БИОФИЗИКА, 2020, том 65, № 5, с. 932-948
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 577.3
МОДЕЛЬ ПРЕДПОСЫЛОК ВИДООБРАЗОВАНИЯ В ПРЕДСТАВЛЕНИЯХ
ТЕОРИЙ ПЕРКОЛЯЦИЙ И САМООРГАНИЗОВАННОЙ КРИТИЧНОСТИ
© 2020 г. А.Я. Гараева, А.Э. Сидорова, В.А. Твердислов, Н.Т. Левашова
Физический факультет Московского государственного университета имени М.В. Ломоносова,
119991, Москва, Ленинские горы, 1/2
E-mail: sky314bone@mail.ru
Поступила в редакцию 15.02.2020 г.
После доработки 13.05.2020 г.
Принята к публикации 20.05.2020 г.
Рассмотрены модели фиксации мутаций в ходе видообразования для бесполого и полового размно-
жения под управлением случайных процессов, а также модель перколяционной решетки отбора му-
таций в поколениях и триггерная модель фиксации мутаций в популяционном кластере, основан-
ные на случайных и детерминированных процессах. Показано, что для процесса фиксации мутаций
через перколяционную решетку отбора характерно наличие порогов протекания - нижнего (обра-
тимые процессы) и верхнего (необратимые процессы), где верхний порог - это эволюционный шаг
видообразования, соответствующий самоорганизованной критичности. В результате кооператив-
ного эффекта взаимодействия особей-мутантов формируется качественно новое устойчивое состо-
яние (в зависимости от знака мутаций): популяция с новыми признаками как шаг видообразования
или же вымирание популяции.
Ключевые слова: самоорганизованная критичность, флуктуации, бифуркации, перколяции, запрещаю-
щие и разрешающие мутации.
DOI: 10.31857/S0006302920050129
Теория самоорганизованной критичности
кооперативного когерентного взаимодействия
П. Бака [1, 2], возникшая на основе теории кон-
носителей мутаций происходит цепная реакция
денсата [3, 4], получила развитие в различных об-
видообразования - флуктуационно-бифуркаци-
ластях науки: сейсмологии ([5] и др.), материало-
онная траектория биологической эволюции. Оче-
ведении ([6] и др.), астрофизике и физике плазмы
видно, что переход системы в критическое состо-
([7-9] и др.), биологической эволюции [10]. Са-
яние определяется случайными и детерминиро-
моорганизованная критичность - это закономер-
ванными процессами. Поэтому в данной работе
ность поведения сложных динамических систем,
рассмотрены модели фиксации мутаций в ходе
способных в результате кооперативного поведе-
видообразования в режиме случайных процессов
ния неустойчивых элементов системы (взаимо-
для бесполого и полового размножения, а также
действия) в окрестности точки бифуркации при
модель, включающая случайные и детерминиро-
малых флуктуациях самопроизвольно перейти в
ванные процессы, где эффект новых мутаций за-
критическое состояние, характеризуемое каче-
висит от спектра ранее закрепившихся «разреша-
ственно новой структурой, связанной с формиро-
ющих» и «запрещающих» мутаций [16], а их по-
ванием гигантской флуктуации. В биологической
следовательность определяет «вектор отбора» -
эволюции основным условием формирования
обратимость или необратимость случайных про-
«лавин видообразования» [10] является коопера-
цессов после прохождения точки бифуркации.
тивный эффект взаимодействий между особями в
популяции в результате закрепления мутаций. В
САМООРГАНИЗОВАННАЯ КРИТИЧНОСТЬ
ходе «лавины видообразования», размер которой
В МОДЕЛЯХ СЛУЧАЙНЫХ ПРОЦЕССОВ
определяется количеством успешных эволюци-
ФИКСАЦИИ МУТАЦИЙ
онных шагов [10], происходят множественные
мутации [11] в сторону более высокой приспособ-
Самоорганизованная критичность как явле-
ленности. При этом ландшафт «пригодности»
ние с необходимостью сочетает взаимодействие
[12] определяется генотипом [13] и фенотипом, а
детерминированных и случайных процессов. В
также зависит от характеристик других видов в
целом детерминированные процессы определяют
рассматриваемом биоценозе [14, 15]. В результате
направление эволюции системы, тогда как слу-
932
МОДЕЛЬ ПРЕДПОСЫЛОК ВИДООБРАЗОВАНИЯ
933
чайные процессы - фронтальную структуру ее
Для выявления закономерностей в процессе
развития. Так, суперпозиция флуктуаций в не-
фиксации мутации в модели Морана было прове-
равновесной системе может привести к формиро-
дено 10000 численных экспериментов. В резуль-
ванию гигантских флуктуаций, связанных с фор-
тате каждого эксперимента мы получаем пару
мированием качественно новой структуры. По-
симметричных относительно значения N/2 гра-
добный подход может быть распространен на
фиков динамики мутантов и не-мутантов по по-
изучение проблем и механизмов биологической
колениям в популяции. Каждый эксперимент
эволюции, в частности, видообразования. В на-
останавливается, когда популяция полностью со-
шем случае это формирование устойчивой попу-
стоит из особей одного типа - клона мутантов
ляции, способной перейти к созданию вида, а с
или не-мутантов. Для иллюстрации случайных
точки зрения самоорганизованной критичности
процессов в модели Морана на рис. 1 представле-
мы наблюдаем «лавину видообразования» [1, 2].
Нашей задачей является рассмотрение законо-
ны графики динамики численности мутантов и
мерностей формирования самоорганизованной
не-мутантов при различных вариантах соотноше-
критичности как результата случайных процес-
ния мутанты/не-мутанты в исходном поколении
сов в ходе фиксации мутаций.
по поколениям для десяти экспериментов. Оче-
Широко применяемые рандомные модели
видно, что графики динамики численности му-
позволяют рассмотреть характерные закономер-
тантов и не-мутантов по поколениям (n) для соот-
ности фиксации мутаций на основе случайных
ношений 10/90 и 90/10, а также 30/70 и 70/30
процессов. Модель Морана - простой стохасти-
симметричны относительно N/2. Графики де-
ческий процесс, широко используемый для опи-
монстрируют следующие закономерности между
сания вероятностной динамики процесса конку-
поколениями и долей мутантов в исходном поко-
ренции двух аллелей (носители способны воспро-
лении: по мере увеличения доли мутантов возрас-
изводить свои копии) в популяции конечной
тает количество клонов мутантов и уменьшается
численности (постоянного размера N) [17]. На
минимальное количество поколений, в которых
каждом временном шаге случайным образом вы-
достигается 100%-е доминирование клона.
бираются одна особь для воспроизведения себе
подобной и одна особь для элиминации, что га-
Аналогично протекают процессы для
рантирует постоянную численность популяции:
10000 экспериментов. Для постоянной числен-
если в одном клоне одна особь получает в опреде-
ности популяции (N = 100) получены значения
ленном поколении 100%-е доминирование, то в
поколений (n), в которых мутанты достигают
том же поколении в другом клоне из эксперимен-
100%-го доминирования в популяции в зависи-
та удаляется особь. В случае бесполого размноже-
мости от увеличения доли мутантов в исходном
ния
«простое рандомизирование» аналогично
поколении (Nmut0). Так, если в исходном поколе-
«подбрасыванию монет» [18].
нии соотношение «мутанты/не-мутанты» состав-
Рассмотрим процесс фиксации мутаций в со-
ляет 10/90, доминирование мутантов в популя-
ответствии с моделью Морана [17] для популяции
ции достигается к 1719-му поколению (nmin), а
постоянной численности (N =100), размножаю-
при увеличении доли мутантов/не-мутантов до
щейся бесполым способом, где особи представле-
90/10 - к 39-му поколению (рис. 2а). Медианное
ны клонами двух типов - мутанты и не-мутанты.
значение поколений (nmedian, половина экспери-
Рассмотрены следующие варианты соотношения
ментов заканчивалась доминированием мутантов
мутантов и не-мутантов в исходном поколении (в
до этого значения, половина - после), необходи-
долях от общей численности популяции): 10/90,
мое для закрепления нового признака, также па-
30/70, 50/50, 70/30, 90/10. На каждом временном
дает с увеличением Nmut0 (рис. 2б). При этом мак-
шаге количество мутантов может:
симальные количества поколений, в которых
а) остаться неизменным (если для воспроизве-
достигается 100% доминирование (nmax, макси-
дения и элиминации будут выбраны особи одина-
мальное количество поколений, за которое про-
кового типа, в этом случае количество не-мутан-
ходило закрепление мутации в экспериментах),
тов также останется неизменным);
сравнимы, что связано с большим количеством
б) уменьшиться на единицу (если для элими-
экспериментов (рис. 2в). Мы приводим мини-
нации случайным образом будет выбрана мутант-
мальное, медианное и максимальное количество
ная особь, а для размножения не-мутантная, в
поколений, чтобы показать, что распределение
этом случае количество не-мутантов увеличится
количества поколений, необходимых для закреп-
на единицу);
ления нового признака, имеет очень длинный
в) увеличиться на единицу (если для размно-
«хвост» справа (распределение ассиметрично и
жения будет выбрана мутантная особь, а для эли-
даже при высокой доле мутантов/не-мутантов
минации - не-мутантная, в этом случае количе-
(90/10), 100%-е доминирование мутантов в поло-
ство не-мутантов уменьшится на единицу).
вине из 10000 экспериментов достигается в диа-
БИОФИЗИКА том 65
№ 5
2020
934
ГАРАЕВА и др.
В случае полового размножения выбрана ран-
домизация с распределением особей по двум
группам - мутанты и не-мутанты, самцы и самки
[19], что уменьшает системную ошибку отбора.
Для двуполой популяции рассмотрен вариант мо-
дели Райта-Фишера [20], описывающей измене-
ния в частоте одиночной мутации в популяции во
времени. Модель и эксперимент основаны на
следующих условиях: поколения не пересекают-
ся, постоянный размер популяции в каждом по-
колении (N = const), популяция размера N состо-
ит из диплоидных особей (2N копий), имеющих
единственный полиморфный сайт с двумя алле-
лями - один из которых является предковым, а
другой - производным (А и а), случайное спари-
вание. В каждую единицу времени типы особей в
момент времени (n - 1) определяют типы особей
в момент времени (n).
Рассмотрим процесс фиксации мутаций при
случайном спаривании в непересекающихся по-
колениях для двуполой популяции постоянного
размера (N = 100) и постоянного состава - коли-
чество самцов (Nm) и самок (Nf) неизменно в по-
колениях. На каждом временном шаге (n - поко-
ление) каждая пара особей (самец-самка) произ-
водит пару потомков (самец-самка), далее
родители элиминируются из популяции. Переме-
шивание самцов и самок происходит случайным
образом на каждом временном шаге с использо-
ванием генератора случайных чисел (каждой сам-
ке f случайным образом равновероятно ставится в
пару самец m). Передача мутации потомству
определяется следующим образом:
а) если оба родителя не-мутанты, то оба по-
томка - не-мутанты;
б) если оба родителя мутанты, то оба потомка -
мутанты;
в) если один родитель мутант, а другой - не-
мутант, то наследование мутации для каждого по-
томка определяется в зависимости от значения
вероятности pmut.
Результаты моделирования определяются слу-
чайными процессами выборки, вероятностью
фиксации новой мутации (pmut) в следующем по-
колении и соотношением мутанты/не-мутанты в
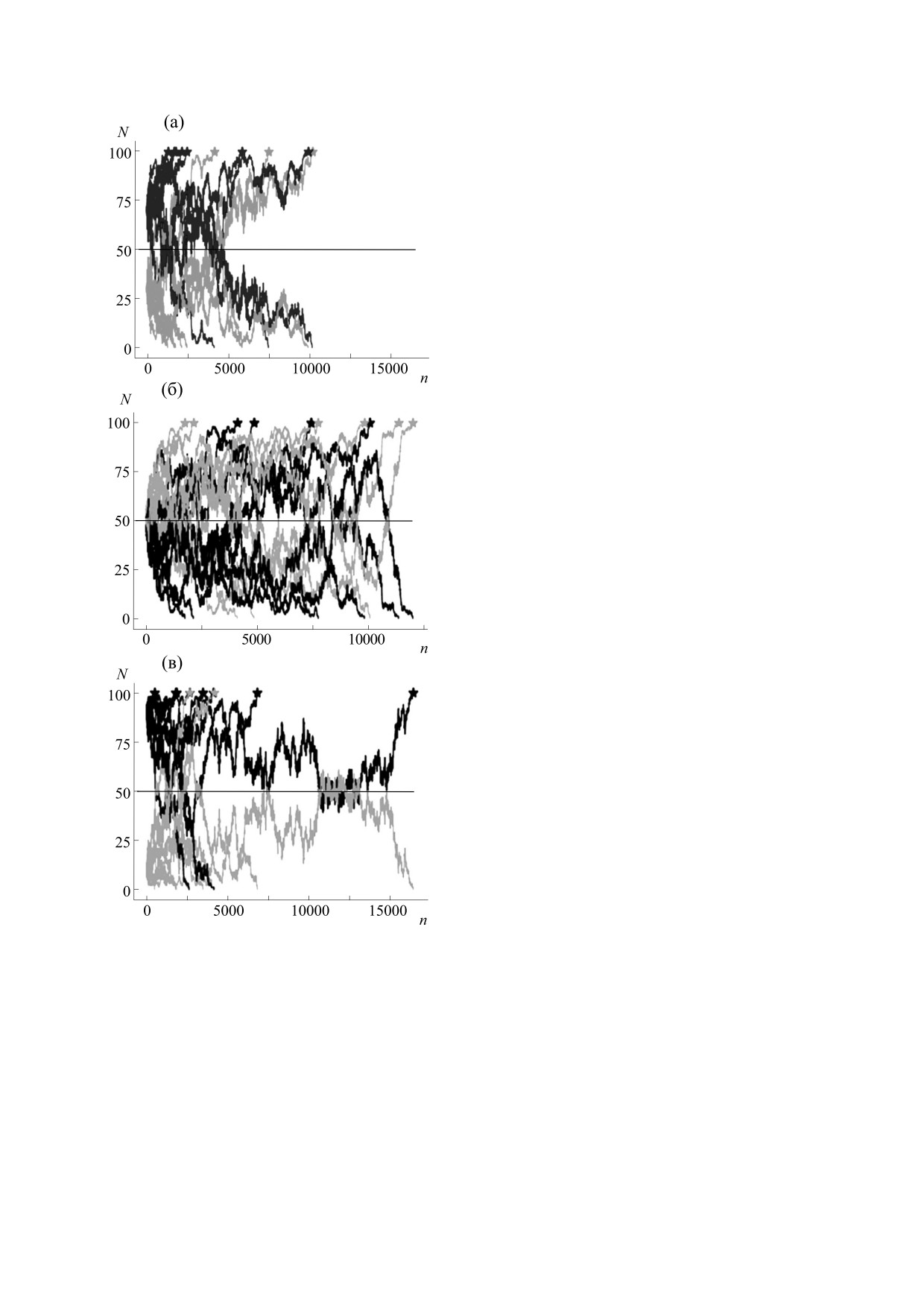
Рис. 1. Результаты численного эксперимента для бес-
исходном поколении. Для модели Райта-Фишера
полой популяции (десять экспериментов) в соответ-
произведено 10000 численных экспериментов для
ствии с моделью Морана для трех вариантов соотно-
различных значений соотношения мутанты/не-
шения «мутанты/не-мутанты»: (а) - 30/70; (б) -
50/50; (в) - 70/30. Светло-серые линии - мутанты,
мутанты в исходном поколении (10/90, 30/70,
темно-серые линии - не-мутанты. Размер популяции
50/50, 70/30, 90/10) и вероятности передачи мута-
N = 100. Звездочками на графиках отмечены поколе-
ции (pmut = 0.1, 0.3, 0.5, 0.7, 0.9). Так же как и в мо-
ния, в которых клоны достигают 100%-го доминиро-
дели Морана, графики динамики количества му-
вания. Черная горизонтальная линия: N = 50.
тантов и не-мутантов по поколениям симметрич-
ны относительно N/2.
пазоне [39÷1313] поколений, в другой половине
Для иллюстрации случайных процессов при
экспериментов - в диапазоне [1313÷44066] поко-
половом размножении на рис. 3 представлены
лений).
графики динамики численности мутантов и не-
БИОФИЗИКА том 65
№ 5
2020
МОДЕЛЬ ПРЕДПОСЫЛОК ВИДООБРАЗОВАНИЯ
935
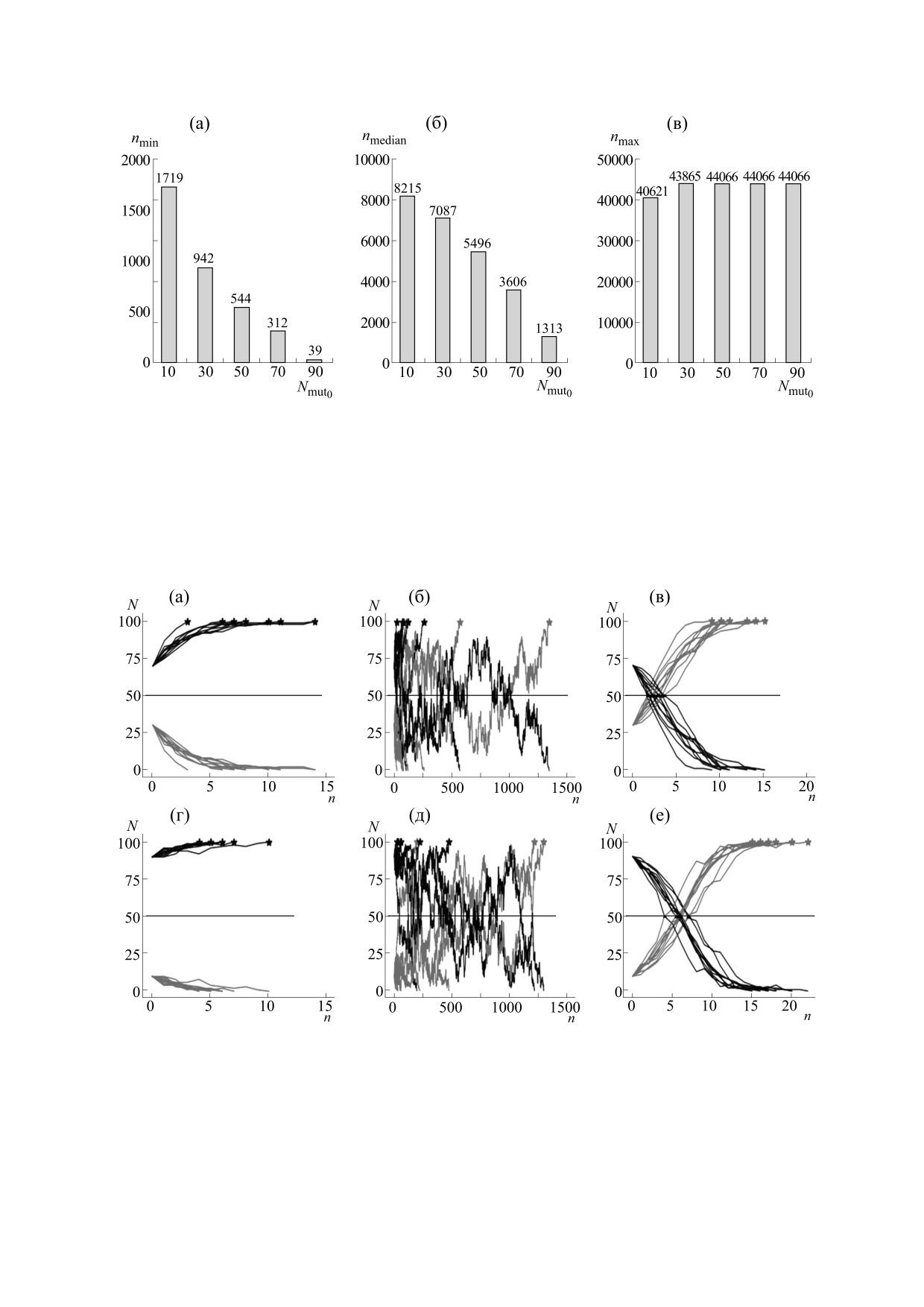
Рис. 2. Графики зависимости количества поколений n, в которых достигается 100%-е доминирование мутантов, от до-
ли мутантов (10, 30, 50, 70, 90) в исходном поколении (Nmut0) для бесполой популяции: (а) - минимальное количество
поколений (nmin); (б) - медианное количество поколений (nmedian); (в) - максимальное количество поколений (nmax).
Размер исходного поколения популяции постоянный (N = 100); 10000 экспериментов. Цифрами над столбцами ука-
зано значение поколений.
Рис. 3. Результаты численного эксперимента для двуполой популяции (10 экспериментов) в соответствии с моделью
Райта-Фишера при различных вариантах соотношения «мутанты/не-мутанты» в исходном поколении и значении ко-
эффициента pmut. Соотношение «мутанты/не-мутанты» от общей численности популяции в исходном поколении -
30/70: (а) - pmut = 0.3; (б) - pmut = 0.5; (в) - pmut = 0.7. Соотношение «мутанты/не-мутанты» от общей численности
популяции в исходном поколении - 10/90: (г) - pmut = 0.3; (д) - pmut = 0.5; (е) - pmut = 0.7. Размер популяции N = 100.
Светло-серые линии - мутанты, темно-серые линии - не-мутанты. Звездочками отмечены поколения, в которых кло-
ны достигают 100%-го доминирования. Черная горизонтальная линия: N = 50.
БИОФИЗИКА том 65
№ 5
2020
936
ГАРАЕВА и др.
Рис. 4. Графики зависимости количества поколений (n), необходимых для закрепления нового признака (100%-го
доминирования мутантов в популяции), от количества мутантов в популяции в исходном поколении (Nmut0) и
вероятности мутации потомства от смешанной пары родителей (pmut): (а) - минимальное количество поколений
(nmin); (б) - медианное (nmedian); (в) - максимальное количество поколений (nmax). Размер популяции N=100;
10000 экспериментов. Цифрами над столбцами указано количество поколений.
мутантов при pmut = 0.5 и различных вариантах
сти до pmut =0.7 - от десяти до двух поколений (при
соотношения мутанты/не-мутанты в исходном
аналогичных соотношениях); с уменьшением ко-
поколении по поколениям для десяти экспери-
эффициента вероятности до pmut = 0.3 - домини-
ментов. В отличие от модели Морана при поло-
рование мутантов не отмечено (рис. 4а). Для меди-
вом размножении случайные выборки учитывают
анных значений количества поколений, в которых
не только клоны мутантов и не-мутантов, но и
фиксируются мутанты, получаем зависимость,
половую принадлежность особей в клонах - сам-
сходную с минимальными значениями количества
цы и самки.
поколений (рис. 4б), а для максимальных значе-
Для полового размножения, аналогично моде-
ний, в отличие от модели бесполого размножения
ли Морана, отмечена зависимость количества по-
(рис. 2в), отмечается наличие пика при соотноше-
колений, необходимых для закрепления нового
нии мутанты/не-мутанты, равном 50/50, что связа-
признака (100%-го доминирования мутантов в
но с увеличением вариабельности при данном рас-
популяции) от начального соотношения мутан-
пределении в двух группах организмов (мутанты и
ты/не-мутанты при постоянном значении коэф-
не-мутанты, самцы и самки) (рис. 4в).
фициента pmut (рис. 3). Количество поколений,
Для визуализации плотности распределения
необходимых для закрепления нового признака,
вероятностей случайной величины (количества
зависит от значения коэффициента вероятности
поколений, в которых достигается доминирова-
фиксации новой мутации (pmut): при увеличении
ние мутантов или не-мутантов в популяции) в мо-
коэффициента вероятности фиксации мутаций
делях бесполого и полового размножения были
количество поколений, необходимых для фикса-
использованы «скрипичные» графики (violin plot)
ции мутаций, сокращается (рис. 3б, в, д, е); при
(рис. 5). (Вероятность того, что доминирование
уменьшении коэффициента вероятности фикса-
произойдет в интервале поколений [n1, n2], равна
ции мутаций количество поколений, необходи-
площади области значений на этом интервале:
мых для фиксации мутаций, увеличивается (рис.
чем больше площадь области, тем выше вероят-
3б,в,д,е) или мутации не фиксируются (рис. 3а, г).
ность.) Чем шире светло-серая область на графи-
Для 10000 экспериментов отмечены аналогич-
ке, тем выше вероятность достижения мутантами
ные зависимости. Минимальное количество поко-
100%-го доминирования в популяции в данном
лений, необходимых для доминирования мутан-
поколении (n) (это же справедливо и для темно-
тов, при значении коэффициента вероятности
серых областей - не-мутантов). С увеличением
pmut= 0.5, уменьшается от 68 поколений при соот-
начальной доли мутантов светло-серая половина
ношении мутанты/не-мутанты 10/90 до двух поко-
«скрипки» (мутанты) сдвигается к оси х: чем
лений при соотношении мутанты/не-мутанты
больше доля мутантов в исходном поколении,
90/10 (рис. 4а); с ростом коэффициента вероятно-
тем меньше поколений необходимо для домини-
БИОФИЗИКА том 65
№ 5
2020
МОДЕЛЬ ПРЕДПОСЫЛОК ВИДООБРАЗОВАНИЯ
937
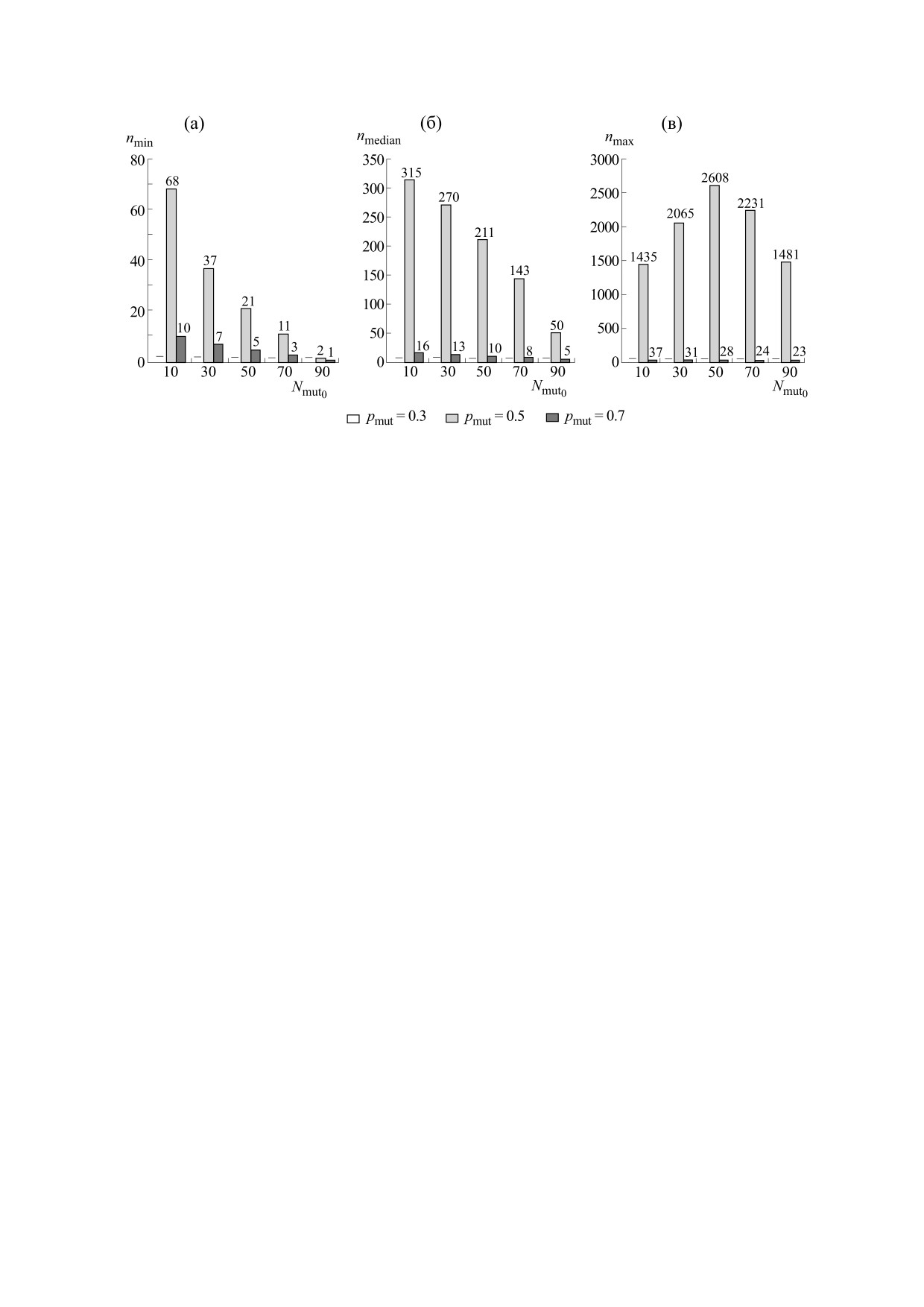
Рис. 5. Скрипичные графики зависимости количества поколений (n), необходимых для закрепления нового признака
(100%-го доминирования мутантов в популяции), от количества мутантов в популяции в исходном поколении (Nmut0)
для различных вариантов начального количества мутантов в популяции: (а) - модель бесполого размножения
Морана, (б)
- модель полового размножения Райта-Фишера. Размер популяции N
= 100; pmut
= 0.5;
10000 экспериментов. Светло-серые области на графике
- мутанты, темно-серые
- не-мутанты. Белыми
горизонтальными линиями показаны медианы распределений.
рования мутантов. Белыми горизонтальными ли-
тем к локальному изменению коллективного со-
ниями показаны медианы распределений. Мас-
стояния малой группы песчинок относительно
штабирование рассчитывали отдельно для каж-
соседних песчинок, что вызывает песочную лави-
дого графика. Для наглядности графики для
ну. При видообразовании способность особей к
модели бесполого размножения Морана построе-
изменению генотипа связана с количеством му-
ны до 15000 поколений, а для модели полового
тантов в исходном поколении и особенностями
размножения Райта-Фишера - до 600: выше этих
размножения - характеристиками особей в попу-
значений для n происходят единичные события
ляции. Если при бесполом размножении форми-
фиксации мутантов или не-мутантов.
рование устойчивого клона (100%-е доминирова-
ние) в поколениях определяется только исход-
Несмотря на то что рандомные модели отлича-
ным соотношением особей-мутантов и не-
ются условиями случайного распределения - на-
мутантов, то при половом размножении добавля-
личием двух групп при бесполом размножении и
ется еще одна характеристика - наличие пола. В
четырех групп при половом размножении, полу-
результате размножения (кооперативного эф-
ченные результаты позволяют выявить опреде-
фекта взаимодействующих особей в процессе
ленные закономерности: с увеличением началь-
размножения) происходит фазовый переход си-
ной доли мутантов уменьшается минимальное
стемы: состояние неустойчивости по мере накоп-
количество поколений, необходимых для фикса-
ления мутаций в поколениях преобразуется в ка-
ции мутации при случайной выборке особей.
чественно новое устойчивое состояние - новый
вид (100%-е доминирование) или вымирание му-
Рассмотренные модели отражают процесс са-
тантов или не-мутантов.
моорганизованной критичности [1, 2]. Примени-
тельно к случайным процессам фиксации мута-
ций это состояние системы, при котором в
ПЕРКОЛЯЦИОННАЯ МОДЕЛЬ ОТБОРА
результате кооперативного эффекта взаимодей-
МУТАЦИЙ. РОЛЬ ДЕТЕРМИНИРОВАННЫХ
ствующих элементов (особей в популяции) и в за-
И СЛУЧАЙНЫХ ПРОЦЕССОВ
висимости от их характеристик происходит на-
копление малых флуктуаций (мутаций), которое
Модели Морана и Райта-Фишера учитывают
при определенных условиях способно перерасти
исключительно случайные процессы, однако при
в гигантскую флуктуацию, т. е. сформировать но-
закреплении мутаций в поколениях существен-
вую структуру - новый вид или вызвать элимина-
ную роль играют также детерминированные про-
цию популяции. В модели «кучи песка» [2] цеп-
цессы. Представленная в данной работе модель
ная реакция, поддерживающая критическое со-
перколяционной решетки как механизмов отбора
стояние системы, определяется неустойчивостью
мутаций в процессе видообразования включает
песчинок - способностью к изменению динами-
как детерминированные, так и случайные про-
ческого состояния сначала одной песчинки, а за-
цессы.
БИОФИЗИКА том 65
№ 5
2020
938
ГАРАЕВА и др.
Перколяция - это явление протекания потока
деляется детерминированными процессами, что
подвижной среды через пористые материалы,
связано с наличием достаточно устойчивых кла-
при котором формируется хотя бы один непре-
стеров носителей новых признаков. Согласно ра-
рывный путь через соседние проводящие узлы ре-
боте [30], в случае фиксации мутации в течение
шетки [21]. Порог перколяции как количествен-
десяти поколений мутация становится закрепив-
ная характеристика протекания зависит от пара-
шейся - это третий уровень, определяемый взаи-
метров среды и доли проводящих узлов, при
модействиями между популяциями в границах
которой возникает перколяционный кластер [22].
общего вида. Иными словами, если для единич-
Иными словами, кластером следует считать сово-
ных особей мутации (малые флуктуации) являют-
купность кооперативно связанных в перколяци-
ся обратимыми, поскольку преимущественно
онном пространстве узлов и каналов. В такой си-
определяются случайными процессами, то при
стеме существенно соотношение случайных и
формировании кластера носителей новой мута-
скоррелированных процессов [22-25]. Процесс
ции (клона) малые флуктуации способны перера-
фиксации мутаций в свою очередь также опреде-
сти в гигантскую. Поэтому на этом уровне фор-
ляется детерминированными и случайными про-
мируются предпосылки видообразования - спе-
цессами. В модели естественный отбор представ-
циализация и дифференциация, а конечный
лен как явление перколяции [26, 27], где перколя-
результат, связанный с наличием достаточно
ционный фильтр
- физический инструмент
устойчивых кластеров носителей новых призна-
отбора оптимальных для видообразования мута-
ков, в значительной степени определяется детер-
ций на популяционном уровне. Детерминиро-
минированными процессами [27]. Устойчивое
ванные взаимодействия мутаций в череде поко-
формирование кластеров является точкой бифур-
лений и случайные процессы дрейфа определяют
кации, при преодолении которой возможны два
коллективное поведение узлов и связей (мута-
варианта: формирование нового вида (при нали-
ций) в пространстве перколяционной решетки
чии мутаций, способствующих видообразова-
популяции.
нию) или элиминация носителей мутации (при
Естественный отбор (подобно фильтру перко-
наличии негативных мутаций). «Знак» мутаций
на популяционном уровне (положительных, от-
ляционной решетки) определяет закономерность
рицательных и нейтральных) определяется их
прохождения потока новых мутаций [27]. Давле-
адаптивной пользой на кратком временном ин-
ние для протока создает пул мутаций, и в резуль-
тервале и в дальнейшем, в зависимости от усло-
тате отбора происходит выборка и фиксация му-
таций. Перколяции, как система связанных эле-
вий среды, может изменить знак на противопо-
ложный, поэтому в модели мутации рассматрива-
ментов (особей
- носителей мутаций), в
ются вне зависимости от их направленности [27].
зависимости от характеристик отбора и потока
мутаций могут стать либо разрастающимися -
Скорость закрепления новой мутации (в долях
устойчивыми, либо - неустойчивыми. При от-
от общего количества мутаций) в поколениях
сутствии детерминированных механизмов не-
определяется детерминированными процессами
устойчивые перколяции стохастичны и отфиль-
отбора и случайными процессами генетического
тровываются отбором, а устойчивые перколяции
дрейфа и может быть описана следующим урав-
определяют эволюционную траекторию популя-
нением [26]:
ции и вида [27]. Фиксация новых мутаций на по-
пуляционном уровне зависит от соотношения ра-
dMС/dt = -lnMС ⋅ pС(MR - MB) Ne/N + kW,
(1)
нее закрепившихся «разрешающих» и «запреща-
где N - численность популяции, Ne - доля особей
ющих» мутаций
[16]. В процессе фиксации
от общей численности популяции, способных к
мутаций преимущество получают мутации, кото-
размножению и являющихся носителями мута-
рым подвержены не единичные особи, а кластеры
ций - кластер, (N - Ne) - количество особей не-
в череде поколений [28, 29].
мутантов. Отношение Ne/N лежит в интервале
Формирование нового вида происходит по-
0.3-0.8 (согласно работе [31], в среднем, мини-
этапно. На первом уровне процесс определяется
мальная величина эффективного размера попу-
точечными взаимодействиями особей - носите-
ляции Ne составляет 30%, а максимальная вели-
лей мутаций, и мутации являются обратимыми,
чина - 60-85% от общей численности популяции
поскольку значителен вклад случайных процес-
[32]). MC - доля новых мутаций от общего коли-
сов. При наличии достаточного количества носи-
чества мутаций (нормированы на единицу), pC -
телей новой мутации происходит формирование
кластера носителей мутаций - это промежуточ-
вероятность фиксации новых мутаций. Суще-
ный уровень локального взаимодействия (конку-
ствуют различные подходы к определению веро-
ренции) кластеров, связанный с началом специа-
ятности мутации, согласно которым, как прави-
лизации и дифференциации
- необходимых
ло, значение pC колеблется в пределах 0.5~1.0 [33,
условий образования вида. На этом уровне ко-
34]. Поскольку для таксонов разного уровня био-
нечный результат в значительной степени опре-
логической сложности вклад случайных процес-
БИОФИЗИКА том 65
№ 5
2020
МОДЕЛЬ ПРЕДПОСЫЛОК ВИДООБРАЗОВАНИЯ
939
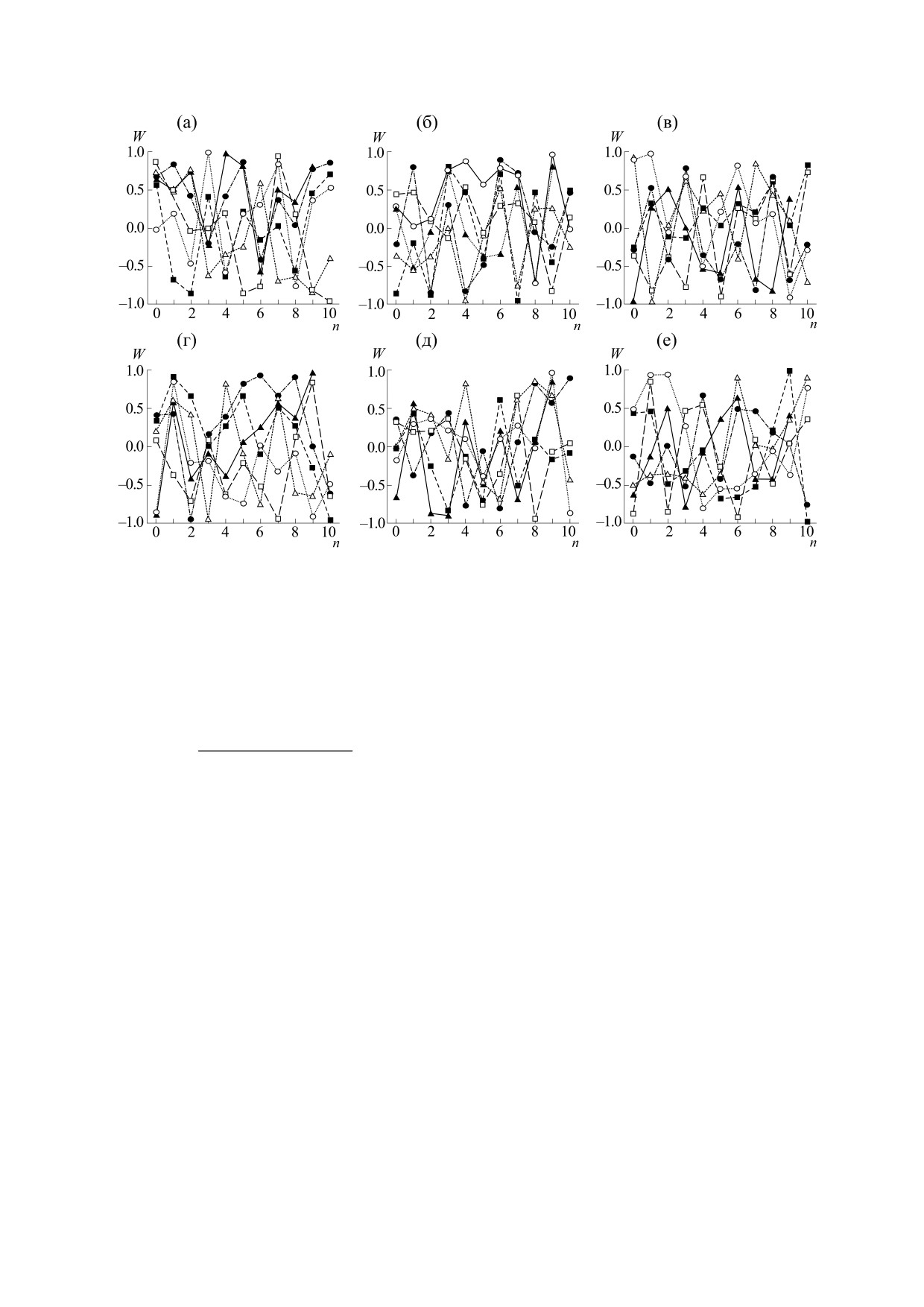
Рис. 6. Выборки случайных чисел для десяти поколений n при различных значениях (Ne/N) для бесполого ((а) - № 1,
(б) - № 2, (в) - № 3) и полового ((г) - № 1, (д) - № 2, (е) - № 3) размножения. Отношение Ne/N равнялось:
0.3 (темные треугольники), 0.4 (светлые треугольники), 0.5 (темные квадраты), 0.6 (светлые квадраты), 0.7 (темные
кружки) и 0.8 (светлые кружки).
сов различен, используя данные работы [35], рас-
коление) в соответствии с выборками случайных
считываем вероятность закрепления новых мута-
чисел для полового и бесполого размножения
ций при отсутствии дрейфа по формуле
(рис. 6).
Начальные условия: MC
= 0.1, популяция со-
Ne
/
N
· s
p
≈
(2)
C
стоит из N = 100 особей, из которых Ne0 = 30 - об-
{
1
-
exp
(
−2Ne
/
Ns
)}
ладатели положительной мутации. Сумма разре-
MR - доля ранее закрепившихся в популяции раз-
шающих и запрещающих мутаций (в долях) опре-
делена как MR + MB = 1. Необходимое условие
решающих мутаций (нормированы на 1). MB -
закрепления новой мутации - наличие преобла-
доля закрепившихся ранее в популяции запреща-
дающей доли поддерживающих мутаций. Полага-
ющих мутаций (нормированы на единицу). Но-
ем MR = 0.51, и тогда MR - MB = 0.02. Уравнение
вая мутация MC может быть передана конечному
(1) принимает следующий вид:
числу особей в популяции в зависимости от соот-
ношения Ne/N, наличия и соотношения разреша-
dMС/dt = -0.02lnMС ⋅ pСNe/N + kW.
ющих (MR) и запрещающих (MB) мутаций, значе-
ний коэффициентов отбора (s) и дрейфа (k). s -
Величина детерминированной части уравне-
Коэффициент отбора (определяет скорость
ния (1) (-lnMС ⋅ pС(MR - MB) Ne/N) изменяется в
уменьшения частоты генотипа: с уменьшением
интервале от 0.007117 (Ne/N = 0.3) до 0.000985
коэффициента отбора уменьшается приспособ-
(Ne/N = 0.8). Отсюда пороговым значением ко-
ленность генотипов и увеличивается давление от-
эффициента дрейфа при MR - MB = 0.02 и Ne/N =
бора), для природных популяций принимаем
0.1 ≤ s ≤ 0.2 [36]. kW - слагаемое, описывающее
0.3 полагаем kth = 0.01: при kth < 0.01 в ходе за-
дрейф, где k - коэффициент дрейфа (определяет
крепления мутаций доминируют детерминиро-
случайные изменения частот аллелей и генотипов
ванные процессы, а при kth > 0.01 - случайные.
в популяции), W - случайная величина, равно-
мерно распределенная на отрезке [-1, 1]. Величи-
Скорость изменения Ne зависит только от ско-
на W изменяется каждые десять шагов (раз в по- рости смерти и рождения мутантов:
БИОФИЗИКА том 65
№ 5
2020
940
ГАРАЕВА и др.
−
−
+
+
dN /dn =−r
d
· (N − Ne) + r
b
· (N − Ne) − r
d
· Ne + r
b
· Ne,
(3)
где rd- - константа скорости смертности не-мутан-
тов, rb+ - константа скорости рождаемости мутан-
тов. В каждом поколении изменение численности
тов, rb- - константа скорости рождаемости не-му-
популяции определяется скоростями рождаемости и
тантов, rd+ - константа скорости смертности мутан-смертности мутантов и не-мутантов:
-
-
+
+
N
n
=-r
d
·(N
n−1
−
Ne
n−1
)
+
r
b
·(N
n−1
−
Ne
n−1
)
−
r
d
·Ne
n−1
+
r
b
·Ne
n−1
+
N
n−1
(4)
Начальные условия: популяция состоит из
смертность не-мутантов в два раза выше их рожда-
N0 = 100 особей, из которых Ne0 = 30 - обладате-
емости, а смертность мутантов в два раза ниже их
рождаемости, и уже в первом поколении все не-
ли положительной мутации, а (N0 - Ne0) = 70 -
мутанты вымирают. Поэтому принимаем следую-
особи, не имеющие мутацию (не-мутанты). Усло-
щие значения констант скорости: а) для полового
вие соответствия между значениями s и Ne0: мута-
размножения: смертность не-мутантов rd- = 0.75,
ция, повышающая приспособленность на 10%
(s = 0.1), поддерживается отбором при численно-
рождаемость не-мутантов rb- = 0.5, смертность му-
сти мутантов от 10 особей [37].
тантов rd+ = 0.33, рождаемость мутантов rb+ = 0.5
Рассмотрим уравнение (4) для бесполого и по-
(рис. 7а); б) для бесполого размножения: смерт-
лового размножения. Если принять, что рождае-
ность не-мутантов rd- = 1.5, рождаемость не-му-
мость в обоих случаях одинакова и разница наблю-
тантов rb- = 1, смертность мутантов rd+ = 0.66,
дается только в смертности, то в этом случае кон-
станты скорости (в долях) равны: смертность не-
рождаемость мутантов rb+ = 1 (рис. 7б). Поскольку
мутантов rd- = 2, рождаемость не-мутантов (rb-)
мы рассматриваем только те мутации, которые
способствуют видообразованию (положительные),
равна рождаемости мутантов (rb+): rb+ = rb-= 1,
то константы скорости смертности не-мутантов в
смертность мутантов rd+ = 0.5. В этом случаеполтора раза больше, чем скорость их рождаемо-
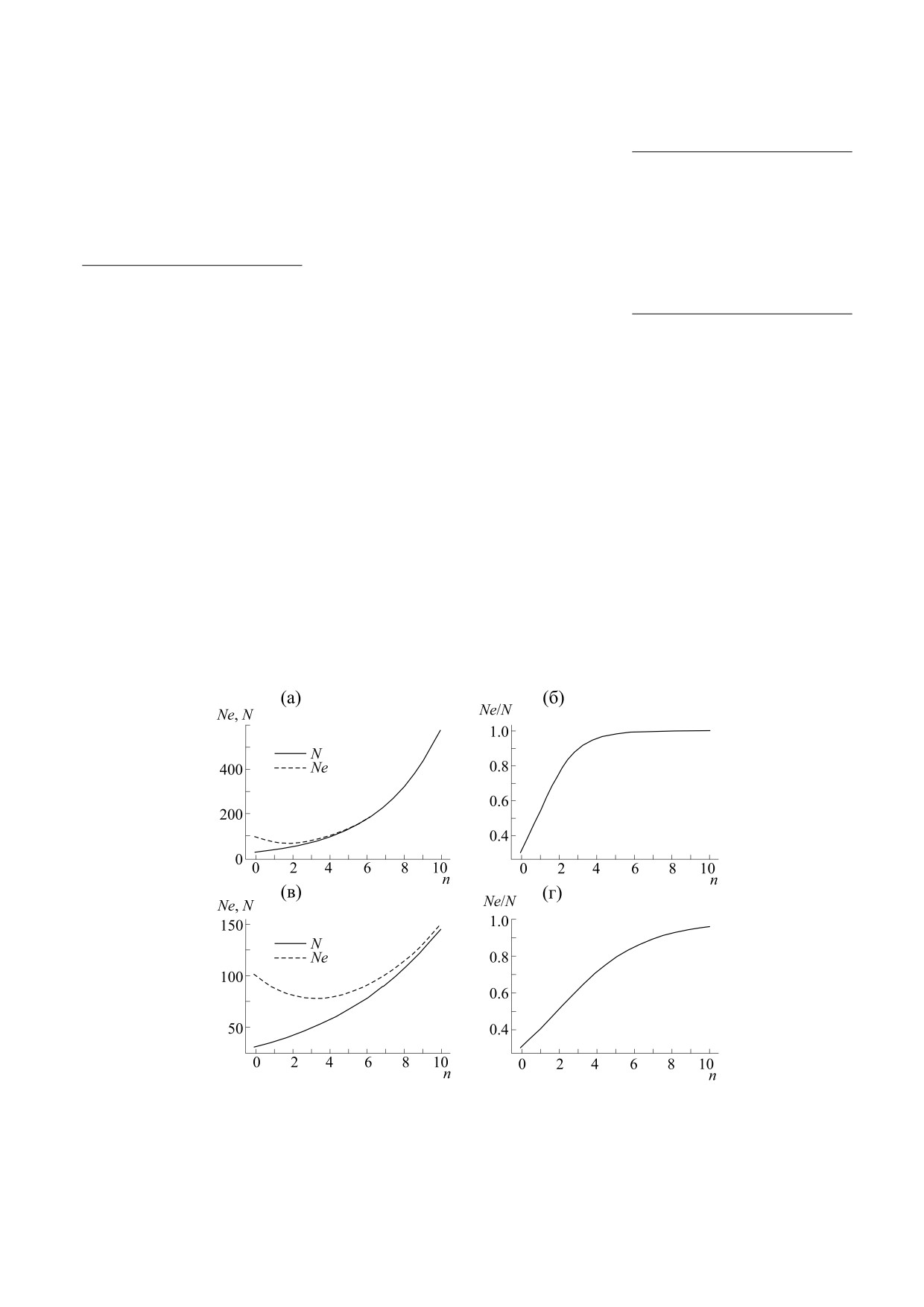
Рис. 7. Графики динамики популяции по поколениям (n). Для бесполого размножения: (а) - динамика численности
популяции (N) и мутантов (Ne), (б) - динамика отношения Ne/N; для полового размножения: (в) - динамика
численности популяции (N) и мутантов (Ne), (г) - динамика отношения Ne/N.
БИОФИЗИКА том 65
№ 5
2020
МОДЕЛЬ ПРЕДПОСЫЛОК ВИДООБРАЗОВАНИЯ
941
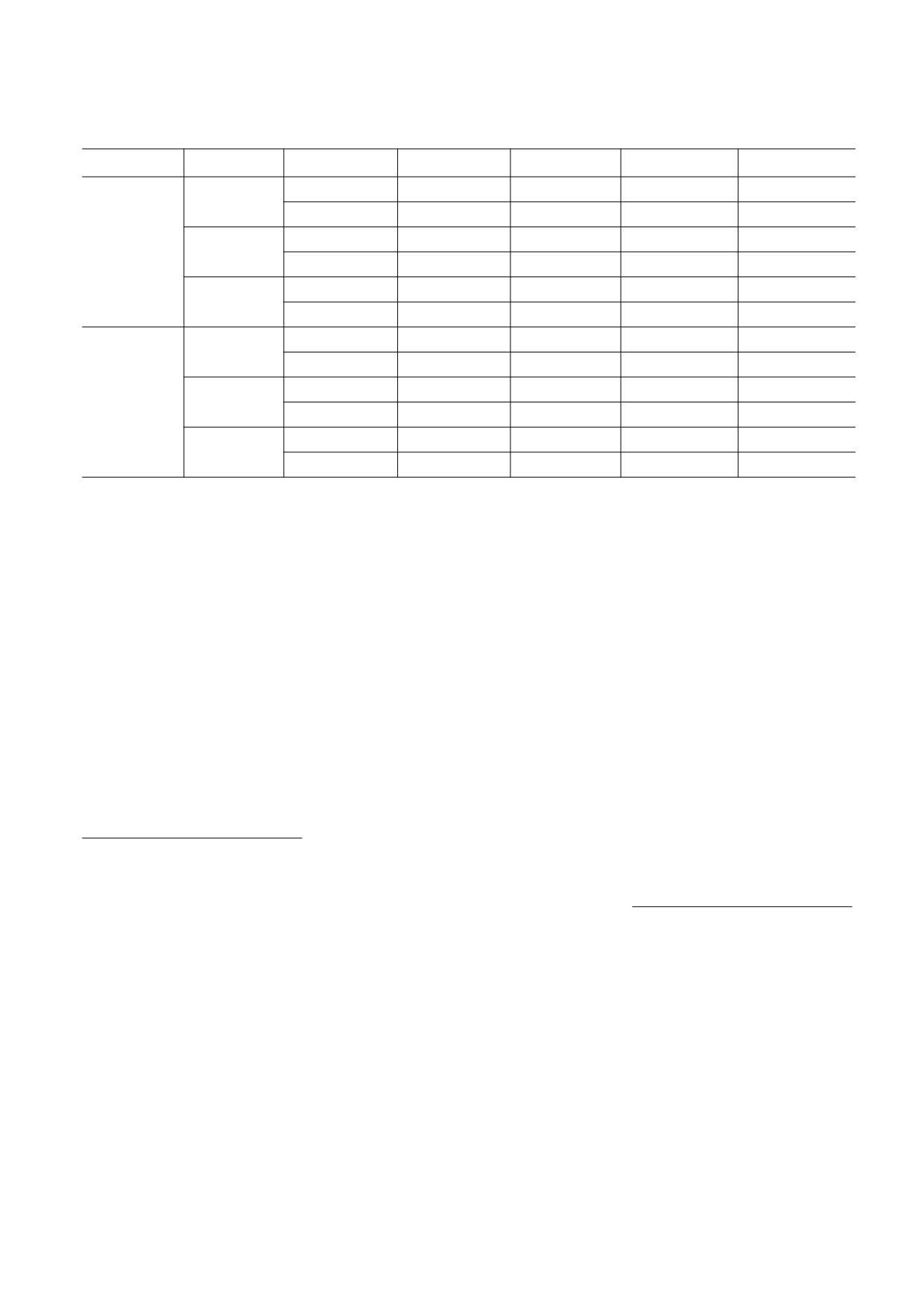
Таблица 1. Параметры условий решения уравнений (1) для полового и бесполого размножения
s
pC
kth
MC
MR
MR - MB
kth
0.1
0.51515
0.01
0.1
0.51
0.02
0.01
сти: не-мутанты оказываются менее приспособ-
жения, согласно уравнению (1), представлены в
ленными к изменившимся условиям, поэтому их
табл. 1-3.
элиминация происходит быстрее.
Для демонстрации роли случайных чисел, в
соответствии с уравнением (1), построены графи-
Полученные графики адекватно воспроизво-
ки динамики фиксации мутаций при половом и
дят динамику популяции при бесполом и поло-
бесполом размножении для трех выборок случай-
вом размножении в условиях достаточного объе-
ных чисел: s = 0.1, (MR - MB) = 0.8, k1 = 0.01,
ма ресурса: для бесполого размножения во время
k2 = 0.6 (рис. 8, табл. 2).
логарифмической фазы происходит более интен-
Графики на рис. 8 демонстрируют следующие
сивный экспоненциальный рост [38] отношения
закономерности. При пороговом значении коэф-
мутантов к численности популяции по поколени-
фициента дрейфа (kth = 0.01) процесс фиксаций
ям (рис. 7б), чем для полового размножения
мутаций в поколениях для двух типов размноже-
(рис. 7г). Ввиду особенностей бесполого размно-
ния и трех выборок случайных чисел (№№ 1-3)
жения численности мутантов и популяции в це-
протекает по одному сценарию (рис. 8а,д). При
лом выравниваются к четвертому поколению,
k = 0.6 (значительно больше kth = 0.01) как при
а при половом размножении - к десятому поко-
половом (рис. 8е-и), так и без бесполом (рис. 8б-
лению.
г) размножении возникают хаотические процес-
Численное решение уравнения (1) для полово-
сы, и прохождение мутаций может происходить в
го и бесполого размножения выполнено на языке
разных поколениях и при разных значениях MС (в
программирования Python 3.7 с использованием
долях), что объясняется не только значением ко-
библиотек NumPy и Matplotlib. Параметры фик-
эффициента дрейфа, но и выборкой случайных
сации мутаций для полового и бесполого размно- чисел.
Рис. 8. Графики динамики фиксации мутации (в долях от общего количества мутаций) в поколениях (n) в зависимости
от начальный значений Ne/N, k и выборки случайных чисел для бесполого ((а)-(г)) и полового ((д)-(з)) размножения.
Бесполое размножение: (а) - k = 0.01, выборки №№ 1-3 (при трех выборках получаются визуально неразличимые
графики); (б) - k = 0.6, выборка № 1; (в) - k = 0.6, выборка № 2; (г) - k = 0.6, выборка № 3. Половое размножение:
(д) - k = 0.01, выборки №№ 1-3 (при трех выборках получаются визуально неразличимые графики); (е) - k = 0.6,
выборка № 1; (ж) - k = 0.6, выборка № 2; (з) - k = 0.6, выборка № 3. Постоянные значения: s = 0.1, MR = 0.9.
Оттенками серого обозначено возрастание доли новых мутаций (от 0 до 1).
БИОФИЗИКА том 65
№ 5
2020
942
ГАРАЕВА и др.
Таблица 2. Параметры решения уравнения (1) при половом и бесполом размножении для разных выборок
случайных чисел при MR = 0.9
Выборка
k
MR
MR - MB
MC(min)
MC(max)
0.01
0.9
0.8
0.05
1.00
№ 1
0.6
0.9
0.8
0.05
1.00
Бесполое
0.01
0.9
0.8
0.05
1.00
размножение
№ 2
0.6
0.9
0.8
0.04388
1.00
s = 0.1
0.01
0.9
0.8
0.05
0.99957
№ 3
0.6
0.9
0.8
0.05
1.00
0.01
0.9
0.8
0.05
1.00
№ 1
0.6
0.9
0.8
0.05
0.7985
Половое
0.01
0.9
0.8
0.05
1.00
размножение
№ 2
0.6
0.9
0.8
0.05
1.00
s = 0.1
0.01
0.9
0.8
0.05
0.9986
№ 3
0.6
0.9
0.8
0.05
1.00
ТРИГГЕРНАЯ МОДЕЛЬ
значения коэффициента дрейфа) процессов, а
ФИКСАЦИИ МУТАЦИЙ
также размера кластера носителей мутаций, при
этом доля (МС) увеличивается от МС(min) в пер-
На популяционном уровне особи, а затем кла-
стеры носителей мутаций, которые, в зависимо-
вом поколении до МС(max) в десятом поколении.
сти от численности особей, а также их генотипи-
Если в первом поколении процесс фиксации му-
ческих и фенотипических характеристик, и спек-
таций можно охарактеризовать как малую флук-
тра ранее зафиксированных и новых мутаций,
туацию, то к десятому поколению снижается уро-
определяют особенности взаимодействия с дру-
вень воздействия случайных процессов (по срав-
гими особями в популяции. Это состояние не-
нению с первым поколением) и формируется
устойчивости постепенно преобразуется в каче-
гигантская флуктуация - новая структура. Таким
ственно новое устойчивое состояние - новый вид
образом, перколяционную решетку фиксации
или вымирание. В модели перколяционной ре-
мутаций можно представить в качестве много-
шетки рассматривается фиксация мутаций в те-
слойной (десять слоев) структуры, где процесс за-
чение десяти поколений под управлением детер-
крепления мутаций в десяти поколениях под дей-
минированных (в зависимости от доли разреша-
ствием отбора и дрейфа определяется стационар-
ющих мутаций) и случайных (в зависимости от
ным решением триггерного уравнения:
2
−u,
M
≤
M
(k
=
0),
∂u
∂
u
С
С
−
D
=
0.3
<
x
<
0.8.
(5)
2
∂t
∂x
1-
u,
M
С
>
M
С
(k
=
0),
Отношение Ne/N лежит в интервале 0.3-0.8,
MR- доля ранее закрепившихся в популяции
откуда x определяется соотношением Ne/N в пре-
разрешающих мутаций; MB - доля ранее закре-
делах x ∈ [0.3; 0.8]. MС > MС(k = 0) - необходимое
пившихся в популяции запрещающих мутаций;
условие закрепления новой мутации. Поскольку
D - коэффициент диффузии прохождения новых
модель (5) определяет только два возможных ва-
мутаций. Согласно данным работы [39] принима-
рианта - прохождение или не прохождение но-
вых мутаций по поколениям, значение MС при
ем D = 0.01 (средняя скорость мутаций на кодиру-
k = 0 сравнивается с MС при k ≠ 0. u - Функция (в
ющий геном за поколение для организмов). От-
условных единицах) доли новых мутаций (MС)
ношение Ne/N лежит в интервале 0.3-0.8. Триг-
герная модель
(5) определяет только два
по поколениям в зависимости от Ne/N (уравне-
ние 1); x определяется соотношением Ne/N в пре-
возможных варианта - прохождение или не про-
делах x ∈ [0.3; 0.8]; t - поколения (n) от 1 до 10.
хождение новых мутаций по поколениям.
БИОФИЗИКА том 65
№ 5
2020
МОДЕЛЬ ПРЕДПОСЫЛОК ВИДООБРАЗОВАНИЯ
943
Таблица 3. Параметры решения уравнения (5) при половом и бесполом размножении
Выборка
k
MR
MC(min)
MC(max)
MR
MС(min)
MС(max)
0
0.6
0.05
0.7989
0.8
0.05
0.98115
0.01
0.6
0.05
0.8143
0.8
0.0.5
0.99075
0.1
0.6
0.05
0.9699
0.8
0.05
1.00
№ 1
0.2
0.6
0.05
1.00
0.8
0.05
1.00
0.3
0.6
0.05
1.00
0.8
0.05
1.00
0.4
0.6
0.05
1.00
0.8
0.05
1.00
0.5
0.6
0.04674
1.00
0.8
0.05
1.00
0.01
0.6
0.05
0.82116
0.8
0.05
0.99365
0.1
0.6
0.05
1.00
0.8
0.05
1.00
Бесполое
размножение
0.2
0.6
0.0448
1.00
0.8
0.05
1.00
№ 2
s = 0.1
0.3
0.6
0.03437
1.00
0.8
0.05
1.00
0.4
0.6
0.021
1.00
0.8
0.0491
1.00
0.5
0.6
0.00788
1
0.8
0.04174
1.00
0.01
0.6
0.05
0.80813
0.8
0.05
0.98767
0.1
0.6
0.05
0.92124
0.8
0.05
1.00
0.2
0.6
0.05
1.00
0.8
0.05
1.00
№ 3
0.3
0.6
0.05
1.00
0.8
0.05
1.00
0.4
0.6
0.044943
1.00
0.8
0.05
1.00
0.5
0.6
0.022169
1.00
0.8
0.05
1.00
0.01
0.6
0.05
0.8197
0.8
0.05
0.993996
0.1
0.6
0.0.5
1.00
0.8
0.05
1.00
0.2
0.6
0.0.5
1.00
0.8
0.05
1.00
№ 1
0.3
0.6
0.0.5
1.00
0.8
0.05
1.00
0.4
0.6
0.03549
1.00
0.8
0.05
1.00
0.5
0.6
0.015687
1.00
0.8
0.05
1.00
0.01
0.6
0.05
0.80983
0.8
0.05
0.991484
0.1
0.6
0.05
0.9565
0.8
0.05
0.981053
Половое
0.2
0.6
0.05
1.00
0.8
0.05
1.00
размножение
№ 2
0.3
0.6
0.05
1.00
0.8
0.05
1.00
s = 0.1
0.4
0.6
0.05
1.00
0.8
0.05
1.00
0.5
0.6
0.05
1.00
0.8
0.05
1.00
0.01
0.6
0.05
0.8026
0.8
0.05
0.98491
0.1
0.6
0.05
0.85736
0.8
0.05
0.981053
0.2
0.6
0.05
0.9209
0.8
0.05
0.981053
№ 3
0.3
0.6
0.02173
1.00
0.8
0.05
1.00
0.4
0.6
0.0036
1.00
0.8
0.04625
1.00
0.5
0.6
0.0007
1.00
0.8
0.0383
1.00
Численное решение получено с помощью ме-
ческое уравнение решается при помощи метода
тода релаксации решения параболической задачи
прямых в соответствии со схемой CROS1 (ком-
к решению соответствующей стационарной зада-
плекснозначной одностадийной схемой Розен-
чи на большом промежутке времени. Параболи-
брока) [40].
БИОФИЗИКА том 65
№ 5
2020
944
ГАРАЕВА и др.
Опишем алгоритм численного решения урав-
ux(0.3; t) = ux(0.8; t) = 0. Эти однородные условия
нения (5). Обозначим
Неймана означают отсутствие влияния на рас-
сматриваемую систему процессов, происходящих
−u,
M
≤
M
(k
=
0),
при Ne/N ≤ 0.3 или Ne/N ≥ 0.8.
C
C
f u)
=
1-
u,
M
>
M
(k
=
0)
На отрезке x ∈ [0.3; 0.8] введем равномерную
C
C
сетку из Nx + 1 точек: x0 = 0.3,…, xNx = 0.8. Заме-
Зададим начальные условия задачи как u(x, 0) =
ним вторую производную по координате разност-
= 1 + th(x - 0.5). Эти условия отвечают случаю,
ной производной и перейдем к системе обыкно-
когда фазовый переход происходит при эффек-
венных дифференциальных уравнений. Добавим
тивном размере популяции Ne/N=0.5. Зададим
краевые условия. Получаем следующую систему
краевые
условия
следующим образом: обыкновенных дифференциальных уравнений:
−
y
0
+
y
1
=
0;
∂y
i
y
i+1
−
2
y
i
+
y
i−1
=
D
−
f
(y
)
=:
F
(y),
i
=
1,...,N
−
2;
(6)
2
i
i
∂t
h
−
y
N
−
2
+
y
N
−1
=
0.
Здесь h = (0.8 - 0.3)/Nx - шаг сетки по перемен-
му одностадийной комплекснозначной схемы
ной x, а время задается как t = τ × (количество ша-
Розенброка вектор
находится
(
)
0
1
1
ω=
ω
,
ω ωN-
гов по времени), τ - шаг по времени.
из системы линейных алгебраических уравнений:
Решение задачи на каждом следующем вре-
менном слое рассчитывается как
y = y + τReω;
1+
i
∂F
i
E
−
τ
ω=
F
(y
)
(7)
i
y
=
(y
,
y
,...,yN
) - решение на текущем времен-
0
1
-1
2
∂y
j
ном слое;
y
- решение на следующем временном
Распишем схему (7) подробно с учетом крае-
слое. Вектор
τReω- это приращение сеточных
вых условий для вектора
ω, вытекающих из крае-
значенийy искомой функции в приближении ис-
пользуемой численной схемы. Согласно алгорит- вых условий для вектора y (см. выражение (6)):
1+
i
1+
i
−
1+
τ
ω
+
τω
=
0,
(
)
0
1
2
2
2
1+
i D
1
+
i
2
∂
f
1+
i
ε
y
i+1
−
2
y
i
+
y
i−1
τ ω
−
1
+
τ
D
+
(y
)
ω
+
τ ω
=-D
+
f
(y
)
2
i−1
2
i
i
2
i+1
2
i
2
h
2
h
∂
y
2
h
h
1+
i
1
+
i
−
τ
ω
−
1
−
τ
ω
=
0.
(
)
N
−
2
(
)
N
−1
2
2
Численное решение уравнения (5) как оценка
Графики на рис. 9 и 10 демонстрируют, что в
процессов фиксации новых мутаций (в долях) по
результате детерминированных (в модели при
поколениям и при коэффициенте отбора, харак-
s = const определяются значения MС в зависимо-
терном для природных популяций (s = 0.1 [36]),
сти от значений k и MR) только часть мутаций
выполнено на языке программирования
способна сохраниться в кластере в течение десяти
Python 3.7 с использованием библиотек NumPy и
поколений. Для бесполого размножения отмече-
Matplotlib.
но закрепление мутаций в течение десяти поколе-
На рис. 9 и 10 представлено графическое реше-
ний при MR = 0.6: для выборки № 1 - при Ne/N =
ние триггерной модели (5) при постоянном зна-
= 0.7 (рис. 9а (№ 1), k = 0.01 и рис. 9б (№ 1),
чении коэффициента отбора (s = 0.1), MR = 0.6,
k = 0.5); для выборки № 2 - при Ne/N = 0.8
MR = 0.8 и различных значениях коэффициента
(рис. 9а (№ 2), k = 0.01); для выборки № 3 - при
дрейфа (k) для полового и бесполого размноже-
Ne/N = 0.8 (рис. 9а (№ 3), k = 0.01); при MR = 0.8:
ния (рис. 9). Расчетные данные представлены в
для выборки № 1 - при Ne/N = 0.7 (рис. 9а (№ 1),
табл. 3. В модели сравниваются текущие значения
k = 0.01); для выборки № 2 - при Ne/N = 0.8
параметров модели с исходными: k = 0 (без учета
(рис. 9а (№ 2), k = 0.01). Для полового размноже-
случайных процессов).
ния отмечено закрепление мутаций в течение де-
БИОФИЗИКА том 65
№ 5
2020
МОДЕЛЬ ПРЕДПОСЫЛОК ВИДООБРАЗОВАНИЯ
945
Рис. 9. Графики прохождения новых мутаций MC (в долях от общего количества мутаций) по поколениям для
бесполого размножения при постоянном значении коэффициента отбора (s = 0.1) в зависимости от начального
соотношения Ne/N, значений MR и k: верхний ряд соответствует MR = 0.6 для трех выборок случайных чисел; нижний
ряд соответствует MR = 0.8 для трех выборок случайных чисел. Светло-серые квадраты - новые мутации (в долях от
общей численности мутаций в популяции), закрепленные в поколениях; темно-серые - отсутствие закрепившихся
новых мутаций. Светло-серые непрерывные полосы в интервале от первого до десятого поколения означают
закрепившуюся мутацию в течение десяти поколений.
сяти поколений при MR = 0.6: для выборки № 1 -
череде последующих поколений (n > 10) также
возможна фиксация (например, при Ne/N = 0.5
при Ne/N = 0.5 (рис. 10а (№ 2), k = 0.01 и рис. 10б
(рис. 9а (№ 1), k = 0.01 или при Ne/N = 0.6
(№ 2), k = 0.5); при MR = 0.8: для выборки № 1 -
(рис. 10а (№ 3), k = 0.01).
при Ne/N = 0.5 (рис. 9а (№ 1), k = 0.01). Осталь-
ные мутации в ряду поколений 1 ≤ n ≤ 10 не сохра-
Таким образом, при MR = 0.6 и MR = 0.8 для
няются в кластере. Поскольку в модели (5) учи-
всех выборок случайных чисел в случае бесполого
тываeтся прохождение мутаций для десяти поко-
размножения фиксируется в два раза больше му-
лений, в принципе существует вероятность, что в таций, чем в случае полового размножения. По-
Рис. 10. Графики прохождения новых мутаций MC (в долях от общего количества мутаций) по поколениям для
полового размножения при постоянном значении коэффициента отбора (s = 0.1) в зависимости от начального
соотношения Ne/N, значения MR и k: верхний ряд соответствует MR = 0.6 для трех выборок случайных чисел; нижний
ряд соответствует MR = 0.8 для трех выборок случайных чисел. Светло-серые квадраты - новые мутации (в долях от
общей численности мутаций в популяции), закрепленные в поколениях; темно-серые - отсутствие закрепившихся
новых мутаций. Светло-серые непрерывные полосы в интервале от первого до десятого поколения означают
закрепившуюся мутацию.
БИОФИЗИКА том 65
№ 5
2020
946
ГАРАЕВА и др.
лученные нами данные для десяти поколений
отбора мутаций может существенно снижаться
коррелируют с экспериментальными данными,
(табл. 3). С ростом доли поддерживающих мута-
приведенными в работе [41], где было показано,
ций (MR = 0.8) усиливается детерминированная
что при одинаковом темпе мутирования в поло-
составляющая в ходе фиксации мутаций, поэтому
вых популяциях за 1000 поколений закрепляется
по сравнению с MR = 0.6 отмечено уменьшение
в пять раз меньше мутаций, чем в бесполых попу-
разбросов значений MС(min) - нижнего порога
ляциях такой же численности. Согласно класси-
перколяционной решетки отбора мутаций
ческим представлениям половое размножение
(табл. 3).
помогает отбору закреплять полезные мутации.
Анализ модели показывает, что важной осо-
Фиксации негативных и нейтральных мутаций
бенностью системы могут быть как обратимый,
при бесполом размножении происходят за счет
так и необратимый режимы поведения нижнего
«генетического автостопа» [42] (отбираться могут
порога перколяционной решетки отбора (фикса-
только целые геномы, и многие вредные мутации
ции мутаций), в зависимости от параметров (k,
могут закрепляться за счет полезных мутаций,
MR, Ne). При сохранении мутации в десяти поко-
расположенных в других местах генома) и кло-
нальной интерференции [37, 43] (конкуренция
лениях в популяции возникают необратимые
полезных мутаций в разных генах часто приводит
процессы, вызванные новым устойчивым состоя-
к потере большинства полезных мутаций в попу-
нием, - это верхний порог перколяционной ре-
ляции).
шетки фиксации мутаций, определяемый долей
новых мутаций в кластере в десятом поколении,
Перколяционные процессы характеризуются
например, при MR = 0.6, k = 0.5 и Ne = 0.7 (рис. 9);
особыми критическими точками, в которых ос-
при MR = 0.8, k = 0.01 и Ne = 0.5 (рис. 9). Верхний
новные параметры системы претерпевают каче-
ственные изменения - это порог протекания си-
порог перколяционной решетки фиксации мута-
ций - это эволюционный шаг видообразования
стемы [22, 44]. Для процесса фиксации мутаций
через перколяционную решетку отбора также ха-
[10], т.е. критический порог накопления мута-
рактерно наличие порогов протекания - нижнего
ций, который соответствует состоянию самоорга-
и верхнего.
низованной критичности. С точки зрения теории
самоорганизованной критичности в данной мо-
Как уже было сказано, если мутация сохрани-
дели, по сравнению с рассмотренными моделями
лась в кластере в течениe десяти поколений, то в
случайных процессов видообразования, добавля-
дальнейшем это может привести либо к форми-
ется еще одна характеристика особей - наличие и
рованию вида (положительная на данном этапе
соотношение ранее зафиксированных в популя-
видообразования мутация), либо элиминации ее
ции поддерживающих мутаций. Эта детермини-
носителей (негативная мутация). Для сохранив-
рованная составляющая моделей перколяцион-
шейся в течение десяти поколений (минимальное
ной решетки отбора и триггерной модели фикса-
количество поколений, необходимых для устой-
ции мутаций существенно снижает роль
чивой фиксации мутации в кластере [30]) нижний
случайных процессов в ходе образования.
порог перколяционной решетки фиксации мута-
ций определяется долей зафиксированных мута-
ций (MС) для воспроизводящей численности кла-
ЗАКЛЮЧЕНИЕ
стера носителей новой мутации (Ne) в одном по-
Для анализа процесса самоорганизованной
колении - MС(max). Иными словами, нижний
критичности рассмотрены модели случайных
порог перколяционной решетки отбора мутаций
процессов, а также случайных и детерминирован-
определяется минимальным количеством узлов
ных процессов фиксации мутаций при бесполом
(носителей мутаций в одном поколении) и связей
и половом размножении, позволяющие каче-
(мутаций). На этом уровне детерминированные и
ственно оценить вероятность возникновения но-
случайные процессы работают на уровне отдель-
вого вида. Показано, что для этих моделей пара-
ных особей в кластере, поэтому мутации (малые
метры взаимодействия особей в популяции в про-
флуктуации) в последующих поколениях могут
цессе самоорганизованной критичности зависят
не закрепиться, т.е. являются обратимыми. Свет-
от их генотипа и фенотипа, начального соотно-
ло-серые непрерывные полосы в течение десяти
шения «мутанты/не-мутанты» (определяет коли-
поколений (рис. 9, 10) означают, что мутации, за-
чество случайным образом выбранных контак-
крепившиеся в первом поколении, сохраняются в
тов) и половой принадлежности.
последующих поколениях. Как видно из графи-
Аналогично модели «кучи песка» [2] цепная
ков на рис. 9 и 10, а также данных табл. 3, при по-
реакция, поддерживающая критическое состоя-
стоянных значениях доли поддерживающих му-
ние системы, определяется неустойчивостью
таций (MR) с ростом коэффициента дрейфа (k)
особей в популяции - способностью к измене-
усиливается роль случайных процессов, и значе-
нию динамического состояния (фиксации мута-
ние нижнего порога перколяционной решетки
ции) сначала одной особи, а затем к локальному
БИОФИЗИКА том 65
№ 5
2020
МОДЕЛЬ ПРЕДПОСЫЛОК ВИДООБРАЗОВАНИЯ
947
изменению коллективного состояния кластера
12. S. Wright, Evolution 36, 427 (1982).
носителей мутаций относительно соседних пес-
13. S. A. Kauffman and S. Johnsen, J. Theor. Biol. 149, 467
чинок, что вызывает «лавину видообразования».
(1991).
При видообразовании способность особей к из-
14. L. A. Van Valen, Evol. Theory 1, 1 (1973).
менению генотипа связана с количеством мутан-
15. N. W. Watkins and M. P. Freeman, Science 320 (5874),
тов в исходном поколении, соотношением под-
323 (2008).
держивающих и запрещающих мутаций в популя-
16. T.N. Starr. L. K. Picton, and J. W. Thornton, Nature
ции, случайными процессами и особенностями
549 (7672), 409 (2017).
размножения. Полученные нами результаты
17. P. A. P. Moran, Math. Proc. Cambridge Philosoph.
триггерной модели подтверждаются представ-
Soc. 54 (1), 60 (1958).
ленными в научной литературе эксперименталь-
ными моделями оценки скорости мутаций одно-
18. K. F. Schulz and D. A. Grimes, Lancet 359 (9305), 515
клеточных эукариотов (Saccharomyces cerevisiae)
(2002).
[45-48]. Представленная нами модель может
19. D. Moher, S. Hopewell, K. F. Schulz, et al., Br. Med. J.
быть использована для анализа формирования
340, 869 (2010).
новых видов организмов в процессе биологиче-
20. S. Wright, Genetics 16 (2), 97 (1931).
ской эволюции.
21. А. Л. Эфрос, Физика и геометрия беспорядка (Нау-
ка, М., 1982).
ФИНАНСИРОВАНИЕ РАБОТЫ
22. J. Balogh and B. G. Pittel, Random Struct. Alg.,
No. 1-2, 257 (2007).
Работа выполнена при финансовой поддержке
23. C. McDiarmid, Math. Progr. Study, No. 13, 17 (1980).
Российского научного фонда (проект № 18-11-
00042).
24. H. L. Frisch and J. M. Hammersley, J. SIAM, No. 11,
894 (1963).
25. N. Fountoulakis, Internet Mathematics 4 (4), (2007).
КОНФЛИКТ ИНТЕРЕСОВ
26. A. E. Sidorova and V. A. Tverdislov, Moscow Universi-
Авторы заявляют об отсутствии конфликта
ty Phys. Bull. 67 (2), 213 (2012).
интересов.
27. A. Sidorova, N. Levashova, A. Garaeva, and V. Tverdis-
lov, Biosystems 193-194, 104 (2020).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
28. N. L. Komarova, Proc. Natl. Acad. Sci. USA 111,
10789 (2014).
Настоящая работа не содержит описания ка-
29. L. Wagstaff, G. Kolahgar, and E. Piddini, Trends Cell
ких-либо исследований с использованием людей
Biol. 23, 160 (2013).
и животных в качестве объектов.
30. В. Эбелинг, А. Энгель и Р. Файстель, Физика про-
цессов эволюции (Эдиториал УРСС, М., 2001).
СПИСОК ЛИТЕРАТУРЫ
31. М. Nei and Y. Imaizumi, Heredity 21, 183 (1966).
1. Р. Bak, С. Tang, and К. Wiesenfeld, Phys. Rev. Lett. 59
32. J. F. Crow and N. E. Morton, Evolution 9, 202 (1955).
(4), 381 (1987).
33. T. Bäck, in Proc. 5th Int. Conf. on Genetic Algorithms
2. P. Bak, How nature works: the science of self-organized
(1993), pp. 2-8.
criticality (Springer Science & Business Media, 2013).
34. J. Hesser and R. Männer, in Proc. Conf. “Parallel Prob-
3. H. J. Jensen, Self-Organized Criticality: Emergent Com-
lem Solving from Nature” (1990), pp. 23-32.
plex Behavior in Physical and Biological Systems (Cam-
35. B. Charlesworth, Nature Rev. Genet. 10 (3),
195
bridge University Press, Cambridge, 1998), v. 10.
(2009).
4. G. Pruessner, Self-Organised Criticality (Cambridge
36. V. Grant, Organismic evolution (H. Freeman and Co.,
University Press, 2012).
San Francisco, 1977).
5. P. Bak, et al., Phys. Rev. Lett. 88 (17), 178501 (2002).
37. S. F. Levy, J. R. Blundell, S. Venkataram, et al., Nature
6. E. Altshuler, et al., Phys. Rev. Lett. 86 (24), 5490
519, 181 (2015). DOI: 10.1038/nature14279
(2001).
38. C. Prats, D. López, A. Giró, et al., J. Theor. Biol. 241,
7. R. O. Dendy and P. Helander, Phys. Rev. E 57 (3), 3641
939 (2006).
(1998).
39. M. Lynch, et al., Nature Rev. Genet. 17 (11), 704
8. S. C. Chapman and N. W. Watkins, Plasma Physics and
(2016).
Controlled Fusion 51 (12), 124006 (2009).
40. H. H. Калиткин и П. В. Карякин, Численные мето-
9. A. T. Y. Lui, et al., Geophys. Res. Lett. 27 (7), 911
ды. Кн. 2. Методы математической физики (Акаде-
(2000).
мия, М., 2013).
10. K. Sneppen, P. Bak, H. Flyvbjerg, and M. H. Jensen,
41. M. J. McDonald, D. P. Rice, and M. M. Desai, Nature
Proc. Natl. Acad. Sci. USA 92, 5209 (1995).
531 (7593), 233 (2016).
11. S. J. Gould and N. Eldredge, Nature 366, 223 (1993).
42. J. M. Smith and J. Haigh, Genet. Res. 23 (1), 23 (1974).
БИОФИЗИКА том 65
№ 5
2020
948
ГАРАЕВА и др.
43. M. J. Wiser, N. Ribeck, and R. E. Lenski, Science 342,
46. H. T. Tran, D. A. Gordenin, and M. A. Resnick, Mol.
1364 (2013).
Cell. Biol. 19, 2000 (1999).
44. H. Kesten, Percolation Theory for Mathematicians
47. C. N. Greene and S. Jinks-Robertson, Genetics 159, 65
(Birkhauser, Boston, 1982).
(2001).
45. A. Morrison, A. L. Johnson, L. H. Johnston, and
48. A. J. Herr, et al., PLoS Genetics 7 (10), el002282
A. Sugino, EMBO J. 12, 1467 (1993).
(2011).
A Model of Speciation Preconditions in the Notions of Percolation
and Self-Organized Criticality Theories
A.Ya. Garaeva, A.E. Sidorova, V.A. Tverdislov, and N.T. Levashova
Faculty of Physics, Lomonosov Moscow State University, Leninskie Gory 1/2, Moscow, 119991 Russia
The model is presented which describes features of the formation of the upper threshold of percolation selec-
tion lattice—a share of the fixed mutations, the excess accumulation of which lead to irreversible processes in
the population. It is shown that a state of instability caused by a cooperative effect of interaction between mu-
tant individuals and the formation of their clusters, evolves into a qualitatively new stable state—a new species
or extinction of the population. Obtained results are confirmed by available in scientific literature experimen-
tal models for estimation of mutation rates in multicellular and unicellular eukaryotes. Models of random
processes of the fixation of mutations which happen asexually and in sexually reproducing species are pre-
sented to describe the process of self-organized criticality. With the use of these models it is possible to esti-
mate qualitatively the probability of occurrence of a new species.
Keywords: natural selection, self-organization, fluctuations, bifurcations, percolations, prohibitive and permitting
mutations
БИОФИЗИКА том 65
№ 5
2020