БИОФИЗИКА, 2020, том 65, № 5, с. 920-923
БИОФИЗИКА КЛЕТКИ
УДК 576.32/.36
ЛИГАНДЫ РЕЦЕПТОРОВ СИГМА-1 - ХЛОРПРОМАЗИН
И ТРИФЛУОПЕРАЗИН - ИНГИБИРУЮТ ТРАНСПОРТ Na+
В ЭПИТЕЛИИ КОЖИ ЛЯГУШКИ
© 2020 г. А.В. Мельницкая*, З.И. Крутецкая*, В.Г. Антонов**, Н.И. Крутецкая*
*Санкт-Петербургский государственный университет, 199034, Санкт-Петербург, Университетская наб., 7/9
**Военно-медицинская академия им. С.М. Кирова, 194044, Санкт-Петербург, ул. Академика Лебедева, 6
E-mail: z.krutetskaya@spbu.ru, avmelnitskaya@yandex.ru
Поступила в редакцию 30.11.2019 г.
После доработки 30.11.2019 г.
Принята к публикации 04.06.2020 г.
С использованием метода фиксации потенциала впервые показано, что антагонисты рецепторов
сигма-1 - нейролептики хлорпромазин и трифлуоперазин - подавляют транспорт Na+ в эпителии
кожи лягушки. Полученные данные свидетельствуют о возможном участии рецепторов сигма-1 в
регуляции трансэпителиального транспорта Na+ в коже лягушки.
Ключевые слова: рецепторы сигма-1, транспорт Na+, хлорпромазин, трифлуоперазин.
DOI: 10.31857/S0006302920050105
Кожа амфибий и другие изолированные эпи-
ляемые Na+-проводящие каналы, блокируемые
телиальные системы являются классическими
диуретиком амилоридом. Deg/ENaC универсаль-
модельными объектами для исследования меха-
ны для всех многоклеточных организмов; экс-
низмов транспорта ионов через биологические
прессируются в различных возбудимых и невоз-
мембраны. По способности к транспорту элек-
будимых тканях; участвуют в процессах болевой
тролитов и реакции на некоторые гормоны кожа
чувствительности, механочувствительности и на-
и мочевой пузырь амфибий сходны с дистальны-
правленного переноса Na+ [2]. Несмотря на то
ми отделами почечных канальцев, что позволяет
что представители суперсемейства Deg/ENaC
использовать данные, получаемые на этих объек-
функционально гетерогенны, они обладают сход-
тах, для выяснения механизмов транспорта воды
ными биофизическими свойствами и структур-
и ионов в клетках почки [1]. Транспорт Na+ в эпи-
ной организацией.
телиальных тканях представляет собой сложную,
Рецепторы сигма-1 представляют собой уни-
многокомпонентную систему, в работе которой
кальные лигандрегулируемые молекулярные ша-
принимают участие Na+-транспортирующие бел-
пероны, локализованные в плазматической мем-
ки и сигнальные каскады, локализованные в раз-
бране и в мембране эндоплазматического ретику-
личных мембранах клетки. Белковые компонен-
лума на границе с митохондриями. Эти рецеп-
ты этой системы являются мишенью для дей-
торы широко экспрессированы в центральной
ствия широкого спектра гормонов и фар-
нервной системе и в периферических тканях, в том
макологических агентов.
числе в клетках почки и печени [3, 4]. Их лиганда-
ми являются эндогенные стероиды, антидепрес-
Ключевую роль в транспорте Na+ в реабсорби-
санты, антипсихотические, противосудорожные
рующих эпителиях играют амилорид-чувстви-
и анальгетические средства [5, 6]. Рецепторы сиг-
тельные эпителиальные Na+-каналы (ENaC).
ма-1 взаимодействуют с многочислен-ными бел-
ENaC являются представителями обширного су-
ками-мишенями, включая ионные каналы и ре-
персемейства дегенерин/эпителиальные Na+-ка-
цепторы, а также участвуют в модуляции многих
налы (Deg/ENaC), объединяющего лигандуправ-
клеточных процессов [7]. Обнаружено, что в ней-
ронах рецепторы сигма-1 модулируют активность
Сокращения: ENaC - амилорид-чувствительные эпители- потенциалзависимых ионных каналов различных
альные Na+-каналы, Deg/ENaC - суперсемейство дегене-
типов [8]. В последнее время появляются данные о
рин/эпителиальные Na+-каналы, ASICs - протон-активи-
руемые ионные каналы, ХП - хлорпромазин, ТФП - три-
том, что рецепторы данного типа участвуют в мо-
флуоперазин.
дуляции активности протон-активируемых ион-
920
ЛИГАНДЫ РЕЦЕПТОРОВ СИГМА-1 - ХЛОРПРОМАЗИН
921
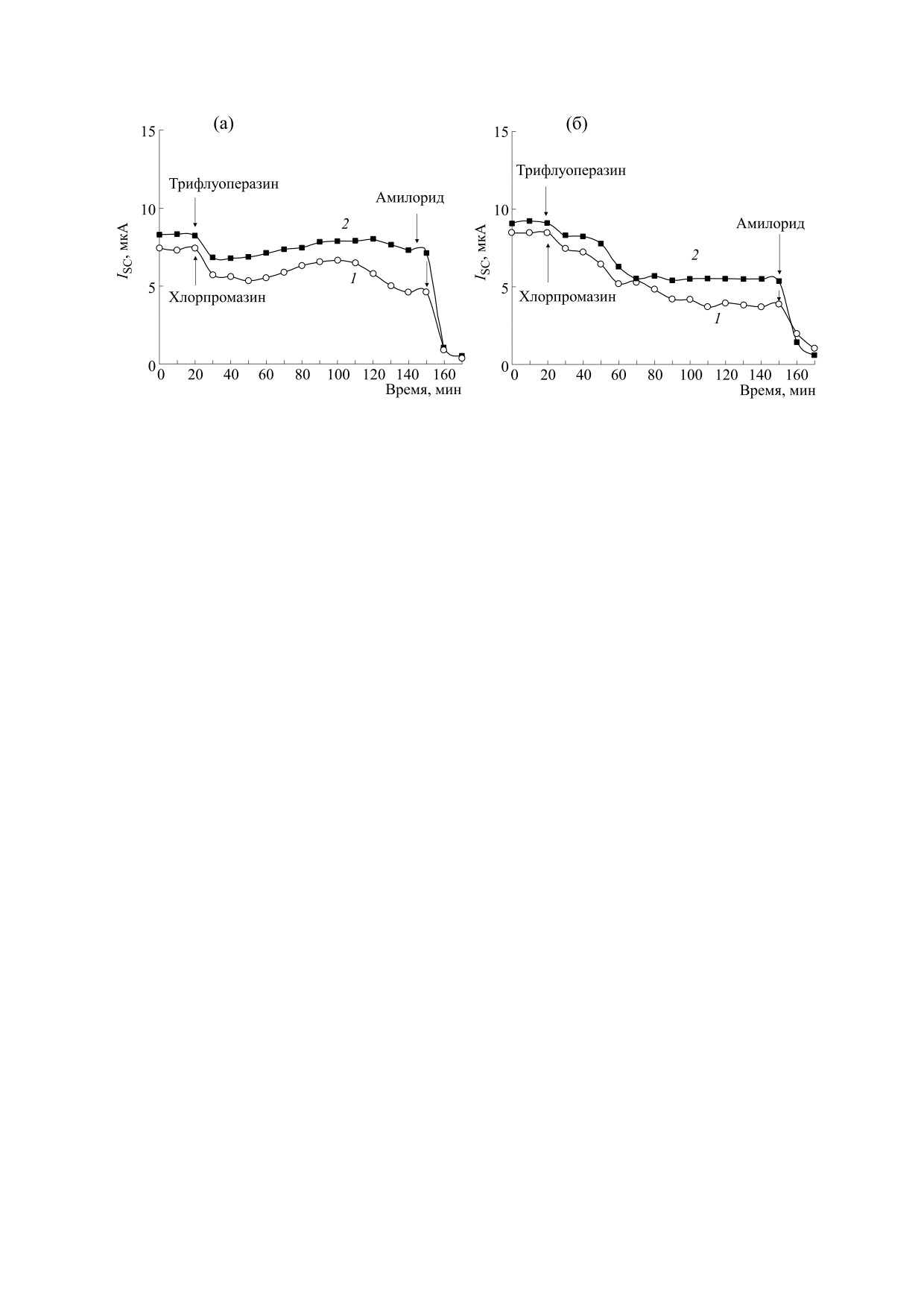
Кинетика изменения тока короткого замыкания ISC через кожу лягушки в ответ на приложение антагонистов
рецепторов сигма-1 - 50 мкг/мл хлорпромазина (1) и 20 мкг/мл трифлуоперазина (2), добавленных со стороны
апикальной (а) и базолатеральной (б) поверхности кожи лягушки. В конце каждого эксперимента в раствор, омы-
вающий апикальную поверхность кожи, добавляли блокатор амилоридчувствительных эпителиальных Na+-ка-
налов - амилорид (20 мкМ).
ных каналов (ASICs) - одного из представителей
держивали при 0 мВ (режим короткого замыка-
суперсемейства Deg/ENaC [9]. В то же время роль
ния) или при потенциале открытой цепи VOC
рецепторов сигма-1 в процессах трансэпителиаль-
(VOC = VT при трансэпителиальном токе IT = 0).
ного транспорта Na+ практически не изучена. В
Из вольт-амперных характеристик определяли
связи с этим представлялось целесообразным ис-
электрические параметры кожи: ток короткого
следовать участие рецепторов сигма-1 в регуляции
замыкания ISC (ISC = IT при VT = 0), VOC и транс-
транспорта Na+ в эпителии кожи лягушки. В экс-
эпителиальную проводимость gT.
периментах были использованы антагонисты ре-
Транспорт Na+ оценивали как амилоридчув-
цепторов сигма-1 - типичные антипсихотические
ствительный ток ISC. В связи с этим в конце каж-
средства - нейролептики фенотиазинового ряда
хлорпромазин (аминазин, ХП) и трифлуоперазин
дого эксперимента в раствор, омывающий апи-
(трифтазин, ТФП) [5, 6].
кальную поверхность кожи, добавляли блокатор
ENaC амилорид (20 мкМ).
Использовали реактивы фирмы Sigma (США).
МАТЕРИАЛЫ И МЕТОДЫ
Маточные растворы ХП
(25 мг/мл), ТФП
(5 мг/мл) и амилорида (10 мМ) готовили на воде.
Эксперименты проводили на самцах лягушки
Фармакологические агенты добавляли к апикаль-
Rana temporaria в период с ноября по март. Кожу с
ной или базолатеральной поверхности кожи ля-
брюшка лягушки срезали и помещали в камеру
гушки.
Уссинга (World Precision Instruments Inc., Герма-
Статистический анализ проводили с примене-
ния) с диаметром внутреннего отверстия 12 мм.
нием t-критерия Стьюдента. Данные представле-
Камеру заполняли раствором Рингера для холод-
ны в виде x ± sx. На рисунке приведены результа-
нокровных, содержащим (в мМ): NaCl - 110,
KCl - 2.5, CaCl2 - 3, Tris-HCl -5, pH 7.4. Опыты
ты типичных экспериментов.
проводили при комнатной температуре
(22-
23°С).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Для измерения электрических параметров ко-
Значения электрических характеристик кожи
жи лягушки использовали автоматизированную
лягушки в контроле в среднем (по данным десяти
установку фиксации потенциала и регистрации
экспериментов) составили: ISC = 9.5 ± 2.34 мкA,
вольт-амперных характеристик [10]. Для измере-
VOC = - 11.71 ± 3.85 мВ, gT = 0.79 ± 0.27 мСм.
ния вольт-амперных характеристик на кожу по-
Мы обнаружили, что обработка кожи лягушки
давали линейно изменяющееся напряжение со
скоростью изменения 20 мВ/с. В интервалах меж-
ТФП или ХП снижает в ней транспорт Na+. В
ду измерениями вольт-амперных характеристик
среднем, изменение электрических характери-
трансэпителиальный потенциал (VT) кожи под-
стик кожи лягушки (здесь и далее по тексту x ± sx,
БИОФИЗИКА том 65
№ 5
2020
922
МЕЛЬНИЦКАЯ и др.
n (число опытов)
= 10) после добавления
В цитируемых работах также высказывалось
20 мкг/мл ТФП было следующим: ISC уменьшил-
предположение о том, что молекулярные меха-
ся на 10.34 ± 3.82 или 31.24 ± 13.11%, VOC умень-
низмы, вовлеченные в регуляцию ТФП транс-
шился на 26.35 ± 12.24 или 34.18 ± 16.22%, а gT
порта Na+ в коже лягушки, различаются в зависи-
уменьшилась на 21.12 ± 8.74 или увеличилась на
мости от приложения агента к апикальному или
базолатеральному домену полярных клеток эпи-
5.21 ± 1.14% при приложении ТФП со стороны
апикальной или базолатеральной поверхности
телия. Так, наиболее вероятно, что влияние ТФП
кожи соответственно. В случае обработки кожи
на ISC в случае приложения агента к апикальной
лягушки 50 мкг/мл ХП изменение электрических
поверхности кожи связано с модуляцией актив-
характеристик в среднем было следующим:
ности ENaC [16] или с изменением внутрикле-
ISC уменьшился на 18.34 ± 6.19 или 39.25 ± 7.18%,
точной концентрации ионов Са2+ [15], тогда как
VOC уменьшился на 20.02 ± 7.35 или 40.18 ±
влияние ТФП на ISC при приложении агента к ба-
± 12.34% , а gT не изменялась или уменьшилась на
золатеральной поверхности кожи опосредуется
10.34 ± 3.85 % при приложении ХП со стороны
Са2+-зависимым синтезом и последующим выде-
апикальной или базолатеральной поверхности
лением из базолатеральной мембраны проста-
кожи соответственно.
гландина Е2, что в свою очередь приводит к сти-
Полученные данные свидетельствуют о том,
муляции транспорта Na+ через кожу лягушки [15,
что степень и кинетика ингибирующего действия
16]. Известно, что ТФП является антагонистом
нейролептиков фенотиазинового ряда на транс-
Са2+-связывающего белка кальмодулина, играю-
порт Na+ различаются в зависимости от приложе-
щего ключевую роль в регуляции процессов
ния агентов со стороны апикальной или базола-
Са2+-сигнализации [17]. Однако имеются данные
теральной поверхности кожи (см. рисунок). Так,
о сходном влиянии на ISC различных производ-
приложение ХП или ТФП к апикальной поверх-
ности кожи вызывает двухфазное изменение ISC:
ных фенотиазина (ХП и ТФП), что свидетель-
подавление ISC, наблюдаемое в течение 30-
ствует о том, что влияние ТФП на транспорт Na+
40 мин после приложения агентов, сменяющееся
в коже лягушки осуществляется, по-видимому,
некоторым увеличением ISC, наблюдаемым в те-
без участия комплекса «Са2+-кальмодулин» [16].
чение второго часа после приложения ТФП или
ХП. В случае добавления ТФП или ХП со сторо-
Известно, что транспорт Na+ в эпителиях
ны базолатеральной поверхности кожи наблюда-
представляет собой сложную многокомпонент-
ется постепенное снижение ISC в течение первого
ную систему, в работе которой принимают уча-
часа после приложения агентов. Фаза увеличения
стие многочисленные Na+-транспортирующие
ISC в этом случае не наблюдалась. Можно пред-
белки, локализованные в различных мембранах
положить, что влияние антагонистов рецепторов
клетки. Введение в конце каждого эксперимента
в раствор, омывающий апикальную поверхность
сигма-1 на транспорт Na+ в коже лягушки осу-
кожи лягушки, блокатора ENaC амилорида
ществляется при участии различных белковых и
(20 мкМ) вызывало полное подавление ISC (см.
липидных сигнальных комплексов, ассоцииро-
рисунок), что свидетельствует о том, что влияние
ванных с апикальным и базолатеральным доме-
нами поляризованных эпителиальных клеток.
антагонистов рецепторов сигма-1 ХП или ТФП
на транспорт Na+ связано преимущественно с
Полученные нами результаты согласуются с
модуляцией активности ENaC. Полученные ре-
литературными данными. Так, способность ХП
зультаты могут представлять интерес в связи с ли-
ингибировать активность Na+-K+-ATФазы [11] и
тературными данными о способности рецепторов
снижать внутриклеточную концентрацию Na+
сигма-1 модулировать активность ионных кана-
обнаружена в клетках печени крысы (Rattus nor-
лов ASICs - одного из представителей суперсе-
wegicus) и жабы (Bufo marinus) [12]. Было показа-
мейства Deg/ENaC, к которому принадлежат и
но, что ХП ингибирует активность ENaC, экс-
ENaC, играющие кючевую роль в транспорте Na+
прессированных в ооцитах лягушки Xenopus [13].
в реабсорбирующих эпителиях. Так, на клетках
Обнаружено также, что приложение 100 мкМ
эмбриональной почки человека (линия НЕК-
ТФП со стороны базолатеральной мембраны, вы-
293) обнаружено, что возможно как прямое взаи-
зывает снижение ISC и VOC в мочевом пузыре жа-
модействие между рецепторами сигма-1 и ASICs,
бы [14]. Двухфазное дозозависимое изменение
с образованием комплекса со стехиометрией 1 ре-
ISC при воздействии ТФП показано для изолиро-
цептор сигма-1 : 1 субъединица ASIC [9], так и
ванной кожи лягушки Rana esculenta [15, 16]. Од-
опосредованное влияние агонистов/антагони-
нако в этом случае добавление ТФП вызывало
стов рецепторов сигма-1 на ASICs при участии
стимуляцию транспорта Na+ через кожу лягушки.
дополнительных сигнальных молекул, таких как
БИОФИЗИКА том 65
№ 5
2020
ЛИГАНДЫ РЕЦЕПТОРОВ СИГМА-1 - ХЛОРПРОМАЗИН
923
гетеротримерные G-белки и комплекс кальци-
СПИСОК ЛИТЕРАТУРЫ
нейрина с адаптерным белком AKAP150 [18].
1. Ю. В. Наточин, Основы физиологии почки (Наука,
Таким образом, как ранее [19], так и в настоя-
Л., 1982).
щей работе нами показано, что различные анта-
2. S. Kellenberger and L. Schild, Physiol. Rev. 82, 735
гонисты рецепторов сигма-1, в том числе нейро-
(2002).
лептики фенотиазинового ряда ХП и ТФП, моду-
3. C. G. Rousseaux and S. F. Greene, J. Recept. Signal.
лируют транспорт Na+ в коже лягушки, что
Trans. 36, 327 (2016).
свидетельствует об участии рецепторов сигма-1 в
4. S. B. Hellewell, A. Bruce, G. Feinstein, et al., Eur. J.
Pharmacol. 268, 9 (1994).
регуляции транспорта Na+ в эпителии кожи ля-
гушки. Полученные нами данные о влиянии ан-
5. E. J. Cobos, J. M. Entrena, F. R. Nieto, et al., Curr.
тагонистов рецепторов сигма-1 на трансэпители-
Neuropharmacol. 6, 344 (2008).
6. Y. Itzhak, M. Ruhland, and H. Krahling, Neurophar-
альный транспорт Na+ способствуют более де-
macol. 29, 181 (1990).
тальному пониманию молекулярных механизмов
7. B. Penke, L. Fulop, M. Szucs, et al., Curr. Neurophar-
фармакологического действия производных фе-
macol. 16, 97 (2018).
нотиазина, широко применяемых в клинической
практике в качестве антипсихотических, миоре-
8. T.-P. Su, T. Hayashi, T. Maurice, et al., Trends Phar-
macol. Sci. 31, 557 (2010).
лаксирующих и седативных средств, а также для
лечения шизофрении и других психических забо-
9. S. M. Carnally, M. Johannessen, R. M. Henderson,
et al., Biophys. J. 98, 1182 (2010).
леваний.
10. Z. I. Krutetskaya, O. E. Lebedev, A. V. Melnitskaya,
et al., Dokl. Akad. Nauk 421 (5), 709 (2008).
ФИНАНСИРОВАНИЕ РАБОТЫ
11. R. W. Van Dyke and B. F. Scharschmidt, Am. J.
Работа выполнена в рамках плановых тем ка-
Physiol. 253, 613 (1987).
федры биофизики Санкт-Петербургского государ-
12. P. Else and K. Mansfield, Biochem. Pharmacol. 54,
ственного университета и кафедры клинической
275 (1997).
биохимии и лабораторной диагностики Военно-
13. M. S. Awayda, W. Shao, F. Guo, et al., J. Gen. Physiol.
медицинской академии им. С.М. Кирова (Санкт-
123, 709 (2004).
Петербург), а также Договора на выполнение на-
14. S. D. Levine, W. A. Kachadorian, D. N. Levin, et al.,
учно-исследовательских работ № 28-12-38.
J. Clin. Invest. 67 (3), 662 (1981).
15. H. F. Bjerregaard and R. Nielsen, Acta Physiol. Scand.
127 (1), 75 (1986).
КОНФЛИКТ ИНТЕРЕСОВ
16. H. F. Bjerregaard and R. Nielsen, Acta Physiol. Scand.
Авторы заявляют об отсутствии конфликта
134 (1), 43 (1988).
интересов.
17. M. D. Feldkamp, S. E. O'Donnell, L. Yu, et al., Pro-
teins 78 (10), 2265 (2010).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
18. Y. Herrera, C. Katnik, J. D. Rodriguez, et al., J. Phar-
macol. Exp. Ther. 327, 491 (2008).
Все применимые международные, националь-
19. А. В. Мельницкая, З. И. Крутецкая, С. Н. Бутов
ные и институциональные принципы ухода и ис-
и др., в кн. Рецепторы и внутриклеточная сигнали-
пользования животных при выполнении работы
зация, под ред. В. П. Зинченко и А. В. Бережнова
были соблюдены.
(Fix-Print, Пущино, 2017), т. 1, сс. 55-58.
Sigma-1 Receptor Ligands Chlorpromazine and Trifluoperazine Inhibit Na+ Transport
in Frog Skin Epithelium
A.V. Melnitskaya*, Z.I. Krutetskaya*, V.G. Antonov**, and N.I. Krutetskaya*
*Saint Petersburg State University, Universitetskaya nab. 7/9, St. Petersburg, 199034 Russia
**Kirov Military Medical Academy, ul. Akademika Lebedeva 6, St. Petersburg, 194044 Russia
Using voltage-clamp technique, we have shown for the first time that sigma-1 receptor antagonists - neuro-
leptics chlorpromazine and trifluoperazine - attenuate Na+ transport in the frog skin epithelium. The results
suggest the possible involvement of sigma-1 receptors in the regulation of transepithelial Na+ transport in frog
skin.
Keywords: sigma-1 receptors, Na+ transport, chlorpromazine, trifluoperazine
БИОФИЗИКА том 65
№ 5
2020