БИОФИЗИКА, 2020, том 65, № 5, с. 949-966
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 574.34:575.174.4
ЭКОЛОГО-ГЕНЕТИЧЕСКИЕ МОДЕЛИ
В ПОПУЛЯЦИОННОЙ БИОФИЗИКЕ
© 2020 г. Е.Я. Фрисман*, О.Л. Жданова**, Г.П. Неверова**
*Институт комплексного анализа региональных проблем ДВО РАН, 679016, Биробиджан, ул. Шолом-Алейхема, 4
**Институт автоматики и процессов управления ДВО РАН, 690041, Владивосток, ул. Радио, 5
E-mail: axanka@iacp.dvo.ru
Поступила в редакцию 25.11.2019 г.
После доработки 10.06.2020 г.
Принята к публикации 16.06.2020 г.
Динамика численности популяции и изменение ее генетической структуры находятся в сложных
причинно-следственных связях. В работе на простом модельном примере показано, что эволюци-
онный процесс плотностно-независимого естественного отбора по приспособленностям, опреде-
ляемым одним диаллельным локусом, может привести к изменению параметров роста популяции и
связанной с этим смене динамических режимов ее численности. Обсуждаются возможные механиз-
мы и направленность этих изменений. Результаты наглядно демонстрируют, что эволюционное из-
менение частот аллелей, сопровождающееся ростом средней приспособленности популяции, мо-
жет привести к циклическим, квазипериодическим и хаотическим режимам динамики ее числен-
ности. Эффекты, наблюдаемые в рассматриваемых моделях, во многом появляются в силу простого
объединения (суперпозиции) двух моделей: естественный отбор приводит к эволюционному росту
приспособленности, а плотностное регулирование при увеличении репродуктивного потенциала
приводит к известным бифуркациям, обеспечивающим различные колебания численности. Только
теперь эти флуктуации появляются (и проявляются) в ходе генетической эволюции. Вместе с этим
при объединении моделей появляются новые режимы, которые не наблюдались в каждой из моде-
лей отдельно: колебания частот генов, связанные с мультирежимностью рассматриваемых систем и
возникновением новых устойчивых аттракторов.
Ключевые слова: математическое моделирование, эволюция, естественный отбор, динамика попу-
ляции, ловушка бистабильности.
DOI: 10.31857/S0006302920050130
В 1798 г. Т. Мальтус опубликовал работу «Essay
эволюции Р. Фишер, С. Райт, Дж. Холдейн и
on the Principle of Population» («Опыт о законе на-
С.С. Четвериков [3-6] фактически применяли
родонаселения») [1], в которой привел обоснова-
модель Мальтуса, полагая мальтузианский пара-
ние модели, описывающей рост численности на-
метр зависимым от генотипа, а основоположники
селения земли по геометрической прогрессии со
теоретической экологии (динамической теории
знаменателем, названным впоследствии мальту-
экосистем) П. Ферхюльст и В. Вольтерра (а также
зианским параметром. Эта работа не только по-
Г.Ф. Гаузе) [7-10] рассматривали модель Мальту-
служила основой для многих демографических и
са, в которой этот параметр являлся функцией
мировоззренческих концепций, но и стимулиро-
численности популяции (или нескольких чис-
вала развитие основополагающих биологических
ленностей взаимодействующих популяций).
теорий.
Таким образом, были сформулированы базовые
Так, Ч. Дарвин приводит и подробно анализи-
концепции общей биологии, связанные с эволю-
рует модель Мальтуса в первой части своей зна-
цией популяций и динамикой экосистем. Однако
менитой книги: “On the Origin of Species by Means
полного синтеза экологических и генетических
of Natural Selection, or the Preservation of Favored
представлений и концепций нет до сих пор.
Races in the Struggle for Life” («Происхождение
Вместе с тем уже в 60-е годы прошлого столе-
видов путем естественного отбора, или сохране-
тия появились представления о плотностно- и ча-
ние благоприятных рас в борьбе за жизнь») [2], а
стотно-зависимых составляющих в действии
затем использует ее в качестве одного из обосно-
естественного отбора [11, 12], а также о плотност-
ваний своей концепции естественного отбора.
но-независимых формах естественного отбора,
Создатели синтетической «генетической» теории
действие которых в условиях экологического ли-
949
950
ФРИСМАН и др.
митирования способно вызвать изменение харак-
родных популяциях происходит в процессе их
тера динамики численности популяций [13-16].
эволюции под действием естественного отбора, а
Работы в рамках эколого-генетического направ-
точнее, под действием плотностно-независимого
ления моделирования, рассматривающего взаи-
естественного отбора, повышающего среднюю
мосвязанные изменения численности и генети-
приспособленность популяции в соответствии с
ческой структуры популяции, позволили охарак-
фундаментальной теоремой естественного отбора
теризовать действие эволюционных факторов, в
Р. Фишера. Если это так, то в процессе естествен-
первую очередь естественного отбора, на измене-
ной эволюции природной популяции с выражен-
ние генетической структуры, а соответственно и
ной сезонностью жизненного цикла должен про-
изменений характера динамики популяций, ко-
исходить закономерный переход от равновесных
торые сталкиваются с естественным ограничени-
режимов динамики численности к колебаниям и
ем экологических ресурсов (т.е. находящихся под
хаосу (псевдостохастическому поведению).
действием лимитирующих факторов окружаю-
В данной работе мы на простом модельном при-
щей среды) [17-22].
мере рассмотрим, как эволюционный процесс есте-
Даже простейшие модели лимитированных
ственного отбора по приспособленностям особей
популяций, построенные на классических для
может привести к изменению параметров роста по-
математической биологии уравнениях Рикера,
пуляции и связанной с этим смене динамических
Смита-Фейгенбаума, Мэя-Хассела, демонстри-
режимов ее численности. Обсудим возможные ме-
руют удивительно сложную динамику [23, 24].
ханизмы и направленность этих изменений.
При этом важно учитывать, что большинство
биологических популяций имеет ярко выражен-
ХАРАКТЕР ДИНАМИКИ ЧИСЛЕННОСТИ
ную неоднородность, представленную возраст-
ПОПУЛЯЦИЙ
ной или стадийной структурой. В популяцион-
С НЕПЕРЕКРЫВАЮЩИМИСЯ
ной экологии для описания динамики структури-
ПОКОЛЕНИЯМИ
рованных популяций широко используются
матричные модели Лесли-Лефковича [25-28].
Для однолетних растений, многих видов насе-
Все эффекты, связанные с плотностно-зависи-
комых, некоторых видов рыб, земноводных и
мой регуляцией роста популяции и наблюдающи-
пресмыкающихся каждая отдельная популяция
еся в одномерных моделях неструктурированных
представляет собой один возрастной класс, и
популяций, отмечаются и в системах динамики
смежные поколения такой популяции не пере-
структурированных популяций. Наиболее иссле-
крываются. Если условия среды меняются от по-
дованными моделями, демонстрирующими раз-
коления к поколению не очень сильно, то чис-
нообразные типы динамического поведения,
ленность некоторого поколения будет опреде-
являются двумерные системы рекуррентных
ляться только численностью предыдущего
уравнений или двумерные отображения, преиму-
поколения. Обозначив через Nn численность п-го
щественно описывающие динамику двухвозраст-
поколения, можно записать следующее детер-
ной популяции [29]. Такая структура характерна
министическое уравнение, описывающее дина-
для организмов с небольшим временем жизни,
мику численности такой одновозрастной попу-
включающим два-три периода размножения.
ляции: Nn+1=F(Nn). Простейший вид этого урав-
Примерами могут служить мелкие млекопитаю-
нения для F(Nn) = rNn фактически является
щие (мышевидные грызуны, беличьи и т.п.),
моделью Мальтуса, где r — некоторая постоян-
быстросозревающие рыбы (такие как корюшка,
ная (т.е. каждая особь оставляет в следующем по-
навага и др.), многие насекомые, двух- и трехлет-
колении r потомков независимо от численно-
ние растения [30-33]. Именно мышевидные гры-
сти родительской популяции):
зуны, быстросозревающие виды рыб и насекомые
обладают сложной флуктуирующей динамикой и
Nn+1 = rNn.
(1)
являются наиболее частыми объектами исследо-
ваний в экспериментальной и яркими примерами
Решение этого уравнения представляет собой
в теоретической популяционной биологии.
геометрическую прогрессию со знаменателем r и
начальным членом N0, что фактически идентич-
В теоретических исследованиях двухвозраст-
но экспоненциальному росту численности по-
ных популяций неоднократно было показано, что
пуляции в отсутствие лимитирующих факторов.
увеличение репродуктивного потенциала и пара-
метров, характеризующих выживаемость особей,
Хорошо известно, что достаточно долгий экс-
проводит к усложнению динамики численности
поненциальный рост численности в природе ни-
от стабильных режимов до нерегулярных колеба-
когда не наблюдается. Рано или поздно сказыва-
ний [34-36]. Однако механизмы роста репродук-
ется действие лимитирующих факторов, поэто-
тивного потенциала, как правило, не рассматри-
му коэффициент r в уравнении (1) оказывается
вались. Вместе с тем можно предположить, что
функцией численности. Положим r = af(N), где
рост потенциальной плодовитости особей в при-
f(N) - функция, описывающая лимитирование,
БИОФИЗИКА том 65
№ 5
2020
ЭКОЛОГО-ГЕНЕТИЧЕСКИЕ МОДЕЛИ
951
и а - параметр, называемый репродуктивным
потенциалом популяции и характеризующий
скорость роста популяции в пустоту (т.е. f(N) вы-
бирается так, чтобы выполнялось f(0) = 1). Те-
перь вместо уравнения (1) получаем:
Nn+1 = a Nn f(Nn).
(2)
Именно такие модели исследовали А.П. Ша-
пиро [23] и Р. Мэй [24]. Они показали, что дина-
мика численности популяции, описываемая
уравнением (2), может быть весьма сложной, если
функция F(N) = aNf(N) убывает при больших N
достаточно быстро (например, быстрее, чем
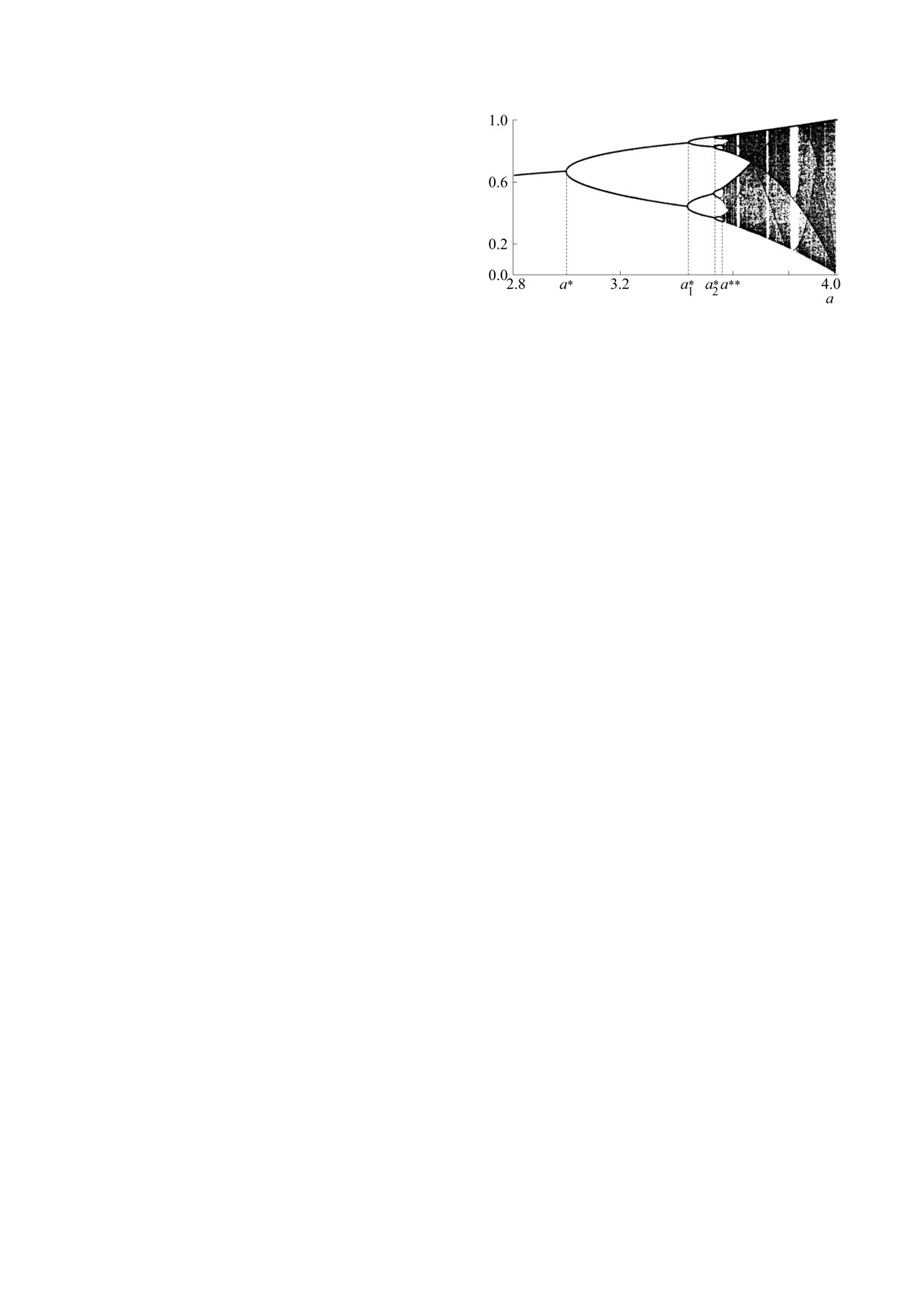
Рис. 1. Бифуркационная диаграмма, характеризую-
щая предельные траектории уравнения xn+1 = axn(1 -
1/N2). Рассмотрим кратко характер этой динами-
- xn) в зависимости от величины коэффициента a.
ки.
Пусть N(a) - нетривиальное стационарное
значение численности, т.е. решение уравнения
характер и становится хаотическим. Однако зоны
N = F(N). При малых значениях коэффициента а
хаотического поведения численности перемежа-
(1 < а < а0; F'(N(а0)) = 0; 0 < F'(N(a)) < 1) каждое
ются с «окнами» периодического, т.е. регулярно-
решение уравнения (2) ведет себя аналогично ре-
го поведения. Отличить эти режимы можно, рас-
шению соответствующего дифференциального
считав значение показателя Ляпунова; при
уравнения - его можно изобразить последова-
этом регулярной динамике соответствует отри-
тельностью точек (n, Ν(n)), лежащих на S-образ-
цательный показатель, квазипериодическим ре-
ной кривой в плоскости {n, Ν}. Каждая из таких
жимам - нулевой, а хаосу - положительный [39].
последовательностей монотонно сходится к рав-
Конкретизируя вид функции f(N), для уравне-
новесному значению численности N(а). Однако
ния (2) численно можно построить бифуркаци-
при больших значениях коэффициента а динами-
онную диаграмму, характеризующую предельные
ческая картина существенно меняется. Так, при
траектории в зависимости от величины коэффи-
а0 < а < a*(F'(N(а*) = -1) решения уравнения (2)
циента а (рис. 1). Хорошо изучены дискретный
сходятся к равновесию N(a) уже не монотонно, а
аналог модели Ферхюльста, для которого f(N) =
в виде затухающих колебаний.
= 1 - kN, и модель, предложенная канадским их-
тиологом У. Рикером, для которой f(N) = exp(-
При а > а* (но а < а**, см. ниже) решения
kN). При исследовании уравнения (2) обычно ис-
уравнения (2) уже не сходятся к N(a) (если на-
ключают масштабный параметр k и переходят к
чальное значение не совпадало с Ν(a)), а дают
безразмерным переменным:
«относительным»
картину устойчивых незатухающих колебаний.
численностям x = kN. В этом случае дискретный
Потеря устойчивости равновесной точки N(a)
аналог модели Ферхюльста имеет вид: xn+1 =
при a = а* сопровождается возникновением пре-
= axn(1 - xn), а модель Рикера: xn+1 = a xnexp(-xn).
дельного цикла длины 2, который представляет
собой единственное устойчивое решение уравне-
ния (2) при а* < а < а1*.
F-ОТБОР В ПОПУЛЯЦИИ
При а1* < а < а2* решения уравнения (2) схо-
С НЕПЕРЕКРЫВАЮЩИМИСЯ
дятся к устойчивому предельному циклу длины 4,
ПОКОЛЕНИЯМИ. ИЗМЕНЕНИЕ ХАРАКТЕРА
при а2* < а < а3* - к предельному циклу длины 8
ДИНАМИКИ ЧИСЛЕННОСТИ В ПРОЦЕССЕ
и т.д. Последовательность значений параметра a
ЭВОЛЮЦИИ ЛИМИТИРОВАННОЙ
{ak*}, при которых происходит удвоение периода,
ПОПУЛЯЦИИ
удовлетворяет закону М. Фейгенбаума [37, 38].
Мы рассматривали характер динамики чис-
ленности при фиксированных значениях пара-
Описанные изменения характера динамики
численности, связанные с изменением параметра
метров. Однако эволюционный процесс и изме-
модели и заключающиеся в возникновении серии
нения в окружающей среде могут привести к из-
устойчивых циклических траекторий с длинами
менению параметров конкретной популяции.
Обсудим возможные механизмы и направлен-
циклов, равными целым степеням числа 2, при-
ность этих изменений.
нято называть первой серией бифуркаций. Эта
серия заканчивается при а = а**, называемой
Одной из первых задач математической попу-
«точкой накопления». Если параметр а превосхо-
ляционной генетики, рассматриваемой Р. Фише-
дит точку накопления a**, то появляются области
ром [3], С. Райтом [4] и Дж. Холдейном [5], ока-
его значений, в которых поведение численности
зался количественный анализ результатов дей-
популяции теряет сколько-нибудь регулярный
ствия отбора в изолированной популяции
БИОФИЗИКА том 65
№ 5
2020
952
ФРИСМАН и др.
диплоидных организмов. Предполагалось, что
2
2
w
n
=w
11
q
n
+
2w
12
q
n
(1−q
n
)
+w
22
(1−q
n
)
популяция не лимитирована внешними ресурса-
ми, и динамика ее численности не рассматрива-
– средняя приспособленность популяции.
лась. Мы попытались перенести полученные в
ходе решения той задачи результаты на лимити-
Пусть действие отбора не зависит от численно-
сти (плотности населения) популяции, однако бу-
рованные популяции и неожиданно обнаружили,
что чисто количественные изменения генетиче-
дем учитывать влияние плотностно-зависимых
ской структуры приводят к существенным каче-
факторов, лимитирующих рост популяции. Такое
ственным изменениям динамики их численно-
представление, по-видимому, вполне соответству-
сти.
ет реальному действию отбора на те признаки, при-
родная генотипическая изменчивость по которым
Для понимания основных закономерностей эво-
никак не коррелирована с изменениями плотности
люции динамического поведения численности ли-
в популяции. Подобным образом должны, по-ви-
митированной популяции ограничимся подроб-
димому, отбираться многие физиологические мута-
ным рассмотрением простой модельной ситуации,
ции, влияющие на общую жизнеспособность.
когда все адаптивное разнообразие в популяции
Можно привести опубликованные данные о плот-
определяется одним диаллельным A локусом с ал-
ностно-независимом поведении генетической
леломорфами A1 и A2, причем фенотип особи жест-
структуры популяции. Например, Л.Д. Готлибом
ко определяется ее генотипом; популяция панмик-
показано отсутствие какой-либо корреляции меж-
тична, в ней действуют менделевские правила на-
ду изменениями численности и динамикой частот
следования, а смежные поколения особей не
аллелей для трех (из пяти изученных) полиморф-
перекрываются. В этом случае действие отбора
ных белковых локусов в четырех сильно флукту-
можно описать количественно, поставив в соответ-
ирующих популяциях степных полевок Micro-
ствие каждому из генотипических классов особей
tus ochrogaster [13], М.С. Грайнесом обнаружена
A1A1, A1A2, и A2A2 по одному коэффициенту w11, w12
стабильность генетической структуры по пяти
и w22 соответственно, называемому приспособлен-
полиморфным ферментным локусам в геогра-
ностью (fitness) особей данного генотипа. Будем
фически изолированной популяции однолетнего
считать, что wij равно среднему числу потомков,
растения Stephanomeria exigua (Compositae) [42].
произведенных одной особью данного генотипиче-
Аллельные частоты этих локусов практически не
ского класса и доживших до репродуктивного воз-
изменились в течение четырех последовательных
раста (давших вклад в следующее поколение). Это
поколений несмотря на то, что численность по-
определение приспособленности формально экви-
пуляции испытывала в этот период значитель-
валентно определению Р. Фишера [3, 40].
ные изменения.
Несмотря на заведомое максимальное упро-
Это означает, что относительные приспособ-
щение, моногенная модель отбора не является
ленности (отношения wij/w*, где wij - приспособ-
полностью оторванной от реальности. В качестве
ленность ij-го генотипа, a w* - приспособлен-
яркого примера можно привести моногенный ха-
ность одного из генотипов, принятая за эталон)
рактер наследования размера помета у песцов
не зависят от величины численности популяции,
(Alopex lagopus). В работе [41] на основе комплекс-
т.е. их можно считать константами. Если популя-
ного сегрегационного анализа типа наследования
ция находится под действием плотностно-зави-
размера приплода в расширенной генеалогии
симых лимитирующих факторов, то абсолютные
фермерских песцов было показано, что данный
приспособленности каждого генотипического
заведомо адаптивный (напрямую определяющий
класса должны зависеть от численности популя-
приспособленность) признак является аутосом-
ции:
ным признаком, а его наследование можно опи-
сать в рамках моногенной модели с контролем
(4)
w
ij
(N
)
=a
ij
f (N ),
малого размера приплода по рецессивному типу.
Перейдем теперь к математическому описа-
где f(N) - функция, характеризующая плотност-
нию процесса эволюции популяции. Введем сле-
ную регуляцию численности, одинаковую для
дующие обозначения: q - частота гамет, несущих
каждого генотипа, аij - коэффициенты, равные
аллель A1; N - численность популяции. Сделан-
относительным генотипическим приспособлен-
ные предположения позволяют получить рекур-
ностям. Функцию f(N) мы будем считать моно-
рентные уравнения, связывающие значения этих
тонно убывающей с ростом численности, причем
переменных в смежных поколениях [40]:
f(0) = 1. Такое действие отбора мы предлагаем на-
зывать F-отбором, подчеркивая этим тот факт,
N
=
w
n
N
n+1
n
что отбор осуществляется при постоянных отно-
,
(3)
сительных приспособленностях (fitnesses). Легко
q
n+1
=
q
n
(w
11
q
n
+
w
12
(1−
q
n
)) / wn
видеть, что в случае F-отбора уравнения динами-
где n - номер поколения,
ки (3) преобразуются к виду:
БИОФИЗИКА том 65
№ 5
2020
ЭКОЛОГО-ГЕНЕТИЧЕСКИЕ МОДЕЛИ
953
N
=
a
N f(N
)
что аn зависит от n, это связано с тем, что аn равно
n+1
n n
n
(5)
среднему значению относительных приспособ-
q
=
q
(a
q
+
a
(1-
q
)) /
a
n+1
n
11
n
12
n
n
ленностей (аij) в популяции в n-м поколении, т.е.
Уравнение для динамики численности (N)
2
2
a
=a
q
+
2a
q
(1−q
)
+a
(1−q
)
Заметим,
рассматриваемой популяции, находящейся под
n
11
n
12
n
n
22
n
действием F-отбора, фактически может описы-
что без потери общности модель (5) может быть
ваться моделями Рикера (f(N) = exp(-bN)), Ферх-
записана в относительных значениях численно-
юльста (f(N) = 1 - kN) и др. Разница в том лишь,
сти:
х
n+1
=
a
n n
х f(х
n
)
2
2
,
(5*)
q
=
q
(a
q
+
a
(1−
q
)) /
a
q
+
2
a
q
(1−
q
)
+
a
(1−
q
)
(
)
1
11
12
11
12
22
n
n
n
n
n
n
n
n
+
при этом в случае дискретного аналога модели
подробнее на новых эффектах, связанных с нали-
Ферхюльста f(xn) = 1 - xn, а в случае модели Рике-
чием плотностного лимитирования роста чис-
ра - f(xn) = exp(-xn).
ленности популяции.
Уравнения, характеризующие изменение ге-
При движущем типе отбора относительная
нетической структуры популяции, при F-отборе
приспособленность гетерозиготы занимает про-
не зависят от ее численности и, следовательно,
межуточное положение между приспособленно-
могут быть исследованы отдельно. Кроме того,
стями гомозигот (min{a11, a22} < a12 < max{a11,
они совпадают с точностью до обозначений с
a22}). В этом случае в ходе эволюции популяции
уравнением динамики генетической структуры
происходит вытеснение неоптимальных аллелей
нелимитированной популяции (3). Следователь-
и средняя относительная приспособленность по-
но, здесь справедливо утверждение теоремы Фи-
пуляции монотонно увеличивается, стремясь к
шера, доказанное для дискретного случая Кинг-
наибольшей приспособленности гомозигот. При
маном [43]: ап+1 ≥ ап, т.е. среднее значение отно-
этом, пока не произойдет полного вытеснения
сительных приспособленностей аi может лишь
одного из аллелей, в популяции будет наблюдать-
возрастать в процессе эволюции, независимо от
ся полиморфизм (т.е. будут присутствовать все
их конкретных значений и начального состояния
три генотипа), но этот полиморфизм будет «пере-
популяции. Это означает, что репродуктивный
ходящим» - он исчезнет с фиксацией аллеля,
потенциал лимитированной популяции возрас-
обеспечивающего большую относительную при-
тает в процессе эволюции так же, как средняя
способленность. Действительно, в случае такого
приспособленность свободной. Однако такой
отбора система имеет только одно устойчивое мо-
монотонный рост репродуктивного потенциала,
номорфное состояние, в котором и должна ока-
вызванный динамикой генетической структуры,
заться популяция вне зависимости от начальных
может привести к изменению характера динами-
условий. Однако при этом может существенно
ческого поведения численности даже однород-
поменяться динамическое поведение численно-
ной популяции и обусловить возникновение ко-
сти популяций. Пока значение средней приспо-
лебаний и нерегулярной динамики численности.
собленности невелико численность популяции
Таким образом, в лимитированных популяциях
будет стремиться к устойчивому равновесию, со-
с неперекрывающимися поколениями прогрес-
ответствующему этому значению. Однако если
сивное возрастание средней приспособленности
значение средней приспособленности перейдет
может оказаться в диссонансе со стабильностью
через некоторое бифуркационное значение, то
роста популяции. Этот факт находится в явном
это может привести к возникновению колебаний
противоречии с интуитивным представлением об
численности, а при дальнейшем росте средней при-
увеличении стабильности популяции с ростом ее
способленности - и к хаотическим режимам дина-
средней приспособленности.
мики численности мономорфной популяции
(рис. 2). Отметим, что для идентификации типа ди-
намического режима были использованы показате-
ТИПЫ ОТБОРА В ЛИМИТИРОВАННЫХ
ли Ляпунова (L1, L2); в частности, регулярная дина-
ПОПУЛЯЦИЯХ
мика представлена на рис. 2г (оба показателя отри-
По аналогии с классической моделью нелими-
цательны: L1
= -0.08, L2
= -0.56), хаосу
тированной популяции для модели (5) также
соответствует один положительный показатель
можно выделить три типа естественного отбора в
(рис.
2а (L1 = 0.494, L2
= -0.002), рис.
2б
зависимости от соотношения репродуктивных
(L1 = 0.497, L2 = -0.296), рис. 2в (L1 = 0.494,
потенциалов генотипов на основе анализа усло-
L2 = -0.72), рис. 2д (L1 = 0.362, L2 = -0.588));
вий существования и устойчивости неподвижных
точек рассматриваемых моделей. Остановимся
квазипериодическим режимам - один нулевой и
БИОФИЗИКА том 65
№ 5
2020
954
ФРИСМАН и др.
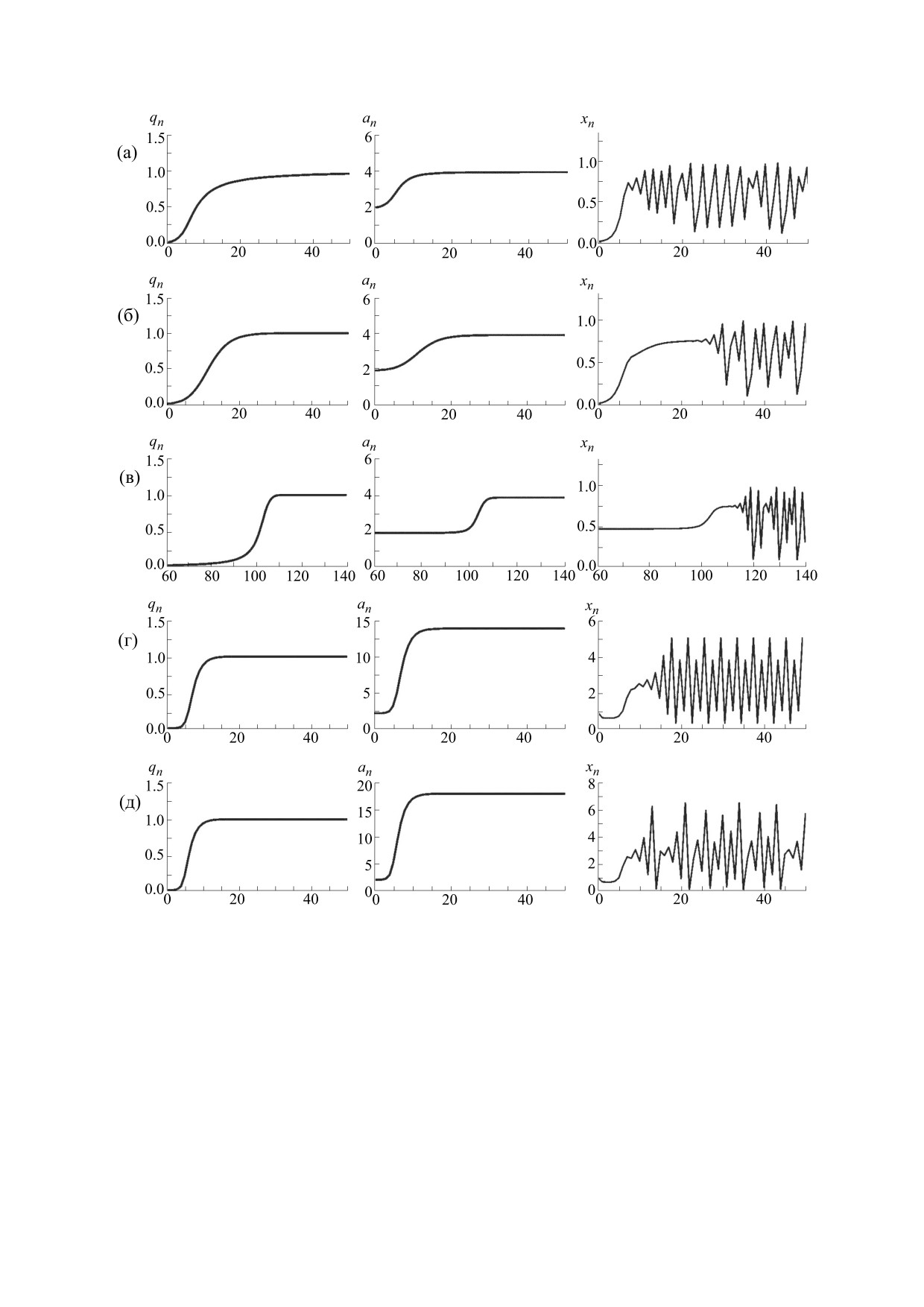
Рис. 2. Динамика частоты аллеля A1 (q), средней приспособленности (а) и относительной численности популяции (х),
полученная по модели (5*). Для панелей (а), (б) и (в): f(x) = 1 - x, a11 = 3.9, a22 = 1.9, q0 = 0.01, х0 = 0.01. Панель (а) - аллель
A1 доминантен, a12 = 3.9; панель (б) - аллель A1 кодоминантен, a12 = 2.9; панель (в) -аллель A1 рецессивен, a12 = 1.9.
Панель (г): f(x) = exp(-x), a11 = 14, a12 = 8, a22 = 2, q0 = 0.001, х0 = 1. Панель (д): f(x) = exp(-x), a11 = 18, a12 = 10, a22 = 2,
q0 = 0.001, х0 = 1.
один отрицательный показатель, а гиперхаосу -
фиксации этого аллеля, быстрому росту средней
два положительных показателя [44].
приспособленности и быстрому переходу чис-
ленности популяции от стабильного режима к не-
Как видно на рис. 2а, появление нового доми-
нантного аллеля, обеспечивающего большую
регулярным колебаниям. Если появившийся ал-
приспособленность гомозиготы (и соответствен-
лель доминирует не полностью (гетерозигота
но, гетерозиготы), приводит к весьма быстрой
имеет приспособленность промежуточную между
БИОФИЗИКА том 65
№ 5
2020
ЭКОЛОГО-ГЕНЕТИЧЕСКИЕ МОДЕЛИ
955
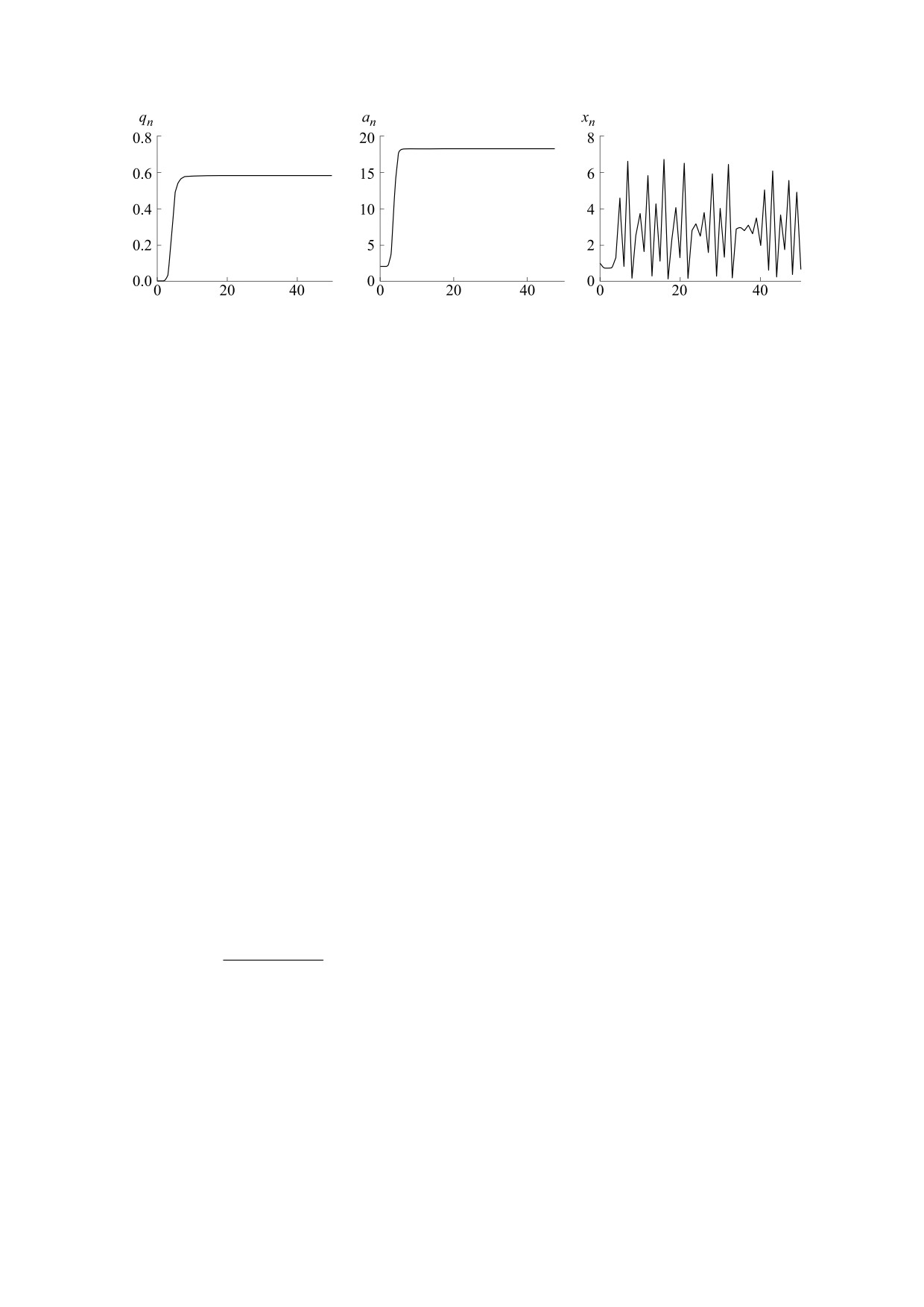
Рис. 3. Динамика частоты аллеля A1 (q), средней приспособленности (а) и относительной численности популяции (х),
полученная по модели (5*) при сверхдоминировании: f(x) = exp(-x), a11 = 10, a12 = 30, a22 = 2, q0 = 0,001, х0 = 1.
приспособленностями гомозигот), эти процессы
ности в предельных режимах всегда присутствуют
идут заметно медленнее (рис 2б). Они идут совсем
все три генотипа в стабильных соотношениях.
медленно, если появившийся аллель оказывается
При пониженной приспособленности гетерози-
рецессивным: рост средней приспособленности и
готы (а12 < min{а11, а22}) полиморфное равновес-
равновесного значения численности начинает
ное значение частот аллей q*, определяемое вы-
проявляться где-то к сотому поколению (рис. 2в).
ражением (6), существует, но оказывается не-
По-видимому, реальная ситуация роста средней
устойчивым. Популяция стремится к одному из
приспособленности оказывается еще более мед-
двух мономорфных состояний (q = 0 или q = 1),
ленная и существенно менее регулярная. Редко
причем к какому именно - определяется началь-
возникающие положительные мутации могут
ными условиями. Если в начальный момент вы-
обеспечивать очень небольшую прибавку при-
полнено соотношение q0 < q*, то будет вытес-
способленности, и, следовательно, небольшое
няться аллель A1 и в итоге популяция окажется мо-
медленное увеличение равновесной численности
популяции. Однако даже такой медленный рост
номорфной по генотипу A2A2 (q = 0ì), если q0 > q*,
средней приспособленности вполне может при-
то будет вытесняться аллель A2 и в итоге популя-
вести к тому, что она превзойдет некоторое би-
ция окажется мономорфной по генотипу A1A1. вне
фуркационное значение и равновесная числен-
зависимости от соотношения величин приспо-
ность популяции окажется неустойчивой: воз-
собленностей a11 и a22 (рис. 4).
никнут популяционные колебания. Дальнейший
рост приспособленности приведет к их усложне-
Таким образом, здесь возникает эффект, кото-
нию и, возможно, скатыванию в хаотический ре-
рый можно назвать «ловушкой бистабильности».
Этот эффект возникает только в случае, когда
жим динамики. Заметим, что, как видно на
приспособленность гетерозиготы меньше при-
рис. 2г,д, последовательность описываемых про-
цессов принципиально не зависит от конкретной
способленности каждой из гомозигот и заключа-
модели плотностно-зависимого лимитирования
ется в том, что более перспективная форма не мо-
роста численности популяции.
жет естественным образом вытеснить явно более
слабого (по приспособленности) генетического
Сверхдоминирование или повышенная приспо-
конкурента.
собленность гетерозигот (а12 > max{а11, а22}) все-
Дело в том, что при низких значениях репро-
гда приводит к установлению сбалансированного
дуктивного потенциала гетерозиготы популяция
полиморфизма: частота аллеля A1 стремится к не-
находится в режиме бистабильности, когда оба
тривиальному равновесному значению
мономорфных состояния устойчивы и характе-
a
−
a
ризуются своими областями притяжений. Если в
21
22
q*
=
,
(6)
некоторый «начальный» момент времени в моно-
2a
−
a
−
a
21
11
22
морфной популяции появляется новый аллель,
и в популяции всегда присутствуют все три гено-
обеспечивающий большую приспособленность
типа. При этом в популяции за счет отбора проис-
гомозиготы (но с пониженной приспособленно-
ходит монотонный рост средней приспособлен-
стью гетерозиготы), то этот аллель никак не смо-
ности, а численность сначала стремится к более
жет закрепиться в популяции. Действительно,
высокому значению равновесия, а потом, при
начальная концентрация нового аллеля (мутанта
переходе средней приспособленности через би-
или мигранта) не может быть высокой, она близ-
фуркационное значение, начинает колебаться
ка нулю и заведомо меньше q*. Именно поэтому
(рис. 3). Подчеркнем, что в случае свехдоминиро-
текущие значения частот оказываются в бассейне
вания в популяции независимо от уровня числен-
притяжения исходного мономорфного равнове-
БИОФИЗИКА том 65
№ 5
2020
956
ФРИСМАН и др.
численность репродуктивной части популяции.
Период размножения заканчивается появлением
новорожденных особей следующего поколения.
Будем предполагать, что времени, протекающего
между двумя последовательными периодами раз-
множения, достаточно для развития особей млад-
шего возраста до половозрелого состояния, а но-
ворожденных (или личинок) до состояния млад-
шего возраста. Допустим, что выживаемость и
репродуктивная способность половозрелых осо-
бей не зависит от их хронологического возраста.
Это правомерно для организмов с небольшим
временем жизни, включающим два-три периода
размножения, как у многих насекомых, рыб, мел-
Рис. 4. Изменение частоты аллеля А1 в модели (3) при
ких млекопитающих, двух- и трехлетних расте-
пониженной относительной приспособленности гете-
розигот и различных начальных условиях.
ний и др. Можно записать следующую систему
уравнений, описывающую относительные чис-
ленности рассматриваемых возрастных групп в
сия, и даже большое значение приспособленно-
смежных поколениях:
сти гомозиготы по новому аллелю не способно
X
=
A(X
,Y
)Y
привести к смещению частот в бассейн притяже-
n+1
n n n
,
(7)
ния другого мономорфного равновесия с боль-
Y
=
B
X
,Y
X
+C
X
,Y
Y
n+1
(
n n
)
n
(
n n n
)
шой приспособленностью особей. Как результат,
где A - рождаемость, B и С - выживаемости млад-
изменение режима динамики численности также
шего и старшего возрастного классов соответ-
не происходит: популяция остается стабильной
ственно.
по численности и мономорфной по генотипу с
низким значением приспособленности.
Регуляция роста численности популяции пу-
тем плотностно-зависимого лимитирования
Для выхода из описанной ловушки необходи-
рождаемости наблюдается у многих животных,
мо внешнее воздействие на популяцию. Падение
особенно у мелких млекопитающих, когда рожда-
численности популяции и связанные с этим слу-
емость заметно снижается по мере увеличения
чайные процессы (известные, как «прохождение
численности популяции. Основным проявлени-
через бутылочное горлышко») могут привести к
ем плотностно-зависимой регуляции рождаемо-
флуктуации генетического состава и «перескоку»
сти является стресс-синдром, приводящий к сни-
популяции из одного бассейна притяжения в дру-
жению половой активности и уменьшению пло-
гой (т.е. к случайному увеличению текущей ча-
довитости особей, вплоть до рассасывания части
стоты нового аллеля до величины, превышающей
заложенных эмбрионов. В частности, это харак-
q*). А это уже приведет к прогрессивному росту
терно для видов, подверженных сильным колеба-
средней относительной приспособленности, к
ниям численности, например леммингов, поле-
росту численности, к росту равновесных значе-
вок и др. [30]. При моделировании динамики по-
ний численности и, возможно, к возникновению
пуляций таких видов выживаемости можно
колебательных режимов при большой приспо-
считать константами B(X,Y) = b, C(X,Y) = c, а рож-
собленности.
даемость, по аналогии с моделью Рикера, можно
Далее рассмотрим более сложные нелинейные
записать в виде: A(X,Y) = r⋅exp(-αX-βY), где r -
модели динамики популяций с возрастной
репродуктивный потенциал, α и β - коэффици-
структурой и покажем, как влияет наличие воз-
енты, характеризующие интенсивности воздей-
растной структуры или стадийности развития на
ствия численностей (плотностей) неполовозре-
результаты действия естественного отбора.
лого и половозрелого возрастного класса на
уменьшение рождаемости. С учетом этого и после
ДИНАМИКА ЧИСЛЕННОСТИ ПОПУЛЯЦИИ
перехода к безразмерным переменным - относи-
С ДВУМЯ СТАДИЯМИ РАЗВИТИЯ
тельным значениям численностей x = bβX и
y = βY - модель (7) может быть преобразована к
Рассмотрим популяцию с возрастной структу-
виду:
рой, которая к началу очередного сезона размно-
жения может быть представлена совокупностью
x
=
r ⋅ y
⋅
exp(−ρ ⋅
x
−
y
n+1
n
n
n
двух возрастных классов: младшего, включающе-
),
(8)
y
n+1
=
x
n
+
c⋅ y
n
го неполовозрелых особей, и старшего, состоя-
щего из особей, участвующих в размножении.
где параметр ρ = α/(bβ) характеризует относи-
Обозначим через Xn численность младшего воз-
тельный вклад младшей возрастной группы в ли-
растного класса в n-й сезон размножения, а Yn -
митирование воспроизводства.
БИОФИЗИКА том 65
№ 5
2020
ЭКОЛОГО-ГЕНЕТИЧЕСКИЕ МОДЕЛИ
957
В работах [34, 35] проведено детальное иссле-
Рождаемость и выживаемость особей старшей
дование динамики численности двухвозрастной
возрастной группы в (7) будем считать константа-
популяции, описываемой моделью (8), где пока-
ми: A(X,Y) = r и C(X,Y) = c. Перейдем далее от аб-
зано, что падение рождаемости с ростом числен-
солютных численностей возрастных классов к от-
ности особей оказывается эффективным меха-
носительным (x = kX, y = kY) и получим следую-
низмом регуляции роста численности. Однако
щую систему уравнений, описывающую
при больших потенциальных репродуктивных
относительные численности рассматриваемых
возможностях и большой выживаемости особей
возрастных групп в смежных поколениях:
падение рождаемости с ростом численности осо-
x
n+1
=
ry
n
бей может привести к потере устойчивости и воз-
,
(9)
никновению колебаний численности, имеющих
y
=
x
(1−
x
)
+
cy
n+1
n
n
n
весьма сложную временную организацию.
где r - репродуктивный потенциал особей стар-
В частности показано, что при относительно
шей возрастной группы, а c - их выживаемость на
больших значениях параметра ρ (ρ ≥ 1), т.е. в слу-
последующих годах жизни.
чае, когда вклад в лимитирование рождаемости
осуществляется преимущественно младшим воз-
F-ОТБОР В ПОПУЛЯЦИИ С ДВУМЯ
растным классом, потеря устойчивости, как и в
СТАДИЯМИ РАЗВИТИЯ
исходной одномерной модели Рикера, сопровож-
дается появлением двухгодичных колебаний и их
В случае, когда репродуктивный потенциал r
последующими бифуркациями по сценарию
определяется генетически одним адаптивным
Фейгенбаума. При этом чем больше ρ, тем при
аутосомным диаллельным локусом, уравнения
более низком значении репродуктивного потен-
динамики численностей возрастных групп попу-
циала наблюдаются колебания.
ляции преобразуются очевидным образом и к
ним добавляются уравнения динамики частот ге-
При малых же значениях параметра ρ, т.е. в
нов, характеризующие изменение генетической
случае, когда лимитирование рождаемости осу-
структуры в ходе эволюции двухвозрастной попу-
ществляется в основном численностью взрослых
ляции [36]. При плотностно-зависимом лимити-
половозрелых особей, потеря устойчивости мо-
ровании рождаемости уравнения имеют следую-
жет произойти только при комплексно-сопря-
щий вид [46]:
женных корнях характеристического уравнения,
при переходе |λ| через 1, и сопровождается появ-
x
n+1
=
r
yn n
exp(
−ρx
n
−
y
n
)
лением предельных инвариантных кривых. Соот-
y
n+1
=
x
n
+
cy
n
ветственно уменьшение рождаемости с ростом
p
(r
p
+
r
(1
−
p
))
численности взрослых особей способно привести
n
11
n
12
n
q
=
,
(10)
n
+1
к возникновению достаточно сложно организо-
w
n
ванных колебаний численности. Именно этот ме-
q
x + cp
y
n n
n n
ханизм является, по-видимому, определяющим в
p
n+
1
=
y
n+1
поведении численности мелких млекопитающих,
таких как лемминги и некоторые полевки [45].
а в случае лимитирования выживаемости млад-
шего возрастного класса:
Рассмотрим еще случай, когда регуляция ро-
ста численности популяции осуществляется пу-
x
=
r
y
n
+1
n n
тем плотностно-зависимого лимитирования вы-
y
n
+1
=
x
n
(1
−
x
n
)
+
cy
n
живаемости особей младшего возрастного клас-
p
(r
p
+
r
(
1
−
p
))
са. По аналогии с моделью Ферхюльста будем
n
11
n
12
n
q
=
,
(11)
n
+1
полагать B(X) = 1 - kX. Ситуация, когда выжива-
r
n
емость приплода в большей степени зависит от
x
n
(1
−
x
)
n n
q + cy
n n
p
его собственной численности и практически
p
=
n
+1
(либо совсем) не зависит от величины поло-
y
n+1
возрелой группы, широко распространена в при-
где pn и qn - это частота аллеля А1 в старшем и млад-
роде. Так, у видов с выраженной стадийностью
шем возрастном классах соответственно; парамет-
возрастные группы могут быть разделены гео-
ры r11, r12 и r22 характеризуют репродуктивные
графически и развиваются в разных условиях, не
потенциалы генотипов A1A1, A1A2 и A2A2 старшего
вступая в конкурентные отношения между со-
2
бой. Примером могут служить многие виды рыб,
возрастного класса;
r
=r
p
+
2r
p
(1− p
)
+
n
11
n
12
n
n
которые, отложив икру, мигрируют в другие ме-
2
+ r
(1− p
)
- средний репродуктивный потенциал.
ста, мальки же развиваются отдельно и присо-
22
n
единяются к основному стаду, лишь достигнув
В моделях (10, 11) на основе анализа условий
определенной стадии зрелости.
существования и устойчивости неподвижных то-
БИОФИЗИКА том 65
№ 5
2020
958
ФРИСМАН и др.
Рис. 5. Динамика частот аллеля А1 и численностей возрастных классов, полученных по модели (10) при начальных
условиях q0 = p0 = 0.05, x0 = y0 = 1. Панель (а): ρ=1.2; c = 0.1, r11 = 28, r12 = 15, r22 = 2; панель (б): ρ = 0.2, c=0.1, r11 = 18,
r12 = 10, r22 = 2.
чек также можно выделить три типа естественно-
(квазипериодические режимы) (рис. 5б), не на-
го отбора в зависимости от соотношения репро-
блюдающиеся в моделях популяций без возраст-
дуктивных потенциалов генотипов. Остановимся
ной структуры.
подробнее на новых существенных эффектах,
Сверхдоминирование или повышенный репро-
связанных с наличием возрастной структуры.
дуктивный потенциал гетерозигот (r12 > max{r11,
При движущем типе отбора репродуктивный
r22}) всегда приводит к установлению полимор-
потенциал гетерозиготы занимает промежуточ-
физма, при этом популяция за счет отбора дости-
ное положение относительно репродуктивных
гает большего среднего репродуктивного потен-
потенциалов гомозигот (min{r11, r22}
< r12 <
циала, а численность, как и при движущем отбо-
< max{r11, r22}) и происходит вытеснение неопти-
ре, сначала стремится к более высокому
значению равновесия, а потом начинает коле-
мальных аллелей с возникновением колебаний
баться (рис. 6).
численности при достаточно высоком среднем
репродуктивном потенциале популяции (рис. 5).
Более того, при сверхдоминировании в случае
В частности, при плотностно-зависимой регуля-
лимитирования выживаемости младшего воз-
ции рождаемости и при относительно больших
растного класса, т.е. в модели (11), эволюционное
значениях параметра ρ (ρ ≥ 1) такая потеря устой-
увеличение среднего репродуктивного потенциа-
чивости в модели (10) происходит, как показано
ла может не только вызвать колебания численно-
на рис. 5а, подобно потери устойчивости в моде-
сти, но и привести к возникновению колебаний
ли (5*) и сопровождается появлением двухгодич-
генетического состава.
ных колебаний и их последующими усложнения-
Рассмотрим возможные сценарии динамики
ми по сценарию Фейгенбаума (т.е. подобно дина-
модели (11) при фиксированных значениях пара-
мике, приведенной на рис. 2д). При малых же
метров (c = 0.8, r11 = 1.1, r22 = 1.05) и изменении
значениях параметра ρ, т.е. в случае, когда умень-
параметра r12. Если репродуктивный потенциал
шение рождаемости происходит с ростом числен-
гетерозиготы r12 меняется в интервале от 1.1 до
ности взрослых особей, в ходе эволюции популя-
ции увеличение среднего репродуктивного по-
1.736, то в популяции устанавливается устойчи-
тенциала популяции приводит к бифуркации
вый генетический полиморфизм, а численность
Неймарка-Сакера и возникают достаточно
достигает своего устойчивого равновесия. При
сложно организованные колебания численности
переходе r12 через 1.736 происходит бифуркация
БИОФИЗИКА том 65
№ 5
2020
ЭКОЛОГО-ГЕНЕТИЧЕСКИЕ МОДЕЛИ
959
Рис. 6. Динамика системы (10) в случае сверхдоминирования при различном репродуктивном потенциале
гетерозиготы: ρ = 0.2, c = 0.1, r11 = 10, r12 = 25, r22 = 3, q0 = p0 = 0.05, x0 = y0 = 1.
Неймарка-Сакера (пара комплексно-сопряжен-
них начальных условий популяция приходит к
ных собственных значений становятся больше
устойчивому полиморфному равновесию частот
единицы по модулю) и появляются флуктуации
генов и колебаниям численности, из других - к
численности при постоянном генетическом со-
устойчивым колебаниям как численностей, так и
ставе.
генетического состава (рис. 7).
При дальнейшем увеличении параметра r12,
С ростом r12 наблюдаемый цикл с периодом 7
примерно при r12 = 2.787, происходит новая би-
быстро зашумляется, и динамика всех перемен-
фуркация - появляется еще один устойчивый ре-
ных, начинающихся в бассейне притяжения это-
жим динамики: цикл длины 7 - устойчивые коле-
го цикла, становится нерегулярной. Еще боль-
бания как численности, так и генетического со-
ший рост r12 приводит к мультистабильности.
става с периодом 7, которые сосуществуют с
Помимо имеющихся аттракторов с устойчивым
устойчивым стационарным состоянием генети-
полиморфным равновесием генотипов и нерегу-
ческого состава. Другими словами, при r12 > 2.787
лярной динамикой всех переменных появляется
в модели (11) наблюдается бистабильность: из од-
еще один цикл с периодом 7, сменяющийся цик-
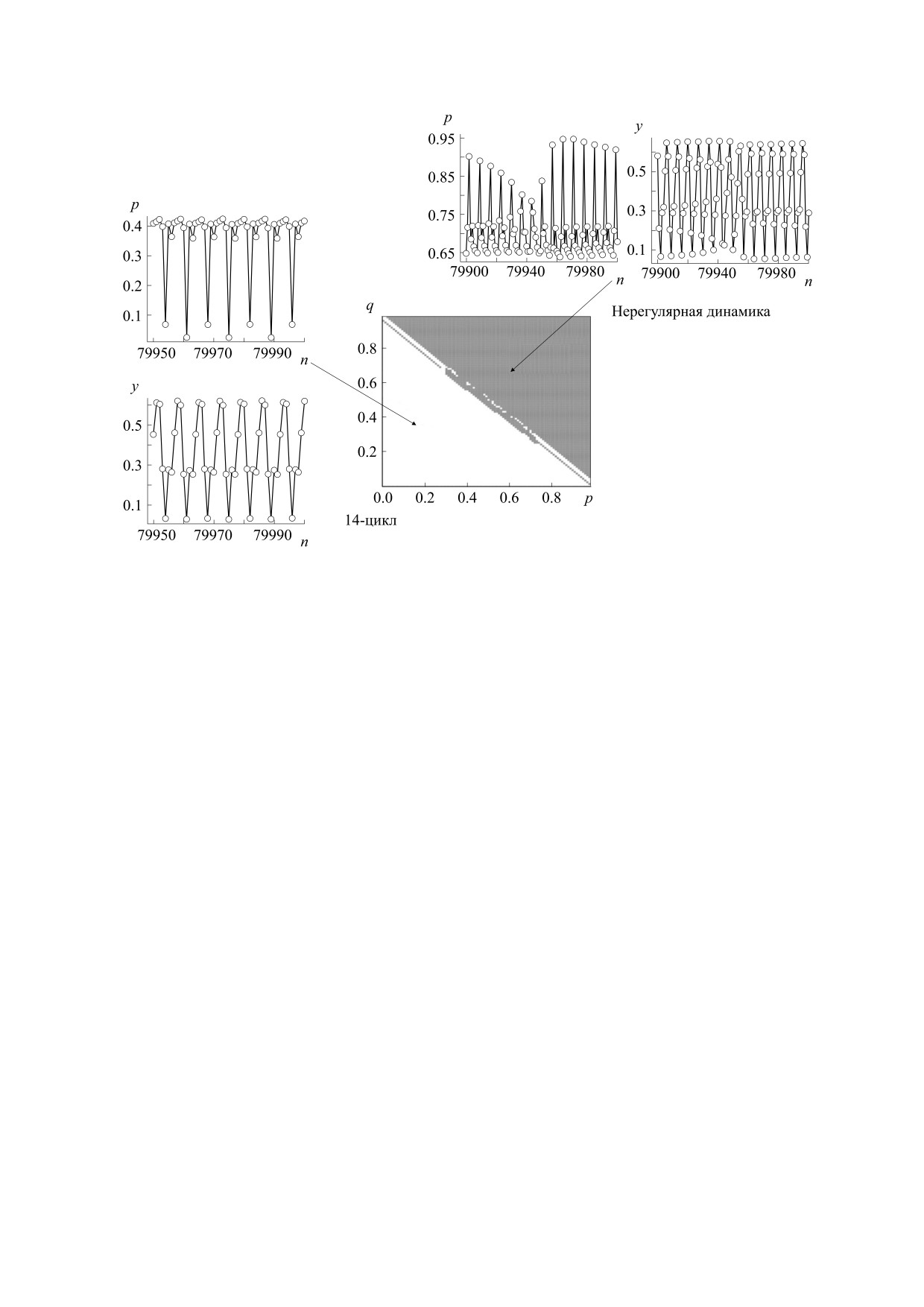
Рис. 7. Два динамических режима (бистабильность) модели (11) при с = 0.8, r11 = 1.1, r22 = 1.05 и r12 = 2.7871. Из одних
начальных условий (бассейн показан темно-серым цветом) популяция приходит к устойчивому полиморфному
равновесию частот генов и колебаниям численности, из других (бассейн показан светло-серым цветом) - к
устойчивым колебаниям, с периодом 7 как для численностей, так и для генетического состава.
БИОФИЗИКА том 65
№ 5
2020
960
ФРИСМАН и др.
Рис. 8. Два динамических режима модели (11) при с = 0.8, r11 = 1.1, r22 = 1.05 и r12 = 2.82. Из одних начальных условий
(бассейн показан белым цветом) популяция приходит к устойчивым колебаниям с периодом 14 как численностей, так
и генетического состава, из других (бассейн показан серым цветом) - к нерегулярным флуктуациям и численностей
и частот аллелей.
лом с периодом 7, и бассейн притяжения этого
Здесь также возникает эффект «ловушки би-
цикла фактически захватывает почти все началь-
стабильности», который заключается в том, что
ные условия, из которых популяция стремилась к
более перспективная форма не может естествен-
устойчивому равновесию генетического состава
ным образом вытеснить явно более слабого по ре-
при флуктуирующей численности (рис. 8). Таким
продуктивным показателям генетического кон-
образом, в зависимости от начальных условий в
курента.
популяции фактически устанавливаются либо
Вместе с тем при пониженной приспособлен-
устойчивые периодические колебания, либо апе-
ности гетерозигот в моделях с возрастной струк-
риодические колебания как численности, так и
турой наряду с выявленной и хорошо понятной
генетического состава.
бистабильностью, связанной с устойчивостью
При пониженной приспособленности гетерози-
двух мономорфных состояний, несколько неожи-
готы (r12 < min{r11, r22}) оба мономорфных состо-
данно обнаруживается еще один - третий устой-
чивый динамический режим. Этот режим оказы-
яния могут быть устойчивы, и система окажется в
вается возможен при низких значениях коэффи-
одном из них в зависимости от начальных усло-
циента выживания взрослых половозрелых
вий. При этом если в популяции без возрастной
особей и заключается в циклических колебаниях
структуры зависимость от начальных условий вы-
генетической структуры, при которых в четные
глядит предельно просто: при q0 < q* она достиг-
годы в популяции преобладает один из аллелей, а
нет равновесия (q = 0), при q0 > q* - q = 1 (рис. 4),
в нечетные годы другой (рис. 10). Наличие такого
то модели структурированных популяций уже
режима означает возможность устойчивой пер-
имеют весьма сложно очерченную область притя-
вичной генетической дивергенции (дифференци-
жения соответствующих мономорфных равнове-
ации) особей разных поколений. Выявленный
сий (рис. 9). Следовательно, результат отбора в
периодический режим реализуется при опреде-
лимитированной популяции зависит не только от
ленных начальных условиях, т.е. имеет свой бас-
начальной частоты рассматриваемого аллеля в
сейн притяжения, и тем более выражен, чем боль-
популяции, но и от численности и ее возрастного
ше изоляция между поколениями, т.е чем меньше
состава.
половозрелых особей выживает. В качестве край-
БИОФИЗИКА том 65
№ 5
2020
ЭКОЛОГО-ГЕНЕТИЧЕСКИЕ МОДЕЛИ
961
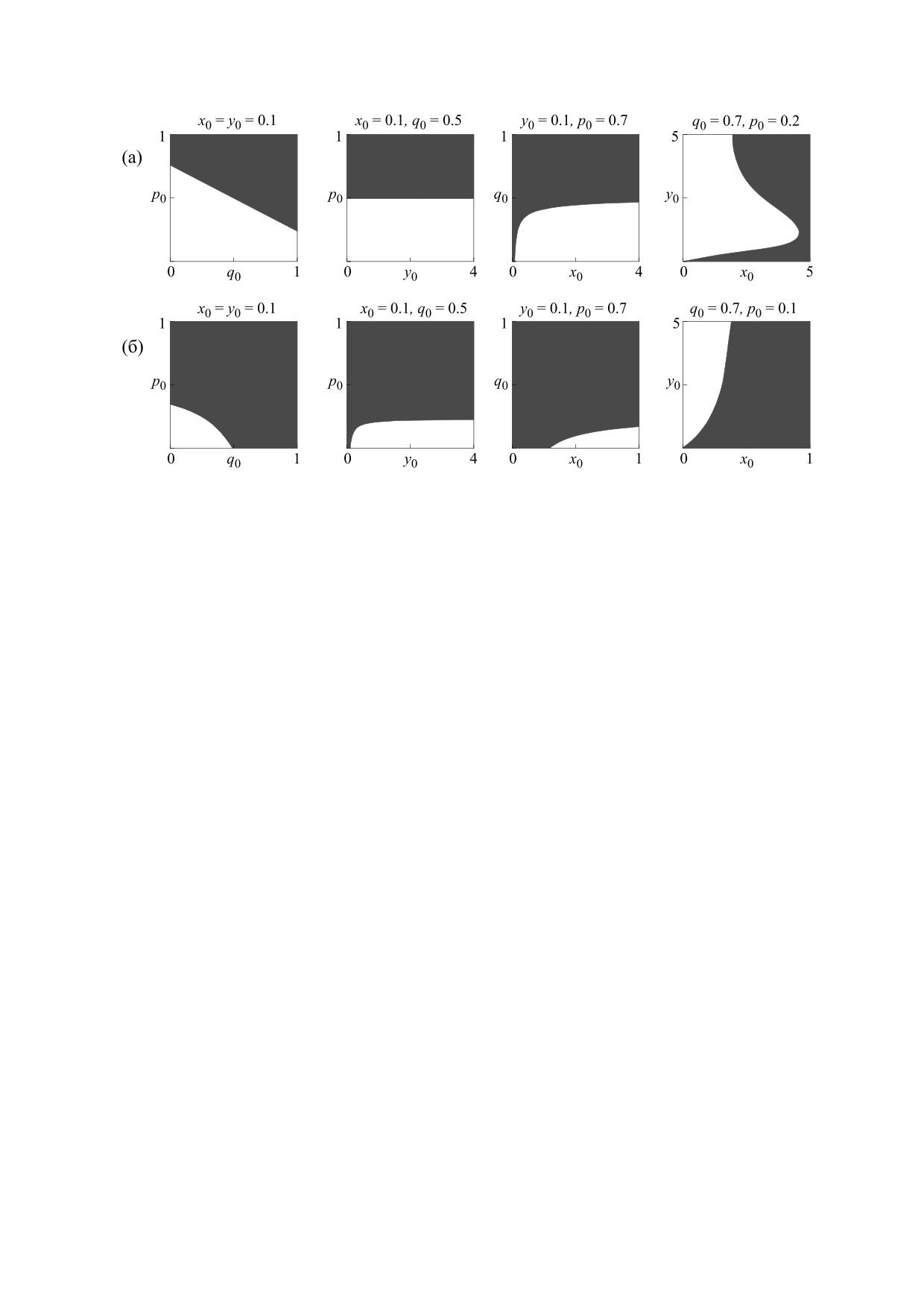
Рис. 9. Бассейны притяжения мономорфных равновесий модели (11) в области бистабильности. Белые области
-
-
соответствуют областям притяжения мономорфного решения
q = 0, –
= 0, а черные -
q = 1, –
= 1. Значения
параметров: панель (а) - r22 = 5, панель (б) - r22 = 3, r11 = 5, r12 = 2, с = 0.2, ρ = 0.2.
него примера проявления этого режима динами-
фиксированным периодом размножения. Посто-
ки (при полной изоляции между поколениями)
янство параметров модели означает, что к началу
можно отметить выраженную аллозимную диф-
следующего периода популяция оказывается в
ференциацию популяций тихоокеанской горбу-
тех же условиях окружающей среды, которые бы-
ши (Oncorhynchus gorbuscha), вида, нерестующего
ли и в начале предыдущего. Другими словами, все
на втором году жизни. Благодаря этой особенно-
потребляемые популяцией ресурсы успевают
сти в одной реке размножаются особи четных и
полностью восстановиться. Полученным на мо-
нечетных генераций, между которыми существу-
делях с дискретным временем трансформациям
ет жесткая репродуктивная изоляция. В результа-
режимов динамики при изменении параметров
те две субпопуляции четных и нечетных лет по-
модели можно дать следующую содержательную
рождают двухлетний цикл. На основе данных об
интерпретацию. Численность популяций видов с
аллозимной изменчивости было показано нали-
небольшим репродуктивным потенциалом (не-
чие заметных генетических различий между по-
большой средней приспособленностью) успевает
колениями четных и нечетных лет, которые и де-
монотонно перейти в равновесие, которое обес-
монстрируют выявленный эффект генетической
печивается существующими условиями среды.
дифференциации [47, 48]. Таким образом, мы ви-
Для видов с несколько бóльшим репродуктивным
дим, что наряду с известными и ожидаемыми ре-
потенциалом такой переход происходит в виде за-
жимами эволюционной динамики в рамках даже
тухающих колебаний. Виды с большим репродук-
простейших моделей отбора проявляются новые
и интересные эффекты, требующие дальнейшего
тивным потенциалом при численности популя-
исследования и осмысления.
ции вблизи равновесия дают слишком большое
количество приплода, которое подрывает ресурс-
ную базу, и численность падает. Возникают неза-
ОБСУЖДЕНИЕ
тухающие осцилляции. Интуитивно казалось, что
естественный отбор должен приводить к тому,
Колебания численности и хаотические режи-
чтобы виды как можно полнее использовали име-
мы популяционной динамики в рассматривае-
ющие ресурсы жизнедеятельности и давали боль-
мых моделях в большой степени связаны с дис-
шую равновесную численность при заданных па-
кретным представлением времени. Такое пред-
ставление времени связано с биологическими
раметрах экологической ниши. Фактически так и
особенностями видов, динамика популяций ко-
происходит, но оказывается, что рост равновес-
торых рассматривается в данной работе. Предпо-
ной численности в конечном итоге приводит к
лагается, что эти виды имеют выраженный вре-
потере ее устойчивости. Особи начинают давать
менной цикл развития (например, годовой) с
слишком много потомства, и популяция переска-
БИОФИЗИКА том 65
№ 5
2020
962
ФРИСМАН и др.
Рис. 10. Три устойчивых динамических режима (и бассейны их притяжений) модели (11) при пониженной
приспособленности гетерозигот. При одних начальных условиях (бассейн показан белым) в популяции
-
-
устанавливается мономорфизм
q = 0, –
= 0, при других (бассейн показан черным цветом) - мономорфизм
q = 1,
–
= 1, а при третьих (бассейн показан серым цветом) - устанавливаются устойчивые колебания частот генов с
периодом, равным двум, причем во всех трех случаях численность популяции стремится к устойчивому равновесному
уровню.
кивает равновесное состояние со всеми вытекаю-
собленностей. Наличие возрастной структуры
щими последствиями.
обеспечивает возможность двух сценариев: из-
вестный из одномерных моделей сценарий Фей-
В работе развита концепция плотностно-неза-
генбаума, приводящий к «пилообразным» коле-
висимого отбора в экологически лимитирован-
баниям численности, весьма редко встречаю-
ных популяциях (F-отбора). Показано, что
щимся в природе, и сценарий рождения
результаты такого отбора не отличаются от ре-
зультатов отбора по «фишеровским» приспособ-
инвариантной кривой (аналог предельного цик-
ла), приводящий к более-менее плавной модель-
ленностям в свободно размножающихся популя-
циях. Плотностное лимитирование приводит к
ной цикличности, похожей на то, что часто на-
тому, что при достижении больших средних
блюдается в природе. Эффекты, полученные в
репродуктивных потенциалов популяций их
наших моделях, во многом являются простым
равновесная численность теряет устойчивость и
объединением (суперпозицией) эффектов двух
возникают колебания. При этом генетическая
моделей: естественный отбор приводит к эволю-
структура определяется соотношением приспо-
ционному росту приспособленности (здесь к ро-
БИОФИЗИКА том 65
№ 5
2020
ЭКОЛОГО-ГЕНЕТИЧЕСКИЕ МОДЕЛИ
963
сту репродуктивного потенциала), а плотностное
чае. Нередко в динамике природных популяций
регулирование при увеличении репродуктивного
выявляются квазипериодические режимы. При-
потенциала приводит к известным бифуркациям,
рода этой квазипериодики чаще всего вызывает
обеспечивающим устойчивые различные флукту-
дискуссию и, как правило, объявляется «зашум-
ации численности. Только теперь эти флуктуа-
ленной» периодичностью. Вместе с тем при учете
ции появляются (и проявляются) в ходе генетиче-
возрастной структуры квазипериодические ре-
ской эволюции.
жимы появляются в детерминированной модели
при вполне естественных значениях параметров,
Принципиально новое и, по-видимому, самое
характерных для популяций многих видов, обла-
интересное это то, что здесь обнаружилось воз-
дающих возрастной структурой и стадийностью
никновение устойчивых колебаний не только
развития. Такие режимы наблюдаются в моделях
численности, но и частот генов. Вообще-то F-от-
в случаях, когда экологическое лимитирование
бор не является плотностно-зависимым отбором:
осуществляется путем уменьшения рождаемости
репродуктивные потенциалы генотипов являют-
с ростом численности половозрелых особей или
ся константами и не зависят ни от уровня числен-
путем уменьшения выживаемости молоди с ро-
ности, ни от уровня плотности. Ожидалось, что
стом численности молоди. Эти виды регуляции
F-отбор вызовет монотонное изменению частот,
популяционного роста весьма распространены в
но при этом может привести и к изменению дина-
природе и именно при такой регуляции чаще все-
мического режима популяции и создать условия
го наблюдаются квазипериодические режимы ди-
для r- и К-отбора. Но оказалось, что в рамках
намики численности.
только F-отбора возможны бифуркации, которые
приводят к возникновению устойчивых колеба-
Дальнейший рост средней приспособленности
ний частот аллелей. При этом уже нарушается
теоретически может вызвать как возникновение
принцип простого объединения (суперпозиции)
хаотических режимов, так и увеличение амплиту-
результатов двух моделей: плотностно-независи-
ды квазипериодических колебаний, что, в свою
мого естественного отбора и плотностно-зависи-
очередь, может привести к достижению столь ма-
мой регуляции роста численности; появляются
лых значений численности, при которых детерми-
режимы, которые не наблюдались отдельно в
нистическое описание оказывается недостаточно
каждой из моделей: на фоне колебания численно-
адекватно. При вероятностном (стохастическом)
сти возникают колебания частот генов, связан-
описании процесса динамики экологически лими-
ные с мультирежимностью рассматриваемых
тированных популяций учениками А.А. Ляпунова
систем.
Ю.Г Каревым и С.А. Терсковым было показано,
что однородные лимитированные популяции об-
Флюктуации колебаний численности харак-
речены на вымирание [49]. Заметим, что большин-
терны для подавляющего большинства природ-
ство существовавших в природе популяций вымер-
ных популяций биологических видов, имеющих
ло, и многие существующие обречены на вымира-
высокий репродуктивный потенциал и сезонный
ние. Вместе с тем в весьма изящном исследовании,
режим жизненного цикла: насекомые, многие ко-
выполненном учениками В.А. Ратнера М.А. Коро-
роткоцикличные виды рыб, многие мелкие мле-
стышевским и М.Р. Штабным, было показано, что
копитающие и т.д. Причины этих колебаний бы-
необходимыми условиями, требуемыми для «не-
ли, есть и будут объектом многих версий и дис-
вымирания» популяции, являются, в частности,
куссий. Но их существование не вызывает
процессы, обеспечивающие выполнение триады
сомнений.
Ч. Дарвина - изменчивости, наследственности и
Проведенное исследование показывает, что
естественного отбора [50, 51].
теоретически эти флуктуации изначально вполне
В наших моделях наследственность и есте-
могли возникнуть в ходе эволюционного процес-
ственный отбор присутствуют, а изменчивость
са в экологически лимитированных популяциях в
неявно предполагается в виде мутаций, обеспечи-
результате независимого от плотности населения
вающих начальное распределение генотипов.
естественного отбора в случаях, когда рост при-
Вместе с тем, поскольку численность популяции
способленности приводит к тому, что интенсив-
в модельных расчетах может падать до весьма
ное воспроизводство популяции не позволяет ей
низких значений, возможность вырождения
перейти к устойчивому равновесию.
остается. Долгое успешное существование при-
Выявление хаотических режимов динамики в
родных флуктуирующих популяций можно объ-
реальных популяциях крайне сложная задача.
яснить в частности следующими соображениями.
Прежде всего, нужны длинные ряды наблюде-
Как показано в данной работе, плотностно-неза-
ний, по которым можно было бы надежно рас-
висимый отбор (F-отбор) в лимитированных по-
считать показатели хаотичности (например, по-
пуляциях увеличивает среднюю приспособлен-
казатели Ляпунова). Конечно, всегда остаются
ность особей, увеличивает равновесную числен-
сомнения, как в надежности показателей, так и в
ность и вначале увеличивает среднее время жизни
причинах хаотизации в каждом конкретном слу-
(до вырождения) популяции, но затем приводит к
БИОФИЗИКА том 65
№ 5
2020
964
ФРИСМАН и др.
флуктуациям численности и росту вероятности
лиморфизму; пониженная относительная приспо-
вырождения. Однако возникновение и наличие
собленность гетерозигот приводит к бистабильно-
флуктуаций само по себе становится новой аре-
сти мономорфных состояний.
ной для действия отбора. В результате мутаций
Плотностное лимитирование приводит к тому,
появляются новые генотипы, чувствительные к
что при достижении больших средних репродук-
уровню плотности и дающие преимущества их
тивных потенциалов популяций их равновесная
обладателям либо в фазах роста, либо в фазах па-
численность теряет устойчивость и возникают
дения численностей. Колебания частот этих ге-
колебания. При этом генетическая структура
нотипов синхронизируются с колебаниями чис-
определяется соотношением приспособленно-
ленности, а затем регуляризируют и сглаживают
стей: при движущем отборе - мономорфизм,
эти колебания. Это позволяет либо сильно увели-
при сверхдоминировании - полиморфизм, при
чить время жизни популяции, либо вообще избе-
пониженном репродуктивном потенциале гете-
жать процесса вырождения. Принципиальным
розигот - ловушка бистабильности.
здесь является то, что рост средней приспособ-
ленности, вызванный плотностно-независимым
Наличие возрастной структуры обеспечивает
отбором, может оказаться причиной возникнове-
возможность двух сценариев: известный из одно-
ния колебаний, которые, уже в свою очередь, мо-
мерных моделей сценарий Фейгенбаума, приво-
гут привести к плотностно-зависимому отбору и
дящий к «пилообразным» модельным колебани-
процессам регуляризации. Другими словами,
ям численности, весьма редко встречающимся в
плотностно-зависимый отбор может быть полез-
природе, и сценарий рождения инвариантной
ным следствием и модификатором возникших
кривой (аналог предельного цикла), приводящий
колебаний численности, но не их причиной.
к более-менее плавной модельной циклике, по-
хожей на то, что часто наблюдается в природе.
ЗАКЛЮЧЕНИЕ
Эффекты, наблюдаемые в наших моделях, во
многом появляются в силу простого объединения
Проведенные ранее исследования нелиней-
(суперпозиции) двух моделей: естественный от-
ных моделей динамики популяций с возрастной
бор приводит к эволюционному росту приспо-
структурой показали, что увеличение средней ин-
собленности (здесь к росту репродуктивного по-
дивидуальной приспособленности приводит к
тенциала), а плотностное регулирование при уве-
колебаниям численности, а затем к возникнове-
личении репродуктивного потенциала приводит
нию хаотических аттракторов, структура и раз-
к известным бифуркациям, обеспечивающим
мерность которых меняются при изменении па-
различные флуктуации численности. Только те-
раметров модели. В частности, увеличение рож-
перь эти флуктуации появляются (и проявляют-
даемости и уменьшение смертности возрастных
ся) в ходе генетической эволюции.
групп приводят к усложнению структуры аттрак-
торов и росту фрактальной размерности [33-36].
Принципиально новое и, по-видимому, самое
интересное это то, что здесь обнаружилось воз-
Здесь мы обобщили все результаты, свидетель-
никновение устойчивых колебаний не только
ствующие о том, что перечисленные типы дина-
численности, но и частот генов. Вообще-то F-от-
мики численности могли бы последовательно
бор не является плотностно-зависимым отбором:
возникать в эволюции лимитированной популя-
репродуктивные потенциалы генотипов являют-
ции под действием плотностно-независимого
ся константами и не зависят от уровня численно-
естественного отбора, повышающего среднюю
сти. Ожидалось, что F-отбор вызовет монотонное
приспособленность популяции в соответствии с
изменение частот, но при этом может привести и
фундаментальной теоремой естественного отбора
к изменению динамического режима популяции
Р. Фишера. Эффект совокупного одновременно-
и создать условия для r- и К-отбора. Но оказа-
го взаимодействия плотностно-независимого от-
лось, что в рамках F-отбора возможны бифурка-
бора и плотностно-зависимых неселективных
ции, которые приводят к возникновению устой-
экологических лимитирующих факторов, назван
чивых колебаний частот аллелей.
нами F-отбором.
Показано, что результаты F-отбора в экологи-
При этом нарушается принцип простого объ-
чески лимированных популяциях не отличаются
единения (суперпозиции) результатов двух моде-
от результатов отбора по «фишеровским» приспо-
лей: плотностно-независимого естественного от-
собленностям в свободно размножающихся попу-
бора и плотностно-зависимой регуляции роста
ляциях: промежуточные значения приспособлен-
численности; появляются режимы, которые не
ностей гетерозигот, относительно припособленно-
наблюдались отдельно в каждой из моделей: ко-
стей гомозигот (движущий отбор), приводят к
лебания частот генов, связанные с мультирежим-
установлению мономорфизма по аллелям, обеспе-
ностью рассматриваемых систем, которая возни-
чивающим больший репродуктивный потенциал;
кает в результате бифуркационного появления
сверхдоминирование приводит к устойчивому по-
новых устойчивых аттракторов.
БИОФИЗИКА том 65
№ 5
2020
ЭКОЛОГО-ГЕНЕТИЧЕСКИЕ МОДЕЛИ
965
Другой парадокс F-отбора заключается в том,
17. I. Hanski, Metapopulation ecology (Oxford University
что он, будучи независимым от плотности, при-
Press, N.Y., 1999).
водит к колебаниям и хаотическим режимам ди-
18. В. И. Евсиков, Г. Г. Назарова и В. Г. Рогов, Сибир-
намики численности, которые создают условия
ский экологич. журн. 1, 59 (1999).
для плотностно-зависимого отбора, такого как r-
19. Г. В. Гречаный, А. Я. Никитин, В. М. Корзун и др.,
и К-отбор.
Эколого-генетическая детерминация динамики чис-
ленности популяций (ФГБОУ ВО «ИГУ», Иркутск,
2004).
ФИНАНСИРОВАНИЕ РАБОТЫ
20. B. С. Артамонова и А. А. Махров, Генетика 42 (3),
Работа выполнена в рамках государственных
310 (2006).
заданий Института автоматики и процессов
21. U. Dieckmann and J. A. J. Metz, Theor. Popul. Biol. 69
(3), 263 (2006).
управления ДВО РАН и Института комплексного
22. M. Traykov and I. Trenchev, Rus. J. Genet., 52 (9), 985
анализа региональных проблем ДВО РАН, а так-
(2016).
же при частичной финансовой поддержке Рос-
23. А. П. Шапиро, Управление и информация 3, 96
сийского фонда фундаментальных исследований
(1972).
(проект № 18-04-00073а).
24. R. M. May, J. Theor. Biol. 51 (2), 511 (1975).
25. P. H. Leslie, Biometrika 33 (3), 183 (1945).
КОНФЛИКТ ИНТЕРЕСОВ
26. P. H. Leslie, Biometrica 35 (3/4), 213 (1948).
27. L. P. Lefkovitch, Biometrics 21 (1), 1 (1965).
Авторы заявляют об отсутствии конфликта
28. H. Caswell, Matrix Population Models: Construction,
интересов.
Analysis, and Interpretation (Sinauer Associates Inc.,
Sunderland, MA, 2001).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
29. Е. Я. Фрисман, М. П. Кулаков, О. Л. Ревуцкая и
др., Компьютерные исследования и моделирова-
Настоящая работа не содержит описания ка-
ние 11 (1), 119 (2019).
ких-либо исследований с использованием людей
30. Р. Дажо, Основы экологии (Прогресс, М., 1975).
и животных в качестве объектов.
31. Ю. М. Свирежев и Д. О. Логофет, Устойчивость
биологических сообществ (Наука, М., 1978).
СПИСОК ЛИТЕРАТУРЫ
32. Е. Я. Фрисман, С. П. Луппов, И. Н. Скокова и др.,
в кн. Математические исследования в популяцион-
1. T. R. Malthus, The works of Thomas Robert Malthus
ной экологии (ДВО АН СССР, Владивосток, 1988),
(Pickering & Chatto Publishers, London, 1986).
сс. 4-18.
2. C. Darwin, On the origin of species by means of natural
33. Е. Я. Фрисман и Е. И. Скалецкая, Обозрение при-
selection or the preservation of favoured races in the strug-
кладной и промышленной математики 1 (6), 988
gle for life (Oxford University Press, H. Milford, 1859).
(1994).
3. R. A. Fisher, The genetical theory of natural selection
34. E. Ya. Frisman, G. P. Neverova, and O. L. Revutskaya,
(Clarendon Press, Oxford, 1930.
Ecol. Model. 222, 1943 (2011).
4. S. Wright, J. Hered. 21, 340 (1930).
35. Е. Я. Фрисман, Г. П. Неверова, О. Л. Ревуцкая и
5. J. B. S. Haldane, The causes of evolution (Longman
др., Изв. вузов. Прикладная нелинейная динамика
Green, London, 1932).
18 (2), 111 (2010).
6. С. С. Четвериков, О некоторых моментах эволюци-
36. Е. Я. Фрисман и О. Л. Жданова, Генетика 45 (9),
онного процесса с точки зрения современной генети-
1277 (2009).
ки (Госиздат., 1926).
37. M. J. Feigenbaum, J. Stat. Phys. 19 (1), 25 (1978).
7. P. F. Verhulst, Nouv. Mem. Acad. Roy. Soc. Bellelettr.
38. М. Фейгенбаум, Успехи физ. наук 141 (10), 343
Bruxelles 18, 1 (1845).
(1983).
8. В. Вольтерра, Математическая теория борьбы за
39. С. П. Кузнецов, Динамический хаос (Физматлит,
существование (Наука, М., 1976).
M., 2006).
9. Г. Ф. Гаузе, Зоол. журн. 14 (2), 243 (1935).
40. В. А. Ратнер, Математическая популяционная гене-
10. Г. Ф. Гаузе, в кн. Экология и эволюционная теория
тика (Наука, Новосибирск, 1977).
(Наука, Л, 1984), сс. 5-105.
41. T. I. Axenovich, I. V. Zorkoltseva, I. R. Akberdin, et al.,
11. L. C. Birch, Evolution 9, 389 (1955).
Heredity 98, 99 (2007).
12. B. Charlesworth, Ecology 52, 469 (1971).
42. M. S. Graines, L. R. McClenaghay, and R. R. Rose,
13. L. D. Gottlieb, Genetics 76 (3), 551 (1974).
Evolution 32 (4), 723 (1978).
14. M. S. Gaines, Jr, L. R. McClenaghan, and R. K. Rose,
43. J. F. C. Kingman, Proc. Can. Phill. Soc. 57, 574 (1961).
Evolution 32 (4), 723 (1978).
44. А. П. Кузнецов, С. П. Кузнецов, М. В. Поздняков
15. Е. Я. Фрисман, Первичная генетическая диверген-
и др., Нелинейная динамика 8 (3), 461 (2012).
ция (теоретический анализ и моделирование)
45. Ф. Б. Чернявский и А. Н. Лазуткин, Циклы леммин-
(ДВНЦ АН СССР, Владивосток, 1986).
гов и полевок на Севере (ИБПС ДВО РАН, Магадан,
16. Е. Я. Фрисман, Вестн. ДВО РАН 4, 97 (1995).
2004).
БИОФИЗИКА том 65
№ 5
2020
966
ФРИСМАН и др.
46. Г. П. Неверова, О. Л. Жданова и Е. Я. Фрисман,
ронцова (Наука, Сибирское отделение, Новоси-
Генетика 56 (6), 714 (2020).
бирск, 1975), сс. 5-10.
50. М. А. Коростышевский, М. Р. Штабной и
47. С. П. Пустовойт, Вавиловский журн. генетики и
В. А. Ратнер, в кн. Вопросы математической гене-
селекции 15 (3), 475 (2011).
тики (ИЦиГ СО АН СССР, Новосибирск, 1974),
48. С. П. Пустовойт, Изв. ТИНРО 188, 162 (2017).
сс. 5-32.
49. Г. П. Карев, А. А. Ляпунов и С. А. Тресков, в кн.
51. M. A. Korostyshevsky, M. R. Shtabnoy and V. A. Rat-
Современные проблемы эволюции, под ред. Н. Н. Во-
ner, J. Theor. Biol. 48, 85 (1974).
Ecological and Genetic Models in Population Biophysics
E.Ya. Frisman*, O.L. Zhdanova**, and G.P. Neverova**
*Institute for Complex Analysis of Regional Problems, Far East Branch of the Russian Academy of Sciences,
ul. Sholom-Aleikhema 4, Birobidzhan, 679016 Russia
**Institute of Automation and Control Processes, Far East Branch of the Russian Academy of Sciences,
ul. Radio 5, Vladivostok, 690041 Russia
There are complex causal relationships between population dynamics and a change in population genetic
structure. In our study a simple model was used to show that the evolutionary process of density-independent
natural selection depending on the fitnesses determined by a single diallelic locus could lead to a change in
population growth parameters changing the dynamic modes in population. Possible mechanisms and direc-
tion of these changes are discussed. Results obtained clearly demonstrate the evolutionary change of allele
frequencies accompanied by an increase in the average fitness in the population that may result in appearance
of periodic, quasiperiodic and chaotic modes of population dynamics. The effects observed in our models are
largely due to the simple combination (superposition) of the two models: natural selection leads to an evolu-
tionary increase in the fitness, and density regulation with an increase in reproductive potential leads to bi-
furcations that provide various fluctuations in population number. At that in eco-evolutional models, these
fluctuations appear (and progress) due to genetic evolution. At the same time, combined models give appear-
ance of new dynamic modes that were not observed in each of the separate models, i.e. fluctuations in gene
frequencies associated with the multi-modality of the considered systems and the emergence of new stable
attractors.
Keywords: mathematical modeling, evolution, natural selection, population dynamics, bistability of trapping posi-
tionkl
БИОФИЗИКА том 65
№ 5
2020