БИОФИЗИКА, 2020, том 65, № 2, с. 390-401
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 577.38:574.34
ОПРАВДАЛСЯ ЛИ ДОЛГОСРОЧНЫЙ ПРОГНОЗ РИСКА ВЫМИРАНИЯ
АЗОВСКОГО СУДАКА?
© 2020 г. Ю.В. Тютюнов*, **, И.Н. Сенина***, Л.И. Титова**, Л.В. Дашкевич*
*Федеральный исследовательский центр «Южный научный центр Российской академии наук»,
344006, Ростов-на-Дону, просп. Чехова, 41
**Южный федеральный университет, 344090, Ростов-на-Дону, просп. Стачки, 200/1
***Collecte Localisation Satellites, rue Hermes 8-10, Ramonville Saint-Agne, 31520 France
E-mail: yuri.tyutyunov@yandex.ru
Поступила в редакцию 08.08.2019 г.
После доработки 19.11.2019 г.
Принята к публикации 24.12.2019 г.
В конце 1990-х годов на основе стохастических экспериментов, проведенных с моделью долгосроч-
ного прогноза популяционной динамики азовского судака, идентифицированной по данным на-
блюдений 1950-1981 гг. за запасом, выловами, температурой и соленостью Азовского моря, был
сделан вывод о высокой вероятности квазивымирания популяции азовского судака в течение 35 лет,
c 1982 по 2016 гг., обусловленного повышением солености моря. Данный вероятностный
прогноз подтвердился — событие риска произошло в 2000-х годах: с 2005 г. численность взрослых
особей судака не превышала 4 млн штук, а с 2017 г. введен полный запрет на вылов этого вида. Ва-
лидация результатов работы модели, проведенная в настоящее время, выявила достаточно хорошее
(качественное и количественное) соответствие модельной траектории данным наблюдений за
1982-2018 гг. При этом обнаружено, что в современный период негативное воздействие, оказывае-
мое на динамику численности стада повышением среднегодовых значений температуры моря, яв-
ляется не менее значительным, чем влияние роста солености. В соответствии с краткосрочным мо-
дельным прогнозом на 2019-2020 гг., в сложившихся условиях значительных положительных ано-
малий температуры и солености, введенный в 2017 г. запрет промысла не позволяет остановить
снижение запаса азовского судака в ближайшие годы.
Ключевые слова: Азовское море, популяционная динамика, Sander Lucioperca L., стохастическая модель,
метод Монте-Карло, риск квазивымирания.
DOI: 10.31857/S0006302920020234
В конце 1990-х годов нами была построена
Кубани в 1972 г. Уменьшение пресного стока,
имитационная модель динамики эксплуатируе-
увеличение затока (объема) черноморских вод
мой рыбной популяции, использованная для
и рост солености Азовского моря привели к су-
проведения стохастических экспериментов в це-
щественному сокращению нерестовых и нагуль-
лях долгосрочного прогноза изменений числен-
ных ареалов судака, леща, тарани, рыбца, чехони
ности и оценки риска квазивымирания популя-
[3, 4].
ции азовского судака (Sander lucioperca L.) [1, 2].
Модель имела достаточно простую структуру и
Выбор судака в качестве модельного объекта обу-
работала с годовым шагом по времени, описывая
славливался тем, что это один из ценных видов,
влияние на воспроизводство стада таких факто-
запасы и уловы которого значительно снизились
ров среды, как температура и соленость Азовско-
после зарегулирования в 1950-х годах Дона, круп-
го моря. Основной задачей данного исследования
нейшей реки бассейна Азовского моря. Наблюда-
являлось обоснование возможных ограничений
ющееся с 1960-х годов падение численности
промыслового изъятия, потенциально способных
популяции судака является типичным послед-
предотвратить чрезмерное падение запасов азов-
ствием широкомасштабного антропогенного
ского судака, обусловленное продолжающимся
преобразования экосистемы Азовского моря -
осолонением моря. Кроме того, ихтиологов инте-
сходные проблемы имеют место и для других або-
ресовал вопрос, насколько опасным для попу-
ригенных полупроходных видов. Дальнейшее
ляции является непреднамеренный прилов го-
ухудшение условий воспроизводства полупро-
довиков, обусловленный недостаточной селек-
ходных рыб произошло после зарегулирования
тивностью сетей. Параметры модели были
390
ОПРАВДАЛСЯ ЛИ ДОЛГОСРОЧНЫЙ ПРОГНОЗ
391
идентифицированы с использованием предо-
использованного подхода, а также предлагается
ставленных Азовским НИИ рыбного хозяйства
минимальная версия математической модели,
данных, образующих временные ряды, характе-
корректно отражающая влияние внешних факто-
ризующие динамику моделируемой экологиче-
ров на воспроизводство и выживаемость азовско-
ской системы с 1950 по 1981 гг. (т. е. 32 осреднен-
го судака и наилучшим образом объясняющая ва-
ные по годовому периоду записи). Ряды включа-
риации численности стада.
ли оценки численностей сеголеток, годовиков и
взрослых особей судака, среднегодовые показате-
ОПИСАНИЕ МОДЕЛИ
ли солености и температуры Азовского моря. С
И РЕЗУЛЬТАТЫ ПРОГНОЗА
идентифицированной моделью проводили ими-
тационные эксперименты по методу Монте-Кар-
Модель представляет собой систему разност-
ло, в которых для сгенерированных случайных
ных уравнений, описывающую динамику трех
значений температуры и солености моря был рас-
t
t
возрастных групп: сеголеток (N
0
, годовиков
N
1
и
считан прогноз динамики численностей возраст-
взрослых особей, достигших возраста в два года и
ных групп судака на 35 лет вперед, т. е. c 1982 по
2016 гг. Пучки траекторий, полученные таким об-
старше (Nt с годовым шагом t = 0,1,2, …:
2
разом для различных стратегий промыслового
2
2
t +
1
t +1
⎛S
−S
0
⎞
⎛T
−T
0
⎞
изъятия, использовались для оценки ожидаемых
t
t
−⎜
⎟
-⎜
⎟
−b
(
N
1
+N
2
)
σ
σ
t+1
t
t
⎝
S
⎠
⎝
T
⎠
(1)
уловов, размеров стада, а также вероятностей со-
N
0
=
(
a
1
N
1
+
a
2
N
2
)
e
e
e
,
бытия квазивымирания (quasi-extinction) популя-
t+
1
t
t+1
ции [5, 6]. На основе проведенных эксперимен-
N
=
p
N
1−
h
,
(2)
1
0
0
(
1
)
тов с критическим уровнем квазивымирания,
t
+1
t
t
t+1
равным 4.8 млн взрослых особей, что соответ-
N
2
=
(
p
1
N
1
+
p
2
N
2
)(
1−
h
2
)
,
(3)
ствует 10% от максимального значения в ряду их-
тиологических оценок запаса, нами был сделан
где pi - доля выживших в течение временного ша-
достаточно пессимистично звучащий вывод о вы-
t+
1
га особей i-й возрастной группы (t = 0,1,2), а
h
i
-
сокой вероятности коллапса популяции азовско-
доля вылавливаемых в течение временного шага
го судака в течение 35 лет, c 1982 по 2016 гг. [1, 2].
(t, t + 1) особей каждой возрастной группы. Вы-
А именно, согласно модельным расчетам, даже
жившие в течение года сеголетки переходят в воз-
при гипотетическом прекращении промысла
растной класс годовиков, а выжившие годовики
риск квазивымирания популяции превышал 70%.
пополняют класс взрослых особей популяции.
При ежегодном промысловом изъятии третьей
Суммарный улов, полученный в течение года t,
части взрослых особей прогнозируемый риск
вычисляется как
квазивымирания азовского судака составлял 86%.
Целью настоящей работы является ретроспек-
t
t
t−1
t
t
−
1
t−1
C
=
w
h
p
N
+
w
h
p
N
+
p
N
,
1 1
0
0
2
2
(
1
1
2
2
)
тивная валидация корректности данного вероят-
ностного прогноза долгосрочной динамики по-
где w1 и w2 - средние индивидуальные веса годо-
пуляции судака Азовского моря с использовани-
вика и взрослой особи судака соответственно.
ем данных наблюдений за 1982-2018 гг. Статья
Как видно из уравнения (1), воспроизводство
организована следующим образом. Вначале дано
описывается модифицированной функцией Рик-
краткое описание математической модели и ис-
кера [7]. Параметры a1 и a2 - соответственно ко-
пользованных для ее идентификации исходных
эффициенты плодовитости годовиков и взрослых
данных, представлены значения идентифициро-
особей; b - коэффициент внутривидовой конку-
ванных параметров, а также ранее полученные на
ренции. В соответствии с законом толерантности
основе этой модели результаты стохастических
Шелфорда
[8-10] влияние на выживаемость
экспериментов. Затем приведены и обсуждены
мальков случайных факторов среды - среднего-
результаты валидации модели по данным наблю-
довых значений солености и температуры моря -
дений за запасами популяции судака и значения-
учтено в уравнении (1) в виде множителей, имею-
ми учитываемых в модели абиотических факто-
щих вид функции распределения Гаусса, где S0 и
ров. В частности, показано, что падение числен-
T0 - оптимальные значения факторов, а σS и σT
ности популяции азовского судака, имевшее
место в начале 2000-х годов, не может быть объяс-
характеризуют толерантность вида к флуктуаци-
нено осолонением Азовского моря. Проверка ги-
ям соответствующих характеристик среды [1].
потезы о критическом влиянии на воспроизвод-
Этот подход был нами ранее успешно использо-
ван при моделировании воспроизводства конку-
ство судака климатических изменений (роста
среднегодовой температуры моря) потребовала
рирующих пелагических видов - азовской тюль-
проведения дополнительных имитационных экс-
ки и азово-черноморской хамсы [11].
периментов с моделью. В заключительной части
Идентифицированные параметры. Простота
статьи обсуждаются достоинства и недостатки
структуры и небольшое количество параметров
БИОФИЗИКА том 65
№ 2
2020
392
ТЮТЮНОВ и др.
Таблица 1. Идентифицированные значения параметров модели (1)-(3)
Параметр
Регрессионные оценки
Результаты бутстрэппинга, m ± σ
Плодовитость годовиков (a1)
6.82
6.99 ± 4.53
Плодовитость взрослых (a2)
12.67
14.35 ± 4.99
Внутривидовая конкуренция (b)
0.039
0.041 ± 0.01
Выживаемость сеголеток (p0)
0.54
0.48 ± 0.08
Выживаемость годовиков (p1)
0.47
0.52 ± 0.09
Выживаемость взрослых (p2)
0.62
Толерантность к солености (σS)
1.26
1.27 ± 0.19
Толерантность к температуре (σT)
1.92
1.91 ± 0.27
Оптимальная соленость (S0)
11.0
Оптимальная температура (T0)
10.0
обеспечили приемлемую точность идентифика-
независимых стохастических траекторий модель-
ции модели на основе имевшихся на момент ее
ного прогноза на указанный период.
создания данных наблюдений за период 1950-
Затем результаты имитационных вычислений
1981 гг. (начальный год t = 0 в расчетах соответ-
анализировали статистическими методами и ин-
ствовал 1949 г.). На этапе идентификации ис-
терпретировали в контексте задачи оптимизации
пользовали численности возрастных групп, вели-
экономического и экологического критериев:
чины уловов, а также среднегодовые значения
максимизации среднегодовых уловов и миними-
определяющих успех воспроизводства судака
зации риска чрезмерного падения запаса судака -
внешних факторов - солености и температуры
квазивымирания [5]. Критический порог квази-
моря. Коэффициент выживаемости взрослых
вымирания популяции был принят равным
особей p2 и оптимальные значения внешних фак-
4.8 млн взрослых особей, что соответствует 10%
торов S0 и T0 (обеспечивающие наилучшие усло-
от максимального значения в ряду ихтиологиче-
вия для воспроизводства) взяты из ихтиологиче-
ских оценок запаса. Оказалось, что ограничение
промысла в модели не позволяет существенно
ских исследований [12-14]. Остальные парамет-
снизить риск квазивымирания азовского судака
ры модели были идентифицированы путем
[1, 2]. В соответствии с изоплетной диаграммой,
приближения модельной траектории к наблю-
приведенной на рис. 1, если при изъятии третьей
денным значениям численностей возрастных
части взрослых особей их численность в течение
групп (табл. 1). Ошибки полученных значений
35 лет снижается ниже критического уровня с ве-
модельных параметров были оценены с исполь-
роятностью 86%, то полный запрет вылова умень-
зованием процедуры бутстрэппинга (residual boot-
шает данный показатель риска лишь до 70%. При
strap), описанной в работе [15]. Детали идентифи-
этом непреднамеренный прилов годовиков ока-
кации представлены в работе [2].
зался не так опасен для популяции, как чрезмер-
Стохастические имитационные эксперименты.
ный вылов взрослых особей. Причиной прогно-
зируемого коллапса популяции в имитационных
На следующем этапе идентифицированная мо-
дель была использована для проведения серии
расчетах являлось увеличение солености моря,
предсказываемое статистической регрессионной
стохастических имитационных экспериментов
моделью Бронфмана и Суркова [16], которую ис-
по методу Монте-Карло на 35-летний период (с
пользовали для генерации рядов случайных зна-
1982 по 2016 гг.). Случайные значения временных
рядов солености моря для этих экспериментов
чений {St} в экспериментах Монте-Карло.
вычислялись на основе имитационного алгорит-
Сегодня можно констатировать, что данный
ма, предложенного в работах [16, 17]. Входные ря-
нами вероятностный прогноз, к сожалению,
ды температурных данных генерировались мето-
оправдался - событие риска произошло в середи-
дом рандомизации значений температуры моря,
не 2000-х годов, c 2005 г. оценка численности
которые наблюдались в течение 1950-1981 гг.
взрослых особей судака не превышала 4 млн штук
С этими случайными факторами и для различных
[18]. Более того, запрет промысла азовского суда-
комбинаций параметров hi, характеризующих го-
ка, который в конце 1990-х годов рассматривался
довое промысловое изъятие годовиков и взрос-
лишь как гипотетический сценарий имитацион-
лых особей судака, были рассчитаны пучки 50-ти
ного эксперимента, сегодня также стал реально-
БИОФИЗИКА том 65
№ 2
2020
ОПРАВДАЛСЯ ЛИ ДОЛГОСРОЧНЫЙ ПРОГНОЗ
393
0.7
0.85
0.6
0.83
0.5
0.80
0.4
0.99
0.78
0.3
0.97
0.74
0.2
0.95
0.72
0.93
0.1
0.89
0.87
0.91
0.70
0.0
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Доля промыслового изъятия взрослых особей
Рис. 1. Изоплетная диаграмма риска квазивымирания популяции (вероятности падения численности взрослых
особей ниже заданного критического уровня) с 1982 по 2016 гг. как функции коэффициентов промыслового изъятия
годовиков и взрослых.
стью - с 2017 года правилами рыболовства для
ке популяционных запасов азовского судака.
Азово-Черноморского рыбохозяйственного бас-
Нами использованы материалы опубликованных
сейна введен полный запрет вылова этого вида
результатов ихтиологических исследований
[19, 20]. Изъятие азовского судака допускается
[18, 21-30], а также Протоколов XIX-XXX сессий
только для целей воспроизводства и/или научно-
Российско-Украинской Комиссии (РУК) по
исследовательских работ по заявкам воспроиз-
вопросам рыболовства в Азовском море [19, 20,
водственных и научных организаций. Аналогич-
31-36].
ный запрет введен для берша и чехони [19, 20].
На вход модели (1)-(3) подавали временные
Заметим, что модель использовали не для рас-
ряды фактических среднегодовых значений соле-
чета одной единственной траектории, которая ед-
ности и температуры Азовского моря с 1982 по
ва ли была полезной для долгосрочного прогноза
2018 гг. [37]. При этом среднюю годовую темпера-
популяционной динамики, определяемой внеш-
туру воды для моря в целом рассчитывали соглас-
ними факторами, имеющими существенную слу-
но работе [38]. Наборы срочных данных наблюде-
чайную составляющую. Целью исследования был
ний (температуры воды) гидрометеорологиче-
именно вероятностный прогноз: при различных
ских станций были усреднены посуточно,
промысловых нагрузках, на основе достаточно
помесячно и далее до среднегодовых значений.
большого количества стохастических экспери-
Среднегодовая соленость моря была рассчитана
ментов с идентифицированной моделью оцени-
на основе данных экспедиционных наблюдений в
вали риски такого нежелательного события, как
открытом море и данных прибрежных гидроме-
падение численности взрослых особей судака ни-
теорологических станций согласно методике, по-
же установленного критического уровня квази-
дробно описанной в работе [39].
вымирания.
Поскольку для 2019-2020 гг. данных фактиче-
Валидация модели и краткосрочный прогноз.
ских измерений температуры воды и солености
Спустя двадцать лет после проведения описан-
ных выше имитационных экспериментов у нас
пока нет, для последних двух лет периода прогно-
появилась возможность валидации работы моде-
за были использованы значения, полученные с
ли (1)-(3) с ранее идентифицированными пара-
помощью боксовой гидродинамической модели
метрами путем расчета динамики численности
Бердникова [40], включающей модули расчета
возрастных групп азовского судака для фактиче-
температурного и ледового режимов, балансовый
ских (а не разыгрываемых моделью) значений
модуль водного обмена и режима солености. Мо-
внешних факторов в период с 1982 по 2018 гг. и
дельные значения солености на 2019 и 2020 гг.
сопоставления данного модельного прогноза с
равны 11.78 и 11.71‰, а температуры - 12.61 и
обновленными данными о многолетней динами-
12.55°С соответственно.
БИОФИЗИКА том 65
№ 2
2020
394
ТЮТЮНОВ и др.
(а)
120
Данные
Модель
100
80
60
40
20
0
1950
196 0
197 0
198 0
199 0
2000
2010
2020
(б)
60
Данные
50
Модель
40
30
20
10
0
1950
196 0
197 0
198 0
199 0
2000
2010
2020
(в)
50
По данным Белоусова
По данным Подойницына
40
По данным протоколов РУК
30
20
10
0
1950
196 0
197 0
198 0
199 0
2000
2010
2020
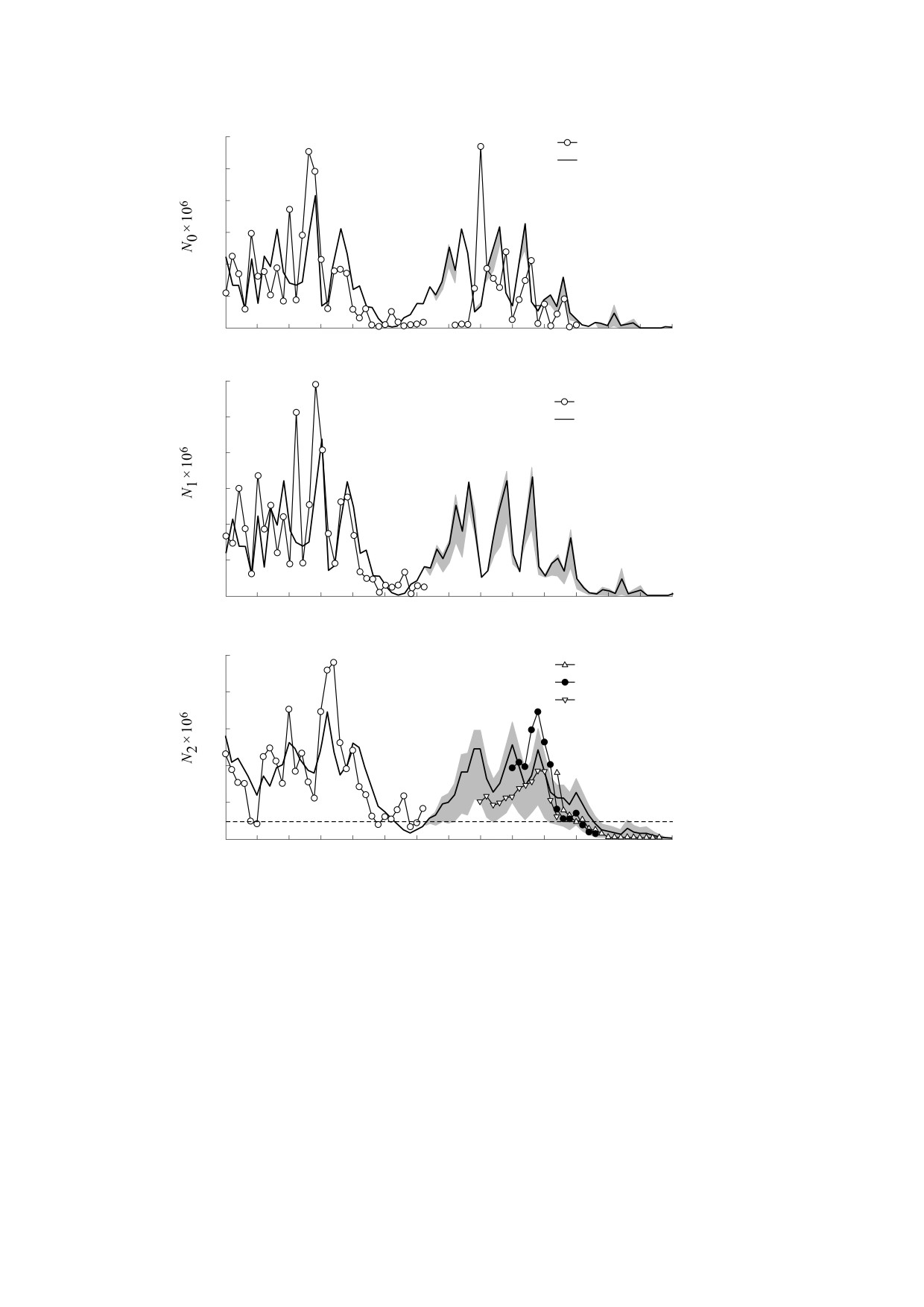
Рис. 2. Модельная и наблюдаемая динамика численности сеголеток (а), годовиков (б) и взрослых особей (в) азовского
судака (млн шт.): 1950-1981 гг. - идентификация; с 1982 г. - валидация модели с коэффициентом изъятия h = 0.07 в
1982-2016 гг. и запретом промысла с 2017 г. Серым цветом показан разброс прогнозируемых численностей возрастных
групп, соответствующий интервалу значений коэффициента промысла 0 ≤ h ≤ 0.3. Пунктиром показан уровень
квазивымирания.
На рис. 2 представлены результаты расчетов,
ловов. Однако фактический промысел мог быть
при проведении которых предполагалось, что до
значительно выше официального [18, 23, 24]. По
введения запрета на промысел судака в 2017 г., с
мнению ихтиологов объем незаконного (соответ-
1982 по 2016 гг. ежегодно вылавливается 7% годо-
ственно, неучтенного) вылова азовского судака
виков и взрослых особей, что равно среднему зна-
всегда был существенным и сопоставимым с ле-
чению коэффициента промыслового изъятия в
гальным промыслом, а после распада СССР доля
1950-1981 гг. Заметим, что значения коэффици-
браконьерского изъятия значительно увеличи-
ента промыслового изъятия в указанные годы вы-
лась [18, 23, 24, 28]. При идентификации модели
числяли на основе официальной статистики вы-
неучтенная промысловая смертность автомати-
БИОФИЗИКА том 65
№ 2
2020
ОПРАВДАЛСЯ ЛИ ДОЛГОСРОЧНЫЙ ПРОГНОЗ
395
чески компенсировалась значениями других па-
тирующим фактором для популяции судака [18]
раметров, прежде всего значениями коэффици-
(соленость моря резко повышается лишь после
ентов выживаемости pi, что обеспечило хорошее
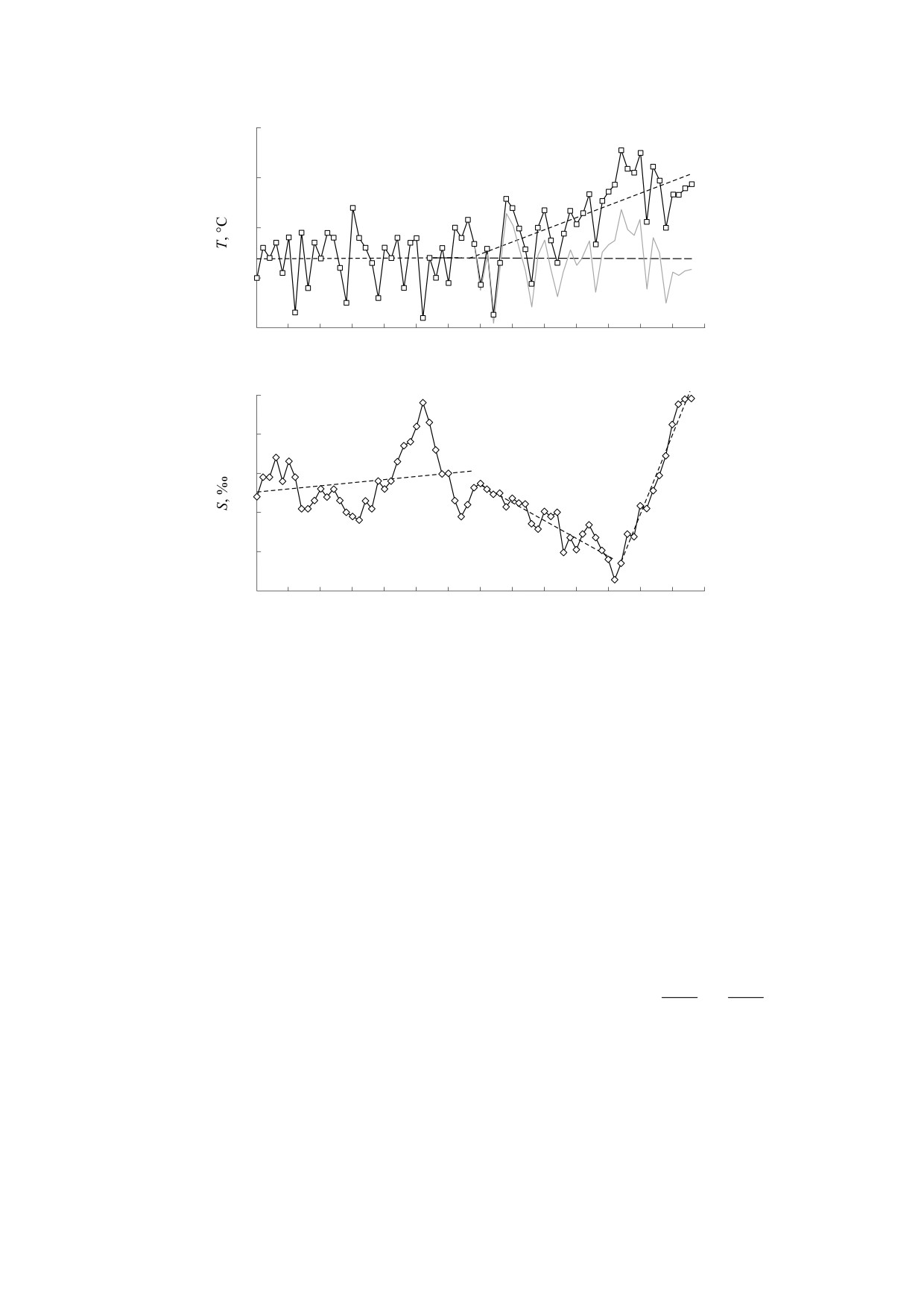
2010 г., см. график на рис. 3).
приближение модельной траектории к использо-
Простота имитационной модели (1)-(3), ко-
ванным при идентификации модели оценкам
торая учитывает минимальное количество внеш-
численности возрастных групп судака (см. иден-
них факторов, оказывающих влияние на популя-
тификационный участок траектории на рис. 2,
ционную динамику, позволяет предположить,
соответствующий периоду с 1950 по 1981 гг.). При
что отмеченное снижение численности судака
отсутствии достоверной статистики реального
может быть вызвано изменениями температуры
промысла в период валидации модели представ-
моря. Действительно, на графике среднегодовых
ляет интерес оценка чувствительности модели к
значений температуры в 1983-2018 гг., изобра-
вариациям параметра промыслового изъятия h.
женном на рис. 3, хорошо виден восходящий
Для этого наряду с модельной траекторией, полу-
тренд, который, по-видимому, связан с глобаль-
ченной при фиксированном коэффициенте изъя-
ными климатическими изменениями. Аномаль-
тия h = 0.07, на рис. 2 представлены результаты
но высокие температуры в таком мелком водое-
дополнительных вычислительных эксперимен-
ме, как Азовское море, являются причиной воз-
тов в виде коридора значений переменных моде-
никновения зон гипоксии, частота образования
ли при варьировании коэффициента изъятия в
которых увеличилась в последнее десятилетие, в
широких пределах (h ∈ [0.0, 0.3]).
том числе, в восточной части моря и Таганрог-
ском заливе [43, 44]. Формирование зон с дефи-
Выполненные вычисления показывают доста-
цитом кислорода оказывает как прямое воздей-
точно хорошее соответствие модельных траекто-
ствие на выживаемость судака, особенно его мо-
рий данным наблюдений в 1982-2018 гг. [18, 21-
лоди, так и опосредованное, вследствие подрыва
30, 41]. Наибольшее отклонение наблюдается в
кормовой базы вида.
1990 г. для численности сеголеток. Однако имен-
но в этом году ихтиологами отмечалась аномаль-
Дополнительные эксперименты и минимальная
но высокая выживаемость сеголеток азовского
модель. Преимуществом модели (1)-(3) является
судака в летний период, которую они объясняли
возможность учета эффекта воздействия темпе-
снижением количества тюльки, в свою очередь
ратуры моря на выживаемость сеголеток при по-
мощи всего лишь двух параметров: оптимального
обусловленным вселением в Азовское море греб-
невика Mnemiopsis leidyi [21, 42]. В результате в
значения температуры T0 и характеристики толе-
июле 1990 г. выживших сеголеток судака остава-
рантности вида к отклонениям от этого значения
лось больше 100 млн шт., что в четыре-пять раз
σT. Чтобы проверить гипотезу о влиянии роста
превышало их количество в предыдущие годы.
температуры на снижение запасов судака, был
Однако уже в сентябре-октябре численность се-
проведен дополнительный имитационный экс-
голеток упала до 10 млн шт. [21, 42], что вполне
перимент, в котором использовали очищенный
соответствует прогнозу нашей модели, которая не
от линейного тренда температурный ряд (показан
включает в себя описание межвидовых отноше-
серым цветом на рис. 3). При этом целесообразно
ний и, работая с годовым шагом, не предназначе-
зафиксировать промысловое изъятие, поэтому, как
на для предсказания подобных краткосрочных
и в предыдущем эксперименте, с 1982 г. были ис-
аномалий. Вот что пишет об этих наблюдениях
пользованы усредненные коэффициенты промыс-
Л.В. Кукарина: «Выживание сеголеток за период
лового изъятия h1 = h2 = 0.07, а с 2017 г. вылов в мо-
«лето-осень» в 1989-1994 гг. составляло 2-25%
дели не осуществлялся. Результаты вычислений
их численности в июле. Высокий процент гибели
представлены на рис. 4.
подросших сеголеток в сложившихся условиях
Как можно видеть, устранение растущего
был обусловлен развитием у судака внутривидо-
тренда в динамике среднегодовой температуры
вой пищевой конкуренции на акватории Таган-
позволяет восстановить численность взрослых
рогского залива. Доля молоди собственного вида
особей до уровня, превышающего критический
в питании увеличилась с 0.2-0.4% до 4-6% в на-
порог квазивымирания популяции в течение все-
чале 1990-х годов» [42].
го валидационного периода с 1982 по 2018 гг. Вме-
Модельные расчеты корректно отображают
сте с тем, согласно расчетам, даже устранение не-
наметившееся в период 1982-2000 гг. восстанов-
гативного воздействия температурного фактора и
ление популяции судака, обусловленное наблю-
прекращение вылова в 2017 г. не позволяет пере-
давшимся в данный период опреснением Азов-
ломить обусловленную аномально высокими
ского моря, однако с начала 2000-х годов начина-
значениями солености моря тенденцию к паде-
ется новая фаза снижения численностей всех трех
нию запаса судака, наблюдающуюся в модели по-
возрастных групп. Падение запасов в этот период
сле небольшого прироста численности взрослых
на первый взгляд выглядит неожиданным, по-
особей в 2013 г. Так, в 2019 и 2020 гг. модель без
скольку в эти годы соленость не являлась лими-
температурного тренда также предсказывает па-
БИОФИЗИКА том 65
№ 2
2020
396
ТЮТЮНОВ и др.
(а)
14
13
12
11
10
1950
1960
1970
1980
1990
2000
2010
2020
Года
(б)
14
13
12
11
10
9
1950
1960
1970
1980
1990
2000
2010
2020
Года
Рис. 3. Среднегодовые значения наблюдений температуры (а) и солености (б) Азовского моря в период 1950-2018 гг.
Пунктирными линиями на графике температуры показаны многолетние тренды, соответствующие периодам 1950-
1982 гг. (y = 0.0007x + 11.38) и 1983-2018 гг. (y = 0.049x + 9.73). Серым цветом показан ряд температур с нулевым
трендом (y = 11.38). Для солености выделены три периода с диаметрально меняющимся трендом: 1950-1984 гг.
(y = 0.016x + 11.52), 1985-2006 гг. (y = -0.09x + 15) и 2007-2018 гг. (y = 0.4x - 13.7).
дение численности половозрелого стада ниже по-
(3), то есть параметров a1, b, p0, h0. Данное обсто-
рога квазивымирания, установленного при про-
ятельство, в частности, объясняет довольно боль-
ведении стохастических экспериментов.
шую ошибку, полученную методом бутстрэппин-
га для параметра a1 (см. табл. 1). Следовательно,
Еще одна гипотеза, которую мы проверили,
связана со структурной устойчивостью мо-
мы можем упростить модель (1)-(3), опуская
дели (1)-(3) - свойством, характеризующим за-
уравнение (2) без ущерба качеству модели в пред-
висимость прогноза модели от изменения ее
сказании динамики популяции судака.
структуры. Нетрудно заметить (см. рис. 2), что
Модель (1)-(3) при этом трансформируется в
временные ряды наблюденных значений сеголе-
следующую минимальную модель с двумя пере-
ток и годовиков обладают высокой степенью кор-
t
t
менными,
N
0
- численность сеголеток и
N
1
-
реляции. Так, для смещенных на один год сеголе-
численность взрослых особей старше одного года
t+1
t
ток
N
0
и годовиков
N
1
коэффициент корреля-
соответственно:
ции r = 0.96. Фактически это означает, что
t +1
t +1
⎛S
−S
0
⎞
⎛T
−T
0
⎞
использование данных по численности годови-
−⎜
⎟
-⎜
⎟
t+1
t
−bN
t
⎝
σ
S
⎠2
⎝
σ
T
⎠2
ков не добавляет информации о динамике попу-
1
N
0
=
aN
1
e
e
e
,
(4)
ляции при минимизации целевой функции (сум-
мы квадратов разностей натурных и модельных
t+1
t
t
t+
1
N
1
=
(
p
0
N
0
+
p
1
N
1
)(
1−
h
)
,
(5)
значений численностей возрастных групп). Сле-
довательно, использование этого временного ря-
где a - плодовитость, b - внутривидовая конку-
да не только усложняет модель, но и снижает точ-
ренция, p1 - выживаемость взрослых особей и h -
ность оценки параметров, контролирующих вли-
промысловое усилие на взрослое стадо. Осталь-
t
яние переменной
N
на динамику системы (1)-
ные параметры те же, что и в модели (1)-(3).
1
БИОФИЗИКА том 65
№ 2
2020
ОПРАВДАЛСЯ ЛИ ДОЛГОСРОЧНЫЙ ПРОГНОЗ
397
По данным Белоусова
По данным Подойницына
50
По данным протоколов РУК
Температура без тренда
40
30
20
10
0
1950
196 0
197 0
198 0
199 0
2000
2010
2020
Года
Рис. 4. Динамика численности взрослых особей азовского судака, рассчитанная по модели (1)-(3) с рядом наблюденных и
очищенных от восходящего тренда температур в сравнении с ихтиологическими оценками размера промыслового стада.
Как видно, по сравнению с системой (1)-(3) мо-
(см. рис. 5). Как и для модели (1)-(3), для двух по-
дель (4)-(5) включает меньше параметров, при
следних лет, соответствующих периоду кратко-
этом все они могут быть довольно точно оценены
срочного прогноза на 2019 и 2020 гг., нами были
в результате минимизации целевой функции (см.
использованы приведенные в предыдущем разде-
табл. 2). В частности, значение параметра p2 было
ле значения солености и температуры, вычислен-
зафиксировано в модели (1)-(3), теперь для
ные с помощью внешней гидродинамической
коэффициента выживаемости взрослых особей
модели Бердникова [40].
старше одного года получена оценка с ошибкой
Помимо результатов валидации и прогноза на
28% (см. табл. 2 и соответствующее ему p1 в моде-
1982-2020 гг., рассчитанных по модели (4)-(5) со
ли (4)-(5)), оцененной методом бутстрэппинга,
значением h = 0.07 (траектории черного цвета),
который подробно описан в работе [2].
на рис. 5 показан разброс прогнозируемых значе-
ний численностей возрастных групп азовского
Как и ожидалось, и полная, и минимальная
модель дают практически одинаковые приближе-
судака, соответствующий интервалу значений ко-
ния временных рядов численностей сеголеток и
эффициента промыслового изъятия 0 ≤ h ≤ 0.3.
взрослых рыб старше одного года: минимальные
Для сравнения и демонстрации структурной
значения целевых функций, включающих в себя
устойчивости модели тонкими линиями изобра-
эти переменные, равны 16608 и 16618 соответ-
жены траектории, соответствующие прогнозу
ственно. При этом модель (1)-(3) объясняет 50 и
полной модели (1)-(3). Также на рис. 5б показана
66% вариаций данных по численности сеголеток
динамика численности взрослых особей, рассчи-
и взрослых соответственно, минимальная
танная по обеим моделям (4)-(5) и (1)-(3) (соот-
модель (4)-(5) - 52 и 66% соответственно. Пред-
ветственно жирная и тонкая линии серого цвета)
сказания модели (4)-(5) согласуются с результа-
для очищенного от восходящего тренда ряда тем-
тами модели (1)-(3) как в период минимизации
ператур. Заметим, что увеличение суммарной
ошибки модельных предсказаний, т.е.
1950-
численности годовиков и взрослых в 2020 г. в
1981 гг., так и в период валидации в 1982-2018 гг.
предполагаемом сценарии стабильного темпера-
Таблица 2. Идентифицированные значения параметров минимальной модели (4)-(5)
Параметр
Регрессионные оценки
Результаты бутстрэппинга, m ± σ
Плодовитость (a)
9.49
10.79 ± 3.64
Внутривидовая конкуренция (b)
0.04
0.04 ± 0.009
Выживаемость сеголеток (p0)
0.63
0.62 ± 0.14
Выживаемость взрослых (p1)
0.49
0.48 ± 0.14
Толерантность к солености (σS)
1.27
1.22 ± 0.15
Толерантность к температуре (σT)
2.22
2.28 ± 0.35
Оптимальная соленость (S0)
11.0
Оптимальная температура (T0)
10.0
БИОФИЗИКА том 65
№ 2
2020
398
ТЮТЮНОВ и др.
(а)
80
Модель (1 3)
Модель (4 5)
60
Без тренда
40
20
0
1950
196 0
197 0
198 0
199 0
2000
2010
2020
Года
(б)
80
60
40
20
0
1950
196 0
197 0
198 0
199 0
2000
2010
2020
Года
Рис. 5. Динамика численности сеголеток (а) и взрослых (б) особей судака Азовского моря в модельных экспериментах
с коэффициентом изъятия h = 0.07 в 1982-2016 гг. и запретом промысла с 2017 г. На графике (б) дополнительно
приведены траектории, рассчитанные с очищенным от восходящего тренда рядом температур. Серым цветом показан
разброс прогнозируемых моделью (4-5) численностей возрастных групп, соответствующий интервалу значений
коэффициента промысла 0 ≤ h ≤ 0.3. На графике (б) дополнительно приведены траектории, рассчитанные по обеим
моделям (1)-(3) и (4)-(5) с очищенным от восходящего тренда рядом температур.
турного режима обусловлено благоприятным для
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
воспроизводства низким значением солености в
Полученные результаты еще раз демонстриру-
краткосрочном прогнозе на 2019 г. (11.78‰), что
ют эффективность и применимость простых сто-
наглядно демонстрирует высокую чувствитель-
хастических моделей для долгосрочного прогно-
зирования динамики промысловых рыбных по-
ность судака к этому внешнему фактору. Пункти-
пуляций (см. также работы [2, 11, 45-51]).
ром показан критический порог квазивымира-
Выполненное исследование позволяет сделать
ния, составляющий 10% от максимальной чис-
вывод о том, что причиной уменьшения запасов
ленности взрослых особей в 1950-1981 гг., для
судака в последние десятилетия, наряду с ростом
модели (4)-(5) равный 7.5 млн шт. Так же как и
солености, может быть увеличение температуры
полная модель (1)-(3), под влиянием наблюдае-
моря. При этом эффект, в настоящее время ока-
мых и прогнозных значений солености и темпе-
зываемый на динамику численности стада потеп-
лением, может быть даже значительнее влияния
ратуры минимальная модель (4)-(5) предсказы-
роста солености моря.
вает падение численности популяции судака зна-
Интересно, что до конца XX века в круг про-
чительно ниже критического уровня, при этом
блем, имеющих отношение к возможному паде-
решающую роль в снижении запаса играет на-
нию запаса судака в Азовском море, включались
блюдающийся с конца 1980-х годов рост средне-
прежде всего негативные последствия зарегули-
годовых температур Азовского моря.
рования пресного стока. Эти последствия рас-
БИОФИЗИКА том 65
№ 2
2020
ОПРАВДАЛСЯ ЛИ ДОЛГОСРОЧНЫЙ ПРОГНОЗ
399
сматривались в контексте антропогенных изме-
специалистами особенностью текущего состоя-
нений экосистем Дона и Азовского моря, обу-
ния Азовского моря, как одновременные значи-
словленных строительством Цимлянского
тельные положительные аномалии температуры
водохранилища [3, 4, 52, 53]. К ним, помимо осо-
и солености вод, которые ранее не наблюдались в
лонения моря и инвазии гребневика мнемиопси-
истории изучения данного водоема [37].
са (Mnemiopsis leidyi) [54], относились перекрытие
Таким образом, мы можем сделать еще одно
путей миграций и сокращение естественных не-
заключение на основании проведенного исследо-
рестовых ареалов проходных и полупроходных
вания. Оно заключается в том, что даже те «песси-
видов промысловых рыб [4, 24]. Не меньшее вни-
мистические» оценки риска квазивымирания су-
мание уделялось неконтролируемому, в частно-
дака, которые были получены нами двадцать лет
сти, браконьерскому промыслу судака, пробле-
назад [1, 2], следует считать заниженными. Это
мам перелова [4, 23] и непреднамеренного вылова
объясняется тем, что псевдослучайные значения
годовиков [2]. В настоящее время запрет промыс-
среднегодовых температур, которые были ис-
ла рассматривается в качестве меры, способной
пользованы в имитационной модели (1)-(3) при
остановить падение запасов данного вида [19, 20].
проведении экспериментов Монте-Карло, гене-
Оценивая потенциал восстановления популяции
рировались на основе ранее наблюдаемых данных
при полном запрете промысла судака, авторы ра-
и потому не могли содержать экстремально высо-
боты [27] пришли к выводу, что кормовая база в
ких значений, характерных для сегодняшнего пе-
Азовском море не является лимитирующим фак-
риода.
тором, способствующим сокращению его запа-
сов. Однако ретроспективный анализ воздей-
ствия среднегодовой температуры на динамику
ВЫВОДЫ
популяции азовского судака меняет оценку эф-
Выполненное исследование позволяет сделать
фективности запрета промысла и перспектив
следующие общие выводы:
восстановления промыслового стада. В этой свя-
1) выявлено достаточно хорошее качественное
зи следует заметить, что хотя воздействие темпе-
и количественное соответствие прогноза модели
ратуры моря на воспроизводство и выживаемость
реальной динамике численности возрастных
судака признавалось важным, что и обусловило
групп азовского судака;
необходимостью ее включения в модель (1)-(3),
2) наблюдающееся с начала
2000-х годов
данный фактор не рассматривался специалиста-
устойчивое снижение полученных ихтиологами
ми-ихтиологами в качестве критического,
оценок численности популяции, которое явилось
способного подорвать популяцию. Тем не менее
основанием введения полного запрета промысла
модель (1)-(3) смогла достаточно корректно от-
азовского судака, подтверждает правильность
разить негативные последствия роста температу-
сделанного двадцать лет назад долгосрочного ве-
ры Азовского моря, наблюдающегося с конца
роятностного прогноза квазивымирания популя-
1980-х годов.
ции;
Дополнительные эксперименты с минималь-
3) в течение последнего десятилетия, помимо
ной моделью (4)-(5) показали, что предложен-
осолонения, негативное воздействие на воспро-
ный подход к моделированию динамики популя-
изводство судака оказывает наблюдаемый рост
ции судака Азовского моря устойчив по отноше-
среднегодовой температуры Азовского моря;
нию к структурным изменениям модели. Это
4) модельные эксперименты показывают, что
говорит о том, что информация о динамике и па-
в сложившихся условиях значительных положи-
дении численности заложена в использованных
тельных аномалий температуры и солености, вве-
при идентификации модели данных за 1950-
денный в 2017 г. запрет промысла обладает огра-
1981 гг. При этом очевидно, что влияние факто-
ниченным потенциалом восстановления запаса
ров внешней среды, в особенности температуры,
судака.
на прогнозируемые падения численности судака
до уровня квазивымирания является, несомнен-
но, решающим. Несмотря на то что точное значе-
ФИНАНСИРОВАНИЕ РАБОТЫ
ние параметра промысловой нагрузки не может
быть определено, модельные расчеты с варьируе-
Публикация подготовлена в ходе реализации
мым в широких пределах коэффициентом изъя-
ГЗ ЮНЦ РАН, тема «Разработка методов геоин-
тия h ∈ [0.0, 0.3] показывают, что потенциал вос-
формационного моделирования морских и на-
становления популяции от введенного в 2017 г.
земных экосистем», № госрегистрации проекта
запрета на промысел азовского судака невелик,
01201363188 (Тютюнов, Дашкевич); базовой части
тем более что вплоть до недавнего времени имела
ГЗ ЮФУ, тема «Фундаментальные и прикладные
место высокая, не соответствующая уровню вос-
задачи математического моделирования»,
№
производства, интенсивность как официального,
1.5169.2017/8.9 (Титова); проекта Российского
так и браконьерского промысла судака [4, 18, 23,
фонда фундаментальных исследований № 18-01-
24, 41]. Ситуация ухудшается такой отмеченной
00453а
«Мультистабильные пространственно-
БИОФИЗИКА том 65
№ 2
2020
400
ТЮТЮНОВ и др.
временные сценарии в популяционных системах»
16.
А. М. Бронфман и Ф. А. Сурков, Труды ВНИРО
(Тютюнов).
118, 62 (1976).
17.
Д. Я. Раткович, Водные ресурсы, № 1, 5 (1977).
18.
Д. А. Подойницын, Изв. вузов. Сев.-Кавк. регион.
КОНФЛИКТ ИНТЕРЕСОВ
Естеств. науки, № 4, 111 (2010).
Авторы заявляют об отсутствии конфликтов
19.
Протокол XXIX сессии Российско-Украинской Ко-
миссии по вопросам рыболовства в Азовском море
интересов.
(Ростов-на-Дону, 2018).
20.
Протокол XXX сессии Украинско-Российской
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
Комиссии по вопросам рыболовства в Азовском
море (Ростов-на-Дону, 2019).
Настоящая работа не содержит описания ка-
21.
В. Н. Белоусов, в сб. Основные проблемы рыбного хо-
ких-либо исследований с использованием людей
зяйства и охраны рыбохозяйственных водоемов Азо-
и животных в качестве объектов.
во-Черноморского бассейна (1998-1999 гг.) (Изд-во
БКИ, Ростов-на-Дону, 2000), сс. 88-96.
22.
В. Н. Белоусов, в сб. Основные проблемы рыбного хо-
СПИСОК ЛИТЕРАТУРЫ
зяйства и охраны рыбохозяйственных водоемов Азо-
во-Черноморского бассейна (2000-2001 гг.) (Вопро-
1. Ю. В. Тютюнов, И. Н. Сенина и Л. И. Титова, в кн.
сы рыболовства, М., 2002), сс. 279-291.
Компьютерное моделирование. Экология: учебное по-
собие, под ред. Г. А. Угольницкого (Вузовская кни-
23.
В. Н. Белоусов, в сб. Основные проблемы рыбного хо-
га, М., 2000), сс. 58-78.
зяйства и охраны рыбохозяйственных водоемов Азо-
во-Черноморского бассейна (2002-2003 гг.) (Медиа-
2. Yu. Tyutyunov, I. Senina, C. Jost, and R. Arditi, Ecol.
Пресс, Ростов-на-Дону, 2004), сс. 131-137.
Modelling 149, 297 (2002).
24.
В. Н. Белоусов, Автореф. дис. … канд. биол. наук
3. В. В. Дроздов, Ученые записки Российского госу-
(КубГАУ, Краснодар, 2004).
дарственного гидрометеорологического универ-
25.
Д. А. Подойницын, в сб. Основные проблемы рыбно-
ситета 15, 155 (2010).
го хозяйства и охраны рыбохозяйственных водоемов
4. П. А. Балыкин, Д. Н. Куцын и А. М. Орлов, Океа-
Азово-Черноморского бассейна (2004-2005 гг.) (Изд-
нология 59 (3), 396 (2019).
во «Диапазон», Ростов-на-Дону, 2006), сс. 199-
5. L. R. Ginzburg, L. B. Slobodkin, K. Jonson, and
207.
A. G. Bindman, Risk Analysis 2, 171 (1982).
26.
Е. П. Цуникова, Т. М. Попова, Е. А. Порошина и
6. H. R. Akçakaya and L. R. Ginzburg, in Species conser-
Е. А. Нефедова, в сб. Основные проблемы рыбного
vation: a population biological approach, Ed. by A. Seitz
хозяйства и охраны рыбохозяйственных водоемов
and V. Loeschke (Birkhäuser Verlag, Basel, 1991),
Азово-Черноморского бассейна (2006-2007 гг.) (Ме-
pp. 78-87.
диа-Пресс, Ростов-на-Дону, 2008), сс. 270-293.
7. W. E. Ricker, J. Fish. Res. Board of Canada 11, 559
27.
В. А. Гетманенко, Е. П. Губанов и Л. В. Изергин,
(1954).
Рибне господарство України: науково-виробни-
чий журнал, № 3, 4 (2012).
8. V. E. Shelford, Animal communities in temperate Ameri-
ca: as illustrated in the Chicago region: a study in animal
28.
А. А. Живоглядов и С. В. Лукьянов, Вопр. рыбо-
ecology (No. 5) (University of Chicago Press, Chicago,
ловства 19 (4), 405 (2018).
Illinois, 1913).
29.
С. В. Лукьянов и А. А. Живоглядов, в сб. Экологи-
9. Э. Пианка, Эволюционная экология (Мир, М., 1981).
ческая, промышленная и энергетическая безопас-
ность - 2018 (Севастопольский гос. ун-т, Севасто-
10. В. Г. Ильичев, Устойчивость, адаптация и управле-
поль, 2018), сс. 739-743.
ние в экологических системах (Физматлит, М.,
30.
С. В. Лукьянов и А. А. Живоглядов, в сб. Конфликт
2009).
природопользования: роль в эволюции ноосферы (Аст-
11. I. Senina, Yu. Tyutyunov, R. Arditi, J. Appl. Ecol. 36,
раханский гос. ун-т, Астрахань, 2019), сс. 78-83.
297 (1999).
31.
Протокол XIX сессии Российско-Украинской Комис-
12. Н. П. Гоптарев, А. И. Симонов, Б. М. Затучная и
сии по вопросам рыболовства в Азовском море
Д. Е. Гершанович, Гидрометеорология и гидрохимия
морей СССР. Т. 5. Азовское море (Гидрометеоиздат,
document/902114750
Санкт-Петербург, 1991).
32.
Протокол XX сессии Российско-Украинской Комис-
13. А. М. Бронфман, С. П. Воловик, С. В. Козлитина
сии по вопросам рыболовства в Азовском море (Бер-
и др., Статистическая структура океанологиче-
ских и биологических параметров экосистемы Азов-
ru/products/ipo/prime/doc/2067519
ского моря (Изд-во Рост. ун-та, Ростов-на-Дону,
33.
Протокол XXI сессии Российско-Украинской Комиссии
1979).
по вопросам рыболовства в Азовском море
14. А. М. Бронфман, В. Г. Дубинина и Г. Д. Макарова,
(пос. Рожок Ростовской области, Россия,
2009).
Гидрологические и гидрохимические основы продук-
тивности Азовского моря (Пищевая промышлен-
index. htm
ность, М., 1979).
34.
Протокол XXIII сессии Российско-Украинской Комиссии
15. B. Efron and R. J. Tibshirani, An introduction to the
по вопросам рыболовства в Азовском море (Темрюк,
bootstrap (Chapman and Hall, London, 1993).
БИОФИЗИКА том 65
№ 2
2020
ОПРАВДАЛСЯ ЛИ ДОЛГОСРОЧНЫЙ ПРОГНОЗ
401
fishindustry.net/govermental-bodies/2164-rezultaty-xxiii-
43. А. В. Никитина, Т. В. Руднева, Т. В. Камышникова
sessii-rossiysko-ukrainskoy-komissii-po-voprosam-
и др., Современные проблемы науки и образова-
rybolovstva-v-azovskom-more.html
ния,
№ 1,
1
35. Протокол XXIV сессии Российско-Украинской Ко-
education.ru/ru/article/view?id=19509
миссии по вопросам рыболовства в Азовском море
44. В. В. Кулыгин, В. С. Герасюк и Л. В. Дашкевич,
(Бердянск, Украина, 2012).
Информация и космос, № 1, 120 (2018).
36. Протокол XXVIII сессии Российско-Украинской Ко-
45. Yu. Tyutyunov, R. Arditi, B. Buttiker, et al., Ecol. Mod-
миссии по вопросам рыболовства в Азовском море
elling 69, 19 (1993).
46. А. Б. Медвинский, А. В. Русаков, А. Е. Бобырев
novosti/xxviii-sessiya-rossiysko-ukrainskoy-komissii-po-
и др., Биофизика 54 (1), 120 (2009).
voprosam-ryibolovstva-v-azovskom-more-zavershila-
47. L. V. Nedorezov, Population Dynamics: Analysis,
svoyu-rabotu-v-kieve-ukraina/
Modelling, Forecast 2 (4), 129 (2013).
37. Л. В. Дашкевич, С. В. Бердников и В. В. Кулыгин,
48. А. И. Абакумов, Информатика и системы управле-
Водные биоресурсы и среда обитания 2 (2), 7
ния 54 (4), 12 (2017).
(2019).
49. В. Г. Ильичев и О. А. Ильичева, Биофизика 63 (2),
38. Л. В. Дашкевич и В. В. Кулыгин, Материалы Между-
373 (2018).
народной конференции «ИнтерКарто. ИнтерГИС.
Геоинформационное обеспечение устойчивого раз-
50. В. Г. Ильичев и Л. В. Дашкевич, Компьютерные
исследования и моделирование 11 (5), 879 (2019).
10.35595/2414-9179-2019-2-25-112-120
51. G. P. Neverova, A. I. Abakumov, I. P. Yarovenko, and
39. Л. В. Дашкевич, С. В. Бердников и В. В. Кулыгин,
E. Ya. Frisman, Nonlinear Dynamics 94 (2), 827
Водные ресурсы
44
(5),
563
(2017).
(2018).
52. И. И. Ворович, А. С. Горелов, А. Б. Горстко и др.,
40. С. В. Бердников, в сб. Закономерности экосистем-
Рациональное использование водных ресурсов бассей-
ных процессов в Азовском море, под ред. Г. Г. Мати-
на Азовского моря. Математические модели (Наука,
шова (Наука, М., 2006), сс. 137-229.
М., 1981).
41. Д. А. Подойницын, Автореф. дис. … канд. биол.
53. Г. Г. Матишов, Ю. М. Гаргопа, С. В. Бердников и
наук (ЮФУ, Ростов-на-Дону, 2010).
С. Л. Дженюк, Закономерности экосистемных про-
42. Л. В. Кукарина, в сб. Основные проблемы рыбного
цессов в Азовском море, под ред. Г. Г. Матишова
хозяйства и охраны рыбохозяйственных водоемов
(Наука, М., 2006).
Азово-Черноморского бассейна (АзНИИРХ, Ростов-
54. S. V. Berdnikov, V. V. Selyutin, V. V. Vasilchenko, and
на-Дону, 1996), сс. 183-186.
J. F. Caddy, Fisheries Res. 42 (3), 261 (1999).
Long-Range Prediction of the Risk of Extinction Faced by the Pikeperch
in the Azov Sea: Has the Prediction Come True?
Yu.V. Tyutyunov*, **, I.N. Senina***, L.I. Titova**, and L.V. Dashkevich*
*Federal Research Centre “The Southern Scientific Centre of the Russian Academy of Sciences”,
prosp. Chekhova 41, Rostov-on-Don, 344006 Russia
**Southern Federal University, prosp. Stachki 200/1, Rostov-on-Don, 344090 Russia
***Collecte Localisation Satellites, Space Oceanography Division (в русском варианте отсутствует),
rue Hermes 8-10, Ramonville Saint-Agne, 31520 France
In the late 1990s, based on stochastic simulations with a model of a long-range prediction of the dynamics of
the Azov Sea pikeperch population identified from the 1950-1981 observational data including stocks, catch-
es, temperature and salinities in the Azov sea, because the salinity of the seawater increases, a high probability
of a risk of quasiextinction faced by the pikeperch population in the Azov Sea within 35 years from 1982 to
2016 was calculated. This probabilistic prediction has come true - the risk event occurred in the 2000s. Since
2005, the abundance of adult pikeperch did not exceed 4 million individuals. Since 2017, the pikeperch catch
in the Azov Sea was completely forbidden. Nowadays, retrospective validation demonstrates that the model
prediction is in a good (qualitative and quantitative) correspondence with the 1982-2018 observation data on
dynamics of the pikeperch population. It has also been found that currently observed rise in annual average
sea temperature has a negative impact on fish population dynamics and this phenomenon is no less signifi-
cant than increase in salinity. According to a short-range prediction for 2019-2020, due to existing positive
temperature and salinity anomalies, the ban on fishing introduced in 2017 is not enough to rebuild stocks of
the Azov Sea pikeperch population in the coming years.
Keywords: Azov Sea, population dynamics, Sander Lucioperca L., stochastic model, Monte-Carlo method, risk
of quasi-extinction
БИОФИЗИКА том 65
№ 2
2020