БИОФИЗИКА, 2020, том 65, № 3, с. 565-573
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 62-506: 591.181
ВОСПРИЯТИЕ И СПЕЦИФИЧНОСТЬ РЕАГИРОВАНИЯ
МЕДОНОСНЫХ ПЧЕЛ, БУМАЖНЫХ ОС И РЫЖИХ ЛЕСНЫХ МУРАВЬЕВ
НА НИЗКОЧАСТОТНОЕ ЭЛЕКТРИЧЕСКОЕ ПОЛЕ
© 2020 г. Е.К. Еськов
Российский государственный аграрный заочный университет,
143900, Балашиха Московской области, ул. Ю. Фучика, 1
E-mail: ekeskov@yandex.ru
Поступила в редакцию 10.02.2019 г.
После доработки 18.02.2020 г.
Принята к публикации 19.02.2020 г.
Пчелы, осы и муравьи не имеют специализированных рецепторов восприятия электрического
поля. Адекватное реагирование на природные электрические поля у пчел и муравьев связано с
воздействием атмосфериков, усиливающихся при приближении грозового фронта. К восприятию
насекомыми низкочастотных электрических полей высокой напряженности имеют отношения
первичные преобразователи механорецепторов, реагирующих на смещение. Неспецифический
механизм восприятия электрических полей основан на раздражении насекомых наведенными
токами, протекающими в местах их контактирования между собой и/или токопроводящими
поверхностями. Частотная характеристика чувствительности к электрическому полю определяется
в основном величиной наводимого им тока и интенсивностью его контактного действия. Величина
тока, наводимого в покровах тела насекомых, нелинейно связана с частотой электрического поля.
Область с наибольшей чувствительностью к электрическим полям приходится примерно на 500 Гц,
что согласуется с максимальной величиной наводимого тока. При этом порог чувствительности к
электрическому полю у ос составляет около 0.04, у пчел - 0.45 кВ/м. Муравьи реагируют
замедлением локомоций на действие электрического поля напряженностью 7-10 кВ/м. Магнитные
поля и ионизация, сопутствующие генерации электрического поля, напряженность которого
достигает 15-20 кВ/м, не стимулируют изменения поведения насекомых.
Ключевые слова: пчелы, осы, муравьи, электрическое поле, чувствительность, аномалии поведения,
адекватное реагирование.
DOI: 10.31857/S0006302920030175
цепторов ЭП, влияют на их физиологическое со-
Биосфера Земли подвергается природным
стояние [1-6].
электромагнитным воздействиям широкого ча-
стотного диапазона - от медленных изменений
В зависимости от образа жизни и уровня орга-
магнитного и электрических полей до гамма-из-
низации животные по-разному реагируют на ЭП,
лучений. Природные электромагнитные процес-
защищаясь от их неблагоприятного воздействия.
сы дополняются возрастающими по диапазону и
Высокоорганизованные социальные насекомые
от различных экстремальных воздействий защи-
интенсивности антропогенными воздействиями
на биосферу. Антропогенные электрические
щаются обычно скоординированными усилиями
больших групп или всех взрослых особей семьи.
(ЭП) и магнитные поля (МП) по интенсивности
На типичные физические или биотические сти-
сравнимы с естественными ЭП и МП, а в некото-
мулы у социальных насекомых прослеживается
рых ситуациях превосходят их.
адекватное реагирование. Так, на изменения
У животных ЭП и МП воздействуют на элек-
внешней температуры пчелы и осы, находящиеся
трические процессы, протекающие в клетках,
в своих гнездах, реагируют изменением генера-
тканях и органах. Поэтому природные и антропо-
ции и выделения тепла [7-10]. Рыжие лесные му-
равьи изменяют наклон поверхности холмика к
генные ЭП и МП, независимо от наличия или от-
солнечным лучам [9, 11, 12]. Коллективные обо-
сутствия у животных специализированных ре-
ронительные реакции стимулирует у всех соци-
Сокращения: ЭП - электрические поля, МП - магнитные
альных насекомых нападение на их жилища вра-
поля, ЛЭП - линия электропередачи.
гов и грабителей. При приближении грозового
565
566
ЕСЬКОВ
фронта, чему сопутствует возрастание интенсив-
частности, одна пчела, находящаяся на изолиро-
ности атмосфериков [12], муравьи заделывают
ванной поверхности, независимо от ориентации
входные отверстия муравейников, а пчелы пре-
тела по отношению к силовым линиям ЭП не
кращают полеты и возвращаются в ульи [9-12].
проявляет никаких признаков реагирования на
низкочастотное ЭП высокой напряженности, до-
Медоносные пчелы наряду с защитными реак-
стигающей 50 кВ/м. Незначительная активиза-
циями на переменные ЭП приспособились ис-
ция локомоций и соответственно увеличение по-
пользовать электростатическое поле в системе
требления О2 происходит под действием ЭП на-
внутриульевых коммуникаций. Статическое по-
ле, возникающее на теле пчелы-сигнальщицы
пряженностью 100 кВ/м.
при трении о поверхность сот, используется мо-
Соответственно при увеличении численности
билизуемыми ею пчелами. Они среди множества
пчел, поддерживающих между собой тактильные
других пчел, находящихся на сотах, по отклоне-
контакты, степень их реагирования на ЭП одина-
нию антенн под действием статического заряда
ковой частоты и напряженности возрастает. У
обнаруживают сигнальщицу и поддерживают с
двух пчел, находящихся на электрически изоли-
нею дистанцию, необходимую для восприятия
рованной поверхности (в стеклянном сосуде), ЭП
относительно слабого акустического сигнала.
частотой 500 Гц при напряженности 50 кВ/м сти-
Посредством этого сигнала пчела-сигнальщица
мулирует активизацию и увеличение потребле-
информирует приблизившихся к ней пчел о рас-
ния О2 в 1.5 ± 0.14 раза, а у десяти пчел - в
стоянии до обнаруженной цели полета, например
1.8 ± 0.17 раза. Резкую активизацию у 100 пчел
источника корма или воды [14].
стимулирует ЭП напряженностью 14 ± 3 кВ/м.
Что касается антропогенных ЭП и МП, то они
Частотная характеристика чувствительности
для насекомых и других видов животных являют-
пчел к ЭП установлена на их семьях [10, 17, 18].
ся новыми, нетипичными факторами среды. На
Семьи, содержавшие от 5 до 30 тыс. взрослых ра-
них высокоорганизованные животные с большой
бочих особей, находились в экранированной аку-
продолжительностью жизни не могли приобре-
стически изолированной камере. Применение
сти адекватных форм реагирования.
акустической изоляции пчелиных семей требова-
Настоящей работой предпринят сравнитель-
лось для точной регистрации изменений звуково-
ный анализ реагирования на ЭП трех видов пере-
го шума семьи, регистрируемого в диапазоне
пончатокрылых насекомых (Hymenoptera), раз-
400-500 Гц. С усилением этого частотного диапа-
личающихся по образу жизни, - медоносных
зоны шумов пчелиной семьи связана активиза-
пчел, бумажных ос и рыжих лесных муравьев. Эти
ция пчел [19]. Таким же способом на семьях зем-
виды социальных насекомых, приспособленные
ляных ос (Paravespula vulgaris L.), содержавших от
к разным условиям жизни, сооружают свои гнез-
1200 до 2000 взрослых особей, изучали их чув-
да из материалов, различающихся по электропро-
ствительность к ЭП.
водности. Основу гнездовых построек у пчел
Пчелы и осы, находящиеся в своих гнездах,
представляют восковые, а у ос - бумажные соты.
обладают максимальной чувствительностью к ЭП
Муравьи сооружают гнезда в земле, возводя зем-
частотой 500 ± 50 Гц. Но семьи ос, отличаясь от
ляные холмики, включающие растительные ча-
пчел меньшей численностью взрослых особей,
стички.
примерно на порядок превышают пчелиные се-
Изучение специфики реагирования пчел, ос и
мьи по чувствительности к ЭП. На частоте около
500 Гц порог чувствительности ос находится на
муравьев на низкочастотное ЭП важно для пони-
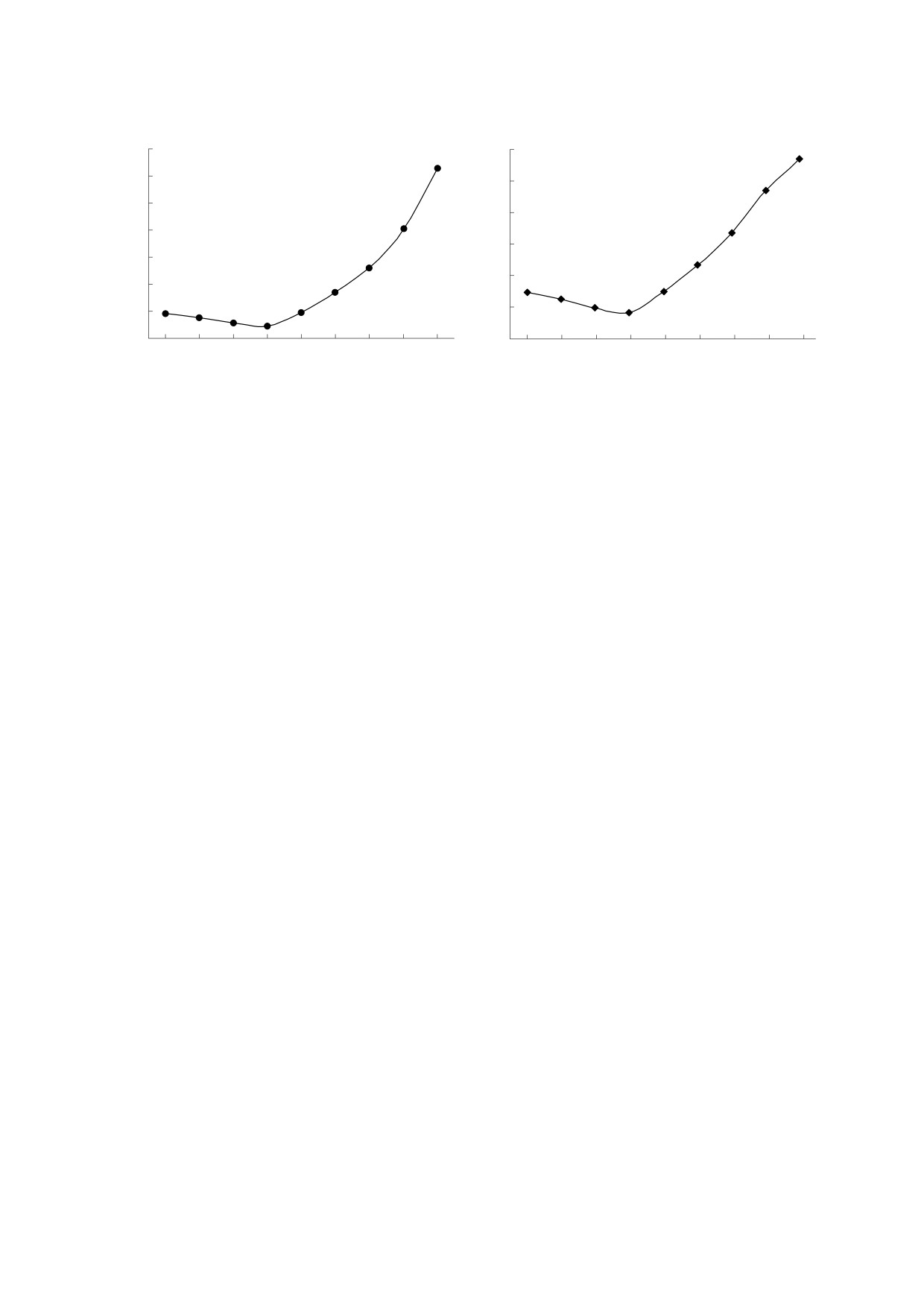
уровне 0.03-0.05 кВ/м (рис. 1б), а у пчел - 0.4-
мания физиологических механизмов восприятия
этого стимула животными, отличающимися по
0.5 кВ/м (рис. 1а).
образу жизни и экологической валентности. У
Чувствительность к ЭП неравномерно изме-
медоносных пчел семья, а не особь, представляет
няется в ответ на понижение или повышение его
собою размножающуюся биологическую едини-
частоты. И у ос, и у пчел при уменьшении частоты
цу, подлежащую действию естественного отбора.
ЭП в 50 раз (от 500 до 10 Гц) порог чувствительно-
Бумажные осы живут сезонными семьями, кото-
сти к этому стимулу возрастает примерно в два
рые размножаются половозрелыми самками. Ры-
раза. При повышении частоты ЭП в десять раз (от
жие лесные муравьи, образуя большие семьи, мо-
500 до 5000 Гц) порог чувствительности у ос воз-
гут размножаться почкованием или половозре-
растает в четыре раза, а у пчел - в двенадцать раз
лыми особями.
(рис 1а,б).
Рыжие лесные муравьи (Formica rufa L.) харак-
теризуются относительно низкой чувствительно-
ЧУВСТВИТЕЛЬНОСТЬ
стью к ЭП. Если порог чувствительности опреде-
К ЭЛЕКТРИЧЕСКИМ ПОЛЯМ
лять по понижению локомоторной активности,
Для пчел (Apis mellifera L.) установлены груп-
то некоторое ее замедление у муравьев, выходя-
повые эффекты реагирования ЭП [10, 15, 16]. В
щих из муравейника, наблюдается под действием
БИОФИЗИКА том 65
№ 3
2020
ВОСПРИЯТИЕ И СПЕЦИФИЧНОСТЬ РЕАГИРОВАНИЯ
567
(а)
(б)
Е, кВ/м
Е, кВ/м
7
0.20
6
0.17
5
0.14
4
0.11
3
0.08
2
1
0.05
0
0.02
10
50
100
500 1000 2000 3000 4000 5000
10
50
100
500 1000 2000 3000 4000 5000
f, Гц
f, Гц
Рис. 1. Частотная зависимость чувствительности медоносных пчел (а) и бумажных ос (б) к электрическому полю (по
работе [17], с изменениями).
ЭП частотой 500 Гц при напряженности, достига-
вышенной двигательной активностью. Они при-
ющей 8-10 кВ/м [20].
ближаются и вступают в кратковременные так-
тильные контакты не только с подлетающими
пчелами, что характерно для сторожей, но и с
РЕАГИРОВАНИЕ
особями, выходящими из улья. Тактильным кон-
НА ЭЛЕКТРИЧЕСКИЕ ПОЛЯ
тактам всегда сопутствует повышение локомо-
Находясь в разных условиях жизни и обладая
торной активности пчел.
разной чувствительностью к низкочастотному
На пороговые и близкие к ним напряженности
ЭП, пчелы, осы и муравьи по-разному реагируют
ЭП пчелиная семья реагирует незначительным
на этот стимул. Поскольку порог чувствительно-
повышением интенсивности спектральных со-
сти этих насекомых к ЭП приходится примерно
ставляющих в диапазоне 400-500 Гц. Этот диапа-
на 500 Гц, то эта частота использована для стиму-
зон акустического шума пчелиной семьи связан с
ляции ответных реакций.
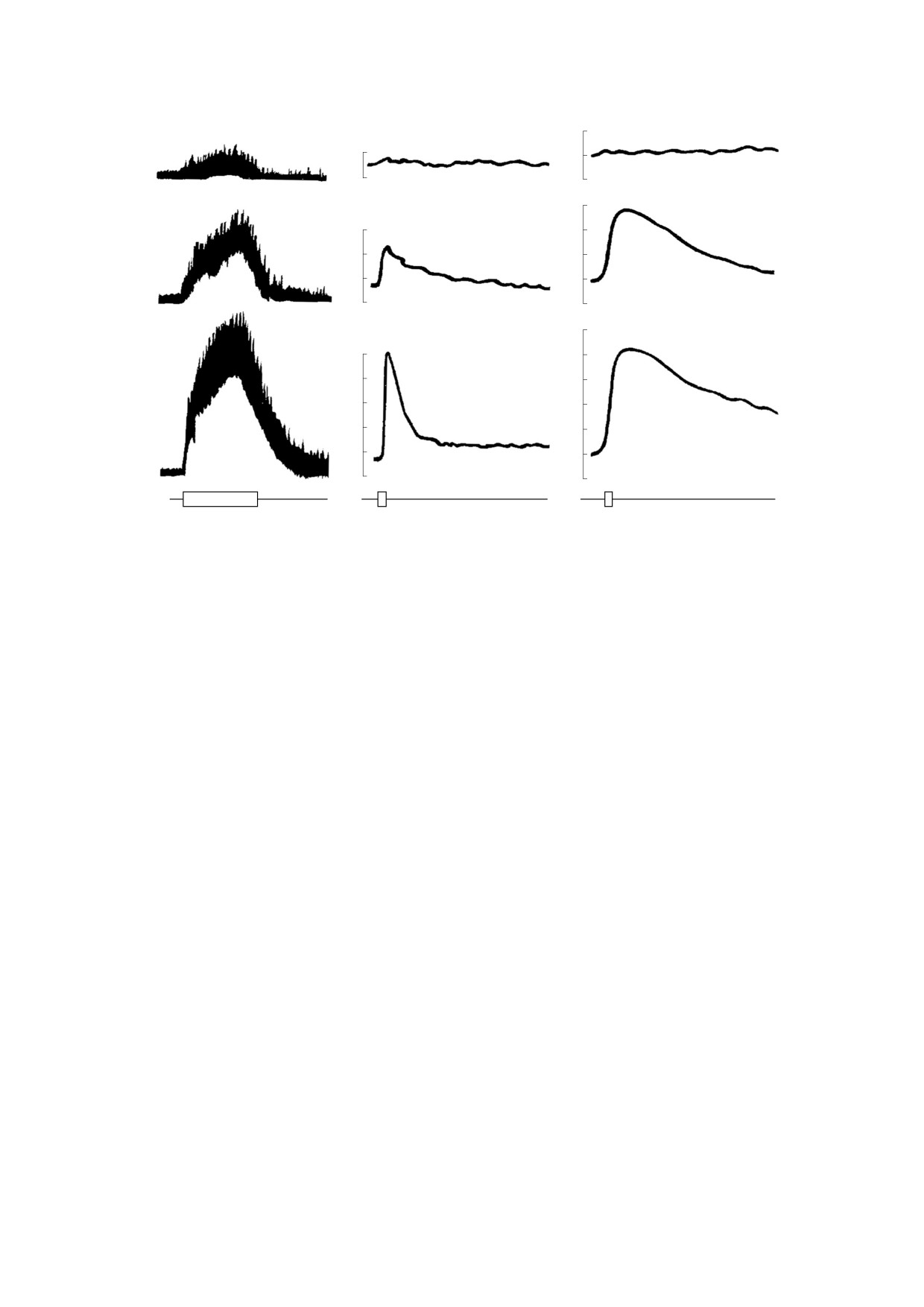
активизацией пчел [19]. Всего на 0.5-0.9 дБ воз-
Пчелы. Пчелы, находящиеся в гнезде, реагиру-
растает интенсивность спектральных составляю-
ют на ЭП повышением локомоций и активности
щих пчелиного шума в диапазоне 450 ± 50 Гц под
метаболизма [21]. Заметную активизацию локо-
действием ЭП напряженностью 1.5-2 кВ/м. При
моций стимулирует ЭП напряженностью
1-
увеличении напряженности ЭП до 5 и 10 кВ/м
2 кВ/м. Возбужденные пчелы начинают выходить
интенсивность этих составляющих возрастает со-
из улья, когда напряженность ЭП достигает 10-
ответственно на 1.7-1.8 и 2.1-2.3 дБ (рис. 2а).
15 кВ/м, а при 20-30 кВ/м сотни возбужденных
При высокой напряженности ЭП, достигаю-
пчел выходят на переднюю стенку улья. Некото-
щей 35 ± 3 кВ/м, сильному возбуждению пчел со-
рая их часть взлетает и кружится на расстоянии
путствует значительное изменение интенсивно-
1-5 м от улья. Летающие пчелы набрасываются и
сти и спектральной структуры акустического шу-
жалят людей и животных, оказавшихся вблизи
ма пчелиной семьи. Частота пика интенсивности,
улья.
локализующегося в области 400-500 Гц, повыша-
Через 4-7 мин от начала воздействия на пче-
ется на 25-35 Гц, а общая интенсивность шума
линую семью ЭП напряженностью 20-30 кВ/м
возрастает на 4-5 дБ. К исходному состоянию
локомоторная активность пчел начинает пони-
интенсивность и спектральная структура пчели-
жаться. С этого времени ранее гиперактивные
ных звуков возвращаются через 3.6-4.2 ч после
пчелы группируются на поверхности сот, образуя
отключения ЭП [10, 19].
неровные ряды, ориентированные преимуще-
Активизации локомоций под действием ЭП
ственно вдоль или под небольшим углом к сило-
сопутствует повышение температуры и выделе-
вым линиям ЭП. В нижней части сот и у дна улья
ния СО2 (рис 2б,в). Незначительное повышение
пчелы образуют беспорядочные малоподвижные
скопления.
температуры (на 0.1-0.3°С) и СО2 (на 0.05-0.1%)
Поведение пчел, находящихся у летка во время
над центром расплодной зоны гнезда происходит
действия ЭП, сходно с поведением сторожей при
под действием ЭП напряженностью 1.5-2 кВ/м.
При повышении напряженности ЭП до 5 кВ/м
нападении на семью врагов или грабителей. Но в
отличие от сторожей эти пчелы отличаются по-
температура и концентрация СО2 в указанной зо-
БИОФИЗИКА том 65
№ 3
2020
568
ЕСЬКОВ
(а)
(б)
(в)
21
1
20
0
19
23
3
22
2
21
1
20
0
19
25
5
24
4
23
3
22
2
21
1
20
0
19
ЭП
ЭП
ЭП
3 мин
30 мин
30 мин
Рис. 2. Изменение интенсивности звуков, генерируемых пчелиной семьей в полосе 400-500 Гц (а), концентрации
СО2 в центре гнезда (б) и температуры над гнездом (в) под действием ЭП частотой 500 Гц при его напряженности 1.5-
2 кВ/м (верхний ряд), 5 кВ/м (средний ряд) и 10 кВ/м (нижний ряд); ЭП - периоды действия поля [21].
не гнезда возрастает на 2.6-2.8°С и 1.6-1.8%, а
Длительное действие ЭП не вызывает у пчел
при 10 кВ/м - соответственно на 4.1-4.3°С и 3.7-
полного привыкания, хотя активность реагирова-
4.1%.
ния со временем уменьшается [10, 23]. Действие
ЭП, напряженность которого примерно на поря-
Значительные изменения микроклимата в
док превосходит пороговое значение чувстви-
пчелином гнезде происходят под действием ЭП
тельности, дестабилизирует внутригнездовую
напряженностью 20-25 кВ/м. На 10-минутное
температуру. Например, у пчелиных семей, кото-
воздействие такого стимула температура в центре
рые в течение 45 суток находились под крайней
гнезда повышается на 7-9°С, а концентрация
фазой линии электропередачи (ЛЭП) напряже-
СО2 - на 4.4-6,1%. Через 2-3 ч после отклю-
нием 500 кВ при напряженности ЭП около
чения поля температура снижается всего на 0.5-
7.5 кВ/м, температура в разных зонах гнезда скач-
кообразно повышалась на 2-4°С, а затем опуска-
1.3°С и возвращается к исходному значению че-
лась до исходных значений. Периоды подъема и
рез 15-18 ч. Но концентрация СО2 нормализуется
понижения температуры не имели строгой упо-
всего за 50-70 мин.
рядоченности. Продолжительность той и другой
Эффективность ЭП зависит от его генерации в
фаз находилась в пределах от нескольких минут
до десятков часов.
форме пакетов импульсов или непрерывных си-
нусоидальных колебаний. При подаче ЭП в им-
ЭП ЛЭП влияет также на дестабилизацию
пульсном режиме имеет значение скважность им-
концентрации СО2 в разных зонах пчелиного
пульсации. В частности, ЭП частотой 300 Гц при
гнезда. В дни, благоприятные для полетов, кон-
напряженности 8 кВ/м, подаваемое в форме си-
центрация СО2 в центральной части гнезда в те-
нусоидальных колебаний в течение 10 мин, сти-
чение ночи повышалась в среднем до 1.8 ± 0.4%, а
мулирует повышение температуры над гнездом
в прилетковой зоне - до 1.1 ± 0.3%. К середине
пчелиной семьи на 2.1 ± 0.4°С. За такое же время
дня в центральной части гнезда происходило по-
ЭП, подаваемое в форме импульсов длительно-
нижение концентрации СО2 примерно в два раза,
стью 30 мс со скважностью, равной 2, стимулиру-
а в прилетковой- повышение в полтора раза. В то
ет повышение пчелами температуры на
же самое время в гнездах семей, находившихся в
1.2 ± 0.2°С, а при скважности, равной 10, - на
20-50 м от линии, суточные колебания темпера-
0.4 ± 0.1°С [21, 22].
туры не выходили за пределы 0.23-0.37% [23].
БИОФИЗИКА том 65
№ 3
2020
ВОСПРИЯТИЕ И СПЕЦИФИЧНОСТЬ РЕАГИРОВАНИЯ
569
К специфическим реакциям пчел на длитель-
рировать кормушку, подвергаемую действию ЭП,
ное воздействие ЭП относится интенсивное по-
и собираются на соседней кормушке.
крытие воском и прополисом леткового прохода
Осы. У ос, находящихся в гнезде, резкое повы-
и примыкающих к нему частей улья. Такое пове-
шение двигательной активности стимулирует ЭП
дение свойственно пчелам в конце лета - начале
напряженностью 1-2 кВ/м. Возбуждение взрос-
осени при подготовке к зимовке. Но в прилетко-
лых особей происходит в процессе тактильных
вой зоне ульев, находившиеся в 10-14 м от край-
контактов. Повышение локомоций приводит к
ней фазы ЛЭП и под нею, к концу лета пчелы на-
повышению внутригнездовой температуры на
копили соответственно по 11 ± 1.3 и 37 ± 4.6 г вос-
0.6-0.9°С [18].
ко-прополисовой смеси. В ульях семей,
В отличие от пчел, у ос увеличение напряжен-
находившихся в это время в 50-60 м от ЛЭП, мас-
ности ЭП не приводит к повышению внутригнез-
са накопленной воско-прополисовой смеси со-
довой температуры, потому что активизировав-
ставляла всего 0.9 ± 0.3 г [24]. Иногда пчелы се-
шиеся осы вылетают из гнезда. Под действием
мей, находящихся под ЛЭП, полностью заделы-
ЭП напряженностью 6-8 кВ/м все взрослые осы
вают прополисом летковые проходы, обрекая тем
покидают гнездо в течение одной-трех минут.
самым себя на гибель [25].
Поэтому температура в гнезде понижается.
ЭП ЛЭП влияет на поведение пчел, вылетаю-
Осы, вылетающие из гнезда, кружатся у его
щих из ульев [23]. Им свойственна повышенная
леткового входа. Летающие осы не проявляют
агрессивность и неупорядоченность вылетов
признаков агрессии. После отключения ЭП осы в
большими группами, которые кружатся некото-
течение двух-трех минут возвращаются в гнездо.
рое время на расстоянии нескольких метров от
Многократные повторные включения ЭП стиму-
ульев. Вылеты этих пчел не связаны и не имеют
лируют стереотипные реакции ос без выражен-
сходства с ориентировочными и очистительными
ных признаков адаптации.
облетами, а полеты пчел-фуражиров отличаются
Муравьи. Обнаружено специфическое реаги-
пониженной продуктивностью. Так, если у фура-
рование муравьев на ЭП частотой от 50 до 500 Гц
жиров, возвращающихся в ульи, которые находи-
при размещении одного из электродов (металли-
лись в 50-60 м от ЛЭП, масса содержимого медо-
ческой пластины размером 40×40 см) над мура-
вых зобиков составляла 28.8 ± 3.9 мг, то под край-
вейником параллельно поверхности почвы и за-
ней фазой линии она была меньше примерно в
глублении в почву на 40 см второго электрода
полтора раза.
[20]. В зависимости от напряженности ЭП пове-
дение муравьев существенно изменяется. В част-
Пчелы, находящиеся у источника кома, поки-
ности, через 7-10 с после включения ЭП напря-
дают его под действием ЭП. Группы из 50-
женностью 7-10 кВ/м происходит замедление
120 пчел, наполняющие медовые зобики 50-60%-
локомоций муравьев, находившихся на поверх-
м раствором сахарозы, под действием ЭП напря-
женностью 40-50 кВ/м взлетают в течение 3-10 с.
ности муравейника. В дальнейшем некоторое по-
вышение активности муравьев, выражающееся в
Летающие пчелы кружатся между электродами и
нормализации их поведения, происходит через
за их пределами, не проявляя признаков агрес-
15-20 мин после выключения ЭП.
сии. После отключения ЭП пчелы в течение 0.5-
1.5 мин вновь собираются на кормушке [10].
Под действием ЭП напряженностью
9-
11 кВ/м муравьи, выходившие из муравейника,
Поведение пчел у кормушки не меняется при
замедляют движение. Это сходно с поведением
десяти кратных включениях ЭП с 5-, 20- и 30-ми-
пчел, оказавшихся па поверхности металличе-
нутными интервалами. При длительном одно-
ской пластины, подключенной к конденсатору,
кратном включении ЭП пчелы, покружившись
заряженному от постоянного источника тока на-
над кормушкой в течение трех-пяти минут, начи-
пряжением 300 В [18]. Но, в отличие от пчел, му-
нают возвращаться на нее и наполнять зобики.
равьи, медленно передвигаясь к вершине холми-
Этих пчел возбуждают взаимные соприкоснове-
ка, останавливаются и принимают динамические
ния и стимулируют взлеты. Но покружившись
позы угрозы - переворачиваются на спинку и
некоторое время, пчелы вновь возвращаются на
поднимают вверх брюшко. Такую позу муравьи
кормушку.
удерживают в течение одной-трех минут, а затем
На действие ЭП у пчел-фуражиров быстро вы-
продолжают движение к вершине холмика.
рабатывается отрицательный условный рефлекс.
Повышение напряженности ЭП до
24-
При наличии двух кормушек, расположенных на
26 кВ/м активизирует у муравьев оборонитель-
расстоянии 1 м, пчелы быстро начинают отличать
ные реакции. Муравьи собираются на вершине
кормушку, которая находится в ЭП. Для выра-
муравейника, поднимают вверх брюшко и
ботки отрицательного рефлекса достаточно 8-
выбрызгивают струйки кислотного секрета, что
14 включений ЭП. После этого все пчелы, кото-
используется обычно для защиты от нападения
рым известно место подкормки, начинают игно-
врагов. Но эта типичная защитная реакция при
БИОФИЗИКА том 65
№ 3
2020
570
ЕСЬКОВ
27
(а)
0.7
(б)
0.6
26
0.5
3
25
3
0.4
2
24
2
0.3
1
23
1
0.2
22
0.1
15
30
45
60
75
90
120
150
15
30
45
60
75
90
120
150
Время, ч
Время, ч
Рис. 3. Изменение температуры (а) и концентрации СО2 (б) на глубине 10 (кривая 1), 20 (кривая 2) и 40 мм (кривая 3)
от вершины муравейника на ЭП напряженностью 10 кВ/м, включенного с 30-й по 60-ю минуту (по работе [20], с
изменениями).
воздействии ЭП дополняется взаимной агрессив-
Сходные изменения температуры и содержа-
ностью муравьев, приводящей к гибели части из
ния СО2 в муравейнике происходили под дей-
них. За 15 мин действия ЭП погибало около 3%
ствием ЭП напряженностью 25 кВ/м. Эти изме-
особей, собиравшихся на вершине муравейника.
нения происходят примерно в таких же пределах,
Этого не наблюдается при разорении муравейни-
как при воздействии ЭП напряженностью
ка, активизирующего защитные реакции мура-
10 кВ/м, но с большей скоростью, что обуславли-
вьев.
валось активизацией локомоций.
В муравейнике под действие ЭП происходит
изменение температуры и содержания СО2
ФИЗИЧЕСКИЕ ПРОЦЕССЫ,
(рис. 3). При температуре внешнего воздуха и по-
СОПУТСТВУЮЩИЕ ГЕНЕРАЦИИ ЭП
верхности муравейника в пределах 19-23°С под
К процессам, сопутствующим генерации ЭП,
влиянием тридцатиминутного воздействия ЭП
относится возникновение магнитного поля (МП)
напряженностью 10 кВ/м температура в средней
и увеличение концентрации аэроионов. Возмож-
части холмика повышается в среднем на
ны также вибрации опорных субстратов (стенок
1 ± 0.15°С, в верхней - на 0.5 ± 0.09°С, а в
ульев, сотов и др.), на которых находятся пчелы.
нижней - опускается на 0.3 ± 0.06°С. В течение
По некоторым сведениям пчелы, изолированные
15 мин после отключения ЭП температура в верх-
от семьи, предпочитают локализоваться в обла-
ней части холмика понижается на 0.2 ± 0.07°С, в
сти повышенного магнитного потока [26]. Порог
средней - на 0.8 ± 0.14°С и в нижней - на
чувствительности пчел к МП находится на уровне
0.7 ± 0.12°С. В последующие 30-45 мин активно-
260 мТл [27]. Имеются сведения, согласно кото-
му передвижению муравьев в глубь муравейника
рым, отрицательные аэроионы влияют на повы-
и из него на поверхность холмика сопутствуют
шение двигательной активности пчел [28,29].
периоды повышения, а затем понижения темпе-
Магнитное поле. Величина магнитного поля,
ратуры (рис. 3а).
сопутствующего действию ЭП, определяется вы-
Динамике температуры в муравейнике под
πfr
ражением
B
=
E
sin(2πft),
где r - расстояние
действие ЭП сопутствует изменение концентра-
2
C
ции СО2 (рис. 3б). За 30 мин действия ЭП содер-
до прямой, проходящей через центры электродов
жание СО2 в верхней и средней частях холмика
(металлические пластины, размером 40 × 40 см);
возрастает примерно в два раза. После отключе-
f - частота ЭП; Е - его напряженность; С - ско-
ния ЭП концентрация СО2 возвращается к ис-
рость света; t - время [30]. Отсюда при воздей-
ствии на семью ЭП частотой 500 Гц и напряжен-
ходному уровню через 40-60 мин. В нижней ча-
ностью 15 кВ/м величина МП составляет около
сти холмика в период действия ЭП содержание
СО2 возрастает в среднем в 1.2 раза, а через 40-
1.3 · 10-10 Тл, или 1.3 · 10-6 Гс. МП Земли на сред-
них широтах приближается к 0.5 Гс, а его суточ-
50 мин после отключения поля уменьшается в
1.6 раза.
ные вариации достигают 10-3 Гс [31], что пример-
БИОФИЗИКА том 65
№ 3
2020
ВОСПРИЯТИЕ И СПЕЦИФИЧНОСТЬ РЕАГИРОВАНИЯ
571
но на три порядка превышает МП, возникающее
ного субстрата, возникающие под действием ЭП,
при генерации ЭП напряженностью 15 кВ/м.
напряженность которого многократно превосхо-
дит порог чувствительности к этому стимулу.
На суточные вариации МП Земли реагируют
пчелы-сигнальщицы (танцовщицы). Они в тече-
ние дня «ошибаются» в указании направления на
ЗАКЛЮЧЕНИЕ
указываемую цель полета, отклоняя от нее до 30-
35° направление прямолинейного пробега. Роль
Природные ЭП и МП существовали на Земле
влияния вариаций МП на «ошибки» пчел-сиг-
задолго до появления на ней жизни. В процессе ее
зарождения и эволюции эти физические факто-
нальщиц доказана тем, что в случае компенсации
ры, взаимодействуя с биоэлектрическими про-
МП Земли до 0-5% угол максимального отклоне-
цессами, оказывали влияние на состояние и
ния направления от реального уменьшается в
функционирование организмов.
среднем в 3.4 раза [32].
К флуктуациям некоторых естественных ЭП
Но на МП, соответствующие пороговым зна-
животные приобрели адаптацию, что выражает-
чениям чувствительности пчел и ос, эти насеко-
ся, например, в реагировании пчел и муравьев на
мые не реагируют. Не обнаружено, чтобы пчелы,
усиление атмосфериков, происходящее при при-
содержавшиеся по 200 особей в стеклянных ци-
ближении грозового фронта. Для этих насекомых
линдрах, помещаемых в соленоид, реагировали
усиление атмосфериков служит сигналом при-
на МП частотой 50-1000 Гц, индукция которого
ближения неблагоприятных погодных условий,
достигала 10 Гс [33]. Это на 107 или 108 превосхо-
что стимулирует у пчел инстинкт хоминга, а у му-
дило индукцию МП, которая сопутствует генера-
равьев - защиты муравейника от дождя [9, 12].
ции ЭП, стимулирующей активное реагирование
Преимущественно неадекватно по отноше-
пчел или ос.
нию к экологической ситуации пчелы, осы и му-
Ионизация. При воздействии на пчелиную се-
равьи реагируют на интенсивные искусственные
мью ЭП напряженностью 15 кВ/м в течение 15 мин
ЭП антропогенной природы. Низкочастотные
концентрация легких, средних и тяжелых ионов с
ЭП высокой напряженности стимулируют у них
подвижностью свыше 0.001 см/(В с) не превышает
различные аномалии поведения. Насекомые рез-
5 · 103/см3 (измерение проведено счетчиком аэрои-
ко активизируются, покидают свои жилища и
онов ИТ-8401).
проявляют немотивированную агрессивность. У
Судя по внутригнездовой температуре, интен-
пчел некоторое проявление адаптации к ЭП вы-
сокой напряженности при длительном воздей-
сивности и спектральной структуре звуков, гене-
рируемых пчелами, они не реагировали на искус-
ствии выражается в обеспечении электрической
ственное (создаваемое тритиевым ионизатором)
изоляции леткового прохода, который усиленно
увеличение в улье концентрации аэроионов до
покрывается
воско-прополисовой
смесью
[23,24]. Это защищает пчел, находящихся под ВЛ
2 · 106 см3 [33]. Осы также не реагировали на
ЛЭП, от раздражения наведенными токами при
ионизацию внутригнездового пространства.
соприкосновениях с поверхностью леткового
Вибрации опорного субстрата. Отношение пчел
прохода. На теле пчелы, подлетающей к улью под
к вибрациям опорного субстрата прослежено на
ЛЭП с напряжением 500 кВ, наводится ток около
пчелах, содержавшихся примерно по 200 особей в
4 · 10-4 нА, а под ЛЭП с напряжением 750 кВ -
энтомологических садках размером 13 × 116 см.
3 · 10-3 нА [35].
Зависимость колебаний деревянных стенок сад-
ков от частоты и напряженности ЭП выражается
В восприятии насекомыми ЭП участвуют спе-
циализированные механорецепторы и/или
уравнением A = 0.036A1 f-2, где А - амплитуда
неспецифические структуры. Один из механиз-
вибросмещения, А1 - изменение амплитуды виб-
мов восприятия ЭП основан на притяжении или
роускорения, f - частота ЭП.
отталкивании под действием кулоновских сил
При действии ЭП частотой 500 Гц и напря-
первичных преобразователей механорецепторных
женностью 15 кВ/м амплитуда вибросмещения
органов, реагирующих на смещение. Под действи-
стенок садка составляла 0.18 ± 0.02 нм. Такие виб-
ем низкочастотных ЭП колеблются антенны и во-
рации опорного субстрата не возбуждают пчел. У
лоски быстроадаптирующихся трихоидных сен-
них порог чувствительности к колебаниям опор-
силл, выполняющих у пчел функцию фонорецепто-
ного субстрата на частоте 500 Гц составляет около
ров [37].
5 нм [34]. Но это стимулирует у пчел не активиза-
Минимальная напряженность однородного
цию, а торможение локомоций [21].
ЭП, отклоняющего волосок быстроадаптирую-
У ос чувствительность субгенуальных органов
щейся трихоидной сенсиллы пчелы на 1°, состав-
к вибрациям опорного субстрата примерно в три
ляет 20-25 кВ/м. Рост неоднородности ЭП, свя-
раза ниже, чем у пчел [35]. Поэтому осы, как и
занный, например, со сближением головами двух
пчелы, не могут воспринимать колебания опор-
или более пчел, увеличивает угол отклонения во-
БИОФИЗИКА том 65
№ 3
2020
572
ЕСЬКОВ
лосков. При сближении головами двух пчел их во-
С раздражением пчел и муравьев наведенными
лоски отклоняются на 1° при напряженности 7-
токами связана модификация их поведения перед
10 кВ/м [38, 39]. Очевидно, что поэтому одна пче-
грозой. На расстоянии 50 км от грозового фронта
ла, находясь на электрически изолированной по-
напряженность пульсаций ЭП может достигать
верхности, не реагирует, а две реагируют на ЭП
10 кВ/м [13, 40], что превышает порог чувстви-
напряженностью 50 кВ/м.
тельности к этому стимулу. С приближением гро-
Колебания антенн насекомых воспринимает-
зового фронта у пчел-фуражиров, находящихся
ся джонстоновыми органами. У пчел при одина-
на цветках, возрастает частота прерывания на-
ковой напряженности ЭП максимальное откло-
полнений зобика [41]. Это стимулирует у пчел по-
нение антенн происходит под действием ЭП
вышение агрессивности и возвращение в ульи. У
частой 50 Гц. На этой частоте при напряженности
муравьев раздражение наведенными токами ас-
5, 7 и 10 кВ/м антенны отклоняются соответ-
социируется с опасностью, стимулирующей за-
ственно на 2, 3 и 6°. Накопление статического за-
крытие входных отверстий в муравейнике.
ряда на теле пчел, связанное с их активизацией,
Таким образом, пчелы, осы и муравьи реагиру-
влияет на увеличение угла отклонения антенн
ют на искусственные низкочастотные ЭП высо-
[21]. Поэтому ЭП высокой напряженности может
кой напряженности, хотя не обладают для этого
восприниматься джонстоновыми органами ан-
специализированными рецепторами. Но, ис-
тенн.
пользуя преимущественно неспециализирован-
Неспецифический механизм восприятия ЭП
ные средства восприятия ЭП, пчелы приобрели
основан на раздражении насекомых наведенными
инстинкты хоминга, а муравьи - защиты мура-
токами, протекающими в местах их контактирова-
вейника перед ненастной погодой. Предвестни-
ния между собой и/или токопроводящими по-
ками непогоды служат у пчел и муравьев атмо-
верхностями. Частотная характеристика чувстви-
сферики, интенсивность которых возрастает при
тельности к ЭП определяется в основном величи-
приближении грозового фронта, усиливающего
ной наводимого им тока и эффективностью его
вариации напряженность ЭП. Пчелы, обладая
контактного действия. Поскольку ток наводится
совершенными средствами связи, изменяют ука-
в покровах тела, характеризующихся высоким со-
зание координат источника корма соответствен-
противлением, то величина наводимого тока не-
но суточным вариациям магнитного поля Земли.
линейно связана с частотой ЭП. Вероятнее всего,
наводимый ток достигает максимума на частоте
КОНФЛИКТ ИНТЕРЕСОВ
около 500 Гц, что согласуется с областью наиболь-
шей чувствительности к ЭП, хотя чувствительность
Авторы заявляют об отсутствии конфликта
к току с понижением частоты возрастает [22]. Но
интересов.
под действием постоянного ЭП ток не наводится.
Поэтому пчелы не реагируют на ЭП постоянного
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
тока [21].
Высокая вариабельность чувствительности к
Настоящая работа не содержит описания иссле-
ЭП связана с действием ряда случайных факто-
дований с использованием людей и животных в ка-
ров. К ним относится изменчивость величины
честве объектов.
контактного тока. Она во многом зависит от
электропроводности контактирующих поверхно-
СПИСОК ЛИТЕРАТУРЫ
стей, которая определяется их свойствами и со-
стоянием. В местах контакта могут находиться ве-
1. А. С. Пресман, Электромагнитные поля и живая
щества различной электропроводности: корм, ча-
природа (Наука, М., 1968).
стицы пыли, воска, прополиса и т. п. Кроме того,
2. Ю. В. Сербант и М. П. Троянский, Радиоволны и
с увеличением общей массы агрегирующихся
живой организм (Знание, М., 1969).
пчел возрастает величина наводимого тока. В зо-
3. А. П. Дубров, Геомагнитное поле и жизнь (Гидро-
нах скопления пчел создаются значительные ло-
метеоиздат, Л., 1974).
кальные градиенты напряженности, которые мож-
4. Ю. А. Холодов, Реакции нервной системы на элек-
но рассматривать как вторичные поля, выступаю-
тромагнитные поля (Наука, М., 1975).
щие в роли дополнительных стимуляторов
5. О. В. Белянский, М. Б Голант и Н. Д. Девятков,
возбуждения. По этой причине взаимные ужале-
Миллиметровые волны в биологии (Знание, М.,
ния происходят в местах больших скоплений
1988).
пчел. Повышению их агрессивности способству-
ет наличие в ЭП токопроводящих тел. Рефлекс
6. Е. К. Еськов, Успехи соврем. биологии 123 (3), 195
ужаления стимулирует изолированный провод-
(2003).
ник, на который подается напряжение, составля-
7. Е. К. Еськов и А. И. Торопцев, Журн. эволюц. био-
ющее всего несколько десятков вольт.
химии и физиологии 15 (5), 500 (1979).
БИОФИЗИКА том 65
№ 3
2020
ВОСПРИЯТИЕ И СПЕЦИФИЧНОСТЬ РЕАГИРОВАНИЯ
573
8. Е. К. Еськов, Изв. АН СССР. Сер. биол., № 1, 135
25. U. Warnke and R. Paul, Umschau, № 3, 415 (1975).
(1982).
26. W. E. Caldwell and F. Russo, J. Genet. Physiol. 113 (2),
9. Е. К. Еськов, Жилища насекомых (Знание, М.,
233 (1968).
1983).
27. M. M. Walker and M. E. Bitterman, J. Exp. Biol. 145,
10. Е. К. Еськов, Экология медоносной пчелы (Русское
489 (1989).
слово, Рязань, 1995).
28. А. Л. Чижевский, в сб. Труды практической лабора-
11. Г. М. Длусский, Муравьи рода Formica (Наука, М.,
тории по зоопсихологии (М. 1928), вып. 1, сс. 42-47.
1967).
29. G. Altman and U. Warnke, Apidologie 2 (4), 309
(1971).
12. В. К. Дмитриенко и Е. С. Петренко, Муравьи та-
ежных биоценозов Сибири (Наука, М., 1976).
30. Р. Фейман, Р. Лейтон и М. Сендлс, Феймановские
лекции по физике (Мир, М., 1966), т. 6, сс. 201-207.
13. Дж. А. Чалмерс, Атмосферное электричество (Гид-
рометеоиздат, Л., 1974).
31. Н. Г. Бочкарев, Магнитное поле в космосе (Наука,
М., 1985).
14. Е. К. Еськов, Биофизика 63 (3), 561 (2018).
32. M. Lindauer and H. Martin, Z. Vergl. Physiol. 60, 219
15. Е. К. Еськов и А. М. Сапожников, Изв. АН СССР.
(1968).
Сер. биол., № 3, 395 (1979).
33. Е. К. Еськов и Г. А. Миронов, Экология, № 6, 81
16. Е. К. Еськов, Зоол. журн. 64 (4), 606 (1985).
(1990).
17. Е. К. Еськов, Журн. эволюц. биохимии и физиоло-
34. 34. H. F. Little, Ann. Entomol. Soc. Amer. 55, 82
гии 17 (2), 178 (1981).
(1962).
18. Е. К. Еськов, Экология, № 6, 76 (1982).
35. H. Autrum and W. Schneider, Z. Vergl. Physiol. 31 (1),
19. Е. К. Еськов, Акустическая сигнализация обще-
77 (1948).
ственных насекомых (Наука, М., 1979).
36. V. P. Bindokas and B. Grenderg, Dioelectromagnetics
20. Е. К. Еськов, Экология, № 1, 60 (1997).
5, 305 (1984).
21. Е. К. Еськов, Этология медоносной пчелы (Колос,
37. Е. К. Еськов, Биофизика 20 (4), 646 (1975).
М., 1992).
38. Е. К. Еськов и Г. А. Миронов, Докл. АН СССР 309
22. Е. К. Еськов и А. М. Сапожников, Биофизика 24
(1), 233 (1989).
(4), 780 (1979).
39. Е. К. Еськов и Г.А. Миронов, Биофизика 35 (4),
23. Е. К. Еськов и Н. И. Брагин, Журн. общ. биологии
675 (1990).
47 (6), 823 (1986).
40. M. Friedmann, Electrotechnik und Masghinenbau 91
24. Е. К. Еськов и Н. И. Брагин, Пчеловодство, № 2, 9
(10), 510 (1975).
(1986).
41. L Schua, Z. Vergl. Physiol. 34, 258 (1952).
Perception and Specificity of Behavioral Effects observed in Honey Bees, Paper Wasp,
and Red Wood Ants in Response to Low Frequency Electric Field
E.K. Eskov
Russian State Agricultural Correspondence University, ul. Yu. Fuchika 1, Balashicha, Moscow Region, 143900 Russia
Bees, wasps and ants have no specialised receptors for electroreception. An adequate response to naturally oc-
curring electric charge in bees and ants is related to the effects of atmospherics produced due to less distant
thunderstorms. Perception of low-frequency high-voltage electric field in insects involves activation of pri-
mary transduction elements of mechanoreceptors reflecting their movements. The nonspecific mechanism
of electroreception is triggered when insects while in contact with each other and / or electrically conductive
surfaces are stimulated with electric fields. The frequency response of the sensitivity to electric fields is deter-
mined mainly by the magnitude of the electric current used and its intensity between insect bodies and/or
conductive material. The magnitude of the current used for the insect body is nonlinearly related to the elec-
tric field frequency. The region with the highest sensitivity to electric field is at about 500 Hz, which is con-
sistent with the maximum value of the current used. Herewith, the threshold of sensitivity to electric field in
wasps and bees are about 0.04 kV/m and 0,45 kV/m, respectively. Ants exposed to electric fields ranging be-
tween 7-10 kV/m show impairment in locomotion. The magnetic fields and ionization associated with the
generation of electric fields up to 15-20 kV/m do not stimulate behavioral changes in bees.
Keywords: bees, wasps, ants, electric field, sensitivity, behavior anomalies, adequate response
БИОФИЗИКА том 65
№ 3
2020