БИОФИЗИКА, 2020, том 65, № 4, с. 705-712
БИОФИЗИКА КЛЕТКИ
УДК 577.344.2, 577.344.3
ФОТОТЕРМИЧЕСКАЯ ИНАКТИВАЦИЯ МИКРООРГАНИЗМОВ
ПРИ РЕЛАКСАЦИИ ВЫСОКОВОЗБУЖДЕННЫХ
СОСТОЯНИЙ СЕНСИБИЛИЗАТОРОВ
© 2020 г. С.Н. Летута, С.Н. Пашкевич, А.Т. Ишемгулов, А.Н. Никиян
Оренбургский государственный университет, 460018, Оpенбуpг, пpосп. Победы, 13
E-mail: ppsnya@yandex.ru
Поступила в редакцию 08.11.2019 г.
После доработки 08.11.2019 г.
Принята к публикации 22.05.2020 г.
Обсуждается инактивация планктонных бактерий Escherichia coli и Bacillus subtilis ударными
акустическими волнами, возникающими при быстром образовании и схлопывании пузырьков пара
в среде, локально нагретой до температуры кипения. Локальный нагрев среды осуществлялся за
счет тепла, выделяемого в ходе релаксации высоких электронно-возбужденных состояний
экзогенных молекул органических красителей. Красители возбуждались наносекундными
импульсами лазерного излучения видимого диапазона. Высокие электронно-возбужденные
состояния заселялись в результате ступенчатого поглощения молекулами двух квантов лазерного
излучения. Исследована зависимость эффективности инактивации микроорганизмов от
концентрации красителей в растворах, плотности мощности возбуждающего излучения и
расстояния до источника генерации ударных волн.
Ключевые слова: экзогенные термосенсибилизаторы, ударные акустические волны, инактивация
микроорганизмов.
DOI: 10.31857/S0006302920040109
в тонком периферийном слое. Для инактивации
Альтернативные стратегии борьбы с патоген-
бактерий в более глубоких слоях необходимо
ными микроорганизмами становятся все более
«включить» другие физические процессы.
востребованными в связи с увеличением количе-
ства штаммов, резистентных к действию тради-
В настоящей работе представлены результаты
ционных антимикробных препаратов [1]. Среди
исследования повреждений планктонных бакте-
перспективных способов немедикаментозного
рий ударными акустическими волнами. Такие
воздействия на бактерии и грибы, к которому
волны возникают в среде при быстром образова-
микробы не могут выработать иммунитет, выде-
нии и схлопывании пузырьков пара в локально
ляют фотодинамическую обработку - совместное
нагретых областях. Быстрый локальный нагрев
деструктивное действие света и фотосенсибили-
среды происходит при безызлучательной релак-
заторов на клетки. Механизм фотодинамическо-
сации короткоживущих высоковозбужденных
го действия основан на селективном окислении
электронных состояний (ВВЭС) молекул-сенси-
субстрата активным кислородом, генерируемым
билизаторов.
фотосенсибилизаторами [2-5]. В качестве фото-
ВВЭС красителей эффективно заселяются при
сенсибилизаторов обычно используют молекулы
облучении молекул наносекундными лазерными
органических красителей с большим квантовым
импульсами плотности мощности Р ≥ 5 MВт/см2
выходом в триплетное состояние [6, 7].
в результате ступенчатого поглощения двух кван-
тов возбуждающего излучения [10, 11]. Релакса-
Основные ограничения фотодинамического
ция ВВЭС молекул преимущественно безызлуча-
действия связаны с малой глубиной проникнове-
тельная, и поглощенная энергия очень быстро
ния фотосенсибилизаторов и света в биологиче-
трансформируется в тепло. Если термосенсиби-
ские среды. В некоторых из них, например в
лизатор локализован непосредственно в клетке
биопленках, практически нет кислорода [8, 9], и
или ее мембране, тепловыделение может иници-
фотодинамическое действие проявляется только
ировать ее гибель за счет гипертермии. Когда тер-
Сокращениe: ВВЭС - высоковозбужденные электронные
мосенсибилизатор находится вне клетки, тепло
состояния.
передается растворителю с последующим образо-
705
706
ЛЕТУТА и др.
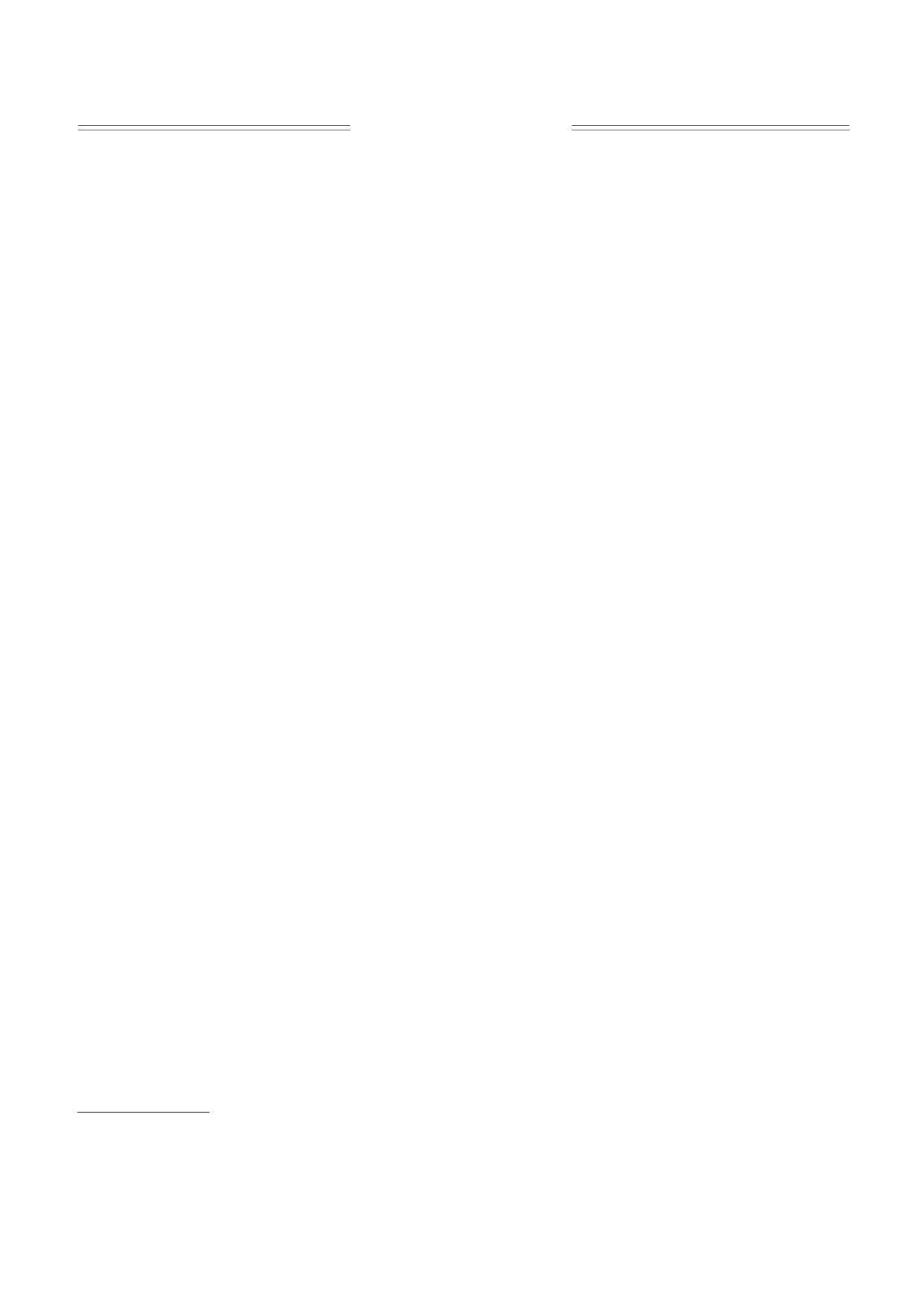
Для фотоинактивации микроорганизмов и
6
изучения ударных волн в растворах красителей
использовали экспериментальную установку,
1
5
4
схема которой показана на рис. 1. Растворы поме-
щали в прямоугольную кварцевую кювету 1 сече-
нием 5 × 10 мм и облучали через ее торцевую стен-
ку. Источником возбуждения служил импульс-
2
3
ный YAG:Nd-лазер
4 (вторая гармоника
6
L
λ = 532 нм, длительность импульса 15 нс).
2
10
Для обеспечения эффективного нагрева среды
L
и формирования пузырьков пара в кювете вблизи
1
границы «стекло-раствор» с помощью собираю-
8
9
щей цилиндрической линзы 6 создавали зону воз-
буждения 2 с поперечным сечением 3.2 × 0.5 мм, в
3
7
которой плотность мощности максимальна в пе-
ретяжке и могла достигать 50 MВт/см2. Протя-
Рис. 1. Экспериментальная установка для инактива-
женность этой зоны вдоль луча возбуждения
ции микроорганизмов и изучения ударных волн: 1 -
кювета с раствором; 2 - область возбуждения раство-
уменьшается с ростом концентрации красителя и
ра; 3 - область зондирования; 4 - твердотельный ла-
увеличивается вместе с плотностью мощности. В
зер, 532 нм; 5 - He-Ne-лазер, 628 нм; 6 - цилиндри-
наших экспериментах излучение накачки полно-
ческая линза; 7 - подвижная платформа; 8 - моно-
стью поглощалось на пути менее 5 мм.
хроматор; 9 - фотоприемник; 10 - фокусирующая
линза. Показано расстояние L между зонами возбуж-
Вдоль перетяжки линзы 6 (или параллельно ей
дения и зондирования. На врезке - конфигурация
установки в плоскости, перпендикулярной зондиру-
на расстоянии L) пропускали зондирующий
ющему лучу.
луч 3 сечением менее 1 мм от маломощного (ме-
нее 1 мВт) He-Ne-лазера 5. Измеряя интенсив-
ность нерассеянной части этого луча, можно су-
ванием и схлопыванием пузырьков пара, в ре-
дить о наведенном поглощении в зоне возбужде-
зультате чего образуются акустические волны,
ния или о наличии рассеивающих центров в виде
способные повредить микроорганизм. Заметим,
пузырьков пара на его пути, а по рефракции
что если в результате каскадной релаксации
луча - о прохождении ударной волны в растворе.
ВВЭС [12, 13] заселяются нижние триплетные
Линза 6 и кювета 1 с раствором располагались на
уровни молекул, то они по-прежнему могут вы-
одной платформе 7, которая могла смещаться
полнять функции фотосенсибилизаторов.
перпендикулярно зондирующему лучу 3. Переме-
щая платформу, можно, не изменяя геометрии
возбуждения, зондировать раствор в кювете на
МАТЕРИАЛЫ И МЕТОДЫ
контролируемом расстоянии L от перетяжки.
Исследованы суточные агаровые культуры кле-
При L = 0 и выключенном He-Ne-лазере свече-
ток Escherichia coli (штамм K12TG1) и Bacillus subtilis
ние зоны возбуждения 2 могло быть собрано на
(штамм 534). Перед облучением бактерии перено-
щели монохроматора 8 (МДР-41, ООО «ОКБ
сили в физиологический раствор. Плотность полу-
Спектр», Санкт-Петербург) с помощью дополни-
ченной суспензии стандартизировали фотометри-
тельной линзы 10, что позволяло получать ин-
чески (длина волны 620 нм, оптическая плотность
формацию о релаксации возбужденных состоя-
D = 0.61 ± 0.01. Для оценки выживаемости бактерий
ний сенсибилизаторов.
после облучения видимым светом в присутствии
Синхронизация запуска лазера и регистриру-
термосенсибилизатора применяли метод подсчета
ющей системы, а также сбор, накопление и обра-
колониеобразующих единиц, количество которых
ботка сигналов производились в автоматизиро-
определялось через 16 ч роста на LB-агаре при тем-
ванном режиме.
пературе 37°С. До облучения и добавления красите-
ля оно составляло (4.50 ± 0.44) ⋅ 107/мл для E. coli и
(32.0 ± 2.3) ⋅ 107/мл для B. subtilis.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Термосенсибилизаторами служили анионные
ксантеновые красители эритрозин и эозин, а так-
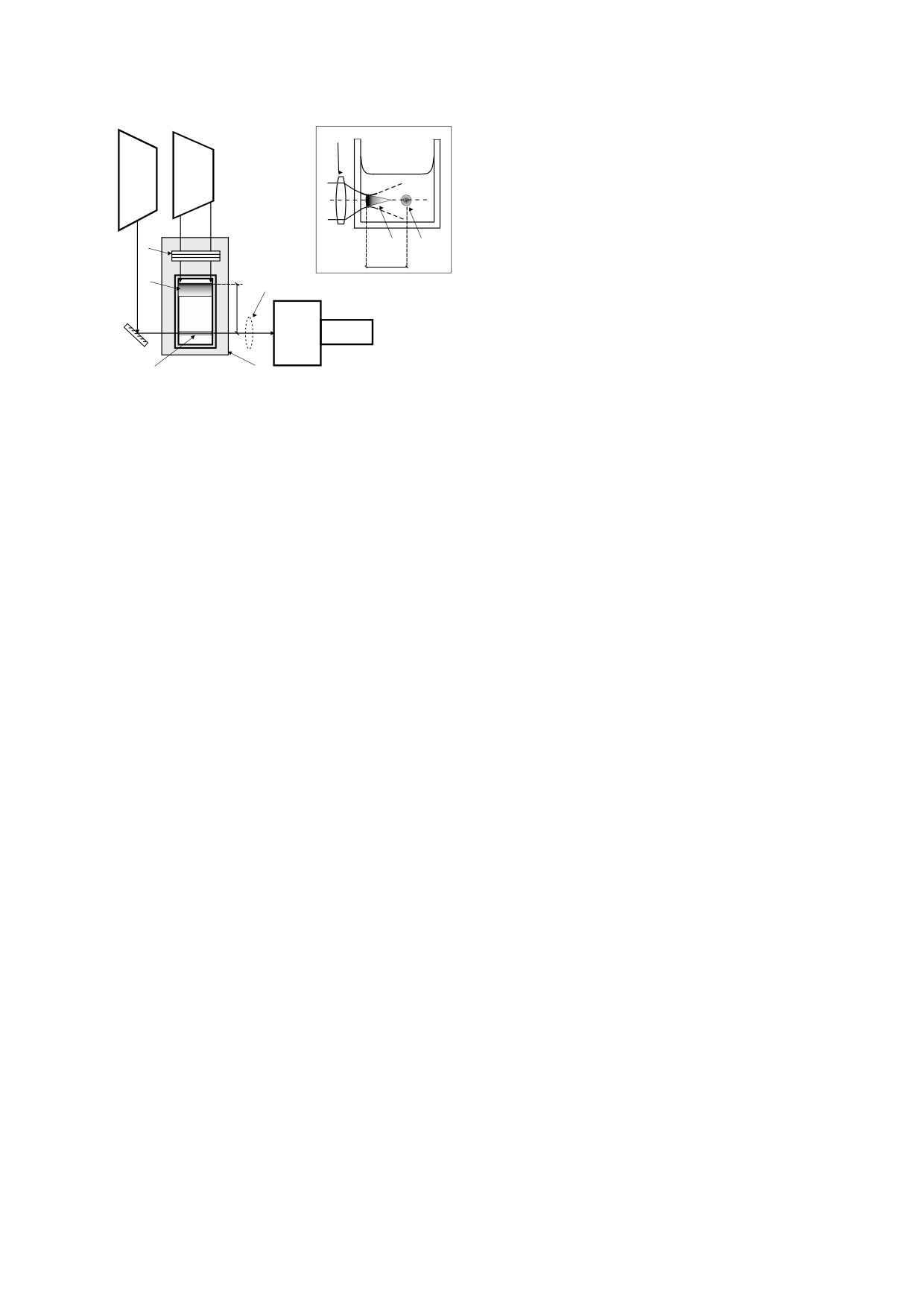
Генерация ударных акустических волн. На
же катионный краситель родамин 6G, которые
рис. 2 представлены кинетические кривые интен-
различаются фотодинамической активностью и
сивности зондирующего луча при импульсном
по-разному взаимодействуют с клеточными стен-
возбуждении эритрозина концентрации 0.25 мМ
ками грамположительных и грамотрицательных
в физиологическом растворе на разных расстоя-
бактерий. Концентрация красителей в растворах
ниях от перетяжки возбуждающего пучка: кривая
составляла 0.01-0.5 мМ.
1 - непосредственно в перетяжке; кривые 2 и 3 -
БИОФИЗИКА том 65
№ 4
2020
ФОТОТЕРМИЧЕСКАЯ ИНАКТИВАЦИЯ МИКРООРГАНИЗМОВ
707
I, отн. ед.
0.30
5
0.25
4
3
0.20
2
6
0.15
4
0.10
1
2
0
0.05
0
2
4
6
8
10
L, мм
0.00
0
1
2
3
4
5
6
7
8
Время, мкс
Рис. 2. Временные зависимости интенсивности нерассеянной части зондирующего луча в физиологическом растворе
с эритрозином (С = 0.25 мМ) на разных расстояниях L от перетяжки возбуждающего пучка (Р = 20 MВт/см2): 1 -
L = 0 мм; 2 - L = 1.2 мм; 3 - L = 2.7 мм; 4 - L = 4.9 мм; 5 - L = 8.5 мм. На врезке - зависимость времени прихода волны
от расстояния до перетяжки.
в зоне возбуждения на некотором расстоянии от
глощение уже не проявляется, что видно из кри-
перетяжки, кривые 4 и 5 - вне зоны возбуждения.
вых 4 и 5 (L = 4.9 и 8.5 мм соответственно).
На всех кривых присутствуют острые провалы,
На интенсивность нерассеянной части зонди-
которые, по нашему мнению, соответствуют мо-
рующего луча оказывают влияние три явления -
ментам пересечения зондирующего луча фрон-
рассеяние света на долгоживущих пузырьках, на-
том ударной волны, источником которой являют-
веденное поглощение возбужденными молекула-
ся родившиеся в перетяжке пузырьки. Косвен-
ми красителя и пересечение луча ударной волной.
ным доказательством этому является график
Рассеяние на пузырьках проявляется только для
зависимости времени появления провала от рас-
луча, зондирующего перетяжку: в этой позиции
стояния L от точки зондирования до перетяжки.
(кривая 1, L = 0) ступенчатое изменение интен-
Обратный наклон этой прямой соответствует
сивности сохраняется вплоть до нескольких мил-
скорости звука в среде, которая измерена нами
лисекунд (не показано на рисунке). Вместе с дол-
при температурах раствора 0, 26 и 80°C. Получен-
гоживущим рассеянием на пузырьках в точке
ные значения вместе со справочными данными
L = 0 имеет место короткоживущее наведенное
для дистиллированной воды при атмосферном
триплет-триплетное поглощение света возбуж-
давлении приведены в таблице.
денными молекулами красителя, которое харак-
терно для кривых 1 и 2 (L = 0, 1.2 мм) и в значи-
Таким образом, при указанных выше диапазо-
тельно меньшей степени для кривой 3 (L = 2.7 мм).
нах концентраций красителей и плотности мощ-
Отметим, что наличие триплетных состояний
ности возбуждающего излучения в кювете с кра-
красителя в присутствии молекулярного кисло-
сителем можно выделить три зоны, в которых
рода в этих областях может повлечь генерацию
возможны различные воздействия на микроорга-
активных форм кислорода. Кривая 3, очевидно,
низмы. В области перетяжки возможна гипертер-
соответствует дальнему краю зоны возбуждения.
мия бактерий, воздействие на них ударной вол-
На расстояниях L ≥ 5 мм триплет-триплетное по-
ной, повреждение активными формами кислоро-
Зависимость скорости волны от температуры раствора
Скорость волны в
Температура, °С
Скорость волны, м/с
дистиллированной воде, м/с [21]
0
1410
1402
26
1505
1499
80
1555
1554
БИОФИЗИКА том 65
№ 4
2020
708
ЛЕТУТА и др.
из которых в триплетное состояние выше, чем с
7
S1-уровня [15-17].
6
В результате баланс концентраций молекул в
5
синглетной и триплетной подсистемах молекул
смещается в сторону увеличения доли триплетов
4
из-за включения последовательности переходов
3
S1 → Sn → Тm при большой плотности мощности. В
свою очередь, к переходам S1 → T1 добавляются
2
переходы Тm → T1, что и приводит к росту концен-
1
трации T1-состояний и увеличению интенсивно-
сти фосфоресценции.
0
0
5
10
15
20
25
30
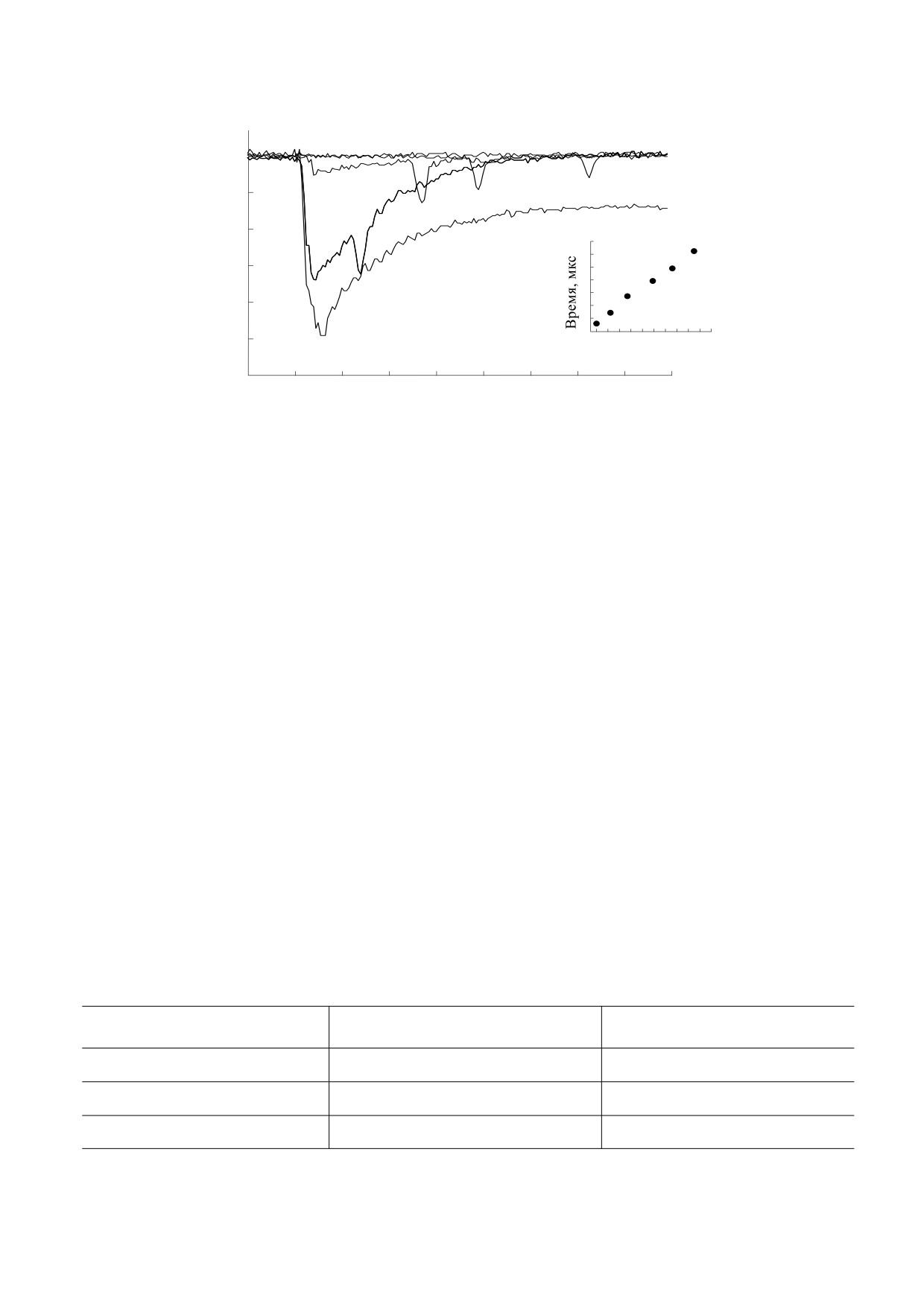
При плотности мощности возбуждающего из-
Р, МВт/см2
лучения Р < 0.5 MВт/см2 ударные волны в раство-
рах с ксантеновыми красителями имеют
Рис. 3. Зависимость интенсивности фосфоресценции
эозина (С = 0.25 мМ) в физиологическом растворе от
очень низкую интенсивность и их эксперимен-
плотности мощности возбуждающего излучения
тальная регистрация затруднена. Поскольку
(λвозб
= 532 нм).
квантовый выход φТ интеркомбинационной кон-
версии S1 → T1 в триплетное состояние в воде у
ксантеновых красителей большой (от 0.71 у эози-
да и перенос энергии с ВВЭС связанных молекул
на до 0.98 у эритрозина [18]), то при длительности
красителей на мембраны или внутриклеточные
импульса 15 нс значительная часть молекул успе-
структуры. В промежуточной области зоны воз-
вает не только перейти в T1-состояние, но и мно-
буждения, где присутствуют молекулы сенсиби-
гократно совершить безызлучательные переходы
лизаторов в триплетном состоянии, гипертермия
T1 ↔ Тm, каждый из которых дает свой вклад в
и перенос энергии маловероятны, но возможно
нагрев среды. Однако при общей малой энергии
химическое повреждение бактерий активными
накачки интенсивность акустических волн неве-
формами кислорода и действие ударной волны.
лика.
Наконец, вне зоны возбуждения единственным
потенциально опасным фактором повреждения
В растворах с родамином 6G при слабой на-
микроорганизмов выступает ударная волна.
качке акустические волны вообще не обнаруже-
Пузырьки пара возникают из-за локального
ны. У родамина 6G квантовый выход φТ интер-
разогрева среды при безызлучательной релакса-
комбинационной конверсии S1 → T1 в триплетное
ции возбужденных состояний молекул красите-
состояние φТ ~ 0.01, а квантовый выход флуорес-
лей. Следует подчеркнуть, что при использован-
ценции перехода S1 → S0 φфл ~ 0.99 [19], поэтому
ных нами плотностях мощности возбуждения су-
при релаксации S1-состояний молекул раствор не
щественный (а в случае родамина 6G - основной)
нагревается. Только когда плотность мощности
вклад в разогрев растворителя вносит релаксация
накачки P достигает примерно 5 MВт/см2 и про-
ВВЭС. Заселение ВВЭС молекул в наших экспе-
цесс поглощения становится двухквантовым
риментах подтверждается зависимостью интен-
hν
hν
сивности фосфоресценции от плотности мощно-
(S
→
S
→
S
), эффективно заселяются высокие
0
1
n
сти накачки, измеренная у эозина (рис. 3).
Sn-состояния молекул, при релаксации которых
На начальном участке интенсивность свече-
возникает локальный нагрев, образуются пузырь-
ния очень быстро нарастает, достигая насыщения
ки пара и появляется ударная волна.
примерно при 5 MВт/см2, что связано с выходом
На рис. 4 показана эволюция сигнала от ударной
на насыщение заселенности S1-уровней молекул
волны при увеличении плотности мощности накач-
и, как следствие, заселенности нижних триплет-
ки родамина 6G в физиологическом растворе. Вид-
ных T1-состояний, образующихся в результате
но, что амплитуда провала на кривой пропускания
интеркомбинационной конверсии S1 → T1.
становится заметной при P > 5.7 MВт/см2.
При дальнейшем увеличении плотности мощ-
ности накачки интенсивность фосфоресценции
Как видно, если P превышает 10 MВт/см2, аку-
эозина вновь начинает заметно расти. Это объяс-
стические волны эффективно формируются и
няется тем, что при мощном возбуждении моле-
уверенно регистрируются как в растворах с ксан-
кулы красителей, находящиеся в S1-состоянии,
теновыми красителями, так и в растворах с рода-
hν
hν
мином 6G. Заметим, что в отсутствие термосен-
поглощают второй квант и по схеме
S
→
S
→
S
сибилизатора в растворе даже при плотности
0
1
n
переходят на более высокие Sn-уровни, эффек-
мощности возбуждающего света
40 MВт/см2
тивность интеркомбинационной конверсии
ударные волны не обнаружены.
БИОФИЗИКА том 65
№ 4
2020
ФОТОТЕРМИЧЕСКАЯ ИНАКТИВАЦИЯ МИКРООРГАНИЗМОВ
709
I, отн. ед.
ветствующим
увеличением
количества
возбуждающих импульсов (времени облучения).
1
0.6
Непосредственно в зоне возбуждения возмож-
но повреждение бактерий в результате фотодина-
2
мического действия; гибель клеток за счет гипер-
0.5
термии при безызлучательной релаксации ВВЭС-
3
сенсибилизаторов, связанных со стенками бакте-
4
рий или находящихся внутри клеток; разрывы хи-
0.4
мических связей биологически важных макромо-
5
лекул при безызлучательном переносе энергии с
6
ВВЭС-красителей, а также разрушение клеток
0.3
ударными волнами. На расстояниях же более
5 мм от зоны возбуждения уже не проявляются
0.2
тепловые эффекты, нет фотодинамического дей-
4.5
5.0
5.5
6.0
6.5
6.0
ствия и переноса энергии. Если имеет место по-
Время, мкс
вреждение бактерий, то оно происходит исклю-
чительно под действием ударных волн, возника-
Рис.
4. Эволюция сигнала от ударной волны в
ющих при быстром образовании и схлопывании
растворе родамина 6G (С = 0.25 мМ) с ростом
пузырьков пара в зоне облучения.
плотности мощности накачки: 1 - 5.7 MВт/см2, 2 -
11.1 MВт/см2, 3 - 18.7 MВт/см2, 4 - 27.0 MВт/см2, 5 -
На рис. 5 показаны зависимости выживаемо-
35.1 MВт/см2, 6 - 41.6 MВт/см2.
сти клеток E. coli и B. subtilis в физиологических
растворах с ксантеновыми красителями
Повреждение бактерий ударными волнами. Для
(0.25 мМ) до и после их облучения импульсами
оценки повреждения бактерий ударными волна-
различной плотности мощности Р при разных
расстояниях до зоны возбуждения.
ми использовали установку, фрагмент оптиче-
ской части которой показан на рис. 1. Отбор бак-
Видно, что непосредственно в перетяжке по-
терий для определения количества колониеобра-
вреждение бактерий E. coli происходит даже при
зующих единиц до и после возбуждения
малых плотностях мощности возбуждающего из-
производили непосредственно в области возбуж-
лучения. По-видимому, это обусловлено фотоди-
дения, а также на расстояниях 5 и 10 мм от зоны
намическим действием. Для гипертермии
возбуждения. В контрольных экспериментах ис-
клеток и безызлучательного переноса энергии
следуемые растворы бактерий с красителями об-
необходимо взаимодействие красителя со стенка-
лучали импульсами с Р < 0.5 MВт/см2, а эквива-
ми бактерий или его проникновение внутрь кле-
лентность дозы облучения обеспечивалась соот-
ток. Ксантеновые красители-анионы не взаимо-
(а)
(б)
5.0
10
4.5
8
4.0
1
6
3.5
3
4
2
1
3.0
2
2
3
2.5
2.0
0
0
10
20
30
40
0
10
20
30
40
Р, МВт/см2
Р, МВт/см2
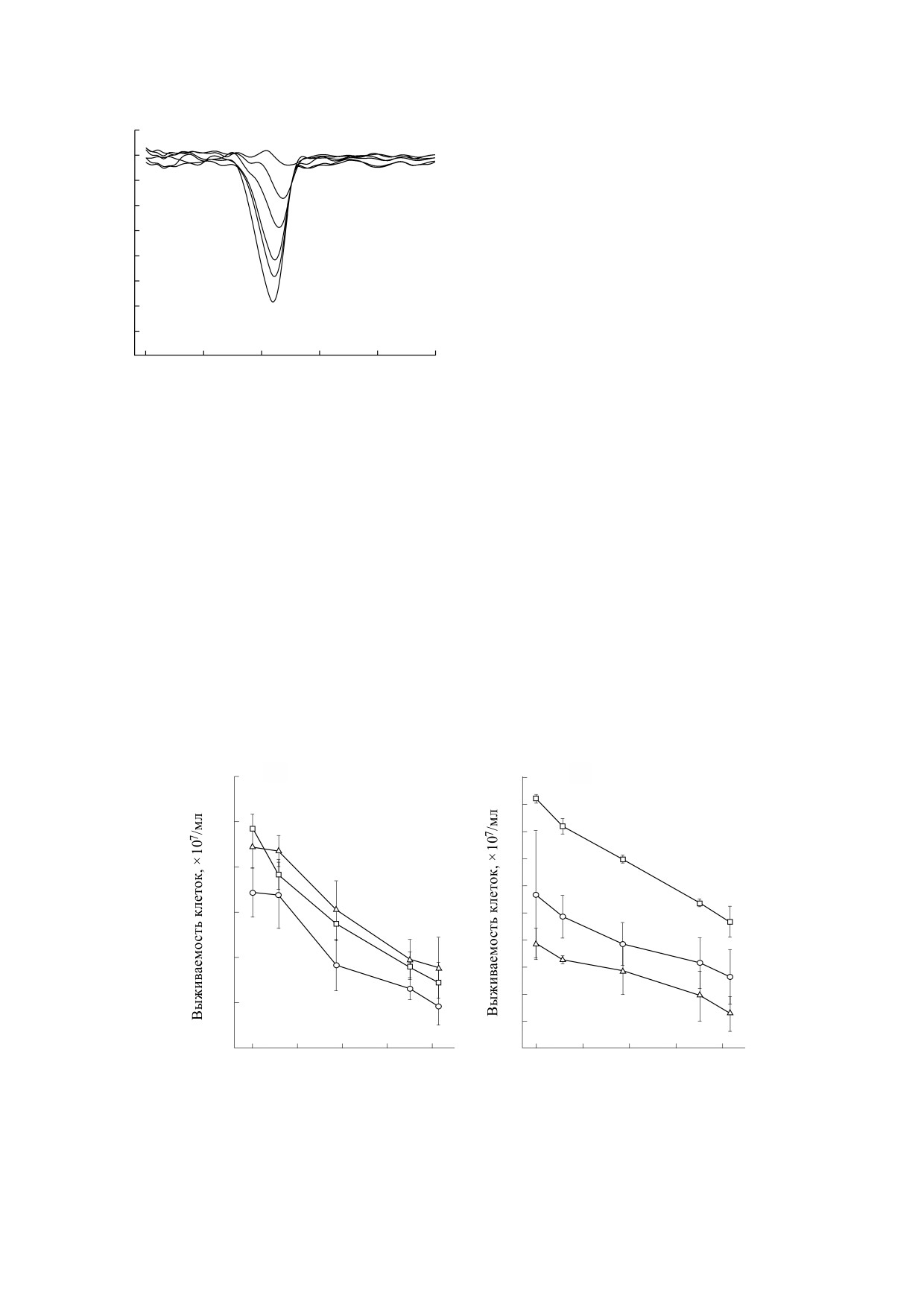
Рис. 5. Зависимости выживаемости клеток Escherichia coli в растворе с эритрозином (a) и Bacillus subtilis в растворе с
эозином (б) от плотности мощности накачки на разном расстоянии от перетяжки при постоянной концентрации
красителей (0.25 мМ) в физиологическом растворе: 1 - 0 мм, 2 - 2.5 мм, 3 - 10 мм.
БИОФИЗИКА том 65
№ 4
2020
710
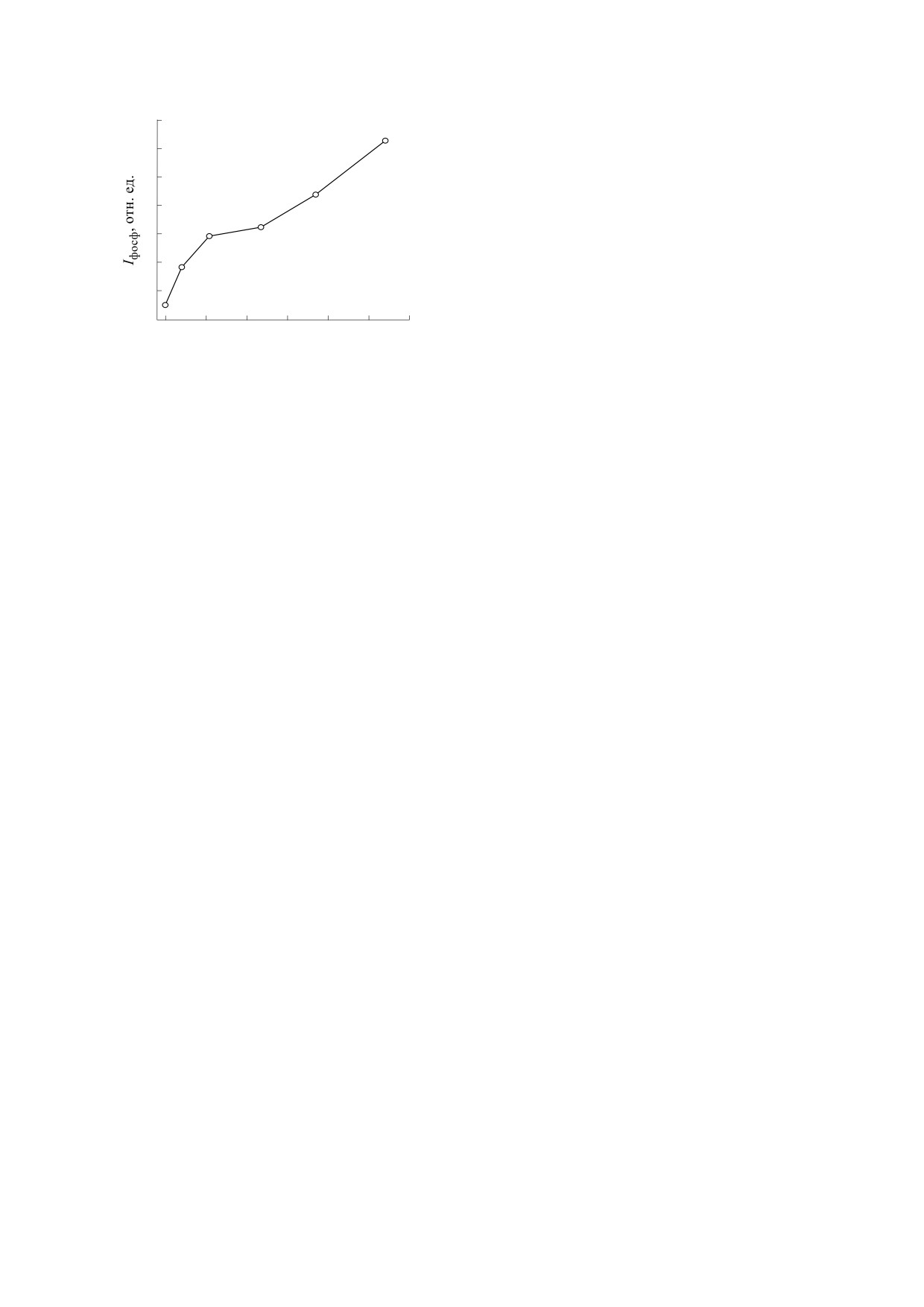
ЛЕТУТА и др.
30
центрации родамина 6G в растворе на расстоя-
нии 10 мм от зоны возбуждения представлена на
2
рис. 6. Видно, что с ростом концентрации термо-
25
сенсибилизатора эффективность повреждения
3
микроорганизмов возрастает.
20
Особенно заметными повреждения становят-
1
ся при Р > 20 MВт/см2. С ростом концентрации
красителя в растворе увеличивается плотность
15
тепловых источников. Это повышает эффектив-
ность генерации ударных волн и, соответственно,
увеличивает вероятность повреждения бактерий.
10
Кроме того, при большой концентрации красите-
лей образуются ассоциаты, квантовый выход
0
10
20
30
40
флуоресценции у которых ниже, чем у мономеров
Р, МВт/см2
[19]. Ассоциаты красителей могут быть дополни-
тельными источниками тепловыделения в рас-
Рис. 6. Зависимости выживаемости клеток Bacillus
творах.
subtilis от плотности мощности накачки на
расстоянии
10 мм от перетяжки при различной
Изображения бактерий до и после облучения в
концентрации родамина
6G в физиологическом
растворе: 1 - 0.1 мМ, 2 - 0.25 мМ, 3 - 0.5 мМ.
присутствие эритрозина, полученные с помощью
атомно-силовой микроскопии, показаны на
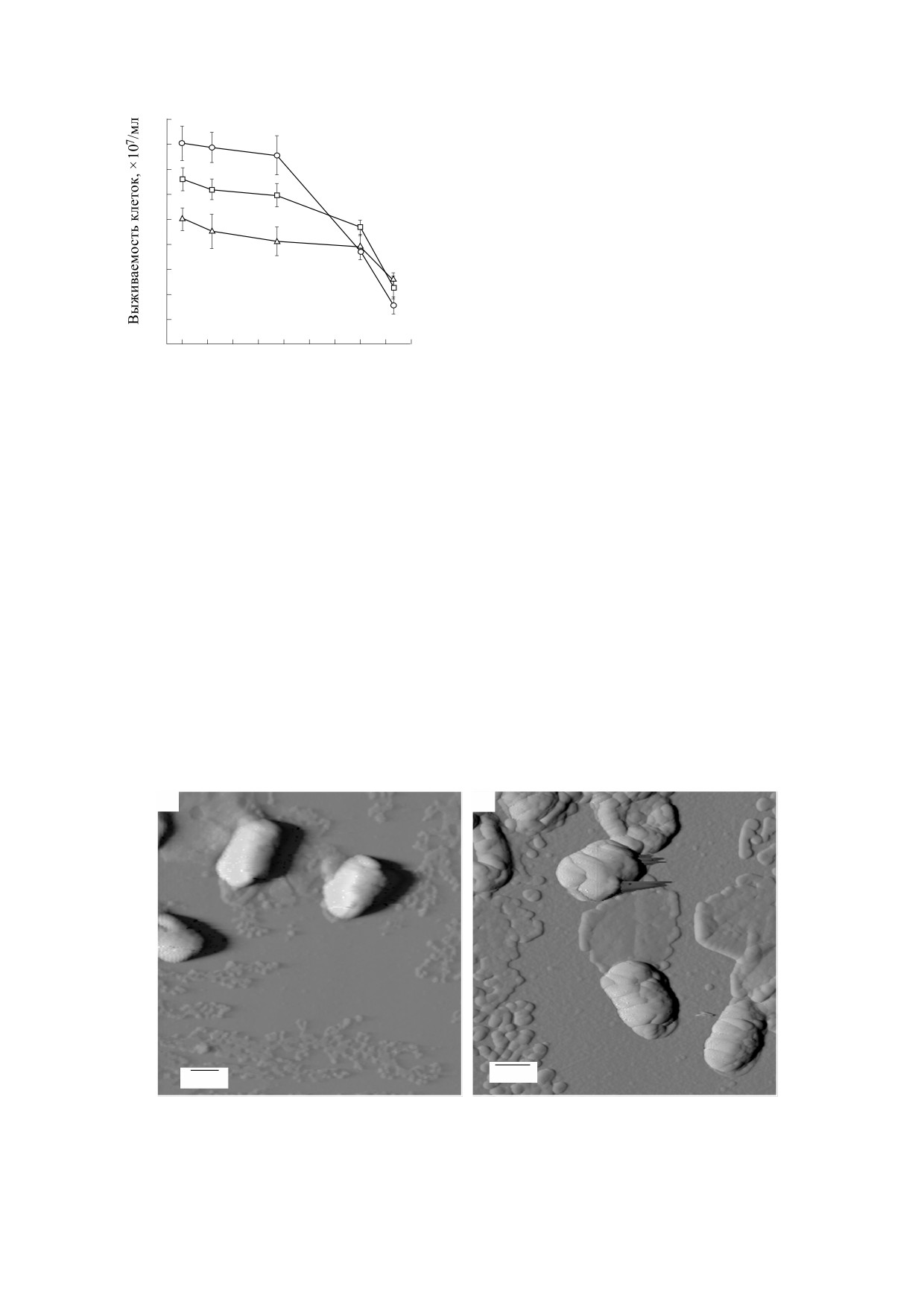
рис. 7. При исследовании образцов, содержащих
действуют с клетками E. coli, стенки которых име-
бактериальные клетки E. coli, до воздействия аку-
ют отрицательный заряд [20]. На расстояниях 5 и
стических волн на поверхности подложки обна-
10 мм от зоны возбуждения повреждения
ружены неоднородные по внешним признакам
клеток E. coli становятся заметными только при
структуры, среди которых можно выделить про-
Р > 10 MВт/см2, когда за счет двухквантового по-
долговатые объекты высотой более 600 нм и объ-
глощения возрастает эффективность генерации
екты разнообразной формы, высотой менее
ударных волн.
100 нм (рис. 7а).
Клетки B. subtilis оказались более чувствитель-
Первые объекты были идентифицированы как
ными к воздействию ударных волн. Их чувстви-
бактериальные клетки со средними размерами
тельность даже к малым дозам облучения, по-ви-
1.76 мкм в длину, 950 нм в ширину и 680 нм в вы-
димому, связана с взаимодействием эозина с бак-
соту. Более мелкие структуры, очевидно, являют-
териями.
ся остатками питательной среды и физиологиче-
Зависимость выживаемости бактерий B. subtilis
ского раствора, использующегося в качестве бу-
от плотности мощности накачки при разной кон-
фера.
(а)
(б)
500 нм
500 нм
Рис. 7. Изображения бактерий до и после облучения в присутствии эритрозина, полученные с помощью атомно-
силовой микроскопии.
БИОФИЗИКА том 65
№ 4
2020
ФОТОТЕРМИЧЕСКАЯ ИНАКТИВАЦИЯ МИКРООРГАНИЗМОВ
711
Результаты визуализации клеток E. coli после
СПИСОК ЛИТЕРАТУРЫ
облучения (λвозб = 532 нм, Р = 35 MВт/см2) в рас-
1. J. O’Neill, Review on Antimicrobial Resistance. Tackling
творе с эритрозином (0.5 мМ), отобранных на
a Global Health Crisis: Initial Steps (Wellcome Trust,
расстоянии 10 мм от зоны возбуждения, показа-
London, 2015).
ны на рис. 7б. Отличительной особенностью
представленных изображений является наличие
2. С. Д. Захаров и А. В. Иванов, Квантовая электро-
на подложке клеток с нарушенной целостностью
ника 29 (3), 192 (1999).
клеточной стенки. При этом высота клеток
3. A. A. Кpаcновcкий (мл.), Биофизика 49 (2), 305
уменьшилась и составила в среднем 580 нм.
(2004).
Для количественного описания изменений кле-
точной стенки оценена ее шероховатость до и по-
4. S. N. Letuta, S. N. Pashkevich, A. T. Ishemgulov, et al.,
сле воздействия. Среднеквадратичная шерохова-
J. Photochem. Photobiol. B: Biology 163, 232 (2016).
тость увеличилась в среднем на 40%, составляя до
5. M. Scholz and R. Dědic, in Singlet Oxygen: Applications
и после облучения в среднем 23 и 32 нм соответ-
in Biosciences and Nanosciences, Ed. by S. Nonell and
ственно.
C. Flors (Roy. Soc. of Chemistry, 2016), Vol. 2,
Аналогичные изменения наблюдаются в клет-
Chapt. 28, pp. 63-81.
ках B. subtilis: в контрольных образцах, помимо
6. A. A. Krasnovsky Jr., Biochemistry 72 (10),
1065
интактных, обнаруживаются также деформиро-
ванные клетки с разной степенью поражения
(2007).
вплоть до полностью дефрагментированных мик-
7. M. R. Hamblin, Curr. Opin. Microbiol. 33, 67 (2016).
роорганизмов.
8. J. W. Costerton, P. S. Stewart, and E. P. Greenberg,
Science 284, 1318 (1999).
ЗАКЛЮЧЕНИЕ
9. G. A. O’Toolе, H. B. Kaplan, and R. Kolter, Annu.
Ударные волны, возникающие при образова-
Rev. Microbiol. 54, 49, (2000).
нии и схлопывании пузырьков пара в физиологи-
10. А. П. Ведута, М. Д. Галанин, Б. П. Кирсанов и
ческом растворе, эффективно повреждают бакте-
З. А. Чижикова, Письма в ЖЭТФ 11 (1), 157 (1970).
рии на расстояниях, многократно превышаюших
радиус фотодинамического действия. Эффектив-
11. H.-B. Lin and M. R. Topp, Chem. Phys. Lett. 47 (3),
ность повреждения микроорганизмов возрастает
442 (1977).
с увеличением концентрации красителей в рас-
12. В. Л. Ермолаев и В. А. Любимцев, Оптика и спек-
творах и, естественно, зависит от плотности мощ-
троскопия 60 (1), 74 (1986).
ности возбуждающего света. Это явление можно
использовать для разработки практического спо-
13. H. Fukumura, K. Kikuchi, K. Koike, and H. Kokubun,
соба борьбы с патогенными микроорганизмами в
J. Photochem. Photobiol. 42 (2-3), 283 (1988).
глубоких слоях тканей. Такой способ воздействия
14. С. Н. Летута, У. Г. Летута и С. Н. Пашкевич, Био-
на микроорганизмы перспективен в средах, где
физика 64 (4), 726 (2019).
малоэффективны традиционные способы - ан-
тибиотикотерапия или фотодинамическое воз-
15. В. В. Рыльков и Е. А. Чешев, Докл. АН СССР 281
действие на патогены.
(3), 648 (1985).
16. В. В. Рыльков и Е. А. Чешев, Оптика и спектроско-
пия 63 (5), 1030 (1987).
ФИНАНСИРОВАНИЕ РАБОТЫ
17. С. Н. Летута, Вестник ОГУ 5, 88 (2002).
Работа выполнена при финансовой поддержке
Министерства образования и науки Российской
18. K. K. Rohatgi-Mukherjee and A. K. Mukhopadhyay,
Федерации, проект FSGU-2020-0003.
Indian J. Pure Appl. Phys. 14 (6), 481 (1976).
19. Л. В. Левшин и А. М. Салецкий, Люминесценция и
КОНФЛИКТ ИНТЕРЕСОВ
ее измерения (Изд-во МГУ, М., 1989).
Авторы заявляют об отсутствии конфликта
20. N. Kashef, Y.-Y. Huang, M. R. Hamblin, Nanopho-
интересов.
tonics 6 (5), 853 (2017).
21. А. А. Александров, В. А. Белогольский, В. И. Лев-
цов и др., ГСССД 190-2000. Таблицы стандартных
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
справочных данных. Вода. Скорость звука при тем-
Настоящая работа не содержит описания ка-
пературах 0…100 градусов Цельсия и давлениях
ких-либо исследований с использованием людей
0,101325…100 МПа, (Издательство стандартов, М.,
и животных в качестве объектов.
2000).
БИОФИЗИКА том 65
№ 4
2020
712
ЛЕТУТА и др.
Photothermic Inactivation of Microorganisms at Relaxation
of Highly Excited States of Sensitizers
S.N. Letuta, S.N. Pashkevich, A.T. Ishemgulov, and A.N. Nikiyan
Orenburg State University, prosp. Pobedy 13, Orenburg, 460018 Russia
This study demonstrates the susceptibility of planktonic bacteria Escherichia coli and Bacillus subtilis to inac-
tivation by shock acoustic waves arising from the rapid formation and collapse of vapor bubbles in the medium
locally heated to boiling point. Local heating of the medium occurred due to heat release through relaxation
of highly excited electronic states of exogenous molecules of organic dyes. Dyes molecules were excited by
nanosecond laser pulses. Highly excited electronic states were formed as a result of stepwise absorption of two
quanta of laser radiation. The dependency of the efficiency of microorganism inactivation on dye concentra-
tion, the excitation power density and the distance from the shock wave source was studied.
Keywords: exogenous thermosensitizers, shock acoustic waves, inactivation of microorganisms
БИОФИЗИКА том 65
№ 4
2020