БИОФИЗИКА, 2020, том 65, № 3, с. 557-564
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 577.3
ФЕНОТИПИЧЕСКАЯ АДАПТАЦИЯ МОРСКИХ ЕЖЕЙ,
ВЫРАЩЕННЫХ В УСЛОВИЯХ ОГРАНИЧЕННОГО ПРОСТРАНСТВА
© 2020 г. Р. Лиу, III. Педлар, А. Од, С. МакДоналд, Дж. Стоун
McMaster University, 1280 Main Street West, Hamilton ON, L8S 4L8, Canada
E-mail: jstoner@mcmaster.ca
Поступила в редакцию 06.06.2019 г.
После доработки 09.01.2020 г.
Принята к публикации 15.01.2020 г.
Морские ежи и их родственники таксономически сгруппированы в класс Echinoidea (тип Echino-
dermata). Эхиноидьт демонстрируют значительные различия в своих фенотипах пятиугольной сим-
метричной формы тела, от сфероидальных панцирей (скелетов) у морских ежей до дискоидных пан-
цирей у плоских щитовидных ежей. Многие исследователи размышляли о морфологических эволю-
ционных переходах между этими фенотипами, но не достигли консенсуса. Авторы разработали и
выполнили эксперимент, в котором морские ежи росли в условиях пространственных ограничений
в вертикальном измерении, физически ограничив их рост в высоту. Особи в этой группе стали более
плоскими по сравнению с особями в контрольной группе по истечении восьми недель. Эти резуль-
таты можно рассматривать как результат фенотипической адаптации, недооцененного феномена,
который может стать первым шагом в эволюции нового фенотипа. Авторы представляют предвари-
тельные предположения, касающиеся фенотипически адаптированных изменений в форме и мор-
фологических эволюционных отношений между иглокожими. Демонстрируя правдоподобный
инициирующий механизм в морфологическом переходе, авторы надеются внести вклад в теорети-
ческие дискуссии о фенотипической эволюции, подчеркивая роль, которую физические факторы
играют в изменении сложных биологических систем.
Ключевые слова: развитие; эволюция; морфология; фенотипическая пластичность.
DOI: 10.31857/S0006302920030163
Класс Echinoidea (тип иглокожих - Echinoder-
менной филогенетической систематики, Недав-
mata) содержит морских ежей («правильные» эхи-
ний филогенетический систематический анализ
ноиды), плоских щитовидных ежей, «песчаных
[1] может быть истолкован как предположение,
долларов», морских плоских ежей (Clypeasteroida)
что сфероидные панцири в наследственной груп-
и их родственников (Cassiduloida), а также серд-
пе превратились в дискоидные, а также в эллип-
цевидных ежей (Spatangoida) и их родственников
соидные и кардиоидные панцири в линиях, веду-
(Holasteroida, Disasteroida). В этих группах пред-
щих к «песчаному доллару», морским плоским и
ставлен спектр фенотипов пентарадиальной фор-
сердцевидным ежам.
мы тела. Две группы могут рассматриваться как
типичные крайности в этом фенотипическом
Морские ежи традиционно считаются «мо-
спектре: «правильные» морские ежи, характери-
дельными организмами» для проведения иссле-
зуемые сфероидальными панцирями (скелета-
дований в области биологии развития (например,
ми), и «неправильные» - «песчаные доллары»,
работы [2-6]). Также можно считать, что морские
характеризуемые дискоидными панцирями,
ежи стали модельными организмами для прове-
морские плоские ежи, характеризуемые эллипсо-
дения исследований в области эволюционной
идными панцирями, и сердцевидные ежи, харак-
биологии развития («evo-devo» research) (напри-
теризующиеся кардиоидными панцирями в ка-
мер, работы [7-10]). Сильная междисциплинар-
честве промежуточных продуктов. Однако от-
ная взаимосвязь между эволюционной биологией
сутствуют общепринятые морфологические
и биологией развития и способность панцирей
эволюционные переходы между разновидностя-
морского ежа на структурную гибкость (описан-
ми панциря (т. е. размерами и формами), которые
ную ниже) делает морских ежей идеальными ор-
можно было бы предположить на основе совре-
ганизмами для изучения морфологических эво-
557
558
ЛИУ и др.
люционных переходов, особенно трансформа-
Апикальная система
ций, связанных с изменениями размера и формы.
Высота
Панцири морского ежа - это эндоскелеты,
каждый из которых находится между нижележа-
щей внутренней целомической подкладкой и вы-
Диаметр
шележащим внешним эпидермисом [11]. Каждый
панцирь состоит из трех компонентов (снизу
вверх в направлении подачи субстрата, рис. 1):
перистома, содержащая щечные (ротовые) пла-
стинки и иногда первичные амбулакральные пла-
Перистома
стинки [11, 12], которые окружают рот, или ари-
стотелев фонарь; корона, содержащая амбула-
Рис. 1. Панцирь морского ежа в боковой перспективе,
кральные и интерамбулакральные пластинки;
показывающий три компонента (снизу вверх) -
апикальная система, состоящая из глазных и ге-
перистому, корону и апикальную систему, и две
нитальных пластинок (и перипрокт у правильных
измеренные переменные - высоту и диаметр.
и ранних неправильных эхиноидов [12]).
Пластины панциря содержат связанный с бел-
Классическая неодарвинистская эволюция
ком магнезиальный кальцит и сшиваются друг с
происходит, когда выгодные модификации воз-
другом коллагеновыми волокнами на балках, на-
никают из-за генетических мутаций (т.е. когда
зываемых трабекулами [13, 14]. Индивидуальный
мутантные аллели кодируют новые фенотипы) и
рост скелета включает пять интегрированных
впоследствии выбираются положительной селек-
процессов: возникновение промежутков между
цией. Фенотипическая пластичность - явление,
пластинами, когда волокна коллагена разрыхля-
при котором отдельные характеристики изменя-
ются; добавление новых пластин в апикальную
ются в ответ на факторы окружающей среды, -
систему; нарастание нового скелетного материа-
может возникнуть при таких сценариях, когда са-
ла на периферии пластин; взаимодействия, про-
ма изменчивость (кодируемая генотипически и
исходящие на границах пластин; рост внутренних
выраженная фенотипически) была выбрана по-
органов, влияющий на стенку тела [15]. Эти ске-
ложительно. Наоборот, явления, известные как
летогенетические процессы происходят в «зонах
фенотипическая и генотипическая аккомодация,
роста», двухсторонних амбулакральных балках,
имеют другую, обратную последовательность.
по обеим сторонам которых находится интерам-
Фенотипы, которые находятся в пределах потен-
булакральная балка [16-19]. Коллагеновые во-
циально реализуемого морфологического диапа-
локна растягиваются во время активного роста,
зона, возникают в ответ на индукторы окружаю-
позволяя возникать промежуткам между пласти-
щей среды; это фенотипическая адаптация (мута-
нами; новые пластины добавляются рядом с глаз-
ными пластинами; это приводит к тому, что более
ция, как другой возможный индуцирующий
старьте пластины в зоне роста оказываются бли-
фактор, здесь не рассматривается как таковая
же к перистоме [20]; новый скелетный материал
[27]). Генотипы, участвующие в экспрессии этих
накапливается вокруг старых пластин периферии
фенотипов (и, следовательно, сами фенотипы),
и размер панциря увеличивается [13, 14, 20, 21].
могут становиться фиксированными в течение
продолжительных периодов времени и несколь-
Рост панциря происходит естественным обра-
ких поколений; если индуктор окружающей сре-
зом в течение активных сезонов, быстро в течение
ды непрерывно передает свои эффекты в течение
нескольких недель после кормления, а затем бо-
этих периодов времени, то изменение генотипа
лее медленно и скачкообразно [22]; у экспери-
(и, следовательно, изменение фенотипа) в конеч-
ментально питаемых морских ежей рост панциря
ном итоге уменьшается, особенно если «затраты»
выражен более четко [23]. Не кормящиеся мор-
будут понесены особями, сохраняющими фено-
ские ежи характеризуются жесткими панцирями;
типическую гибкость; это генотипическая адап-
не кормленые в ходе экспериментов морские ежи
тация (то, что согласованные ответы эквивалент-
характеризуются панцирями, которые со време-
но могут быть исключены, здесь не рассматрива-
нем сжимаются из-за сокращения волокон кол-
ется
[27]). Таким образом, фенотипическая
лагена [14, 24]. Этот прерывистый рост в сочета-
нии с основным пятиступенчатым скелетогене-
аккомодация, сопровождаемая генотипической
зом можно сравнить с линькой [22]. На рост
аккомодацией, включает первоначальное изме-
морских ежей влияют различные условные фак-
нение частоты фенотипа (а не генотипа), которое
торы, связанные с питанием [25] и условиями
зависит от повторного воздействия индуцирую-
окружающей среды [26], как и при линьке у дру-
щего фактора окружающей среды, подлежащего
гих животных.
отбору. Следовательно, фенотипическая адапта-
БИОФИЗИКА том 65
№ 3
2020
ФЕНОТИПИЧЕСКАЯ АДАПТАЦИЯ МОРСКИХ ЕЖЕЙ
559
ция может ускорять быструю морфологическую
продемонстрированы у морских ежей, когда
эволюцию.
ограниченный корм являлся сигналом окружаю-
щей среды, влияющим на изменения морфоло-
Учитывая возможность инициирования рез-
гии [14, 21, 25, 26, 29]. Способность регулировать
кой морфологической эволюции, фенотипиче-
форму в соответствии с условиями окружающей
ская адаптация должна быть тщательно изучена.
среды обеспечивается описанным ранее дискрет-
Однако исследования фенотипической адапта-
ным пятиступенчатым процессом роста панциря
ции, особенно документированные случаи, были
морского ежа.
редкостью, отчасти потому, что соответствующие
механизмы часто включают физические факто-
Подобный динамический ответ на кормление
ры, от которых большинство биологов обычно
с эпизодическим ростом в сочетании с вышеупо-
уклоняются. Это вызывает сожаление, поскольку
мянутой гибкостью, обеспечиваемой волокнами
физические факторы играют важную роль в изме-
коллагена, превращает морских ежей в организ-
нении сложных биологических систем [28], и по-
мы, хорошо подходящие для изучения фенотипи-
нимание этих ролей, особенно с помощью доку-
ческой аккомодации в скелетогенезе в контроли-
ментированных случаев, будет способствовать
руемых экспериментальных условиях. В есте-
теоретическим дискуссиям о фенотипической
ственных условиях химические загрязнители и
эволюции. Тем не менее некоторые случаи и мо-
твердость субстрата могут влиять на формы пан-
менты были описаны.
циря. Было показано, что металлы и детергенты
ингибируют кальцификацию и приводят к сплю-
Фенотипическая адаптация может вызывать
щиванию панцирей [30]. Слабые субстраты могут
изменения в развитии популяции. Например,
препятствовать сцеплению с поверхностями и
пятнистые гиены (Crocuta crocuta) характеризуют-
приводить к укреплению панцирей [21, 31]. Такие
ся наличием ребристых черепов в зрелом воз-
ответы панциря были использованы для разра-
расте; несовершеннолетние же особи не имеют
ботки эксперимента для документирования опи-
гребней. Формирование гребня объясняется си-
санной здесь фенотипической аккомодации.
лами, создаваемыми во время развития мощными
Морских ежей выращивали в искусственной сре-
мышцами, охватывающими кости челюсти, когда
де, которая ограничивала вертикальное измере-
особи постоянно жуют жесткое мясо и кости от
ние, доступное для скелетогенеза, чтобы опреде-
своей добычи [27]. В неволе популяции особей,
лить, может ли рост при геометрическом ограни-
выращенных на мягкой диете, не генерируют ни-
чении вызывать морфологические изменения в
каких сил, и, следовательно, взрослые животные
панцирях, выражающееся в уменьшении высо-
в неволе не имеют гребней черепа, как их моло-
ты/диаметра.
дые коллеги в диких популяциях. Жесткая пища в
дикой природе влияет на использование мышц и
развитие костей - реакция на механическое дав-
МАТЕРИАЛЫ И МЕТОДЫ
ление стала неотъемлемой частью процесса раз-
Вид Arbacia punctulata (Lamarck, 1816) был вы-
вития в естественных популяциях; был ли этот
бран из-за доступности образцов и отсутствия ре-
ответ значим или был преувеличен изменением
генотипа, неизвестно.
продуктивной функции во время проведения
эксперимента (использование неразмножающе-
Фенотипическая адаптация может создать
гося объекта будет гарантировать, что поступаю-
сложное новшество у одной особи без генетиче-
щая через кормление энергия будет отводиться
ских изменений. Например, коза, которая роди-
преимущественно росту). Взрослые особи были
лась с парализованными передними конечностя-
приобретены в Морской лаборатории Gulf Speci-
ми, научилась двигаться, подпрыгивая на задних
men (Панасея, Флорида, США) в течение двух
лапах [27]). Анатомическое обследование после
полевых сезонов (начало января 2010 и 2014 гг.)
ее смерти выявило множество модификаций ко-
для проведения двух экспериментальных испыта-
стей задних конечностей и таза, которые способ-
ний с использованием одной и той же начальной
ствовали фенотипической адаптации. Эти два
популяции. Для каждого испытания особи прохо-
случая: ребристые черепа в пятнистых популяци-
дили короткий период акклиматизации, в тече-
ях гиены и модификации костей, позволяющие
ние которого питание не предоставлялось (т.е.
прыгать отдельной козе, демонстрируют феноти-
период голодания составлял приблизительно од-
пическую экспрессию, вызванную поведением и
ну неделю). Цифровые штангенциркули Samona
включаемую генотипами в ответ на индукторы
(Оттава, Канада) были использованы для измере-
окружающей среды.
ния у каждой особи значений высоты (h) и диа-
Отчетливый ответ на сигнал окружающей сре-
метра (d). Из-за обширной логистики, связанной
ды, влияющий на выживаемость, включает изме-
с протоколом роста (описанным далее), размеры
нение размера тела для снижения затрат на соб-
выборки для контрольной и экспериментальной
ственное обслуживание при оптимизации репро-
групп были ограничены соответственно шестью и
дуктивной мощности; такие случаи были
четырьмя образцами.
БИОФИЗИКА том 65
№ 3
2020
560
ЛИУ и др.
(а)
(а)
(б)
(б)
Рис. 2. Аппарат для ограничения вертикального про-
странственного размера и в конечном счете создания
реактивных сил сжатия, действующих на панцирь
морского ежа: (а) - отдельно, (б) - in situ в аквариуме.
Устройство было разработано для выявления роли,
которую физические факторы могут играть в измене-
нии сложных биологических систем, таких как фор-
мы для панцирей морского ежа.
Отдельных особей содержали в отфильтрован-
ной искусственной морской воде (Oceanic Natural
Sea Salt Mix, Central Garden & Pet Company, Уол-
нат-Крик, Калифорния, США; ρspec = 1.020-
1.023) в аквариумах (TopFin, PetSmart, Финикс,
Рис. 3. Экспериментальные аквариумы для проверки
Аризона, США; максимальный объем 6 л) при
того, влияет ли рост морских ежей при геометриче-
комнатной температуре (Т = 24°С). Обе группы
ских ограничениях на их форму скелета: (а) - образ-
кормили морковью, нарезанной поперек колеч-
цы в контрольном аквариуме (фронтальная перспек-
ками, запасы продовольствия пополняли по мере
тива; особи, помеченные цветной нитью); (б) - об-
разцы в аквариуме воздействия (боковой вид; особи,
истощения. Пополняли воду и очищали аквари-
помеченные цветной нитью). Особи росли в попереч-
умы при необходимости (примерно раз в неде-
ном направлении, увеличиваясь в диаметре, и стано-
лю). Размеры аквариума и объемы морской воды
вились более плоскими, уменьшаясь в соотношении
определяли на основе размеров образцов, чтобы
высоты к диаметру.
обеспечить приблизительно одинаковую плот-
ность особей на объем для контрольной и экспе-
аппарат регулировали в течение всего экспери-
риментальной групп.
ментального периода (т.е. всякий раз, когда по-
Особи с близкими значениями h были включе-
полняли пищу или воду или чистили аквариумы)
ны в экспериментальную группу (без каких-либо
так, чтобы платформа аппарата просто контакти-
других оценок) и помещены в аквариум, который
ровала с растущими образцами. Особи с близки-
был оборудован устройством, ограничивающим
ми по значению h (опять же без каких-либо дру-
вертикальные
пространственные
размеры
гих оценок) были включены и в контрольную
(рис. 2), и в конечном счете (т.е. после того, как
особи выросли так, чтобы их шипы контактиро-
группу. Апикальные колючки на образцах в экс-
вали с платформой аппарата) создающим в экс-
периментальной и контрольной группах были
перименте реактивные силы сжатия, действую-
подпилены до максимальной средней высоты.
щие на панцирь (рис. 3). В случае необходимости
Особи были отмечены нанесением на колючки
БИОФИЗИКА том 65
№ 3
2020
ФЕНОТИПИЧЕСКАЯ АДАПТАЦИЯ МОРСКИХ ЕЖЕЙ
561
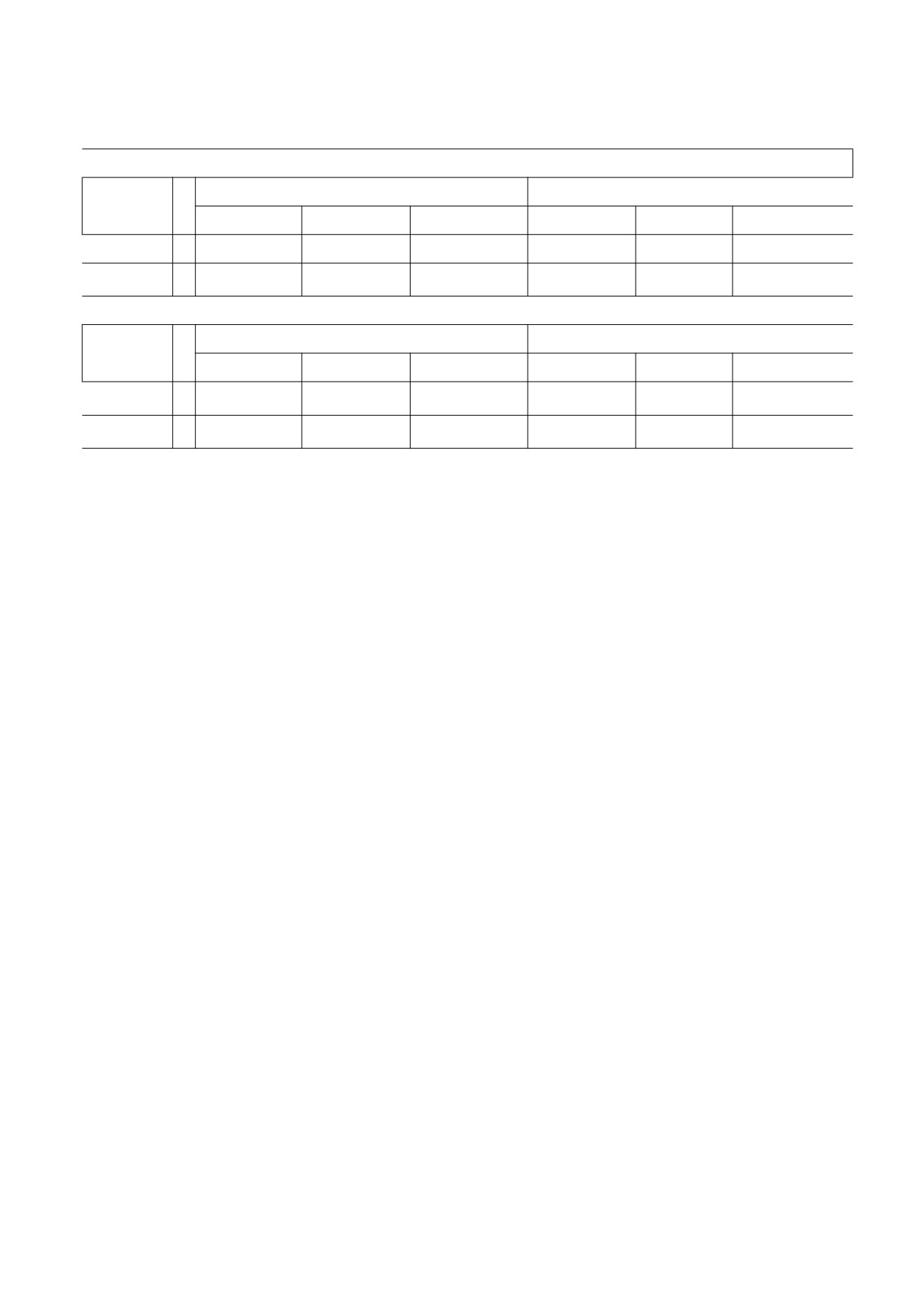
Значения высоты h, диаметра d панциря и отношения высоты к диаметру h/d в контрольной и эксперименталь-
ной группах для образцов Arbacia punctulata в начале и в конце (через восемь недель) эксперимента
Популяция 2010 года
Группа
n
Начальные измерения
Конечные измерения
h
d
h/d
h
d
h/d
Контроль
6
13.215 ± 1.555
21.985 ± 2.250
0.6010 ± 0.03116
14.723 ± 2.266
24.138 ± 2.583
0.6082 ± 0.04173
Воздействие
4
13.635 ± 0.7285
23.213 ± 1.374
0.5877 ± 0.01323
14.220 ± 1.083
26.218 ± 1.640
0.5425 ± 0.02709
Популяция 2014 года
Группа
n
Начальные измерения
Конечные измерения
h
d
h/d
h
d
h/d
Контроль
6
16.323 ± 2.624
28.287 ± 3.649
0.5769 ± 0.04736
18.600 ± 3.308
31.650 ± 3.774
0.5863 ± 0.07531
Воздействие
4
24.325 ± 3.813
31.650 ± 4.071
0.6454 ± 0.07999
22.675 ± 1.866
43.225 ± 2.871
0.5240 ± 0.04929
Примечание. Приведены средние значения ± стандартное отклонение. Различия в средних показателях до и после
эксперимента были проверены с помощью парных одностороних t-критериев. Коэффициент Коэна d был (согласно работе
[32]) рассчитан как разница между средними значениями, деленная на одно стандартное отклонение. Для популяции
2010 года: h для контрольной группы: t = 4.30, Р = 0.00380 (d Коэна = -0.974); h для группы воздействия: t = 3.02, Р = 0.0285
(d оэна = -0.732); d для контрольной группы: t = 3.96, Р = 0.00590 (d Коэна = -0.850); d для группы воздействия: t = 6.26,
Р = 0.00410 (d Коэна = -2.29); h/d для контрольной группы: t = 0.354, Р = 0.369 (d Коэна = -0.208); h/d для группы
воздействия: t = 3.18, Р = 0.0250 (d Коэна = -2,51). Для популяции 2014 года: h для контрольной группы: t = -1.28, Р = 0.1279
(d Коэна = 0.768); h для группы воздействия: t = 0.924, Р = 0.2118 (d Коэна = 0.581); d для контрольной группы: t = -5.51,
Р = 0.00135 (d Коэна = 0.906); d для группы воздействия: t = -4.74, Р = 0.00888 (d Коэна = 3.335); h/d для контрольной группы:
t = -0.364, Р = 0.3889 (d Коэна = 0.154); h/d для группы воздействия: t = -3.40, Р = 0.0212 (d Коэна = 1.878).
нерастворимого в воде лака для ногтей (China
соответствующие прогнозы, означающие «сред-
Glaze©, Чжудун, Северный Тайвань, Китай) или
нее значение hпосле = среднее значение hдо», сред-
завязыванием швейной нити примерно посере-
нее значение «dпосле = среднее значение dдо»,
дине вдоль шипов (Singer Sewing Company, Мерф-
среднее значение «hпосле/dпосле = среднее значе-
рисборо, Теннесси, США) и идентифицированы
ние hдо/dдо» и среднее значение «h/d для кон-
впоследствии на основе цветной маркировки.
Обе группы были обеспечены морковью ad libitum
трольной группы = среднее значение h/d для экс-
в эквивалентных количествах (равной массе мор-
периментальной группы», тестировали, приме-
кови на особь), таким образом, особи могли пи-
няя парные односторонние t-критерии с
таться непрерывно в течение всего эксперимента,
использованием GraphPad Prism Software (Graph-
в течение которого их содержали в одинаковых
Pad Software Inc., Сан-Диего, Калифорния,
условиях, но в ограниченном вертикальном изме-
США).
рении в аквариуме экспериментальной группы.
Значения hud измеряли и записывали один раз
РЕЗУЛЬТАТЫ
перед началом любого кормления (т.е. когда экс-
перимент был начат, сразу после периода голода-
Средние значения h и d были выше в конце
ния) и снова после восьми недель кормления (т.е.
эксперимента, чем в начале, для обеих групп в
когда эксперимент был прекращен). Группы
2010 г.; среднее значение d было больше в конце
сравнивали, рассчитывая величины эффекта как
эксперимента, чем в начале, для обеих групп в
грубую разницу между средними значениями и
2014 г. (таблица). Для группы воздействия сред-
используя коэффициент Коэна d [32]. Данные
также были проанализированы с использованием
нее значение h/d было меньше в конце экспери-
мента, чем в начале, в оба года (рис. 4, таблица;
обычных количественных протоколов: поскольку
для обеих групп предполагалось, что особи в них
величины эффекта, определенные как средние
будут расти, а конкретно в экспериментальной
изменения h/d, составляли 0.0072 и 0.0452 для
группе - что панцири будут более сплюснутыми
группы контроля и воздействия соответственно в
и, следовательно, будут характеризоваться мень-
2010 г. и 0.0094 и 0.1214 для контрольной и экспе-
шим средним значением h/d, была принята нуле-
риментальной группы соответственно в 2014 г.).
вая гипотеза «нет разницы между группами», и
Средние значения h/d между контрольной и экс-
БИОФИЗИКА том 65
№ 3
2020
562
ЛИУ и др.
периментальной группами значительно отлича-
(а)
лись в конце эксперимента в оба года.
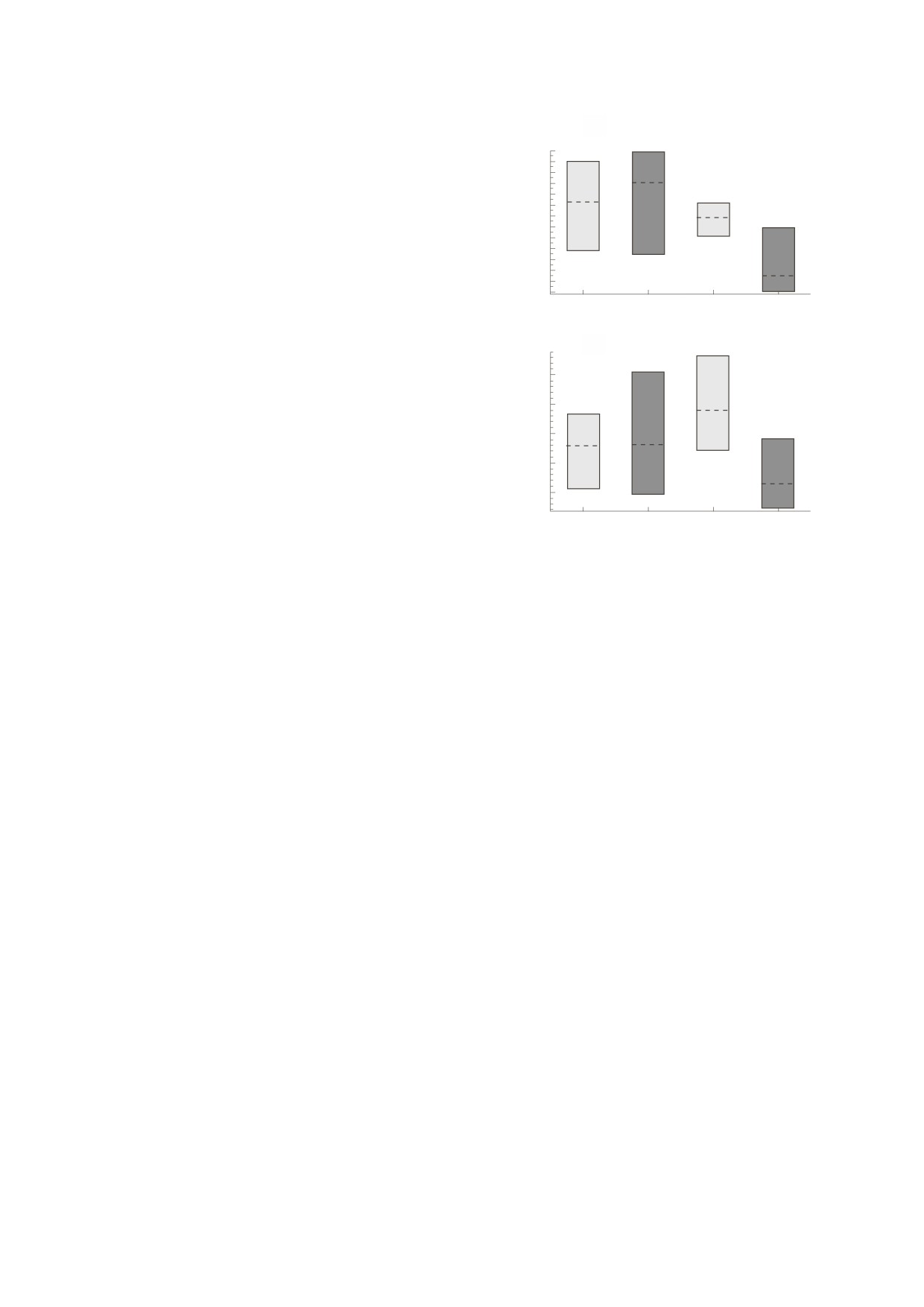
h/d
0.64
ОБСУЖДЕНИЕ
0.62
Среднее значение h/d, характеризующее пан-
0.60
цири морских ежей, которые выросли при гео-
0.58
метрическом ограничении, уменьшилось отно-
0.56
сительно среднего значения h/d, характеризую-
0.54
щего панцири морских ежей, которые выросли
без геометрического ограничения (рис. 4, табли-
0.52
Контроль
Эксперимент
ца). Это можно было обнаружить даже при не-
больших размерах выборки, которые были обу-
словлены трудоемкой логистикой, связанной с
h/d
(б)
экспериментальным дизайном; величина эффек-
0.70
та, связанного с фенотипической адаптацией у
этих морских ежей, предполагает, что такой ответ
0.65
соответствует понятию bona fide. Значение h/d для
всех образцов в экспериментальной группе изме-
0.60
нилось отрицательно в течение восьминедельно-
0.55
го периода в обоих исследованиях (т.е. в 2010 и
2014 гг.), тогда как h/d для образцов в контроль-
0.50
ной группе изменилось неравномерно, три поло-
жительных и три отрицательных случая в обоих
Контроль
Эксперимент
испытаниях.
Средние значения h/d для контрольной груп-
Рис. 4. Значения соотношения высоты и диаметра
пы в 2010 г. увеличились, но среднее значение h/d
панциря (h/d) в контрольной и экспериментальной
группах для образцов морского ежа в начале и через
осталось неизменным (таблица), что указывает на
восемь недель эксперимента (т.е. до и после кормле-
то, что образцы росли изометрическим образом
ния, светлый и темный столбики соответственно);
(т.е. они увеличивались в размерах, но сохраняли
каждый прямоугольник содержит медианное значе-
постоянные пропорции, как и ожидалось у эхи-
ние (пунктирная линия) с верхом и низом (то есть
«усы»), которые охватывают наблюдаемый диапазон:
ноидов [20, 33]), поскольку они были геометриче-
(а) - выборка популяции 2010 г., (б) - выборка попу-
ски не ограничены - особи в группе воздействия
ляции 2014 г.
в 2014 г. росли схожим образом, хотя различий по
h не было обнаружено (таблица). Особи в группе
воздействия в обоих исследованиях (то есть в 2010
Один из вероятных (весьма спекулятивных)
и 2014 гг.) росли аллометрическим образом (т.е.
сценариев морфологического расхождения меж-
увеличивались в размерах, но изменяли пропор-
ду сфероидальными морскими ежами и дискоид-
ции), потому что они были геометрически огра-
ными «песчаными долларами», эллипсоидальны-
ничены (см. таблицу) - увеличение d с уменьше-
ми морскими плоскими ежами, а также кардио-
нием h/d согласуется с фенотипической адапта-
идными сердцевидными ежами заключается в
цией.
наблюдении, что не кормящиеся ночные морские
Гибкие панцири и эпизодическое пищевое по-
ежи часто ищут убежища в дырах или трещинах в
ведение у взрослых морских ежей предрасполага-
скалах или под ними [35]. Морские ежи, как чле-
ют к их использованию в качестве кандидатов для
ны сообщества иглокожих (Echinodermata), ха-
проявления фенотипического аккомодации [27].
рактеризуются наличием изменчивой коллагено-
Мы зафиксировали фенотипическую адаптацию
вой ткани, которая замечательна своей способно-
в одной популяции за два разных года. Если такие
стью быстро изменять жесткость под нейронным
же гибкие панцири и эпизодическое пищевое по-
контролем [36-40]. Изменяющаяся коллагеновая
ведение характеризовали популяции в линиях,
ткань («catch» коллаген) может быть зафиксиро-
ведущих к «песчаным долларам» и морским плос-
вана на месте и укреплять поддерживающие тка-
ким ежам, а также к сердцевидным ежам, то фе-
ни, устраняя необходимость сокращения мышц
нотипическая адаптация обеспечивает правдопо-
при сохранении жизненных позиций (например,
добный механизм, через который проявляется те-
в расщелинах и трещинах на поверхности горных
кущее морфологическое неравенство
[34],
пород). Нахождение на месте в течение длитель-
наблюдаемое среди этих групп. Экологические
ных периодов времени может вызывать феноти-
сигналы или условия, которые могли бы вызвать
пическую адаптацию у особей в популяциях мор-
морфологическое расхождение, практически не-
ских ежей способом, аналогичным описанному
исчислимы.
здесь росту сплющенных панцирей, если поведе-
БИОФИЗИКА том 65
№ 3
2020
ФЕНОТИПИЧЕСКАЯ АДАПТАЦИЯ МОРСКИХ ЕЖЕЙ
563
ние сохранялось непрерывно на протяжении
ном и тетрациклином для визуализации линий
многих поколений. С современной неодарви-
роста [42, 43].
нистской, спекулятивной точки зрения, если бы
подобные геометрические ограничения действо-
ВЫВОДЫ
вали равномерно и выгодно на протяжении мно-
гих поколений, то индивидуальные величины h/d
Морских ежей выращивали в течение восьми
в популяциях могли бы измениться; генотипы,
недель в условиях геометрического ограничения,
лежащие в основе способности к переменному
при котором особи были ограничены в верти-
фенотипу, постепенно стали бы модифицирова-
кальном пространственном измерении. Особи в
ны и смещены в сторону создания более плоских
этой экспериментальной группе становились бо-
панцирей; популяция перешла бы в измененную
лее плоскими по сравнению с особями в кон-
форму.
трольной группе, создавая фенотипическую ак-
комодацию. Подобные геометрические ограни-
С представленными здесь данными могут быть
чения, действующие равномерно на протяжении
связаны два основных ограничения. Во-первых,
многих поколений, могли бы привести к феноти-
размеры выборки были небольшими. Следова-
пической адаптации, создающей более плоские
тельно, было недостаточно степеней свободы для
панцири. Этим можно было бы объяснить несо-
проведения непараметрических тестов Вилкок-
ответствие, наблюдаемое между сфероидальны-
сона с критерием знаковых рангов для сравне-
ми морскими ежами и дискоидными «песчаными
ний, в которых они были бы уместны [41]. Это
долларами».
было неизбежно, учитывая обширную и трудоем-
кую логистику, связанную с протоколом роста и
дизайном эксперимента. Во-вторых, сама точ-
БЛАГОДАРНОСТИ
ность измерения могла зависеть от морфологии.
Ресурсы для разработки материала, представ-
Особи морского ежа изначально обладали шипа-
ленного в настоящей работе, были предоставле-
ми, охватывающими все их панцири, следова-
ны Университетом Макмастера (Department of
тельно, измерение h и d было сложной задачей.
Biology, SHARCNET и Origins Institute). Безгра-
Морские ежи потеряли колючки в течение вось-
ничное трудолюбие в создании ограничивающего
минедельного периода, чем облегчили измерение
аппарата было проявлено Р. Жилем и Д. Оадом; а
h и d. Это различие в измерениях до и после экс-
предложения по расширению экспериментов
перимента могло привести к уменьшению оцен-
были представлены И. Фернандесом и М. Спири-
ки роста в h и d, делая различия меньше, чем они
дисом.
могли бы быть на самом деле. Увеличение разме-
ров выборки и уточнение протокола измерений
приведет к получению данных, которые будут ха-
ФИНАНСИРОВАНИЕ РАБОТЫ
рактеризоваться более значительными экспери-
Финансовые ограничения были преодолены
ментальными эффектами и, таким образом, бо-
за счет средств, предоставленных Советом по
лее заметным изменением формы панциря, чем
естественным наукам и инженерным исследова-
описанная здесь фенотипическая адаптация.
ниям Канады (Discovery Grant 261590).
Потеря шипов указывает на стресс, поэтому
изменение метода применения геометрических
КОНФЛИКТ ИНТЕРЕСОВ
ограничений может увеличить рост, что даст
Авторы заявляют об отсутствии конфликта
больший эффект от эксперимента в будущих ис-
интересов.
следованиях. Информативный последующий
эксперимент включал бы освобождение экспери-
ментальных особей из геометрически ограничен-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
ных условий, чтобы проверить, сохраняют ли они
Настоящая работа не содержит описания ка-
измененные формы и если да, то как долго. Ана-
ких-либо исследований с использованием людей
лизы post mortem на сравнение панцирей в образ-
и животных в качестве объектов.
цах экспериментальной группы с панцирями в
образцах контрольной группы позволили бы вы-
яснить механизм роста, лежащий в основе изме-
СПИСОК ЛИТЕРАТУРЫ
нений формы. Это может проявляться в измене-
1. A. Kroh and А. В. Smith, J. Syst. Pal. 7, 147 (2010).
ниях наложения швов коллагеновых волокон, ро-
2. S. Horstadius, Biol. Rev. 14, 132 (1939).
сте пластин или и в том и в другом. Сшивание
3. R. T. Hinegardner, Biol. Bull. 137, 465 (1969).
коллагеновых волокон можно проанализировать
с применением сканирующей электронной мик-
4. J. E. Mazur and J. W. Miller, Ohio J. Sci. 71, 30 (1971).
роскопии, а рост пластин можно проанализиро-
5. S. B. George, J. M. Lawrence, and A. L. Lawrence,
вать, используя протоколы маркировки кальцеи-
Aquaculture 242, 217 (2004).
БИОФИЗИКА том 65
№ 3
2020
564
ЛИУ и др.
6. M. M. Smith, L. C. Smith, R. A. Cameron, and
24.
O. Ellers, A. Johnson, and P. Moberg, P. Biol. Bull.
L. A. Urry, J. Morphol. 269, 713 (2008).
195, 136 (1998).
7. R. Dalton, Nature 403, 125 (2000).
25.
M. P. Russell, J. Exp. Mar. Biol. Ecol. 220, 1 (1998).
8. C. S. Goodman and В. C. Coughlin, Proc. Natl. Acad.
26.
C. Fernandez, Mar. Ecol. Prog. Ser. 152, 45 (1997).
Sci. USA 97, 4424 (2000).
27.
M. J. West-Eberhard, J. Exp. Zool. 304B, 610 (2005).
9. R. Raff, Nat. Rev. Gen. 1, 74 (2000).
28.
D. W. Thompson, On Growth and Form (Cambridge
10. R. Raff and A. Love, J. Exp. Zool. (Mol. Dev. Evol.)
University, Cambridge, 1917).
302B, 19 (2004).
29.
D. С. C. Lau, S. С. K. Lau, P.-Y. Qian, and J.-W. Qiu,
11. L. H. Hyman, The Invertebrates: Echinodermata (Mc-
J. Shell. Res. 28, 383 (2009).
Graw-Hill Book Company, New York, 1955).
30.
J. Dafni, J. Exp. Mar. Biol. Ecol. 67, 1 (1983).
12. A. B. Smith, Echinoid Palaeobiology (Allen & Unwin,
London, 1984).
31.
J. Dafni and J. Erez, Mar. Biol. 95, 275 (1987).
13. M. Telford, Zoomorphology 105, 114 (1985).
32.
J. Cohen, Psych. Bull. 112, 155 (1992).
14. A. Johnson, O. Ellers, J. Lemire, et al., Proc. R. Soc.
33.
T. A. Ebert, J. Nat. Hist. 22, 1407 (1988).
Lond. В 269, 215 (2002).
34.
S. J. Gould, Paleobiology 17, 411 (1991).
15. M. Abou Chakra and J. R. Stone, Paleobio. 37, 686
35.
J. Dafni, J. Exp. Mar. Biol. Ecol. 47, 259 (2011).
(2011).
36.
T. Motokawa, Biol. Rev. 59, 255 (1984).
16. R. T. Jackson, Boston Soc. Nat. Hist. Mem. 1 (1912).
37.
C. Wilkie, Mar. Behav. Physiol. 11, 1 (1984).
17. B. David and R. Mooi, C. R. Acad. Sci. Paris Ser. 3
319, 577 (1996).
38.
C. Wilkie, in Echinoderm Studies, Ed. by M. Jangoux
and J. M. Lawrence (Balkema, Rotterdam, 1996),
18. B. David and R. Mooi, in Echinoderms: San Francisco,
pp. 61-102.
ed. by R. Mooi and M. Telford (Balkema, Rotterdam,
1998), pp. 21-28.
39.
B. Lelievre, M. Telford, and O. Ellers, in Echinoderm
Research 1995. Proc. 4th Eur. Echinoderms Colloquium,
19. R. Mooi and B. David, Paleontol. Soc. Pap. 3, 305
London, ed. by R. H. Emson, A. B. Smith, and
(1997).
A. C. Campbell (Balkema, Rotterdam, 1995), pp. 103-
20. O. Ellers, Proc. Roy. Soc. Lond. В 254, 123 (1993).
109.
21. J. Dafni, Paleobiology 12, 143 (1986).
40.
M. Telford and O. Ellers, Invert. Biol. 116, 255 (1997).
22. A. S. Johnson and O. Ellers, in Echinoderms: San Fran-
41.
R. R. Sokal and F. J. Rohlf, Biometry, 3rd ed.
cisco, Ed. By R. Mooi and M. Telford (Balkema, Rot-
(W. H. Freeman, N. Y., 1995).
terdam, 1998), p. 705.
42.
J. S. Pearse and V. B. Pearse, Am. Zool. 15, 731 (1975).
23. E. McCarron, G. Burnell, and G. Mouzakitis, Aqua-
culture 288, 83 (2009).
43.
K. Markel, Zoomorphology 97, 31 (1981).
Phenotypic Accommodation in Sea Urchins Grown under Geometric Constraints
R. Liu, Sh. Pedlar, A. Oad, S. McDonald, and J. Stone
McMaster University, 1280 Main Street West, Hamilton ON, L8S 4L8, Canada
Sea urchins and their relatives are taxonomically grouped as Echinoidea (phylum Echinodermata). Echinoids
exhibit considerable modifications of their pentaradially symmetric body plan phenotypes, from spheroid
tests (skeletons) in sea urchins to discoid tests in sand dollars. Researchers have speculated as to morpholog-
ical evolutionary transitions among these phenotypes without reaching consensus. We designed and per-
formed an experiment in which sea urchins grew under geometric constraints manifested through restricted
vertical spatial dimension, physically limiting growth in height. Individuals in this treatment group became
flatter than those in a control group after 8 weeks. These findings can be considered as indicative of pheno-
typic accommodation, an underappreciated phenomenon that may constitute the first step in novel pheno-
type evolution. We present preliminary speculations in regard to phenotypically accommodated changes in
shape and morphological evolutionary relationships among echinoids. We demonstrate a plausible initiating
mechanism in a morphological transition, which can contribute to theoretical discussions about phenotypic
evolution, emphasizing the role physical factors play when complex biological systems change.
Keywords: development; evolution, morphology, phenotypic plasticity
БИОФИЗИКА том 65
№ 3
2020