БИОФИЗИКА, 2020, том 65, № 3, с. 614-624
ДИСКУССИИ
УДК 577.3
ПЕРКОЛЯЦИОННАЯ РЕШЕТКА ЕСТЕСТВЕННОГО ОТБОРА
КАК КОММУТАТОР ДЕТЕРМИНИРОВАННЫХ И СЛУЧАЙНЫХ
ПРОЦЕССОВ В ПОТОКЕ МУТАЦИЙ
© 2020 г. А.Я. Гараева, А.Э. Сидорова, Н.Т. Левашова, В.А. Твердислов
Физический факультет Московского государственного университета имени М.В. Ломоносова,
119991, Москва, Ленинские горы, 1/2
E-mail: sky314bone@mail.ru
Поступила в редакцию 28.12.2019 г.
После доработки 28.12.2019 г.
Принята к публикации 10.02.2020 г.
Рассмотрены основные модели мутационных процессов в ходе отбора и дрейфа. Представлена
принципиально новая модель протекания мутаций через перколяционную решетку отбора на попу-
ляционном уровне. Естественный отбор оптимальных вариантов в эволюционном потоке мутаций
рассмотрен в качестве перколяционного фильтра - физического инструмента, составляющего ме-
ханизм отбора. Модель основана на представлениях о самоорганизации в иерархических структурах
видообразования, где триггерные свойства ячейки (узла) как вентиля определяют детерминирован-
ную компоненту закрепления новых мутаций, а дрейф определяет элемент случайности в процесс
закрепления новых мутаций в потоках мутаций. Получены режимы закрепления мутаций в зависи-
мости от соотношения размера кластера носителей мутаций к общему размеру популяции, коэффи-
циентов отбора и дрейфа в череде поколений на популяционном уровне. Определен нижний порог
перколяционной решетки отбора как доля новых мутаций, зафиксированных под воздействием де-
терминированных процессов отбора для минимально воспроизводящей численности кластера.
Ключевые слова: естественный отбор, самоорганизация, флуктуации, бифуркации, перколяции,
запрещающие и разрешающие мутации
DOI: 10.31857/S0006302920030229
Моделирование мутационных процессов в хо-
фиксации благоприятных мутаций. Новые мута-
де отбора и дрейфа, как правило, базируется на
ции чаще возникают на клональном фоне, и кло-
анализе однотипных мутаций [1, 2] и в основном
нальное доминирование дает мутациям преиму-
положительных [1, 3-6]. Снижению вероятности
щество, особенно если оно сохраняется в течение
фиксации полезных мутаций способствуют взаи-
длительного времени [21, 22]. Но, как показано
модействия между мутациями. Так, взаимодей-
Н.В. Тимофеевым-Ресовским
[23], возможна
ствие отрицательных мутаций [7] может нивели-
фиксация единичной мутации с большим поло-
ровать эффект положительных мутаций [1, 4, 8], а
жительным эффектом, если она затрагивает клю-
взаимодействия между полезными мутациями
чевую функцию и поддержана ранее закрепив-
могут приводить к замедлению процесса адапта-
шимися мутациями - принцип усиления. Адап-
ции и снижению усредненного эффекта полез-
тации могут способствовать как единичные
ных мутаций [9]. В целом клональная интерфе-
положительные мутации с большим эффектом,
ренция между мутациями возможна между мута-
так и множественные положительные мутации с
циями с различной направленностью [10], и ее
небольшим эффектом, которые встречаются ча-
вероятность увеличивается при увеличении чис-
ще, чем единичные мутации с большим эффек-
ленности популяции и частоты мутаций [11]. Рас-
том и способны подавлять фиксацию мутаций с
пределение полезных мутаций в ряде работ свя-
большим эффектом [11, 24, 25]. В первом случае
зывается с эффектами плейотропности [12, 13],
речь идет об узкоспециализированных положи-
Хилла-Робертсона [14], «храповика Мюллера»
тельных мутация, которые способны вытеснять
[15], обратных мутаций [16], эпистаза (так, в по-
менее полезные мутации [9, 26]. Однако необхо-
пуляциях с половым отбором эпистаз может спо-
димо учитывать, что узкоспециализированные
собствовать или не способствовать рекомбина-
положительные мутации с большим эффектом
ции [17-20]), которые при определенных услови-
являются «удачными» на небольшом временном
ях способствуют ускорению или замедлению
интервале, в отличие от широких адаптаций (ре-
614
ПЕРКОЛЯЦИОННАЯ РЕШЕТКА ЕСТЕСТВЕННОГО ОТБОРА
615
зультата взаимодействия спектра мутаций) - бо-
га перколяции, определяемого долей занятых уз-
лее важных для процесса видообразования.
лов, при которой возникает перколяционный
кластер, - связанные элементы в модели проте-
Многие модели мутаций с одинаковым эф-
кания [45]. Перколяции могут наблюдаться в си-
фектом рассматривают приспособленность как
стемах, состоящих из большого числа подобных
бегущую волну с постоянной скоростью (в соот-
элементов, если такие связанные элементы рас-
ветствии с уравнением Фишера-Колмогорова-
пределенных систем находятся подобно триггеру
Петровского-Пискунова) [4, 16, 27-29], где в об-
в одном из двух состояний - устойчивости и не-
ласти действия отбора применяются детермини-
устойчивости. В отсутствие детерминированных
стические законы, а в области генетического
механизмов сеть неустойчивых перколяций сто-
дрейфа - законы случайных чисел [28, 30]. Мо-
хастична и не может обеспечить устойчивое раз-
дель бегущей волны также адекватно описывает
витие системы, поэтому неустойчивые траекто-
обратные и компенсирующие мутации [27, 31] (в
рии мутационного процесса отфильтровываются
случае обратных или компенсирующих мутаций
отбором. Устойчивые (коррелированные) перко-
волна может двигаться либо вправо - храповик,
ляции составляют траекторию эволюции вида
либо влево - реверсия [16]), промежуточную об-
или популяции. Выход популяции/вида на устой-
ласть в профиле волны, определяемую парамет-
чивую траекторию эволюции - это динамическое
рами отбора и дрейфа [16, 28], «стохастический
единство генетического и популяционного уров-
порог» [11, 28, 32], стохастический скачок в «про-
ней, связанных обратными связями [44]. По су-
странстве пригодности» [33] и динамику смеще-
ществу, естественный отбор определяет свойства
ния приспособленности [28, 34], где приспособ-
протекания перколяционной решетки, т.е. опре-
ленность может контролироваться мутациями с
деляет закономерность прохождения новых мута-
достаточно слабым эффектом [33].
ций в зависимости от спектра мутаций, ранее за-
В ряде волновых моделей [35-37] рассматри-
фиксированных в популяции. Давление для про-
вается влияние на мутационный процесс «мута-
тока создает пул мутаций, аналогично давлению
тора» и «антимутатора», наличие которых при
газа или жидкости через перколяционный
определенных условиях может способствовать
фильтр, и в результате отбора-фильтра происхо-
вероятности появления новой мутации [38, 39].
дит выборка и фиксация мутаций. Иными слова-
Вероятность фиксации аллеля определяется раз-
ми, при наличии устойчивых (коррелированных)
мером популяции, параметрами среды, клональ-
мутаций формируется устойчивая траектория
ной интерференцией, частотой «полезных» и
эволюции.
«вредных» мутаций, а также «мутатора» и «анти-
Аналогично перколяциям (случайным и cкор-
мутатора». В целом, естественный отбор способ-
релированным [45, 47-49]), мутационный про-
ствует стабильной частоте мутаций в популяции,
цесс носит детерминированный (определяется
поэтому аллели «мутатора» и «антимутатора», ча-
генотипом и фенотипом в ходе отбора) и случай-
стота которых больше стабильной частоты, как
ный характер (генетический дрейф), а выбор тра-
правило, не поддерживаются отбором [35]. Одна-
ектории развития на популяционном уровне за-
ко понимание механизмов действия «мутатора» и
висит от спектра ранее закрепившихся «разреша-
«антимутатора» в настоящее время остается не-
ющих» и «запрещающих» мутаций [50].
полным.
В значительной степени вероятность фикса-
ции мутаций зависит от спектра ранее закрепив-
МОДЕЛЬ ПЕРКОЛЯЦИОННОЙ РЕШЕТКИ
шихся мутаций. Так, полезные мутации способ-
ОТБОРА В ПОТОКЕ МУТАЦИЙ
ны создавать блоки с вредными и нейтральными
ПОД УПРАВЛЕНИЕМ
мутациями в других локусах, что способствует ге-
ДЕТЕРМИНИРОВАННЫХ И СЛУЧАЙНЫХ
нетической дисперсии и увеличению частоты ре-
ПРОЦЕССОВ
комбинации [40-43]. Поэтому эффект новых му-
В модели на популяционном уровне мутации
таций зависит от спектра ранее закрепившихся, а
рассматриваются вне зависимости от их направ-
последовательность мутаций и их закрепление
ленности, так как вектор их направленности
определяют «вектор отбора» после прохождения
определяется их адаптивной пользой на кратком
точки бифуркации - «критической» точки видо-
временном интервале и в дальнейшем, в зависи-
образования. С физической точки зрения мы
мости от условий среды, может изменить знак.
имеем дело с кинетической необратимостью.
Узлы в перколяционной решетке (особи в попу-
Естественный отбор можно сравнить с явле-
ляции) работают как триггерная вентильная си-
нием перколяции
[44], где перколяционный
стема в потоках мутаций, где особи - носители и
фильтр - составляющий физический инструмент
преобразователи мутаций. Триггерные свойства
отбора оптимальных вариантов в эволюционном
ячейки (узла) как вентиля определяют детерми-
потоке мутаций. Параметры структуры, через ко-
нированную компоненту закрепления новых му-
торую протекает поток подвижной среды, и поро-
таций, а взаимодействие мутаций в череде поко-
БИОФИЗИКА том 65
№ 3
2020
616
ГАРАЕВА и др.
лений и случайные процессы дрейфа определяют
цесс возникновения сложных структур c новыми
коллективное поведение узлов и связей (мута-
параметрами из относительно простых элементов
ций) в пространстве перколяционной решетки.
[51].
В модели мутации (флуктуации) являются ис-
В модели для n поколений приняты следую-
точником, обеспечивающим качественный пере-
щие исходные характеристики.
ход популяции на новую ступень иерархической
Популяция - неоднородная среда, обладаю-
структуры видообразования. Самоорганизация в
щая набором запрещающих, разрешающих и но-
иерархических структурах видообразования под-
вых мутаций.
держивается системой взаимодействий на гори-
зонтальных уровнях. По мере накопления малых
Рассматриваются мутации: новые мутации
флуктуаций (мутаций) и формирования связей
(MC), закрепившиеся ранее мутации - запреща-
между мутациями, малые флуктуации преобразу-
ющие (MR) и разрешающие (MB) вне зависимо-
ются в гигантские, и система переходит на новый
сти от их направленности.
уровень - обретает новую структуру с новыми
функциями. Поэтому формирование нового вида
В отличие от ранее рассмотренной модели [52]
можно рассматривать как трехуровневый про-
в данной случае популяция состоит из N особей,
цесс. На первом уровне постоянного белого шума
Ne - размер кластера носителей мутаций (запре-
за счет наличия многочисленных случайных про-
щающих, разрешающих и новых) в долях от N,
цессов процесс определяется точечными взаимо-
N - Ne - количество особей-немутантов. В сред-
действиями особей - носителей мутаций. На
нем минимальная величина размера кластера Ne
этом уровне мутации для единичных особей мо-
составляет 30% от общей численности популяции
гут являться обратимыми, но при наличии доста-
[53], а максимальная величина варьирует в диапа-
точного количества носителей новой мутации
зоне 60-85% от общего числа особей в популяции
происходит формирование кластеров носителей
[54]. Принимаем 0.3 <Nе < 0.80.
мутаций. На втором уровне имеет место локаль-
Минимальное количество поколений п, необ-
ное взаимодействие кластеров, и начинаются
ходимое для закрепления мутации, принимаем
специализация и дифференциация, которые по-
равным 10. Согласно работам [55, 56], вероят-
лучат завершение при образовании вида. На этом
ность фиксации мутаций в поколениях п сначала
уровне взаимодействия кластеров также имеют
возрастает до п = 10, а затем мутации, лежащие в
место малые флуктуации, но конечный результат
полосе ±10%, ведут себя почти нейтрально, по-
в значительной степени определяется детерми-
этому можно считать, что минимальное количе-
нированными процессами. Переход со второго на
ство поколений, необходимое для закрепления
третий уровень является точкой бифуркации:
мутации на втором популяционном уровне, рав-
возможно формирование нового вида или элими-
но 10. В случае фиксации мутации в течение
нация носителей мутации. На третьем уровне вза-
10 поколений она считается закрепившейся.
имодействия между популяциями новая гигант-
ская флуктуация связана с формированием новой
Скорость изменения общей численности по-
структуры - нового вида. Это элементарный про-
пуляции N равна
dN =−r
·(N −Ne)
+
r
·(N -Ne)
−
r
·Ne + r
·
Ne,
(1)
1
2
3
4
dn
где r1 - константа скорости смертности нему-
Ne зависит только от скорости смерти и рож-
дения мутантов:
тантов, r2 - константа скорости рождаемости
немутантов, r3 - константа скорости смерт-
dNe =−r
·Ne + r
·Ne
(2)
3
4
dn
ности мутантов, r4 - константа скорости
В каждом поколении изменение численности
рождаемости мутантов. Скорость изменения
популяции равно
ΔN = (Nn - Nn-1) = -r1(Nn-1 - Nen-1) + r2(Nn-1 - Nen-1) - r3Nen-1 + r4Nen-1.
Следовательно, численность популяции в поколении n равна
Nn = -r1(Nn-1 - Nen-1) + r2(Nn-1 - Nen-1) - r3Nen-1 + r4Nen-1 + Nn-1.
(3)
БИОФИЗИКА том 65
№ 3
2020
ПЕРКОЛЯЦИОННАЯ РЕШЕТКА ЕСТЕСТВЕННОГО ОТБОРА
617
r3 = 0.1
r3 = 0.2
r3 = 0.3
500
120
120
N
N
N
100
100
400
Ne
Ne
Ne
80
80
300
60
60
200
40
40
100
20
20
0
0
0
0
10
20
30
0
10
20
30
0
10
20
30
Поколение
Поколение
Поколение
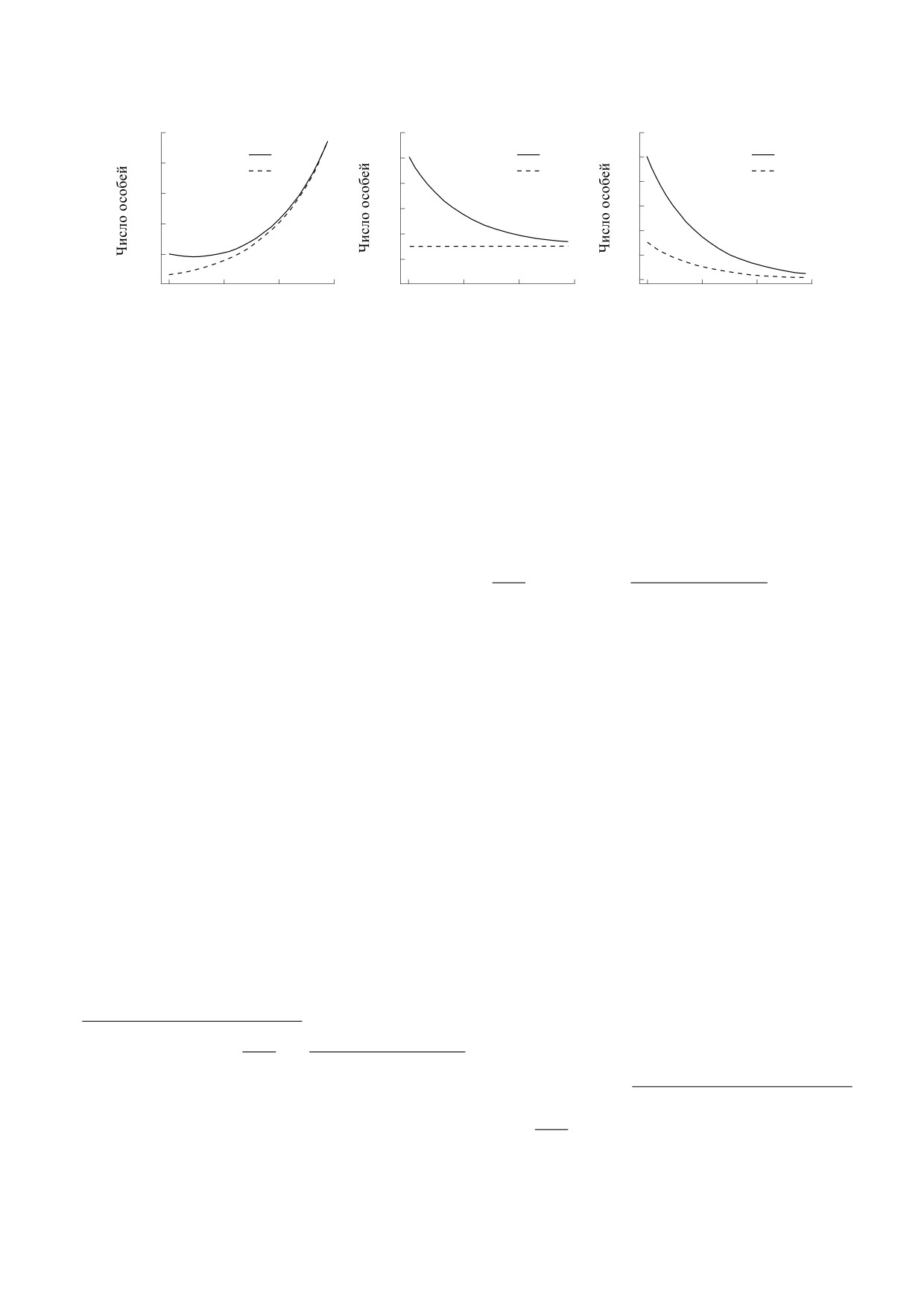
Рис. 1. График зависимости числа особей в популяции (N) и мутантов (Ne) от поколений при различных значениях
констант рождаемости и смертности мутантов и немутантов: (а) - r3 < r4, r1 > r2; (б) - r3 = r1, r1 > r2; (в) - r3 > r1, r1 > r2.
Для части популяции, имеющей мутацию,
Исходно новая мутация MC может быть пере-
имеем:
дана конечному числу особей в популяции, в за-
ΔNe = (Nen - Nen-1) = -r3Nen-1 + r4Nen-1,
висимости от соотношения Ne/N, наличия и со-
отношения разрешающих (MR) и запрещающих
Nen = -r3Nen-1 + r4Nen-1 + Nen-1.
(MB) мутаций, значений коэффициентов отбора
Рассмотрим зависимость числа особей в сово-
(s) и дрейфа (k). Скорость закрепления новой му-
купной популяции N и мутантов Ne от поколений
тации (в долях от общего количества мутаций) в
для следующих случаев:
поколениях определяется как
1. Константа скорости смертности мутантов
dM
(p
M -p
M
)Ne
меньше, чем константа скорости смертности не-
C
R R
B B
=-lnM
·
p
+
kW,
(4)
C C
мутантов (r3 < r4), константа скорости смерти не-
dt
N
мутантов выше, чем константа их скорости рож-
где MC - доля новых мутаций от общего количе-
дения (r1 > r2). В этом случае рост численности
ства мутаций (нормированных на 1); MR - доля
популяции N происходит за счет роста численно-
разрешающих мутаций (нормированных на 1);
сти мутантов Ne. В итоге к тридцатому поколе-
MB - доля запрещающих мутаций (нормирован-
нию немутантов в популяции содержится менее
ных на 1), закрепившихся; Ne - эффективный
0.7% (рис. 1а).
размер популяции (доля особей от общей числен-
2. Константа скорости смертности мутантов
ности популяции, способных к размножению и
равна константе скорости смертности немутан-
являющихся носителями мутаций - кластер);
тов (r3 = r1) и константа скорости смертности не-
pC - вероятность фиксации новых мутаций; pR -
мутантов выше, чем константа скорости их рож-
вероятность наличия поддерживающих мутаций;
даемости (r1 > r2). В этом случае численность по-
pB - вероятность наличия запрещающих мута-
пуляции падает за счет уменьшения численности
ций; kW - слагаемое, описывающее дрейф, где
немутантов, при этом численность мутантов
W - случайная величина, равномерно распреде-
остается неизменной. Популяция выходит на по-
ленная на отрезке [-1, 1], k - коэффициент дрей-
стоянный уровень N = Ne0 = 30 (рис. 1б).
фа. Начальные условия МR = МB = 0.5.
3. Константа скорости смертности мутантов
Для закрепления новой мутации необходимо
больше константы скорости смерти немутантов
выполнение условия pRMR - pBМB > 0, т.е.
(r3 > r1) и константа скорости смертности нему-
pR > 0.5. Принимаем pR = 0.51.
тантов выше, чем константа скорости их рождае-
мости (r1 > r2). В этом случае происходит элими-
В окончательном виде уравнение (3) принима-
нация всей популяции (рис. 1в).
ет следующий вид:
dM
С
Ne/Ns
= -
0.51Ne/Ns
· ln (M
С
)
+
kW
(5)
dt
{1
-
exp(-2Ne
/
Ns)}
При малых стохастических возмущениях
dM
С
Ne
=-0.51p
C
ln(M
С
)
+
kW
(6)
(kW << 0.51pCNe) уравнение (5) принимает вид
dt
N
БИОФИЗИКА том 65
№ 3
2020
618
ГАРАЕВА и др.
Ne/N = 0.3
Ne/N = 0.4
W
W
1.0
1.0
0.5
0.5
0.0
0.0
-0.5
-0.5
-1.0
-1.0
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
Поколение
Поколение
Ne/N = 0.5
Ne/N = 0.6
W
W
1.0
1.0
0.5
0.5
0.0
0.0
-0.5
-0.5
-1.0
-1.0
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
Поколение
Поколение
Ne/N = 0.7
Ne/N = 0.8
W
W
1.0
1.0
0.5
0.5
0.0
0.0
-0.5
-0.5
-1.0
-1.0
0
1
2
3
4
5
6
7
8
9
10
0
1
2
3
4
5
6
7
8
9
10
Поколение
Поколение
Выборка №1
Выборка №2
Выборка №3
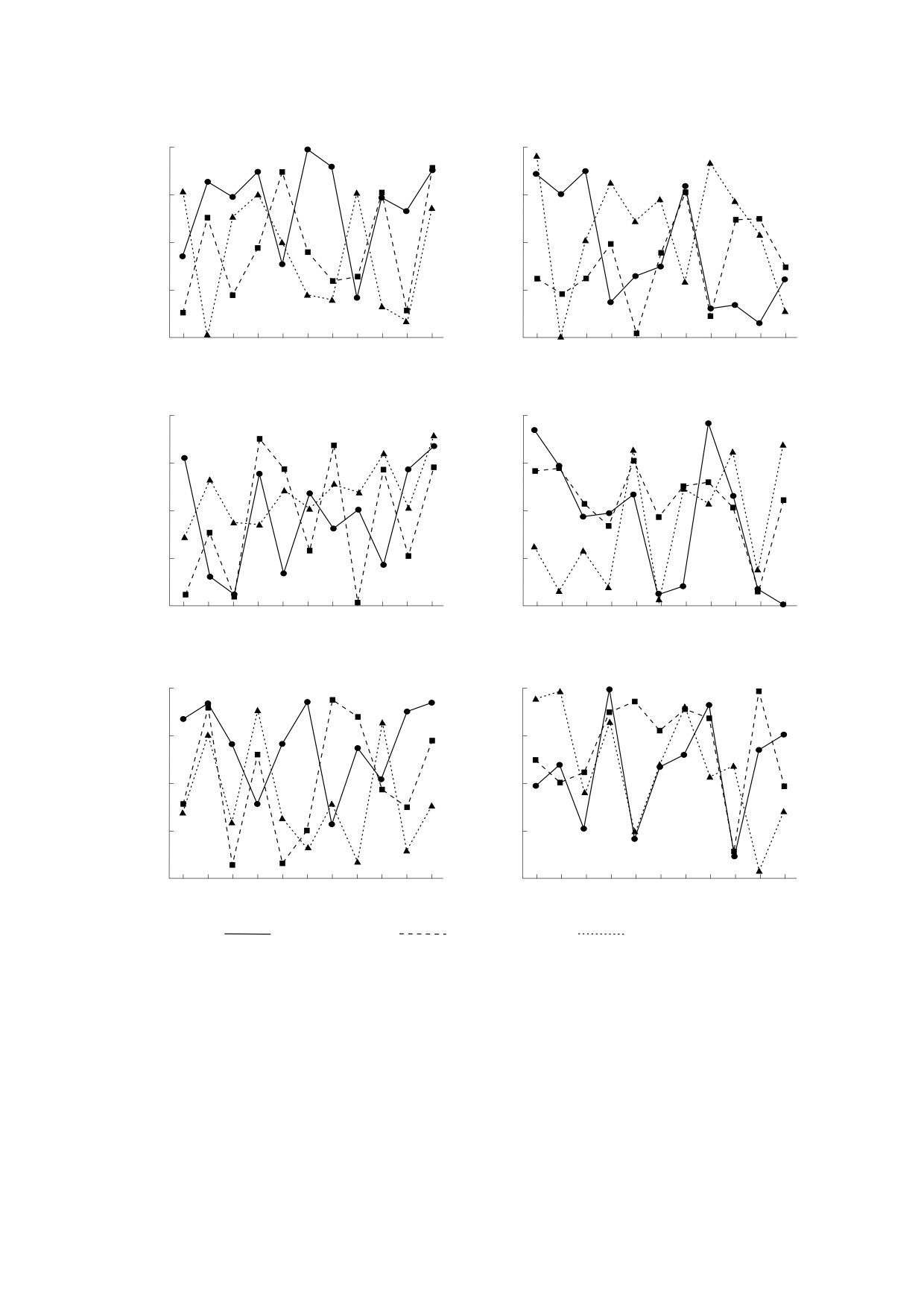
Рис. 2. Выборки из 11 случайных чисел для поколений (n) от 0 до 10 (ось х) при различных значениях Ne/N для трех
выборок случайных чисел.
Согласно нашим расчетам, величина 0.51pCNe
Для численного решения задачи скорости закреп-
изменяется в пределах от 0.08 ≈ 0.1 до 0.23, поэто-
ления новых мутаций как доли общего количества
му пороговым значением мы будем считать вели-
мутаций по поколениям разделим интервал t ∈ [0, t]
чину k = 0.1: при k < 0.1 в ходе закрепления мута-
(где t = 10 поколений) на 100 равных частей. Для ре-
ций доминирует детерминированная составляю-
шения уравнения применим одностадийную схему
щая отбора, а при k ≥ 0.1 - дрейф.
Розенброка с комплексным коэффициентом [57].
БИОФИЗИКА том 65
№ 3
2020
ПЕРКОЛЯЦИОННАЯ РЕШЕТКА ЕСТЕСТВЕННОГО ОТБОРА
619
(а)
(б)
0.55
2.0
0.54
1.6
Т
0.53
1.2
0.52
0.8
<M >С
0.51
0.4
0.50
0.0
0.3
0.4
0.5
0.6
0.7
0.8
0.0
0.2
0.4
0.6
0.8
1.0
Ne/N
Ne/N
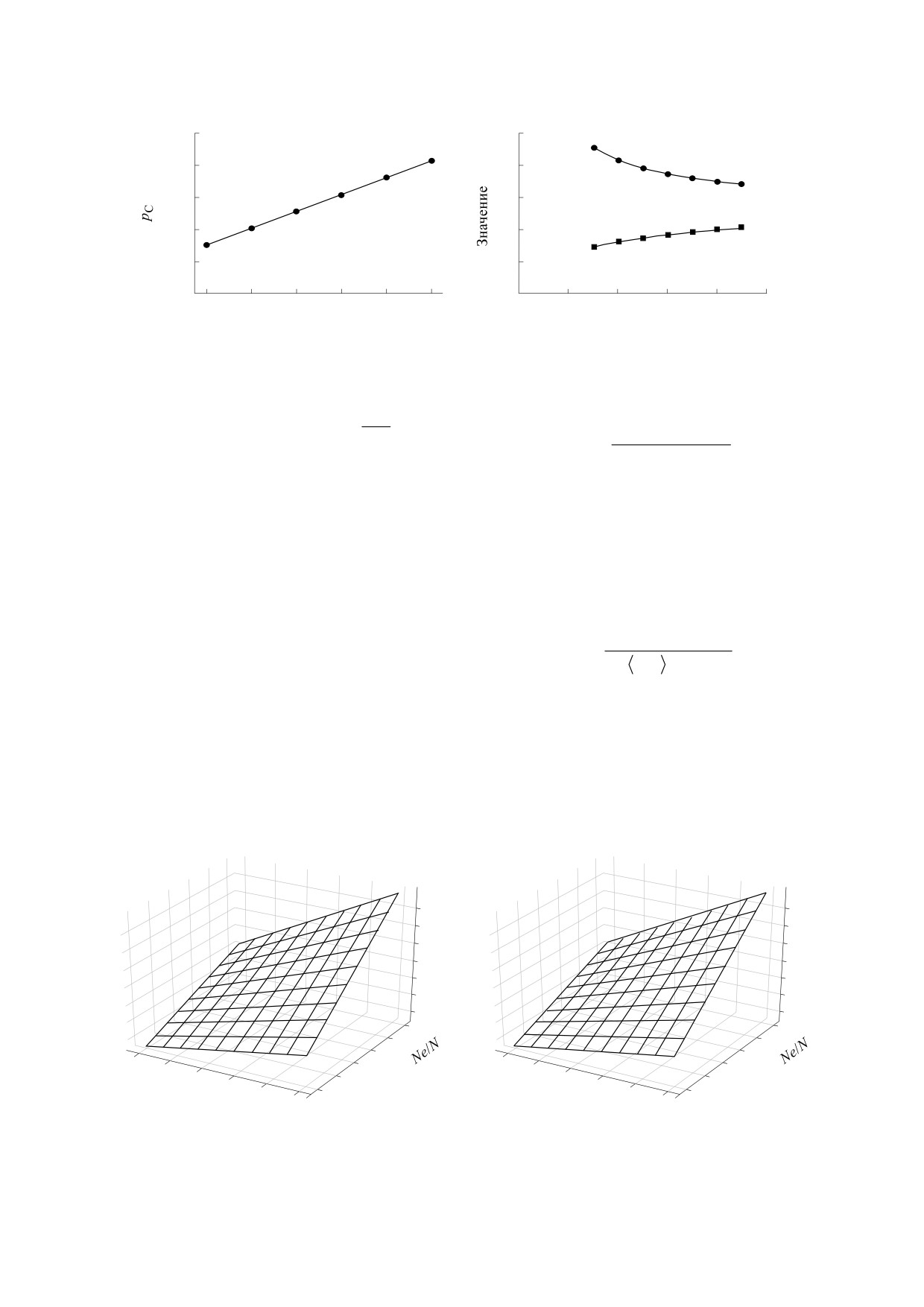
Рис. 3. Графики вероятности закрепления новых мутаций (pС), минимального времени между фиксациями мутаций
(Т) и усредненной доли зафиксировавшихся мутаций (<MC>) при отсутствии дрейфа (k = 0): (а) - зависимость (pC) от
Ne/N; (б) - зависимость Т и <MC> от Ne/N при фиксированных значениях коэффициента отбора (s = 0.1).
M
[
m+1
]
=M
[
m
]
+ τReω,
m=
0,99.
(7)
Ne
/
Ns
C
C
p
≈
(8)
C
{1-
exp(−2N
s)}
e
Величина W изменяется каждые десять шагов
Минимальное время между последовательны-
по времени (раз в поколение). На рис. 2 показаны
ми фиксациями мутации для детерминирован-
выборки случайный чисел, используемые при
ных процессов в зависимости от относительного
расчетах согласно уравнению (5).
размера клона Ne/N, среднего значения доли за-
фиксированных мутаций <MC> для заданного
Ne/N, коэффициента отбора s и отсутствии дрей-
РЕЗУЛЬТАТЫ МОДЕЛИРОВАНИЯ
фа (на основании данных работы [58]) определя-
ется как
При отсутствии дрейфа процесс закрепления
мутаций определяется только закономерностями
exp
(2
sNe
/N
)
-
отбора, т.е. наличием в популяции поддерживаю-
T
≈
1.
(9)
2
M
sNe/N
щих и запрещающих мутаций и их взаимодей-
C
ствием с новыми мутациями. В этом случае веро-
Зависимость Т и <MC> от Ne/N при фиксиро-
ятность закрепления новых мутаций при отсут-
ванных значениях s = 0.1 и k = 0 демонстрирует,
ствии дрейфа рассчитывается по формуле (8) (на
что при увеличении Ne/N растет средняя доля но-
основании данных работы [58]) и растет незначи-
вых мутаций <MC> и уменьшается время их фик-
тельно с ростом относительного размера клона
сации (рис. 3б). При фиксированном значении
при исключительно детерминированных процес-
<MC> представлен трехмерный график зависимо-
сах (рис. 3а):
сти времени фиксации Т от Ne/N и s (рис. 4). На
(а)
(б)
T
T
11.6
1.16
11.4
1.14
11.2
1.12
11.0
1.10
10.8
1.08
10.6
1.06
10.4
1.04
0.8
0.8
0.7
0.7
0.6
0.6
0.100.12
0.5
0.100.12
0.5
s
0.4
s
0.14
0.4
0.14
0.16
0.16
0.3
0.3
0.18
0.20
0.18
0.20
Рис. 4. Графики зависимости Т от Ne/N и s при фиксированном значении <MC>: (а) <MC> = 0.1; (б) - <MC> = 1.0.
БИОФИЗИКА том 65
№ 3
2020
620
ГАРАЕВА и др.
Выборка № 1
s = 0.1
s = 0.15
s = 0.2
МС
10
10
10
1.0
9
9
9
8
8
8
0.8
7
7
7
6
6
6
0.6
5
5
5
4
4
4
0.4
3
3
3
2
2
2
0.2
1
1
1
0
0
0
0.0
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
Ne/N
Ne/N
Ne/N
Выборка № 2
s = 0.1
s = 0.15
s = 0.2
МС
10
10
10
1.0
9
9
9
8
8
8
0.8
7
7
7
6
6
6
0.6
5
5
5
4
4
4
0.4
3
3
3
2
2
2
0.2
1
1
1
0
0
0
0.0
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
Ne/N
Ne/N
Ne/N
Выборка № 3
s = 0.1
s = 0.15
s = 0.2
МС
10
10
10
1.0
9
9
9
8
8
8
0.8
7
7
7
6
6
6
0.6
5
5
5
4
4
4
0.4
3
3
3
2
2
2
0.2
1
1
1
0
0
0
0.0
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
Ne/N
Ne/N
Ne/N
Рис. 5. Графики скорости закрепления новой мутации (в долях от общего количества мутаций) в поколениях в
зависимости от Ne/N при разных значениях коэффициента отбора и допороговых значениях коэффициента дрейфа
(k = 0.01): s = 0.1, s = 0.15 и s = 0.2. На легенде оттенками серого цвета обозначено возрастание доли новых мутаций
(от черного до белого). Графики соответствуют выборкам случайных чисел № 1, 2 и 3.
графике видно, что при фиксированном <MC>
При допороговых значениях коэффициента
время фиксации практически не зависит от s и
дрейфа (k = 0.01) по мере увеличения коэффици-
ента отбора (s) и соотношения Ne/N доля новых
Ne/N. При <MC> = 0.1 Т изменяется примерно на
мутаций (MС) аналогично возрастает по поколе-
одно поколение, при <MCMc> = 1 - на 0.1 поко-
ниям при всех выборках случайных чисел, что
ления. При этом время фиксации для <MC> = 0.1
объясняется слабым воздействием дрейфа
и <MC> = 1 различается на порядок (время между
(рис. 5).
последовательными фиксациями, когда все мута-
При достижении порогового значения коэф-
ции из наличествующих фиксируются, на поря-
фициента дрейфа (k = 0.1) для всех исследуемых
док меньше времени между последовательными
значений коэффициента отбора отмечаются при-
фиксациями, когда только 10% от мутаций фик-
знаки зарождения хаотических процессов
сируются).
(рис. 6).
БИОФИЗИКА том 65
№ 3
2020
ПЕРКОЛЯЦИОННАЯ РЕШЕТКА ЕСТЕСТВЕННОГО ОТБОРА
621
Выборка № 1
s = 0.1
s = 0.15
s = 0.2
МС
10
10
10
1.0
9
9
9
8
8
8
0.8
7
7
7
6
6
6
0.6
5
5
5
4
4
4
0.4
3
3
3
2
2
2
0.2
1
1
1
0
0
0
0.0
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
Ne/N
Ne/N
Ne/N
Выборка № 2
s = 0.1
s = 0.15
s = 0.2
МС
10
10
10
1.0
9
9
9
8
8
8
0.8
7
7
7
6
6
6
0.6
5
5
5
4
4
4
0.4
3
3
3
2
2
2
0.2
1
1
1
0
0
0
0.0
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
Ne/N
Ne/N
Ne/N
Выборка № 3
s = 0.1
s = 0.15
s = 0.2
МС
10
10
10
1.0
9
9
9
8
8
8
0.8
7
7
7
6
6
6
0.6
5
5
5
4
4
4
0.4
3
3
3
2
2
2
0.2
1
1
1
0
0
0
0.0
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
Ne/N
Ne/N
Ne/N
Рис. 6. Графики скорости закрепления новой мутации (в долях от общего количества мутаций) в поколениях в
зависимости от Ne/N при разных значениях коэффициента отбора и пороговом значении коэффициента дрейфа
(k = 0.1): s = 0.1, s = 0.15 и s = 0.2. На легенде оттенками серого цвета обозначено возрастание доли новых мутаций (от
черного до белого). Графики соответствуют выборкам случайных чисел № 1, 2 и 3.
При дальнейшем усилении дрейфа, т.е. при
мированием нижнего порога перколяционной
значении коэффициента дрейфа больше порого-
решетки отбора - долей новых мутаций, которые
вого (k > 0.1), процессы фиксации мутаций стано-
фиксируются в популяции под воздействием де-
вятся хаотическими (рис. 7). В этом случае в зави-
терминированных процессов отбора для мини-
симости от выборки случайных чисел возможно
мально воспроизводящей численности кластера в
как максимальное количество закрепившихся
популяции. В модели значение доли новых мута-
мутаций (выборка №1), так и минимальное (вы-
ций (MC) для Ne/N = 0.3 при отсутствии дрейфа
борка №3).
(k = 0.1) является нижним порогом перколяций
Состояние перколяционной решетки отбора,
(протекания). В этом случае в зависимости от
при котором формируется минимально достаточ-
значения коэффициента отбора в первом поколе-
ное количество носителей новой мутации (мини-
нии значение нижнего порога перколяций меня-
мальный кластер), в нашей модели связано с фор-
ется незначительно, а при увеличении Ne/N отме-
БИОФИЗИКА том 65
№ 3
2020
622
ГАРАЕВА и др.
Выборка № 1
s = 0.1
s = 0.15
s = 0.2
МС
10
10
10
1.0
9
9
9
8
8
8
0.8
7
7
7
6
6
6
0.6
5
5
5
4
4
4
0.4
3
3
3
2
2
2
0.2
1
1
1
0
0
0
0.0
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
Ne/N
Ne/N
Ne/N
Выборка № 2
s = 0.1
s = 0.15
s = 0.2
МС
10
10
10
1.0
9
9
9
8
8
8
0.8
7
7
7
6
6
6
0.6
5
5
5
4
4
4
0.4
3
3
3
2
2
2
0.2
1
1
1
0
0
0
0.0
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
Ne/N
Ne/N
Ne/N
Выборка № 3
s = 0.1
s = 0.15
s = 0.2
МС
10
10
10
1.0
9
9
9
8
8
8
0.8
7
7
7
6
6
6
0.6
5
5
5
4
4
4
0.4
3
3
3
2
2
2
0.2
1
1
1
0
0
0
0.0
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
0.3
0.4
0.5
0.6
0.7
0.8
Ne/N
Ne/N
Ne/N
Рис. 7. Графики скорости закрепления новой мутации (в долях от общего количества мутаций) в поколениях в
зависимости от Ne/N при разных значениях коэффициента отбора (s = 0.1, s = 0.2) и значени коэффициента дрейфа
больше порогового (k = 0.2) в зависимости от выборки случайных чисел: s = 0.1, s = 0.15 и s = 0.2. На легенде оттенками
серого цвета обозначено возрастание доли новых мутаций (от черного до белого). Графики соответствуют выборкам
случайных чисел № 1, 2 и 3.
чается заметный рост доли новых мутаций уже в
ленности определяется их адаптивной пользой на
первом поколении.
кратком временном интервале и в дальнейшем, в
зависимости от условий среды, может изменить
знак. Узлы в перколяционной решетке (особи в
ВЫВОДЫ
популяции) работают как триггерная вентильная
система в потоках мутаций, где особи - носители
Представлена принципиально новая модель
и преобразователи мутаций. Триггерные свойства
протекания мутаций через перколяционную ре-
шетку отбора на популяционном уровне. В моде-
ячейки (узла) как вентиля определяют детерми-
ли мутации (положительные, отрицательные и
нированную компоненту закрепления новых му-
нейтральные) рассматриваются вне зависимости
таций (пропустить/не пропустить). Взаимодей-
от их направленности, так как вектор их направ-
ствие мутаций в череде поколений и случайные
БИОФИЗИКА том 65
№ 3
2020
ПЕРКОЛЯЦИОННАЯ РЕШЕТКА ЕСТЕСТВЕННОГО ОТБОРА
623
процессы дрейфа определяют коллективное по-
7. G. Martin, S. P. Otto, and T. Lenormand, Genetics 172
ведение узлов и связей (мутаций) в пространстве
(1), 593 (2006).
перколяционной решетки.
8. M. Kimura, Genetics Res. 9 (1), 23 (1967).
При допороговых значениях коэффициента
9. P. J. Gerrish and R. E. Lenski, Genetica 102, 127
дрейфа (k < 0.1) закрепление новых мутаций в ос-
(1998).
новном определяется закономерностями их взаи-
10. B. H. Good and O. Hallatschek, Curr. Opin. Microbi-
модействия с поддерживающими (MR) и запре-
ol. 45, 203 (2018).
щающими мутациями (MB). При достижении и
11. C. A. Fogle, J. L. Nagle, and M. M. Desai, Genetics
превышении порогового значения коэффициен-
180 (4), 2163 (2008).
та отбора (k = 0.1) отмечаются признаки зарожде-
12. G. Martin and T. Lenormand, Genetics 179 (2), 907
ния хаотических процессов. При значении коэф-
(2008).
фициента дрейфа больше порогового (k > 0.1)
13. G. Martin, Genetics 197 (1), 237 (2014).
возможно возникновение гигантских флуктуа-
14. G. A. T. McVean and B. Charlesworth, Genetics 155
ций и, как следствие, формирование качественно
(2), 929 (2000).
новой системы: формирование нового вида в
15. I. M. Rouzine, É. Brunet, and C. O. Wilke, Theor.
иерархии видообразования) или вымирание по-
Popul. Biol. 73 (1), 24 (2008).
пуляции под воздействием неблагоприятных му-
таций. Нижний порог протекания мутаций для
16. I. M. Rouzine, J. Wakeley, and J. M. Coffin, Proc.
перколяционной решетки отбора в модели - это
Natl. Acad. Sci. USA 100 (2), 587 (2003).
доля новых мутаций, которые закрепляются в по-
17. N. H. Barton and S. P. Otto, Genetics 169 (4), 2353
пуляции под воздействием детерминированных
(2005).
процессов отбора для минимальной численности
18. W. G. Hill and A. Robertson, Genetics Res. 8 (3), 269
кластера носителей мутаций.
(1966).
Нелинейные системы способны эволюциони-
19. S. P. Otto and N. H. Barton, Evolution 55 (10), 1921
ровать по разным траекториям, проходя через
(2001).
точки бифуркации, которые характеризуются не-
20. M. De Visser, et al., Science 283 (5400), 404 (1999).
устойчивым состоянием систем. Вблизи бифур-
21. N. L. Komarova, Proc. Natl. Acad. Sci. USA 111
кационной точки неравновесная система оказы-
(Suppl. 3), 10789 (2014).
вается особо чувствительной и к незначительным
22. L. Wagstaff, G. Kolahgar, and E. Piddini, Trends Cell
флуктуациям того или иного параметра, и флук-
Biol. 23 (4), 160 (2013).
туации становятся «гигантскими». Так в процессе
23. Н. В. Тимофеев-Ресовский, Избранные труды
биологической микроэволюции происходит ви-
(Наука, М., 2009).
дообразование - разделение во времени и про-
странстве генетически открытой системы (попу-
24. J. B. S. Haldane, Math. Proc. Cambridge Philosoph.
ляции) на генетически закрытые самодостаточ-
Society 23 (7), 838 (1927).
ные системы - виды.
25. D. E. Rozen, J. A. G. M. De Visser, P. J. Gerrish, Curr.
Biol. 12 (12), 1040 (2002).
26. H. A. Orr, Genetics 155 (2), 961 (2000).
КОНФЛИКТ ИНТЕРЕСОВ
27. O. Hallatschek, Proc. Natl. Acad. Sci. USA 108 (5),
Авторы заявляют об отсутствии конфликта
1783 (2011).
интересов.
28. É. Brunet, I. M. Rouzine, and C. O. Wilke, Genetics
179 (1), 603 (2008).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
29. D. S. Fisher, J. Statistical Mechanics: Theory and Ex-
periment 2013 (1), P01011 (2013).
Настоящая работа не содержит описания ка-
ких-либо исследований с использованием людей
30. M. Kimura, et al., Nature 217 (5129), 624 (1968).
и животных в качестве объектов.
31. B. Charlesworth, M. T. Morgan, and D. Charlesworth,
Genetics 134 (4), 1289 (1993).
32. I. M. Rouzine, A. Rodrigo, and J. M. Coffin, Microbiol.
СПИСОК ЛИТЕРАТУРЫ
Mol. Biol. Rev 65 (1), 151 (2001).
1. N. H. Barton, Genetics 140 (2), 821 (1995).
33. O. Hallatschek and L. Geyrhofer, Genetics 202 (3),
2. D. A. Kessler, H. Levine, D. Ridgway, and L. Tsimring,
1201 (2016).
J. Stat. Phys. 87 (3-4), 519 (1997).
34. L. S. Tsimring, H. Levine, and D. A. Kessler, Phys.
3. H. A. Orr, J. Theor. Biol. 238 (2), 279 (2006).
Rev. Lett. 76 (23), 4440 (1996).
4. M. M. Desai and D. S. Fisher, Genetics 176 (3), 1759
35. B. H. Good and M. M. Desai, Genetics 204 (3), 1249
(2007).
(2016).
5. J. H. Gillespie, Theor. Popul. Biol. 23 (2), 202 (1983).
36. J. M. J. Travis and E. R. Travis, Proc. Roy. Soc. Lond.
6. H. A. Orr, Nature Rev. Genetics 6 (2), 119 (2005).
Ser. B: Biol. Sci. 269 (1491), 591 (2002).
БИОФИЗИКА том 65
№ 3
2020
624
ГАРАЕВА и др.
37. A. C. Shaver, et al., Genetics 162 (2), 557 (2002).
48. H. L. Frisch and J. M. Hammersley, J. Soc. Indust.
Appl. Math. 11 (4), 894 (1963).
38. J. B. André and B. Godelle, Genetics 172 (1), 611
(2006).
49. N. Fountoulakis, Internet Mathematics 4 (4),
329
(2007).
39. M. M. Desai and D. S. Fisher, Genetics 188 (4), 997
50. T. N. Starr, L. K. Picton, and J. W. Thornton, Nature
(2011).
549 (7672), 409 (2017).
40. B. Charlesworth, Genetics Res. 55 (3), 199 (1990).
51. P. W. Anderson, Science 177 (4047), 393 (1972).
41. N. H. Barton, Genetics Res. 65 (2), 123 (1995).
52. A. E. Sidorova, N. T. Levashova, V. A. Tverdislov, and
42. S. P. Otto and N. H. Barton, Genetics 147 (2), 879
A. Y. Garaeva, Moscow University Physics Bull. 74 (5),
(1997).
438 (2019).
53. M. Nei and Y. Imaizumi, Heredity 21, 183 (1966).
43. J. Hey, Genetics 149 (4), 2089 (1998).
54. J. F. Crow and N. E. Morton, Evolution 9 (2), 202
44. A. E. Sidorova and V. A. Tverdislov, Moscow Universi-
(1955).
ty Physics Bull. 67 (2), 213 (2012).
55. W. Ebeling, A. Engel, and R. Feistel, in Physik der Evo-
45. J. Balogh and B. G. Pittel, Random Structures & Algo-
lutionsprozesse (Akademie-Verlag, Berlin, 1990), p. 63.
rithms 30 (1-2), 257 (2007).
56. В. Эбелинг, А. Энгель и Р. Файстель, Физика
46. H. Kesten, Percolation theory for mathematicians
процессов эволюции (УРСС, М., 2001).
(Birkhäuser, Boston, 1982).
57. H. H. Rosenbrock, Comput. J. 5 (4), 329 (1963).
47. C. McDiarmid, in Combinatorial Optimization II
58. E. Santiago and A. Caballero, Genetics 204 (3), 1267
(Springer, Berlin, Heidelberg, 1980), pp. 17-25.
(2016).
A Percolation Natural Selection Grid as a Deterministic
and Random Processes Switch in the Mutation Flow
A.Ya. Garaeva, A.E. Sidorova, N.T. Levashova, and V.A. Tverdislov
Faculty of Physics, Lomonosov Moscow State University, Leninskie Gory 1/2, Moscow, 119991 Russia
The basic models of mutation processes during selection and drifting are reviewed. A fundamentally new
model of mutation flow through the percolation lattice of selection at the population level is presented.
The natural selection of optimal variants in the evolutionary flow of mutations is considered as a percolation
filter - a physical tool that forms the selection mechanism. The model is based on concepts of self-organiza-
tion in hierarchical structures of species formation, where trigger properties of a cell (node) as a valve deter-
mine the deterministic component of fixation of new mutations, and the drift determines the element of ran-
domness in the process of fixation of new mutations in mutation flows. Modes of fixation of mutations de-
pending on a ratio of the cluster size of mutation carriers to the general size of a population, selection and drift
coefficients in a sequence of generations at a population level are obtained. The lower threshold of the perco-
lation selection grid was determined as a share of new mutations fixed under the influence of deterministic
selection processes for minimum reproducing cluster size.
Keywords: natural selection, self-organization, fluctuations, bifurcations, percolations, prohibiting and permitting
mutations
БИОФИЗИКА том 65
№ 3
2020