БИОФИЗИКА, 2020, том 65, № 4, с. 670-675
МОЛЕКУЛЯРНАЯ БИОФИЗИКА
УДК 577.323
ИЗМЕНЕНИЕ ВЫХОДНОГО СИГНАЛА ДНК-БИОСЕНСОРА,
ИНДУЦИРОВАННОГО АДСОРБЦИЕЙ ЛИГАНДОВ НА ДУПЛЕКСЫ ДНК
В ФЛУКТУИРУЮЩЕЙ СРЕДЕ
© 2020 г. В.Б. Аракелян*, А.П. Антонян**, М.А. Парсаданян**,
М.А. Шагинян**, П.О. Вардеванян**
*Факультет физики Ереванского государственного университета, ул. А. Манукяна, 1, Ереван, 0025, Армения
**Факультет биологии Ереванского государственного университета, ул. А. Манукяна, 1, Ереван, 0025, Армения
E-mail: v.arakelyan@ysu.am
Поступила в редакцию 25.11.2019 г.
После доработки 14.04.2020 г.
Принята к публикации 17.04.2020 г.
Исследовано связывание лигандов с ДНК-дуплексами в ДНК-биосенсоре, когда под воздействием
флуктуаций среды (внешнего шума) флуктуирует количество лигандов в растворе. Для случая
малого заполнения получено стохастическое дифференциальное уравнение мультипликативного
типа, описывающее изменение во времени числа адсорбированных лигандов на ДНК-дуплексах.
Вычислены средняя величина и время релаксации выходного сигнала ДНК-биосенсора,
обусловленные адсорбцией лигандов на ДНК-дуплексы в флуктуирующей среде. Показано, что
мультипликативный шум уменьшает среднюю величину выходного сигнала ДНК-биосенсора.
Уменьшение сигнала имеет пороговый характер
- при некотором соотношении между
параметрами адсорбции и интенсивностью внешнего шума не происходит уменьшение сигнала.
Показано, что с увеличением интенсивности внешнего шума увеличивается время релаксации
выходного сигнала ДНК-биосенсора.
Ключевые слова: адсорбция лигандов, флуктуация среды, мультипликативный шум, ДНК-биосенсоры.
DOI: 10.31857/S0006302920040067
В настоящее время широко используются ана-
на выходного сигнала ДНК-биосенсора пропор-
литические устройства нового поколения - био-
циональна числу ДНК-дуплексов.
сенсоры. Наиболее важными и актуальными из
Проблеме гибридизации ДНК посвящено
всех видов биосенсоров являются ДНК-биосенсо-
большое число работ и имеются впечатляющие
ры, которые применяются как в фундаментальных
успехи [8-11]. Исследования в области термоди-
исследованиях, так и в прикладных областях - в
намики [8] и кинетики гибридизации ДНК [9]
медицине, сельском хозяйстве, охране окружаю-
представляют большой практический интерес,
поскольку их результаты используются на прак-
щей среды и т. д. [1-5]. Следует отметить, что, в от-
тике при изготовлении и использовании ДНК-
личие от традиционных аналитических устройств,
сенсоров. Следует отметить важную работу [10],
ДНК-биосенсоры обладают высокой избиратель-
где показано, что методы статистической физики
ностью и огромной чувствительностью [6]. Прин-
позволяют исследовать фундаментальную про-
цип работы ДНК-биосенсора чрезвычайно прост
блему биологии - регуляцию генетической экс-
- в основе всех ДНК-биосенсоров лежит регистра-
прессии. Теоретически разработан подход, опи-
ция высокоспецифического распознавания после-
сывающий процесс гибридизации ДНК на мик-
довательностей распределения нуклеиновых кис-
рочипах [11], который служит основой не только
лот [7]. Конструкция ДНК-биосенсора такова, что
для анализа многочисленных эксперименталь-
одноцепочная молекула ДНК иммобилизована на
ных данных по гибридизации ДНК, но и для
подложке и образует слой для распознавания. Ес-
практики изготовления микрочипов.
ли в растворе есть одноцепочные ДНК, компле-
В подавляющем большинстве случаев ДНК-
ментарные иммобилизованной ДНК, то при их ги-
биосенсор «работает» в среде, содержащей боль-
бридизации образуются дуплексы ДНК. Образова-
шое количество разнообразных лигандов, кото-
ние дуплекса активизирует сигнал, который
рые могут адсорбироваться как на дуплексы
передается регистрирующему устройству. Величи-
ДНК, так и на одиночные иммобилизованные
670
ИЗМЕНЕНИЕ ВЫХОДНОГО СИГНАЛА ДНК-БИОСЕНСОРА
671
молекулы ДНК. Наибольший интерес представ-
ганда с ДНК-дуплексом. ДНК-дуплекс предста-
ляет случай, когда изменение выходного сигнала
вим в виде одномерного кристалла с числом ад-
происходит в результате обратимой адсорбции
сорбционных центров N, а лиганд, имеющий на-
лигандов на ДНК-дуплексы. Адсорбированные
много меньшие линейные размеры, при
лиганды неизбежно приводят к изменению вели-
адсорбции занимает n подряд расположенных
чины выходного сигнала ДНК-биосенсора. Рас-
центров адсорбции на ДНК-дуплексе. Если при-
смотрим более естественный случай малости
нять, что число адсорбированных лигандов равно
константы скорости диссоциации ДНК-дуплек-
x, для случая малого заполнения уравнение, опи-
са, когда относительно быстрая адсорбция и де-
сывающее изменение во времени числа адсорби-
сорбция лигандов происходит в условиях посто-
рованных на ДНК-дуплексе лигандов, будет
янства числа ДНК-дуплексов. Отметим, что па-
иметь следующий вид [12, 13]:
раметры среды, в которой
«работет» ДНК-
dx/dt = k1cf(N - (2n - 1)x),
(2)
биосенсор, как правило, не строго постоянны, а
случайно флуктуируют. Эти случайные флуктуа-
где cf - число свободных лигандов в растворе. За-
ции среды (внешний шум) влияют на процесс
метим, что при n = 1 уравнение (2) описывает ад-
формирования выходного сигнала ДНК-биосен-
сорбцию при произвольном заполнении. Чтобы в
сора. Внешние флуктуации не имеют микроско-
дальнейшем при переходе к концентрациям не
пического происхождения и избавиться от них
вводить новые обозначения, будем считать, что
практически невозможно. В работе [12] были
адсорбция происходит в единичном объеме. Рас-
определены характерные особенности выходного
смотрим случай, когда флуктуирует число сво-
сигнала ДНК-биосенсора, когда под воздействи-
бодных лигандов в растворе cf. Чтобы анализ ре-
ем флуктуаций среды флуктуировало число ад-
зультатов сделать однозначным, оставим значе-
сорбционных центров (фактически сродство) на
ния других параметров на уровне их средних
ДНК-дуплексе. Было показано, что шум выход-
значений. В подавляющем больщинстве случаев
ного сигнала ДНК-биосенсора, обусловленный
причиной флуктуации является действие не од-
адсорбцией лигандов на ДНК-дуплексы, являет-
ного выделенного фактора, а действуют множе-
ся ланжевеновским. В данной работе рассмотрен
ство независимых факторов. В этом случае, как
случай, когда под воздействием флуктуаций сре-
известно, их суммарное воздействие, согласно
ды флуктуирует число лигандов в растворе. В
центральной предельной теореме, имеет гауссово
этом случае шум, связанный с адсорбцией и де-
распределение, cf(t) можно представить в виде
сорбцией лигандов на ДНК-дуплексы, оказыва-
суммы среднего c и гауссового шума ξ(τ), сред-
ется мультипликативным. Определены харатер-
f
ные особенности выходного сигнала ДНК-био-
нее значение которого равно нулю, т.е.
ξ(t)
=
0:
сенсора, обусловленные адсорбцией лигандов на
2
ДНК-дуплексы.
c
(t) = c
+σ
⋅ ξ(t),
(3)
f
f
с
где σ2с - интенсивность шума. Примем, что ин-
ТЕОРЕТИЧЕСКАЯ ЧАСТЬ
тенсивность шума не зависит от t, т.е. имеем ста-
ционарный шум. Принимаем, что время корреля-
Чтобы исследовать «работу» ДНК-биосенсора
ции флуктуаций внешней среды намного меньше
в флуктуирующей среде, исключим влияние
характерного времени изменения переменной в
осложняющих факторов. Для этой цели примем,
системе, так что, согласно работе [14], в выраже-
во-первых, что ДНК-биосенсор работает в про-
нии (3) можно перейти к пределу гауссового бело-
странственно-однородной системе, т.е. нет диф-
го шума. Таким образом, принимаем, что имеем
фузионных ограничений в кинетике связывания
гауссов белый шум с характеристиками:
ξ(t)
=
0,
лиганда с ДНК; во-вторых, что ДНК-биосенсор
работает в макроскопически большой системе,
ξ(0)⋅ξ(t)
=δ(t), где δ(t) - дельта-функция. Под-
такой, что можно пренебречь внутренними флук-
ставив выражение (3) в выражение (2), получим
туациями, интенсивность которых уменьшается с
увеличением размеров системы. Квазихимиче-
ская реакция, описывающая связывание лиганда
dx = f x)
c
+σ
c
⋅
g x)⋅ξ(t),
dt
с адсорбционным центром на ДНК-дуплексе, бу-
дет иметь вид
f x)
=
k
c
(N
−
(2n- x)
-
k
x,
(4)
c
1
f
−1
k1
g x)
=
k
1
(N
−
(2n- x).
L + M ↔ LM,
(1)
k-1
Чтобы не вводить новых обозначений в урав-
нении (4) и далее, знак черты усреднения над па-
где LM - комплекс лиганда с адсорбционным
раметрами опущен, оставлен лишь у параметра,
центром на ДНК-дуплексе, k1 и k-1 - константы
флуктуирующего под воздействием внешнего
скоростей образования и распада комплекса ли-
шума. Поскольку, как это видно из уравнения (4),
БИОФИЗИКА том 65
№ 4
2020
672
АРАКЕЛЯН и др.
случайная функция умножается на переменную
нение для моментов. Поскольку флуктуирующий
x, то уравнение (4) относят к классу мультиплика-
параметр аппроксимирован гауссовым белым
тивных стохастических дифференциальных урав-
шумом, то, согласно работе [14], в этом случае
нений. Процедура решения уравнения (4) следу-
стохастическое дифференциальное уравнение (4)
ющая. Вначале для уравнения (4) выписывается
интерпретируется в смысле Стратановича и соот-
соответствующее уравенение Фоккера-Планка,
ветствующее уравнение Фоккера-Планка для не-
а затем, используя это уравнение, получают урав-
го, как показано в работе [14], имеет вид
2
2
2
∂P x,t)
∂
σ
σ
∂
2
с
c
=-
f x)
+
g x)g'(x)
P x,t)
+
g x)P(x,t)
,
(5)
c
2
(
)
∂t
∂x
2
2
∂x
где P(x,t) - вероятность того, что в системе в момент ДНК-дуплексе лигандов. Для получения уравнения
времени t имеется ровно x адсорбированных на для моментов перепишем уравнение (5) в виде
2
∂P x,t)
∂
1
∂
=-
(A x)P(x,t))
+
(B x)P(x,t)),
2
∂t
∂x
2∂x
2
σ
c
A x)
=
c
f x)
+
g x)g'(x),
(6)
2
2
2
B x)
=σ
g x).
c
В работе [15] приводится подробный вывод
Для случая малого шума, когда можно принять
уравнений для моментов произвольного порядка.
α > 0, из уравнения (9) при стремлении времени к
Следуя работе [15], легко получить дифференци-
бесконечности получим стационарное значение
среднего числа адсорбированных на ДНК-дуп-
альное уравнение для
, которое имеет вид
x
лексе лигандов
в виде
( )
x
st
dx = A x).
(7)
2
σ
2
dt
c
k
1
c
f
N
-
k
1
(2n
−1)N
2
Подставив в уравнение (7) значение A(x) из
(
x
)
=
(10)
st
2
σ
2
2
уравнения (6) с учетом явных выражений для fc(x)
c
k
−1
+
(2
n
−1)k
1
c
f
−
k
1
(2n
−1)
и g(x), получим следующее окончательное урав-
2
нение, описывающее изменение во времени
Принимаем, что выходной сигнал ДНК-био-
среднего числа адсорбированных на ДНК-дуп-
сенсора через единицу площади биосенсора I(t),
обусловленный адсорбцией лигандов на ДНК-
лексе лигандов x в виде
дуплексе, пропорциален числу адсорбированных
на ДНК-дуплексе лигандов x(t), т.е. I(t) = γx(t), где
dx
= -αx
+ β,
γ - коэффициент пропорциональности. В этом
dt
случае среднее значение выходного сигнала
2
σ
c
2
2
α=
k
+
(2n
−1)k
c
−
k
(2n
−1)
,
(8)
ДНК-биосенсора равно
I(t)=γx(t), т.е.
−1
1
f
1
2
2
β
σ
2
I(t)
= γ
(1−
exp(−αt
))
(11)
c
β=
k
1
c
f
N
−
k
1
(2n
−1)
N
α
2
а стационарное значение сигнала равно
Принимаем, что в начальный момент времени
на ДНК-дуплексе не было лигандов, т.е. при на-
2
σ
2
c
k
1
c
f
N
-
k
1
(2n
−
1)N
чальном условии
. Из решения уравнения
x(0) = 0
2
(
I
)
= γ
(12)
(8) получим следующее выражение для среднего
st
2
σ
2
2
c
числа адсорбированных на ДНК-дуплексе лиган-
k
+
(2
n
−1)
k
c
−
k
(2n
−1)
−1
1
f
1
2
дов:
Важной характеристикой выходного сигнала
β
ДНК-биосенсора является время релаксации
x(t)
=
(1
−
exp(−αt
))
(9)
сигнала. Из формулы (11) следует, что время ре-
α
БИОФИЗИКА том 65
№ 4
2020
ИЗМЕНЕНИЕ ВЫХОДНОГО СИГНАЛА ДНК-БИОСЕНСОРА
673
0.8
0.6
0.4
0.2
0.0
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Концентрация лигандов в растворе, усл. ед.
Рис. 1. Зависимости сигнала ДНК-биосенсора Y от концентрации лигандов в растворе X. Графики рассчитаны по
формуле (15) при n = 1. Верхняя кривая соответствует детерминированному случаю, когда отсутствует внешний шум
(σc = 0). Средняя кривая соответствует случаю, когда C = D = 0.2, нижняя кривая когда C = D = 0.4.
лаксации выходного сигнала ДНК-биосенсора
шума сигнал отсутствует, когда число лигандов в
равно
растворе меньше некоторого определенного зна-
чения, которое определяется из условия равен-
-1
2
ства нулю числителя в выражении (12). Это значе-
σ
2
2
c
τ=
k
+
(2n
−1)k
c
−
k
(2n
−1)
(13)
−1
1
f
1
ние равно
2
2
*
σ
k
(2n
−
Из уравнения (13) видно, что с увеличением
c
1
c
=
1).
(14)
f
концентрации лигандов в растворе время релак-
2
сации сигнала уменьшается.
Для удобства построения графиков и анализа
результатов, перепишем выражение (12) в безраз-
мерном виде:
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
X -C
Как и следовало ожидать, при отсутствии
Y
=
,
внешнего шума, т.е. при условии σс = 0, стацио-
1+
(2n
−1)X -D
нарное значение среднего числа адсорбирован-
Y
=
I
/ (Nγ),
X =Kc
,
K = k
/
k-
,
(15)
(
)
f
1
1
ных на ДНК-дуплексе лигандов (10) в точности
st
2
2
совпадает с соответствующим решением детер-
σ
Kk
σ
Kk
2
c
1
c
1
C
=
(2n
−1),
D
=
(2n
−1)
,
минированного уравнения (2) (для определения
2
2
стационарного значения адсорбированных на
где Y - безразмерный сигнал ДНК-биосенсора,
ДНК-дуплексе лигандов следует приравнять пра-
X - безразмерная концентрация лигандов в рас-
вую часть уравнения (2) к нулю и найти xst). Как
творе, K - константа равновесия квазихимиче-
следует из уравнений (10) и (11), стационарное
ской реакции (1). Графики зависимостей (15)
значение среднего числа адсорбированных на
представлены на рис. 1. Кривые построены при
ДНК-дуплексе лигандов и среднее значение сиг-
n = 1 (при других значениях n кривые качествен-
нала зависят от интенсивности внешнего шума
но не отличаются от случая n = 1). Из рис. 1 вид-
σ2с, что является прямым следствием мультипли-
но, что наличие внешнего мультипликативного
кативности шума [14]. Заметим, что в случае ад-
шума приводит к уменьшению сигнала ДНК-
дитивного ланжевеновского шума это не так -
биосенсора, обусловленного адсорбцией и де-
стационарное значение среднего числа адсорби-
сорбцией лигандов на ДНК-дуплексы. Из рис. 1
рованных на ДНК-дуплексе лигандов не зависит
также видно, что если интенсивность внешнего
от интенсивности внешнего шума [12]. Из урав-
шума отлична от нуля, то существует область ма-
нения (11) видно, что зависимость среднего вы-
лых чисел лигандов в раствореc (малых концен-
ходного сигнала ДНК-биосенсора, обусловнен-
f
ного адсорбцией лигандов на ДНК-дуплексе, от
траций), при которых отсутствует сигнал ДНК-
интенсивности внешнего шума имеет пороговый
биосенсора. Из выражения (12) также следует, что
характер. Анализ уравнения (11) показывает, что
с увеличением как интенсивности внешнего шу-
при заданном уровне интенсивности внешнего
ма, так и k1 эта область расширяется.
БИОФИЗИКА том 65
№ 4
2020
674
АРАКЕЛЯН и др.
1.5
1.0
0.5
0.0
0.0
0.5
1.0
1.5
2.0
Концентрация лигандов в растворе, усл. ед.
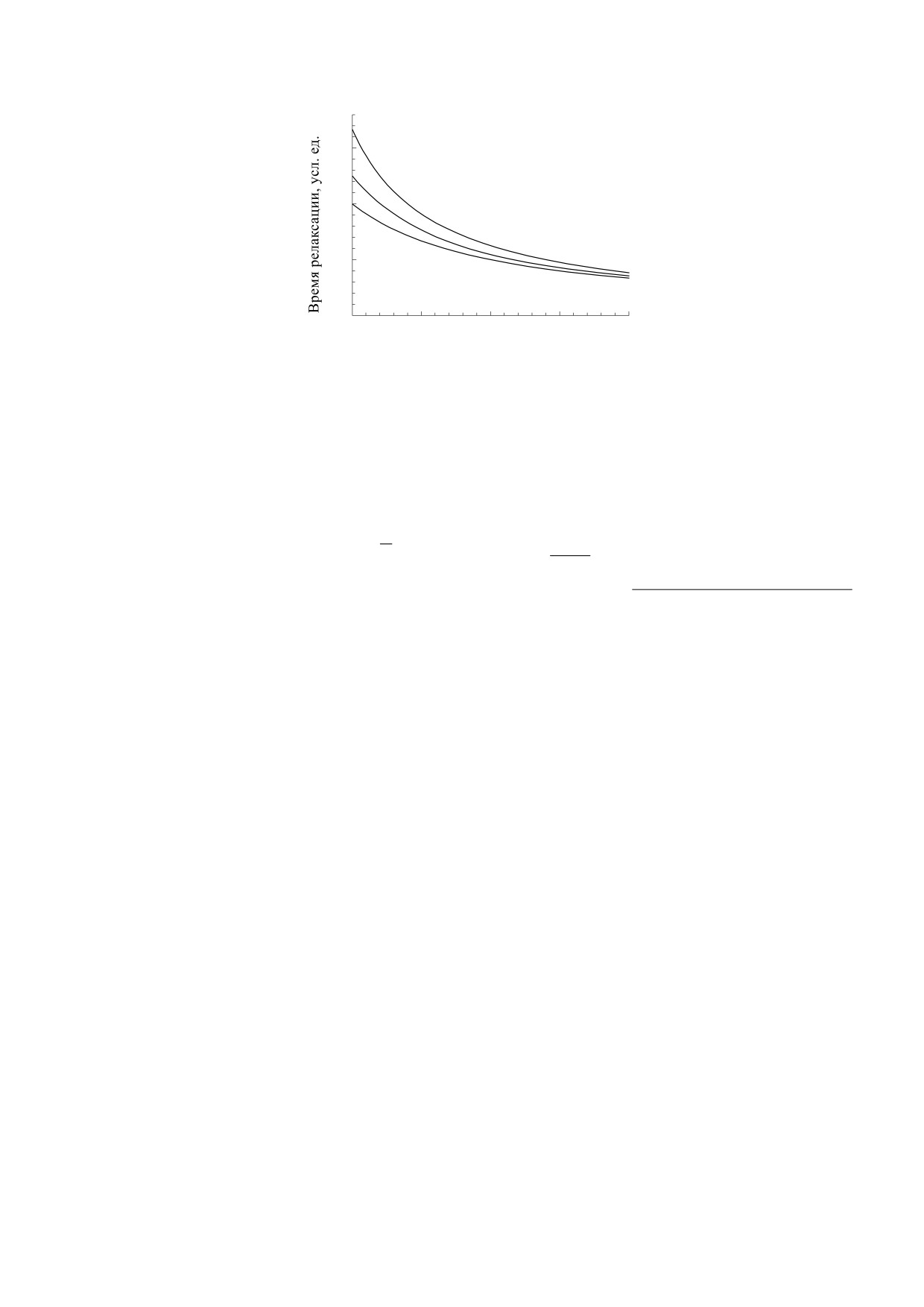
Рис. 2. Зависимости времени релаксации от концентрации лигандов в растворе. Графики рассчитаны по формуле (16)
при n = 1. Нижняя кривая соответствует детерминированному случаю, когда отсутствует внешний шум ((σc = 0). Сред-
няя кривая соответствует случаю, когда C = D = 0.2, верхняя кривая - C = D = 0.4.
Для удобства построения графиков и анализа результатов перепишем уравнение (13) в безразмер-
ном виде:
Y
=
(1 +
(2n
−1)X -
)1
2
(16)
σ
c
Kk
1
2
Y =τk
−1
,
X = Kc
f
,
K = k
1
/
k
−1
,
D
=
(2n
−1)
,
2
где Y - безразмерное время релаксации сигнала,
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
X - безразмерная концентрация лигандов в рас-
Настоящая работа не содержит описания ка-
творе, K - константа равновесия квазихимиче-
ких-либо исследований с использованием людей
ской реакции (1). На рис. 2 представлена зависи-
и животных в качестве объектов.
мость времени релаксации выходного сигнала
ДНК-биосенсора от концентрации лигандов в
растворе. Кривые построены при n = 1 (при дру-
СПИСОК ЛИТЕРАТУРЫ
гих значениях n кривые качественно не отлича-
1. S. K. Metkar and K. Girigoswami, Biocatalysis and Ag-
ются от случая n = 1). Из рис. 2 видно, что с уве-
ricult. Biotech. 17, 271 (2019).
личением интенсивости как внешнего шума, так
2. M. S. Mufamadi and P. R. Sekhejane, in Nanotechnol-
и k1 увеличивается время релаксации, причем
ogy. An Agricultural Paradigm (Springer, Singapore,
при малых концентрациях лигандов это увеличе-
2017), pp. 263-278.
ние проявляется более значительно.
3. T. G. Drummond, M. G. Hill, and J. Barton, Nature
Biotechnol. 21 (10), 1192 (2003).
4. J. Wang, Anal. Chem. Acta 469, 63 (2002).
ЗАКЛЮЧЕНИЕ
5. E. Palecek, M. Fojta, and F. Jelen, Bioelectrochemistry
56, 85 (2002).
Таким образом, в работе в условиях, когда под
6. Ю. А. Ахапкин и др., Биотехника
- новое
воздействием внешнего шума флуктуирует число
направление компьютеризации (Наука, М., 1990).
лигандов в растворе, вычислены среднее число
7. V. Kavita, J. Bioengineer. Biomed. Sci. 7 (2), 222
лигандов, адсорбированных на ДНК-дуплексе, а
(2017).
также средняя величина сигнала ДНК-биосенсо-
8. A. Halperin, A. Buhot, and E. B. Zhulina, J. Phys.
ра, обусловленная адсорбцией лигандов на ДНК-
Condens. Matter 18, S463 (2006).
дуплексах ДНК-биосенсора. Определены неко-
9. B. A. Baker and V.T. Milam, Nucl. Acids Res. 39 (15),
торые характерные особенности выходного сиг-
e99 (2011).
нала ДНК-биосенсора.
10. М. В. Головкин , Ю. Д. Нечипуренко и Г. В. Гурс-
кий, Биофизика 54 (4), 581 (2009).
КОНФЛИКТ ИНТЕРЕСОВ
11. Ю. Д. Нечипуренко, в кн. Физико-химические
механизмы и регуляция процессов трансформации
Авторы заявляют об отсутствии конфликта
энергии в биологических структурах, под ред.
интересов.
Г. Ю. Ризниченко и А. Б. Рубина (Институт
БИОФИЗИКА том 65
№ 4
2020
ИЗМЕНЕНИЕ ВЫХОДНОГО СИГНАЛА ДНК-БИОСЕНСОРА
675
компьютерных исследований, М.-Ижевск, 2017),
14. В. Хорстхемке и Р. Лефевр, Индуцированные шумом
сс. 522-530.
переходы. Теория и применение в физике, химии и
12. А. Т. Карапетян, Г. А. Аветисян и В. Б. Аракелян,
биологии (Мир, М., 1987).
Докл. НАН РА 110 (4), 376 (2010).
15. С. А. Ахманов, Ю. Е. Дьяков и А. С. Чиркин,
13. V. Arakelyan, Y. Babayan, and G. Potikyan, J. Biomol.
Введение в статистическую радиофизику и оптику
Struct. Dyn. 18, 231 (2000).
(Наука, М., 1981).
Adsorption of Ligands on DNA Duplexes under the Influence of the Fluctuating Medium
Causes a Change in DNA-Biosensor Output Signal
V.B. Arakelyan*, A.P. Antonyan**, M.A. Parsadanyan**, M.A. Shahinyan**, and P.O. Vardevanyan**
*Faculty of Physics, Yerevan State University, ul. A. Manoogiana 1, Yerevan, 0025 Armenia
**Faculty of Biology, Yerevan State University, ul. A. Manoogiana 1, Yerevan, 0025 Armenia
In this study we report on ligand binding to DNA-duplexes in DNA-biosensor when the number of ligands
in the solution fluctuates under the influence of the fluctuating medium (external noise). A multiplicative
stochastic differential equation, which takes into account the time-dependent change in the number of bound
ligands to DNA-duplexes is obtained for a system with small amounts of the studying samples. DNA-biosen-
sor output signal average value and relaxation time that depend on adsorption of ligands on DNA-duplexes
in fluctuating medium have been calculated. It is shown that multiplicative noise decreases the average value
of DNA-biosensor output signal. The deterioration in the signal has its threshold level -at a certain ratio be-
tween the adsorption parameters and the intensity of external noise no deterioration in the signal occurs. It is
shown that the relaxation time of the output signal of DNA-biosensor increases along with increasing external
noise intensity.
Keywords: adsorption of ligands, medium fluctuation, multiplicative noise, DNA-biosensors
БИОФИЗИКА том 65
№ 4
2020