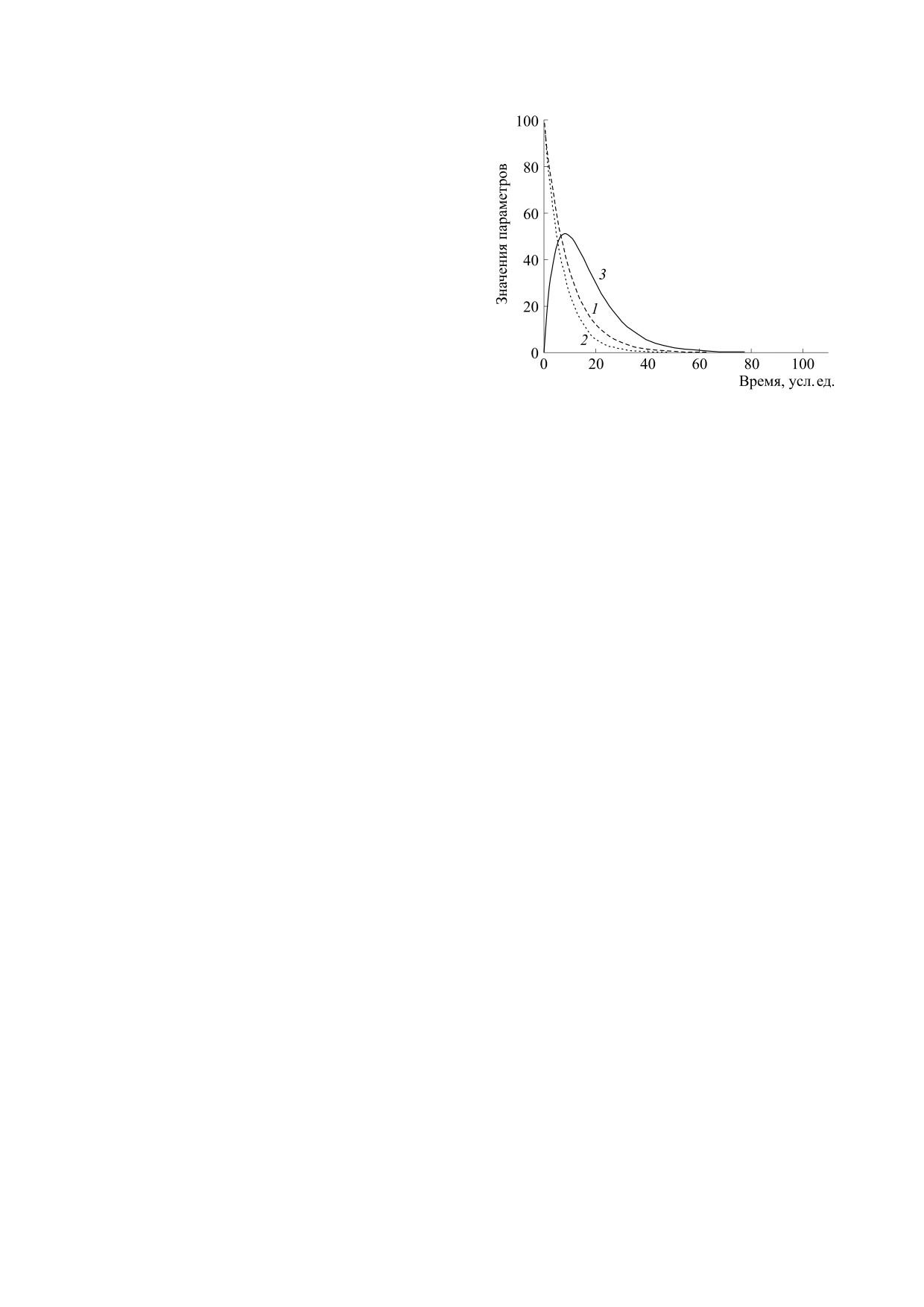
Рис. 1. Модель регуляторного старения как возраст-
ваться системой двух простых линейных диффе-
ной дистрофии тканей при изменении регуляции
клеточного роста. По вертикали - значения парамет-
ренциальных уравнений, используемых для опи-
ров, по горизонтали - время в условных единицах.
сания любых стохастических процессов распада
1 - Количество клеток-стимуляторов (h) для началь-
элементов (например, радиоактивный распад
ного h = 100 при спонтанной гибели 10% за единицу
происходит по тем же фундаментальным меха-
времени; 2 - количество клеток-ингибиторов (s) для
низмам и общим причинам и законам):
начального s =100 при спонтанной гибели 13% клеток
за единицу времени; 3 - содержание итогового регу-
dh/dt = -kh ⋅ h,
(1)
ляторного фактора (F = h - s), с коэффициентом про-
порциональности 5F.
ds/dt = -ks ⋅ s,
где h и s - соответственно количество стимулиру-
смертности населения, была добавлена константа A,
ющих (хелперных) и ингибирующих (супрессор-
равная 0.0007 для выбранной страны.
ных) клеток; kh и ks - коэффициенты интенсив-
Если предположить, что ингибирующие клет-
ности вероятностной гибели соответствующих
ки быстрее разрушаются с возрастом (их функция
типов клеток.
исчерпывается периодом развития: они предна-
Исходя из наиболее простого допущения о
значены для растормаживания при их гибели сти-
том, что продукция некого конечного регулятор-
мулирующих влияний в период быстрого роста
ного фактора F в организме пропорциональна
организма, когда масса тканей увеличивается по
разности между количеством стимулирующих и
сравнению с массой тела при рождении более чем
ингибирующих клеток, получаем соотношение:
на порядок), то из полученных уравнений можно
F = kf (h - s) + С,
(2)
получить динамику моделируемых переменных,
показывающую очень хорошее качественное со-
где kf - коэффициент, С - некоторая константа.
ответствие с реальной кривой смертности челове-
Если считать регуляторный фактор главным
ка (рис. 1 и 2).
фактором жизнеспособности, обеспечивающим
В отличие от первого, чисто стохастического
интегральное функционирование организма как
механизма гибели жизнеспособных элементов
системы, в частности, обеспечивающей регенера-
организма в целом, позволяющего моделировать
цию тканей, то можно допустить, что величина F
только среднюю часть кривой смертности с по-
характеризует жизнеспособность организма и в
мощью уравнения Гомперца, данная модель от-
простейшем случае пропорциональна ей. Тогда
ражает все части кривой истинной смертности -
для смертности как для величины, обратной жиз-
высокую начальную смертность с последующим
неспособности, получим выражение:
снижением и некоторым минимумом в период
m = km ⋅ (1/F),
(3)
роста организма, последующее экспоненциаль-
ное повышение смертности в течение основного
где km - коэффициент пропорциональности.
периода жизни и некоторое снижение в самых
старших возрастах.
Для практических целей компьютерного моде-
лирования использовали следующие количествен-
Для сравнения модели с реальной кривой
ные значения коэффициентов: m = 1/(F ⋅ 375 + C);
смертностью были выбраны несколько стран в
при C = 1; kh = 0.1; ks = 0.13. Для учета смертности от
период относительно благоприятных историче-
внешних причин, всегда влияющих на уровень
ских условий (Австралия, Дания, Канада, Нидер-
БИОФИЗИКА том 65
№ 1
2020
200
ДОНЦОВ, КРУТЬКО
Все это может указывать на определяющую
роль процессов регуляции для старения человека.
Давно известные эффекты гипофизэктомии на
возрастную инволюцию тимуса [15] и разработан-
ные методы трансплантации мозговой эмбрио-
нальной ткани [5] позволяют влиять на восста-
новление истощенных регуляторных программ у
старых животных. Альтернативой являются мето-
ды фармакологической или физиотерапевтиче-
ской активации соответствующих ядер гипотала-
муса, а также создание новых функциональных
регуляторных центров и водителей ритмов, в том
числе с применением (ауто)психотерапевти-
ческих техник, гипноза и пр. На уровне пери-
ферических механизмов наиболее перспектив-
ным представляются иммунофармакологические
средства, влияющие на лимфоциты - регуляторы
роста соматических клеток, а также выделенные
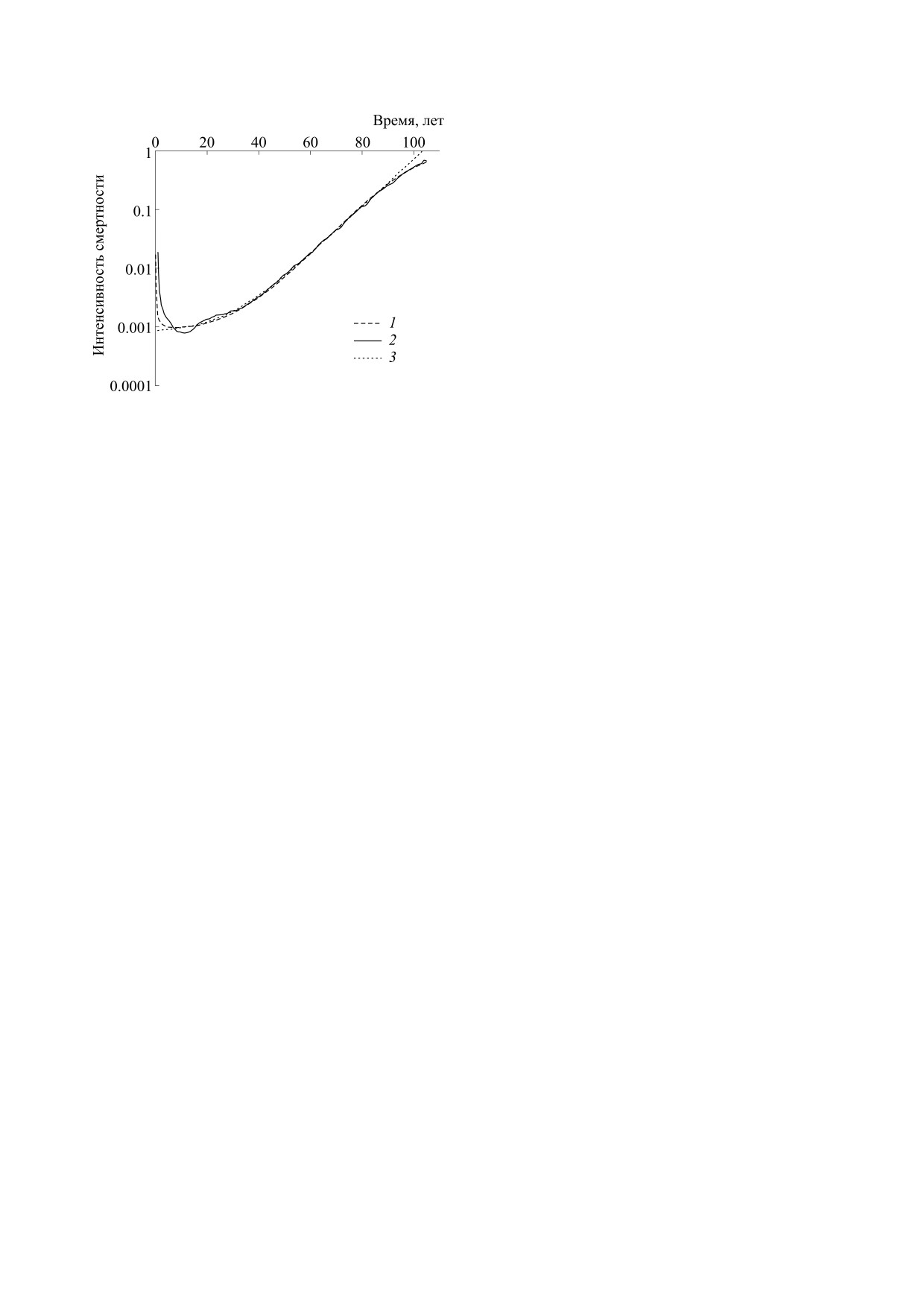
Рис. 2. Математическая модель регуляторной теории
смертности и реальная интенсивность смертности в
из крови молодых растущих животных факторы
Австралии (1940 г.). По оси абсцисс - время в годах,
роста, количество которых выраженно снижается
по оси ординат - интенсивность смертности (в лога-
с возрастом [2, 16].
рифмическом масштабе). 1 - Расчетная величина
смертности согласно предлагаемой модели с указан-
ными в тексте коэффициентами, 2 - реальная интен-
сивность смертности (данные представлены на сайте
ЗАКЛЮЧЕНИЕ
муле Гомперца: m = 7,95E-4 + 6,13E-5*exp(9,37E-2*t).
Разработана математическая модель регуля-
торного механизма старения, с описанием пол-
ной кривой смертности человека и ясной биоло-
ланды и др. в 1910-1940 гг., а также в 1950 г.). На
гической интерпретацией, связанная с представ-
рис. 2 показаны для примера графики для Австра-
лениями о старении как этапе роста и развития
лии (1940 г.): реальный график интенсивности
организма. В основе модели лежит взаимодей-
смертности, расчетный график по формуле Гом-
ствие влияющих на рост тканей регуляторных
перца (отклоняющийся от реального графика в
клеток вегетативных центров регуляции мозга
начальной и конечной части) и расчетный график
стимулирующего и ингибирующего типа, с раз-
по предлагаемой нами формуле, совпадающий с
личной скоростью спонтанной гибели, что поз-
реальной кривой наилучшим образом во всех воз-
воляет моделировать периоды роста, его оконча-
растных интервалах (коэффициент корреляции
ния и процесс старение. Смертность рассматри-
r = 0.999).
вается как обратная величина жизнеспособности,
Качественное соответствие модели с имеющи-
а жизнеспособность принимается пропорцио-
мися реальными демографическими кривыми
нальной скорости клеточного роста как основы
интенсивности смертности и является результа-
самообновления тканей.
том моделирования, оно адекватно поставленной
задаче, носящей прежде всего биологический ха-
Предложенная модель регуляторного старе-
рактер. Реальным морфологическим субстратом
ния впервые позволяет описать одновременно ха-
(параметры h и s клетки) описываемого меха-
рактерные изменения начальной, средней и ко-
низма могут быть регуляторные неделящиеся
нечной частей кривой смертности человека, сов-
клетки гипоталамуса, продуцирующие факторы
падающие с реальной картиной, и может быть
регуляции роста тканей; для периферийных меха-
физиологически интерпретирована.
низмов - разнообразные растущие и самообнов-
Модель указывает на возможную важную роль
ляющиеся делением соматические клетки.
регуляторных механизмов снижения самообнов-
Особую роль могут иметь некоторые механиз-
ления (клеточного деления) тканей с возрастом в
мы регулирования клеточного роста соматиче-
процессе старения человека и животных.
ских клеток: некоторые типы T-лимфоцитов ре-
гулируют не иммунитет, а рост соматических кле-
Так как регуляторные влияния, в отличие от
ток, которые, по нашему мнению, могут
стохастических механизмов, легко поддаются
составлять отдельную специальную систему им-
внешним управляющим воздействиям, это от-
мунного контроля роста соматических клеток [2-
крывает принципиально новые возможности ра-
4], а их возрастной иммунодефицит может лежать
дикального влияния на старение человека и мле-
в основе иммунной теории старения [2].
копитающих.
БИОФИЗИКА том 65
№ 1
2020
МОДЕЛИРОВАНИЕ ПОЛНОЙ КРИВОЙ СМЕРТНОСТИ
201
КОНФЛИКТ ИНТЕРЕСОВ
5. Ф. А. Атамурадова и В. И. Донцов, ДАН СССР 297
(1), 237 (1987).
Авторы заявляют об отсутствии конфликта
6. T. B. Kirkwood and S. Melov, Curr. Biol. 21, 701
интересов.
(2011).
7. R. F. Walker, Rejuvenation Res. 14, 429 (2011).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
8. В. Н. Крутько, В. И. Донцов, О. А. Мамиконова
и др., Мед. новости 2, 25 (2015).
Настоящая работа не содержит описания ка-
9. В. Н. Крутько, В. И. Донцов, О. В. Захарьящева
ких-либо исследований с использованием людей
и др., Авиакосмическая и экологическая медици-
и животных в качестве объектов.
на 48 (3), 12 (2014) .
10. L. Hayflick, PLoS Genet. 3, 220 (2007).
11. I. M. van Leeuwen, J. Vera, and O. Wolkenhauer, Phi-
СПИСОК ЛИТЕРАТУРЫ
los. Trans. R. Soc. Lond. B. Biol. Sci. 365, 3443 (2010).
1. B. Gompertz, Philos. Trans. Roy. Soc. Lond. A 115,
12. M. P. Murphy and L. Partridge, Annu. Rev. Biochem.
513 (1825).
77, 777 (2008).
2. В. И. Донцов, Новая иммунная теория старения:
13. T. A. Rando and H. Y. Chang, Cell 148, 46 (2012).
лимфоциты как регуляторы клеточного роста
14. R. S. Sohal and W. C. Orr, Free Radic. Biol. Med. 52,
(Lambert Acad. Publ., Saarbrucken, 2011).
539 (2012).
3. В. И. Донцов и В. Н. Крутько, Системный анализ
15. D. E. Harrison, J. R. Archer, and C. M. Astie, J. Immu-
и управление в биомедицинских системах 11 (3),
nol. 129, 2673 (1982).
657 (2012).
16. А. Я. Чижов, Е. С. Зенчук, В. Н. Крутько и
4. В. Н. Крутько и В.И. Донцов, Системные
В. И. Донцов, Технологии живых систем 10 (1), 41
механизмы и модели старения (ЛКИ, М., 2008).
(2013).
Postnatal Ontogeny and Aging: Regulatory Model Full Curve of Human Mortality
V.I. Dontsov and V.N. Krut’ko
Institute for Systems Analysis, Federal Research Center «Computer Science and Control» of Russian Academy of Sciences,
prosp. 60-letiya Oktyabrya 9, Moscow, 117312 Russia
A mathematical regulatory model of aging, that allows for the first time to receive the full curve of mortality
in humans and mammals and to have a clear biological interpretation has been created. The model can be
classified as a regulatory model related to ideas about the connection between ageing and processes of growth
and development of the organism. Interaction stimulating and inhibiting regulatory cells with different speed
spontaneous degradation allows to simulate a period of growth and development. Mortality is considered the
reciprocal of the viability, and viability speed is equivalent to cell growth and tissue self-renewal. The model
indicates the importance of regulatory mechanisms in tissues growth and self-renewal reduction with age,
during the aging process in humans and animals. As regulatory impacts, unlike stochastic mechanisms, are
susceptible to external influences, this opens up entirely new possibilities of radical influence on human aging
and mammals.
Keywords: development, aging, mortality, aging patterns, tissue growth
БИОФИЗИКА том 65
№ 1
2020