БИОФИЗИКА, 2020, том 65, № 2, с. 300-309
БИОФИЗИКА КЛЕТКИ
УДК 577.35
АНАЛИЗ УСТОЙЧИВОСТИ СИСТЕМЫ ВНУТРИКЛЕТОЧНОЙ
СИГНАЛИЗАЦИИ PI3K-Akt-mTOR
© 2020 г. Т.С. Сапега*, **, Г.Т. Гурия*, **
*Национальный медицинский исследовательский центр гематологии Минздрава России
125167, Москва, Новый Зыковский проезд, 4
**Московский физико-технический институт,
141700, Долгопрудный Московской обл., Институтский пер., 9
E-mail: guria@blood.ru
Поступила в редакцию 20.11.2019 г.
После доработки 11.12.2019 г.
Принята к публикации 24.12.2019 г.
Каскады системы внутриклеточной сигнализации MAPK и PI3K-Akt-mTOR регулируют
важнейшие клеточные процессы. В рамках текущих представлений о взаимодействии данных
каскадов сформулирована и исследована математическая модель. Анализ устойчивости модели
сигнального пути PI3K-Akt-mTOR позволил установить, что основное состояние сигнальной
системы является метастабильным, оно может быть пороговым образом активировано при
закритической экзогенной стимуляции. Для величины порогового уровня внешней стимуляции
получено явное выражение. Показано, что триггерные явления в динамике сигнальной системы
описываются элементарной катастрофой типа «складка». Обсуждается возможное значение
полученных результатов для молекулярной гематологии и гемостазиологии.
Ключевые слова: PI3K-Akt-mTOR, MAPK, внутриклеточная сигнализация, анализ устойчивости,
активация тромбоцитов.
DOI: 10.31857/S0006302920020118
ставление о ключевых веществах, участвующих в
Каскад внутриклеточной сигнализации PI3K-
передаче сигнала.
Akt-mTOR (PI3K
-фосфоинозитид
3-киназа;
Akt - белки семейства протеинкиназ В, кодируе-
Авторы исходили из того, что метаболические
мые генами AKT, mTOR -мишень рапамицина
сигнальные сети должны отвечать той же сово-
млекопитающих) играет важную роль в регуля-
купности априорных требований, которые при-
ции жизненного цикла клетки в норме [1, 2] и при
нято предъявлять ко всем иным сигнальным се-
тям, в частности электрическим. С этой точки
ряде патологий [3, 4]. Наиболее часто нарушения
передачи сигнала по данному сигнальному пути
зрения метаболические сети, находясь в ждущем
обсуждаются в контексте злокачественного пере-
режиме, должны быть готовы к срабатыванию
при поступлении извне в должной степени выра-
рождения клетки [3-6]. По последним сообще-
женного сигнала и должны быть способны игно-
ниям, PI3K-Akt-mTOR путь участвует в процес-
рировать недостаточно выраженные по амплиту-
сах активации тромбоцитов, а также при разви-
де внешние сигналы. Именно такого рода поро-
тии сосудистых мальформаций [7-9].
говые свойства метаболических сетей, по
Совокупность реакций, обеспечивающих
существующим представлениям, ответственны за
функционирование сигнальных каскадов, при-
их срабатывание в ситуациях, когда это действи-
нято схематически изображать в виде ориентиро-
тельно необходимо [12]. Такое поведение позво-
ванных графов [10, 11]. Такой способ представле-
ляет клеткам экономить ресурсы, игнорируя фо-
ния данных позволяет составить наглядное пред-
новые и случайные сигналы [13]. Другими слова-
ми, основное - «ждущее» - состояние всякой
метаболической системы должно быть устойчи-
Сокращения: PI3K - фосфоинозитид-3-киназа (phosphati-
dylinositol-3 kinase), Akt - белки семейства протеинкиназ вым к достаточно широкому кругу внешних сиг-
В, кодируемые генами AKT, mTOR - мишень рапамицина
налов, амплитуда которых ниже некоторого по-
млекопитающих (mammalian target of rapamycin), MAPK -
митоген-активируемые протеинкиназы (mitogen-activated
рогового уровня. В этой связи анализ устойчиво-
protein kinase).
сти основных состояний каждой конкретной
300
АНАЛИЗ УСТОЙЧИВОСТИ СИСТЕМЫ ВНУТРИКЛЕТОЧНОЙ СИГНАЛИЗАЦИИ
301
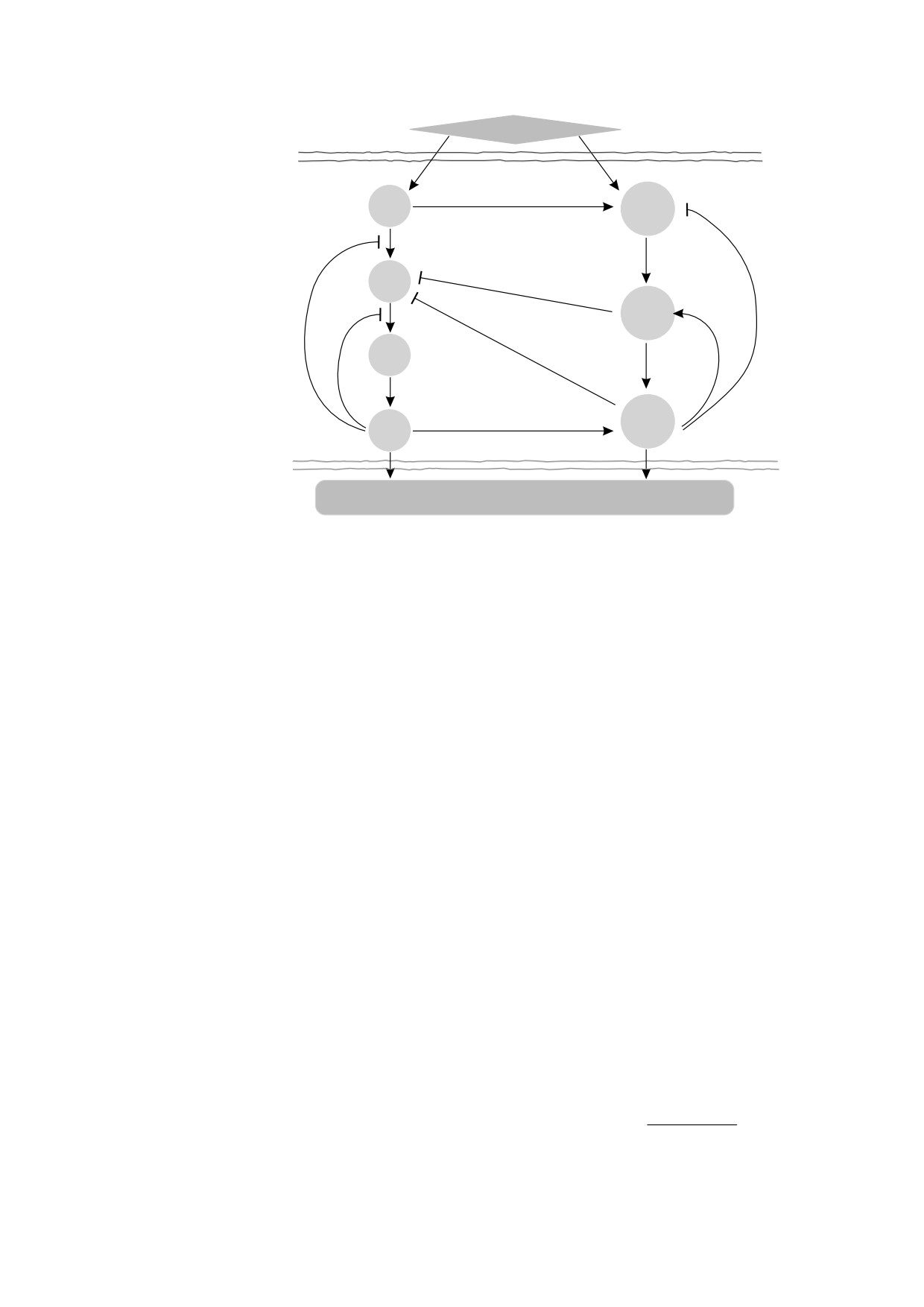
Внешний стимул
Цитоплазматическая
мембрана
RAS
PI3K
RAF
Akt
MEK
mTOR
ERK
Ядерная
мембрана
Регуляция экспрессии генов и белков. Изменение фенотипа
Рис. 1. Схема взаимодействия между сигнальными путями MAPK (белки RAS, RAF, MEK, ERK) и PI3K-Akt-mTOR
(белки PI3K, Akt, mTOR) (адаптировано из [18]). Стрелки отображают стимулирующую передачу сигнала, а линии с
тупым концом соответствуют негативной регуляции, ингибирующей прохождение сигнала.
метаболической сигнальной сети представляет
пути на автономное поддержание сигнала. Пото-
большой интерес. Данный круг вопросов рас-
му условия активации сигнального пути PI3K-
сматривается в концептуальном плане в рамках
Akt-mTOR представляют большой интерес.
теории метаболического контроля, являющейся
В соответствии со схемой, представленной на
сердцевиной современной системной биологии
рис. 1, характер взаимодействия основных элемен-
[14, 15].
тов сигнального пути PI3K-Akt-mTOR можно
В настоящей работе авторы ограничились ана-
представить в сокращенном виде (рис. 2а). При ана-
лизом устойчивости каскада внутриклеточной
лизе кинетики процессов в данном каскаде концен-
сигнализации PI3K-Akt-mTOR, играющего важ-
трации основных белков [PI3K], [Akt] и [mTOR] бы-
ную роль в активации тромбоцитов и целого ряда
ли обозначены как Y1(t), Y2(t) и Y3(t) соответственно.
иных клеток. Количественная оценка изменения
Предполагалось, что концентрации можно рассмат-
функциональных свойств сигнальной системы
ривать как динамические переменные, а константы
клетки при вариации кинетических параметров
скорости соответствующих реакций - k1, k2, k31 и
ее отдельных элементов изучалась с использова-
k32 - в качестве параметров. Для обозначения кон-
нием методов математического моделирования
стант скоростей пассивных утечек были введены па-
[16, 17].
раметры ¨1, ¨2, ¨3. Величина экзогенного стимула
была обозначена как S и рассматривалась как пара-
МАТЕМАТИЧЕСКАЯ МОДЕЛЬ
метр. Учитывая введенные обозначения, диаграмма
принимает вид, представленный на рис. 2б.
Система внутриклеточной сигнализации была
исследована на основе диаграммы взаимодей-
Динамика процессов, схематично представ-
ствия каскадов MAPK и PI3K-Akt-mTOR, пред-
ленных на рис. 2б, описывалась следующей си-
ложенной в работе [18] (см. рис. 1).
стемой обыкновенных дифференциальных урав-
нений:
Стоит отметить, что правая ветвь, содержащая
петлю положительной обратной связи на диа-
Y
=k
S -k
YY
−û
Y
,
(1)
1
0
31 1
3
1 1
грамме (Akt-mTOR-Akt), является важным мар-
кером динамических свойств системы. Это свой-
Y
=
k
Y
−û
Y
+
k
Y
,
(2)
2
1 1
2
2
32
3
ство указывает на принципиальную возможность
û
3
Y
3
возбуждения данного контура. В результате ста-
Y
=
k
Y
−
,
(3)
3
2
2
новится возможным переключение сигнального
(1
+Y
3
/
K
M
)
БИОФИЗИКА том 65
№ 2
2020
302
САПЕГА, ГУРИЯ
(a)
(б)
Внешний стимул
S
Цитоплазматическая
мембрана
k0
Y1
PI3K
κ1
k1
Петля
Y2
Akt
отрицательной
k31
обратной
κ2
связи
Петля
положительной
k32
k2
обратной
связи
Y3
mTOR
κ3
Ядерная
мембрана
Рис.
2. Схематическая диаграмма каскада PI3K-Akt-mTOR, содержащая названия участвующих белков и
обозначения петель обратных связей (а) и введенные символьные обозначения (б).
где символом KM обозначена константа Миха-
фазовая кривая после некоторого приближения к
элиса.
одной из нуль-изоклин будет демонстрировать
тенденцию к неограниченному росту обеих пере-
Случай нулевой внешней стимуляции. Деталь-
менных во времени.
ный анализ модели (см. Приложение) показыва-
ет, что в отсутствии внешних стимулов - S ≡ 0 -
Случай ненулевой внешней стимуляции. Для
динамика рассматриваемой системы (уравнения
анализа ситуации в общем случае были введены
(1)-(3)) может быть сведена к случаю Y1 ≡ 0. Си-
безразмерные переменные:
стема становится двумерной и фактически опре-
k1k
2
деляется парой уравнений:
y
=
Y
,
(6)
1
1
К
M
Y
= -û
Y
+k
Y
,
(4)
2
2
2
32
3
k
2
y
2
=
Y
2
,
(7)
û
3
Y
3
Y
=
k
Y
−
(5)
K
M
3
2
2
(1+Y
3
/
K
M
)
1
y
=
Y
(8)
Эволюция такой подсистемы была рассмотре-
3
3
K
M
на с помощью фазового портрета.
Уровень внешней стимуляции отображен ве-
Если условие
выполняется, нуль-изо-
личиной параметра Ω:
клины пересекаются ровно в двух точках, обозна-
k
k
k
S
0
1
2
Ω
=
(9)
ченных на рисунке символами N и S. По характе-
K
M
ру прилегающих к этим особым точкам траекто-
рий видно, что точка N является устойчивым
В обезразмеренных переменных система урав-
узлом, а точка S - седлом, в которое входят две
нений (1)-(3) принимает следующий вид:
особые линии - АS и ВS. Иными словами, об-
y
1
=Ω-y
1
(û
1
+k
31
K
M 3
y
),
(10)
ласть притяжения состояния, которому соответ-
ствует точка N, ограничивается сепаратрисой
y
= y
−û
y
+k
k
y
,
(11)
2
1
2
2
2
32
3
АSB. При попадании изображающей точки с ко-
ординатами (Y2, Y3) в любую точку области вне
û
3
y
3
y
3
=
y
2
−
(12)
этого «бассейна» притяжения соответствующая
1
+
y
3
БИОФИЗИКА том 65
№ 2
2020
АНАЛИЗ УСТОЙЧИВОСТИ СИСТЕМЫ ВНУТРИКЛЕТОЧНОЙ СИГНАЛИЗАЦИИ
303
k32
Y
=
Y
2
3
y2
1
3
A
2
Y3
S
3
0
Y2 =
y2
k
1 + Y /K3M
2
N
0
B
y3
y3
Рис. 3. Фазовый портрет динамической системы (4)-(5). Линии 1 и 2 - нуль-изоклины. Особые точки: N -
устойчивый узел, S - седло. Сепаратриса ASB разделяет два динамических режима в фазовом пространстве системы:
релаксация к устойчивому исходному состоянию и передача сигнала с нарастанием динамических переменных.
Стационарные состояния системы определя-
û
1
ε
=
,
(19)
ются следующей системой уравнений:
k
K
31
M
0
0
0=Ω-y
(
û
+k
K
y
)
,
(13)
1
1
31
M 3
û
û
2
3
δ
=
−
1
>
0.
(20)
0
0
0
k
2
k
3
0= y
1
−û
2
y
2
+k
2
k
32
y
3
,
(14)
При величине σ меньше определенного уровня
0
0
û
3
y
3
0
=
y
−
(15)
σ#, соответствующее значение
принадлежит
2
0
1+
y
3
интервалу [0, δ]. Выражение для σ# может быть
найдено из уравнения (17) и принимает следую-
Легко показать, что у системы уравнений (13)-
(15) существует единственное нетривиальное ре-
щий вид:
шение, только если выполнено следующее усло-
#
⎡x
(
x
+
ε)(δ
−
x)⎤
вие:
σ
=
max
(21)
0<x<δ
⎢
⎥
⎣
1
+
x
⎦
0
Ω
û
û
y
0
2
3
3
−
+
k
k
y
=
0.
(16)
0
0
2
32
3
В случае гиперстимуляции (σ > σ#) система не
(
û
1
+
k
31
K
M 3
y
)
1
+
y
3
имеет положительных стационарных состояний.
Это означает, что если уровень внешней стимуля-
Условие (16) является основополагающим для
дальнейшего анализа. Оно, по сути, является
ции превышает критическое значение σ#, имеет
условием разрешимости системы уравнений
место триггерная активация сигнального пути и
(13)-(15). Оно может быть представлено в следу-
запуск передачи сигнала.
ющем виде:
При любых значениях параметров δ, ε, σ урав-
0
0
0
0
нение (17) задает кубический полином относи-
σ
(
1+ y
3
)
= y
3
(
ε+ y
3
)(
δ-y
3
)
,
(17)
тельно переменной y0. Его решение дает возмож-
3
где использованы следующие обозначения:
0
ность отыскать
, а затемy0 иy0 через уравнения
3
y
1
2
Ω
(14) и (15). Таким образом, перечень всех воз-
σ
=
,
(18)
k
k
k
K
можных стационарных решений системы уравне-
2
31
32
M
БИОФИЗИКА том 65
№ 2
2020
304
САПЕГА, ГУРИЯ
(а)
S
LHS
N
1
0
y0
RHS
(б)
S
LHS
N
1
0
y0
RHS
Рис. 4. Графическое представление решений уравнения (17). Рассмотрены два варианта расположения точек
пересечения: a) ε < 1 и б) ε > 1. В обоих случаях один из корней нефизичен (отрицательное значение концентрации).
Далее при рассмотрении ограничение на параметр ε > 0.
ний (10)-(12) определяется ровно тремя парамет-
Прямое исследование устойчивости стацио-
рами - δ, ε и σ (уравнения (18)-(20)).
нарных состояний, которым отвечают точки пе-
ресечения прямой с кубической параболой, пока-
Левая часть уравнения (17) по отношению
зало, что для всякого σ из интервала 0 < σ < σ# из
к y0 геометрически представляет собой пря-
3
двух точек пересечения прямой и параболы в не-
мую с коэффициентом наклона σ, проходя-
отрицательной области (см. рис. 4) левая - N -
0
всегда отвечает устойчивому узлу, а правая - S -
щую через ось абсцисс при
y
3
= -1. А правая
седлу. Это значит, что при допороговых уровнях
часть
- кубическую параболу с корнями
0
0
0
внешней стимуляции (σ < σ#) система внутрикле-
y
3
=
-ε,
y
3
=
0,
y
3
= δ. Решениям уравнения
точной сигнализации может находиться в жду-
(17) будут отвечать точки пересечения пря-
щем режиме, в котором значению нормирован-
мой и кубической параболы (рис. 4):
ной концентрации белка mTOR отвечает млад-
ший из положительных корней уравнения (17).
0
Поскольку величина
y
, согласно уравнению
3
(8), соответствует перенормированной концен-
Поскольку расстояние между корнями уравне-
трации белка mTOR, она по смыслу не может
ния (17) может рассматриваться как мера величи-
принимать отрицательные значения. Из рис. 4
ны порога динамической активации, самое высо-
видно, что уравнение (17) может иметь не более
кое значение порога достигается при σ = 0. И на-
двух неотрицательных решений (отмечены на ри-
против, при повышении значения σ до
сунке символами N и S). Так, например, при σ = 0
предельного σ# величина порога динамической
0
0
корнями являются
y
=
0
и
y
= δ
потери устойчивости падает до нуля. Стационар-
3
3
ные состояния системы уравнений
(10)-(12)
При увеличении значения σ до момента
сближаются при увеличении параметра σ. Ветви
касания прямой с дугой кубической парабо-
решений, представляющие собой нормирован-
лы при σ = σ# уравнение (17) будет иметь ров-
ные устойчивые (N) и неустойчивые (S) стацио-
но один неотрицательный корень. Также яс-
нарные концентрации белка mTOR, сливаются
но, что при значении параметра σ > σ# прямая
при σ = σ#. Бифуркационная диаграмма выглядит
на рис. 4 будет проходить выше дуги парабо-
следующим образом (см. рис. 5).
лы, следовательно, уравнение (17) не будет
Для заданных параметров δ и ε можно полу-
0
иметь решений всюду при
, т. е. у си-
y
>
0
3
чить числовое значение σ#. Зависимость уровня
стемы уравнений (10)-(12) не будет стацио-
критической стимуляции σ# от двух других пара-
нарных состояний с неотрицательным значе-
метров модели может быть представлена в виде
0
нием
y
графика в трехмерном пространстве (рис. 6).
3
БИОФИЗИКА том 65
№ 2
2020
АНАЛИЗ УСТОЙЧИВОСТИ СИСТЕМЫ ВНУТРИКЛЕТОЧНОЙ СИГНАЛИЗАЦИИ
305
1.0 S
0.8
0.6
0
y
3
0.4
#
#
#
0.2
N
0.0
0
20
40
60
80
100
120
140
Рис. 5. Бифуркационная диаграмма катастрофы типа складка. Ветвь решений, соответствующая траектории
устойчивого узла N, отображена сплошной линией, а седловой точки S - штриховкой. Решения, отображающие
нормированные устойчивую (N) и неустойчивую (S) стационарные концентрации белка mTOR, сливаются при
критическом значении σ = σ#.
РЕЗУЛЬТАТЫ МАТЕМАТИЧЕСКОГО
чена сверх определенного предела (станет
МОДЕЛИРОВАНИЯ
больше значенияy в точке B, показанной на
3
рис. 3), тогда дальнейшая эволюция системы
Из проведенного анализа следует, что си-
во времени не будет представлять собой ре-
стема, описываемая уравнениями (10)-(12),
лаксацию в исходное состояние. Напротив,
демонстрирует отчетливое триггерное пове-
обе динамические переменные и
, и y будут
y
дение. Действительно, представим себе, что
3
2
демонстрировать асимптотическое стремле-
подсистема (11)-(12) находится в своем ос-
0
ние к неограниченному росту. С точки зре-
новном невозмущенном состоянии
y2
= 0,
ния теории управления, это означает, что
0
y
= 0.
Если в некий момент времени одна из
3
рассматриваемая система демонстрирует
ее динамических переменных (возьмем, для
способность к триггерной динамической ак-
определенности, y3) будет импульсно увели-
тивации.
7000
6000
5000
4000
3000
2000
1000
0
100
80
100
60
60
80
40
40
20
20
0
0
Рис. 6. Зависимость критической величины пороговой стимуляции σ# от кинетических параметров модели δ и ε.
БИОФИЗИКА том 65
№ 2
2020
306
САПЕГА, ГУРИЯ
При этом из рис. 3 видно, что импульсная сти-
рения MAPK-каскад. При анализе диаграммы
муляция по любой из переменных, не выводящая
взаимодействия между сигнальными путями (см.
изображающую точку за пределы бассейна при-
рис. 1) с точки зрения теории управления легко
тяжения, завершится асимптотической релакса-
понять, что ведущая роль принадлежит каскаду
цией в исходное стационарное состояние N.
PI3K-Akt-mTOR. Контур самоусиления между
Можно сказать, что область динамической устой-
Akt и mTOR образует автокаталитическое ядро,
чивости основного состояния фактически опре-
которое отвечает за фундаментальную возмож-
деляется размером прилегающего к точке N бас-
ность порогового возбуждения рассматриваемой
сейна притяжения. Его размер характеризуется
системы, в то время как MAPK играет роль до-
расстоянием между особыми точками N и S (дли-
полнительного стимулятора. Авторы исходили из
ной отрезка NS). При изменении кинетических
следующего: при недостаточно выраженной сти-
параметров системы (c2, c3, k2, k3, KM) длина NS
муляции (по амплитуде или длительности воз-
будет меняться, так что при определенных усло-
действия) сигнал может затухнуть вследствие
виях два корня могут выродиться в один. В таких
ограниченного времени жизни белков, диффун-
случаях основное состояние системы испытывает
дирующих к ядру клетки через насыщенный ци-
параметрическую дестабилизацию, становится
тозоль. При прекращении внешней стимуляции
абсолютно неустойчивым, как следствие, при
процессы передачи сигнала определяются кине-
сколь угодно малом возмущении каскад активи-
тикой автокаталитического ядра. Все остальные
руется.
элементы, обозначенные на диаграмме, не могут
работать автономно - их активация зависит от
Порог динамической дестабилизации - вели-
экзогенных стимулов. При допороговом уровне
чина внешнего импульса, необходимого для
внешней стимуляции каскада PI3K-Akt-mTOR
триггерной активации сигнального пути, - сни-
возможна динамическая потеря устойчивости.
жается по мере увеличения параметра σ до пре-
При этом величина динамического порога акти-
дельного значения σ#: уменьшается расстояние
вации зависит от стимулирующего действия со
между точками пересечения прямой и параболы
стороны сигнального пути MAPK: генерируемый
на рис. 4. Это указывает на то, что бассейн притя-
им импульс, может вызывать пороговое возбуж-
жения устойчивого стационарного состояния
дение автокаталитической петли Akt-mTOR. Ана-
сужается.
лиз дополнительной стимуляции сигального пути
PI3K-Akt-mTOR посредством MAPK-каскад
приведен в Приложении.
ДИСКУССИЯ
Cигнальная система PI3K-Akt-mTOR спо-
В настоящей работе анализ устойчивости ста-
собна находиться в метастабильном стационар-
ционарных состояний сигнального пути PI3K-
ном состоянии: при превышении уровнем внеш-
Akt-mTOR основывался на ориентированных
ней стимуляции порогового значения происхо-
графах, которые используют специалисты по мо-
дит запуск автокаталитического ядра каскада и
лекулярной гематологии [18]. С математической
становится возможной автоволновая передача
точки зрения, анализ устойчивости проводился в
сигнала от внешней мембраны к ядру клетки. В
соответствии со стандартными процедурами,
основном, «ждущем» режиме отфильтровывают-
восходящими к работам A.M. Ляпунова [19]. По-
ся шумовые сигналы, стационарные концентра-
лученные результаты поддаются рациональной
ции белков автокаталитического ядра релаксиру-
интерпретации: в частности, заключение о том,
ют к нулю. Из приведенного в работе анализа яс-
что только три истинных параметра (δ, ε, σ) опи-
но, что основное состояние предоставленной
сывают пороговые явления в сложной регулятор-
себе системы (S ≡ 0) может испытывать как дина-
ной системе, динамика которой зависит от девяти
мическую, так и параметрическую потерю устой-
параметров, представляется очень важным. Най-
чивости. В отличие от динамических неустойчи-
денное выражение для величины порога актива-
востей, развивающихся в ответ на мгновенные
ции, а также оценки размеров области притяже-
импульсные изменения динамических перемен-
ния на фазовой плоскости открывают новые воз-
ных, как в случае стимуляции MAPK-каскадом,
можности для интерпретации данных,
параметрические неустойчивости могут быть
представляющих клинический интерес. Можно
следствием плавных и медленных изменений во
предположить, что аналогичные подходы, не пе-
времени каких-либо кинетических параметров
регруженные деталями, могут быть полезны и для
системы. Такие изменения, по мнению авторов,
изучения взаимодействия между другими сиг-
могут иметь место при генетических мутациях,
нальными путями, интегрированными во внут-
сопровождающихся изменением активности бел-
риклеточную сигнальную сеть [15, 20].
ков, что ведет к количественному изменению со-
В данной работе авторы сконцентрировались
ответствующих кинетических параметров систе-
на анализе поведения изолированного пути
мы. Это отображается в рамках модели изменени-
PI3K-Akt-mTOR, оставив за рамками рассмот-
ем бассейна притяжения к основному состоянию
БИОФИЗИКА том 65
№ 2
2020
АНАЛИЗ УСТОЙЧИВОСТИ СИСТЕМЫ ВНУТРИКЛЕТОЧНОЙ СИГНАЛИЗАЦИИ
307
системы. Таким образом, клетки с мутациями в
ПРИЛОЖЕНИЕ
определенных генах могут стать более чувстви-
I. Отсутствие внешней стимуляции. Анализ си-
тельными к экзогенным стимулам, запускающим
стемы уравнений (1)-(3) показывает, что при от-
каскад: они оказываются способными порогово
сутствии сигнала извне (S ≡ 0) рассматриваемая
реагировать на возмущения, которые в норме иг-
система имеет два стационарных состояния. Од-
норируются.
но из них выглядит как
В последнее время число публикаций, касаю-
0
щихся сигнального пути PI3K-Akt-mTOR в свя-
(S1)
Y1
= 0,
зи с процессами активации тромбоцитов, быстро
растет [21, 22]. Опубликованы новые данные об
0
Y
= 0,
(S2)
ингибировании элементов каскада, подавляюще-
2
го адгезию тромбоцитов и образование тромба
[23]. Таким образом, потеря устойчивости сиг-
0
(S3)
Y3
= 0.
нального каскада PI3K-Akt-mTOR может играть
важную роль в процессах тромбообразования.
а второе -
Стоит отметить, что при активации тромбоци-
0
Y
= 0,
тов, наряду с подробно проанализированной в
1
данной работе кинетической системой внутрен-
ней сигнализации с участием PI3K-Akt-mTOR,
0
k
K
⎡û
û
⎤
32
M
2
3
Y
2
=
−1 ,
(S5)
важную роль играет и межклеточная сигнализа-
⎢
⎥
û
k
k
2
⎣
2
3
⎦
ция. Ниже описаны два механизма активации
тромбоцитов, задействующие рассмотренный
0
⎡û
û
⎤
2
3
Y
3
=
K
M
−1 .
(S6)
сигнальный путь.
⎢
⎥
⎣k
2
k
3
⎦
Одним из этих механизмов является прямая
Легко видеть, что второе решение принадле-
стимуляция тромбоцитов протромботическим
0
0
0
агентом: взаимодействие рецептора тромбоксана
жит областиY
1
≥ 0,Y
2
≥ 0,иY
3
≥
0
лишь при вы-
полнении условия
с тромбоксаном А2 приводит к активации рецеп-
тора, связанного с G-белком. Его димерный ком-
û
2
û
3
плекс Gβγ передает сигнал, вызывая активацию
μ
≡
≥
1.
(S7)
субъединиц PI3K и p44 / 42 MAPK (ERK1/2) [24].
k
2
k
3
При этом сигнальные пути МАРK и PI3K-Akt-
Тогда в отсутствие внешнего стимула рассмат-
mTOR участвуют в процессах секреции гранул
риваемая сигнальная система формально может
тромбоцитов, которые выбрасываются с тром-
находиться в одном из двух стационарных состо-
боксаном А2 [7, 25].
яний: либо в состоянии (S1)-(S3), либо в состоя-
нии (S4)-(S6). Для того чтобы ответить, в каком
Другим механизмом является активация «из-
именно состоянии способна реально находиться
нутри наружу»: связывание коллагена с GPVI ре-
цепторами тромбоцитов стимулирует фосфори-
система, надлежит исследовать оба стационар-
лирование специфических остатков тирозина на
ных состояния на устойчивость.
внутренней стороне мембраны, которые в свою
Прямое исследование по первой
«методе»
очередь рекрутируют сигнальные белки, в том
А.М. Ляпунова [19] при выполнении условия (S7)
числе PI3K [26], что ведет к высвобождению
позволяет найти, что стационарное состояние
тромбоксана А2 во внеклеточную среду [7]. Да-
(S1)-(S3) является асимптотически устойчивым
лее, будучи сильным протромботическим веще-
по отношению к малым отклонениям: все три ля-
ством, продуцируемым активированными тром-
пуновских показателя отрицательны. Таким об-
боцитами, тромбоксан А2 стимулирует актива-
разом, решение (S1)-(S3) является узлом в фазо-
цию и агрегацию новых тромбоцитов.
вом пространстве рассматриваемой системы
уравнений (1)-(3). Аналогичное исследование
Инициирование тромбоксана А2, образую-
стационарного решения (S4)-(S6) показывает,
щимся в ходе реакции по второму механизму, ве-
что из трех показателей Ляпунова один является
дет к реализации первого механизма, который
положительным, а два других отрицательны. Это
сам по себе является автокаталитическим. Такое
значит, что данное решение является седлом в
поведение аналогично проанализированному по-
фазовом пространстве, т.е. оно неустойчиво к
ведению каскадов MAPK и PI3K-Akt-mTOR. Та-
сколь угодно малым возмущениям, а потому не
ким образом, активация тромбоцитов существен-
может быть физически реализовано.
но зависит от самоподдерживающихся процес-
сов, в которых важную роль играет динамическая
Кроме того, из анализа изложенного следует,
дестабилизация системы внутриклеточной сиг-
что при полном отсутствии внешней стимуляции
нализации PI3K-Akt-mTOR.
(случай S ≡ 0) динамика рассматриваемого сиг-
БИОФИЗИКА том 65
№ 2
2020
308
САПЕГА, ГУРИЯ
нального пути фактически определяется парой
где Λ - величина, пропорциональная концентра-
уравнений на Y2 и Y3:
ции белка ERK.
С учетом Λ-члена при поиске стационарного
·
Y
= -û
Y
+k
Y
,
(S8)
состояния одна из нуль-изоклин сместится вниз
2
2
2
32
3
на величину Λ:
·
û
3
Y
3
Y
=
k
Y
−
(S9)
3
2
2
û
3
y
3
(1 +Y
/
K
)
y
=
−Λ.
(S13)
3
M
2
1+
y
3
II. Стимуляция mTOR посредством ERK. Как
Выражение для σ принимает следующий вид:
видно из рис. 1, белок ERK способен напрямую
активировать mTOR. Этот эффект может быть
y
(ε
+
y
)(δ
−
y
)
3
3
3
û
2
σ
=
−
Λ(ε+y
3
)
=
формально учтен, если добавить дополнитель-
1+y
3
k
2
k
32
ный член в уравнение (12). Тогда система (10)-
(S14)
y
(ε
+
y
)(
δ
−
y
)
(12) принимает следующий вид:
3
3
3
=
−
λ
(ε+y
3
),
1+y
3
y
=Ω-
y
(û
+
k
K
y
) ,
(S10)
1
1
1
31
M 3
где введен обезразмеренный параметр
˙
û
2
2
1
y = y -û y +
2
2
2
k k y
32
3
,
(S11)
λ
=
Λ.
(S15)
kk
2
32
˙
û
3
y
3
Условие разрешимости принимает следующий
y
=
Λ
+y
−
,
(S12)
3
2
1
+
y
вид:
3
0
0
0
0
0
0
σ
1+y
= y
ε+y
δ-y
− λ
ε+
y
1
+
y
(S16)
(
3
)
3
(
3
)(
3
)
(
3
)(
3
)
Тогда выражение для значения пороговой сти-
СПИСОК ЛИТЕРАТУРЫ
муляции с учетом λ-члена принимает вид:
1. B. Alberts, A. Johnson, J. Lewis, et al., Molecular Biol-
ogy of the Cell (Garland, NY, 2002).
λ
⎡x(x
+
ε)(δ
−
x
)
⎤
σ
=
max
−λ
(ε+x)
(S17)
2. M. J. Berridge, Cell Signalling Biology
(2012).
0<x<δ
⎢
⎥
⎣
1
+
x
⎦
DOI:10.1042/csb0001012
3. Molecular Oncology, Ed. by M. Bishop and R. A. Wein-
Сравнение выражений (21) и (S17) показывает,
berg (Mosby, 1996).
что в присутствии λ-члена (λ > 0) амплитуда кри-
4. N. Dey, P. De, B. Leyland-Jones, PI3K-mTOR in Can-
тического уровня стимуляции σλ меньше σ#. Чем
cer and Cancer Therapy (Humana Press, NY, 2016).
больше значение λ, тем меньше значение σλ. Сле-
5. J. Bertacchini, N. Heidari, L. Mediani, et al., Cell.
довательно, MAPK-каскад способен обеспечить
Mol. Life Sci. 72 (12), 2337 (2015).
параметрическую дестабилизацию сигнального
6. M. Compagno, Q. Wang, C. Pighi, et al., Nature 542
пути PI3K-Akt-mTOR.
(7642), 489 (2017).
7. Z. Li, M. K. Delaney, K. A. O’Brien, and X. Du, Arte-
riosclerosis, Thrombosis, and Vascular Biology 30 (12),
ФИНАНСИРОВАНИЕ РАБОТЫ
2341 (2010).
8. G. F. Guidetti, I. Canobbio, and M. Torti. Adv. Biol.
Работа выполнена при частичной финансовой
Regul. 59, 36 (2015).
поддержке Российского научного фонда (грант
9. A. Martinez-Lopez, L. Salvador-Rodriguez, T. Mon-
№ 19-11-00260).
tero-Vilchez, et al., Curr. Opin. Pediatrics 31 (6), 747
(2019).
КОНФЛИКТ ИНТЕРЕСОВ
way?map=hsa04151&show_description=show.
Авторы заявляют об отсутствии конфликта
11e8-b939-0ac135e8bacf.
интересов.
12. B. Pfeuty and K. Kaneko. Phys. Rev. E 89 (2), 022707
(2014).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
13. М. Ичас, О природе живого: механизмы и смысл
(Мир, М., 1994).
Настоящая работа не содержит описания ка-
14. B. Ø. Palsson, Systems biology: properties of reconstruct-
ких-либо исследований с использованием людей
ed networks (Cambridge University Press, Cambridge,
и животных в качестве объектов.
2006).
БИОФИЗИКА том 65
№ 2
2020
АНАЛИЗ УСТОЙЧИВОСТИ СИСТЕМЫ ВНУТРИКЛЕТОЧНОЙ СИГНАЛИЗАЦИИ
309
15. B. N. Kholodenko, A. Kiyatkin, F. J. Bruggeman,
20. A. Nair, P. Chauhan, B. Saha, and K. F. Kubatzky, Int.
et al., Proc. Nal. Acad. Sci. USA 99 (20), 12841 (2002).
J. Mol. Sci. 20 (13), 3292 (2019).
21. D. S. Shen, Y. J. Yang, X. J. Kong, et al., Eur. J. Phar-
16. Ю. М. Романовский, Н. В. Степанова и Д. С. Чер-
macol. 852, 1 (2019).
навский, Математическая биофизика (Наука, М.,
1975).
22. Z. Chen, T. Li, K. Kareem, et al., Artificial organs 43
(9), 897 (2019).
17. Г. Ю. Ризниченко, Лекции по математическим мо-
23. T. Später, I. Müller, H. Eichler, et al., Platelets 29 (3),
делям в биологии, изд. 2-е, испр. и дополн. (РХД,
277 (2018).
М., 2011).
24. J. M. Gibbins, S. Briddon, A. Shutes, et al., J. Biol.
18. И. А. Якутик, Л. С. Аль-Ради, Б. В. Бидерман и др.,
Chem. 273 (51), 34437 (1998).
Гематология и трансфузиология 63, 112 (2018).
25. P. Flevaris, Z. Li, G. Zhang, et al., Blood 113 (4), 893
19. А. М. Ляпунов, Общая задача об устойчивости дви-
(2009).
жения (АН СССР, М., 1948).
26. N. Nakahata, Pharmacol. Therap. 118 (1), 18 (2008).
Stability Analysis of PI3K-Akt-mTOR Intracellular Signaling Pathway
T.S. Sapega*, ** and G.Th. Guria*, **
*National Research Center for Hematology, Novozykovskii proezd 4, Moscow, 125167 Russia
**Moscow Institute of Physics and Technology, Institutskiy per. 9, Dolgoprudny, Moscow Region, 141700 Russia
The MAPK and PI3K-Akt-mTOR intracellular signaling cascades regulate important cellular processes. A
mathematical model was designed and used for the analysis based on current knowledge of the interactions
that occur within these cascades. Stability analysis of the PI3K-Akt-mTOR signaling pathway model
showed that the ground state of the signaling pathway is metastable and signal transduction can be activated
by overcritical exogenous stimulation. An explicit expression was obtained for the threshold level of external
stimuli. It was shown that triggering events in the dynamics of the signaling pathway are described by the Fold
catastrophe of the Elementary theory. A discussion about whether the results obtained are significant for mo-
lecular hematology and hemostasiology is presented.
Keywords: PI3K-Akt-mTOR, MAPK, intracellular signaling, stability analysis, platelet activation
БИОФИЗИКА том 65
№ 2
2020