БИОФИЗИКА, 2020, том 65, № 5, с. 978-985
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 638.141
МОДЕЛИРОВАНИЕ ВЕСЕННЕЙ АГРЕГАЦИИ ПЧЕЛ В УЛЬЕ
ПРИ БОЛЬШИХ СУТОЧНЫХ КОЛЕБАНИЯХ ТЕМПЕРАТУРЫ
НАРУЖНОГО ВОЗДУХА
© 2020 г. С.В. Оськин, Д.А. Овсянников
Кубанский государственный аграрный университет им. академика И.Т. Трубилина,
350044, Краснодар, ул. Калинина, 13
E-mail: kgauem@yandex.ru
Поступила в редакцию 15.02.2020 г.
После доработки 15.02.2020 г.
Принята к публикации 05.06.2020 г.
Прохождению пчелами зимнего периода посвящено много исследований, что говорит сложности
понимания такого состояния пчелиной семьи. Однако самым критическим периодом для пчел яв-
ляется конец зимы и весна. В это время наблюдаются большие температурные колебания воздуха,
и пчелы вынуждены время от времени переходить из пассивного состояния в активное и наоборот.
Наиболее явно этот феномен появляется на пчеловодческих пасеках, расположенных в районах Се-
верного Кавказа. В весеннее время положительные высокие температуры воздуха провоцируют
пчел на вылет из улья, и не все пчелы к концу дня возвращаются в улей. Низкие температуры могут
привести к агрегации пчел в неудобном месте, где нет запасов корма или отсутствует возможность
добраться к запасам меда. Данное исследование посвящено именно этим периодам существования
пчел. Моделирование основных физических процессов проводилось в программном обеспечении
Comsol. В ранее разработанные модели были добавлены слагаемые, связанные с переходными про-
цессами и учитывающие теплоемкости элементов улья. Также модели учитывали изменение объема
зимней агрегации в зависимости от окружающей температуры. По результатам моделирования
установлено, что действительно при весенних колебаниях температуры воздуха теплоемкость медо-
вых запасов значительно влияет на внутренний температурный режим улья. Запасенная в меде теп-
ловая энергия сглаживает температурные колебания воздуха внутри улья, что является положитель-
ным фактором для пчел.
Ключевые слова: агрегация пчел, улей, температура, электрический обогрев, моделирование.
DOI: 10.31857/S0006302920050154
Известно, что пчеловоды уделяют много вни-
ные суточные колебания температуры. Наиболее
мания подготовке к зимовке пчелиных семей.
высокие значения суточных температурных коле-
Осенью необходимо правильно определить объем
баний отмечаются на территории Северного Кав-
корма и его месторасположение в улье, предполо-
каза. В данной местности днем температура воз-
жить место, где будет находиться пчелиный клуб.
духа может доходить до плюс 10-14°С, а ночью
Правильно сформированная семья в зиму - это
опускаться до нуля или даже до отрицательных
высокая вероятность успешной зимовки. Осо-
значений - до -5°С. В конце зимы и начале вес-
бенно критический период для пчелиной семьи -
ны возрастает роль электрического подогрева
конец зимы и начало весны. В это время часто за-
ульев, так как это дает возможность пчелам за-
пасы корма на исходе, кроме того, пчелиный клуб
брать мед даже в дальних частях улья и стабилизи-
может сместиться в одну сторону улья, и пчелы не
рует температуру внутри улья [1]. Для того чтобы
могут добраться до медовых рамок, расположен-
создать эффективную систему электроподогрева
ных в другой стороне. Также нужно учитывать,
в ульях, нужно иметь адекватные модели агрега-
что в это время в улье уже есть расплод, и пчелы
ции пчел и теплопроводности элементов улья в
повышают температуру воздуха внутри своей аг-
этот период. Моделированию тепловых процес-
регации, что, естественно, сопровождается более
сов внутри улья зимой был посвящен ряд работ [2,
высоким потреблением корма. Нужно отметить,
3, 4, 5]. Однако весеннее состояние пчел и пара-
что в эти периоды года наблюдаются существен-
метры окружающего воздуха в этот период требу-
978
МОДЕЛИРОВАНИЕ ВЕСЕННЕЙ АГРЕГАЦИИ ПЧЕЛ В УЛЬЕ
979
Рис. 1. Виды геометрической модели объекта исследований.
ет особенного подхода к моделированию. Пчело-
МОДЕЛЬ
воды замечают, что весной в теплый день пчелы
Математическое моделирование велось в сре-
иногда не выходят на очистительный облет. При-
де моделирования Comsol. Ранее были представ-
ходится даже их возбуждать, чтобы они вышли из
лены результаты моделирования основных физи-
улья. Скорее всего, это связано с тем, что после
ческих процессов пчелиной семьи из 15000 пчел,
длительной зимовки с низкими температурами
зимующей в дадановском улье [1, 2]. Зимняя агре-
внутри улья мед принимает температуру, практи-
гация пчел представлена в виде эллипсоида, со-
чески равную температуре окружающей среды
стоящего из нескольких цилиндров разного раз-
мера. Часть цилиндров - это пчелы, размещаю-
(особенно на краях улья). При значительном ко-
щиеся в ячейках сот, и пчелы, расположенные в
личестве меда в улье (может еще доходить до 10-
межсотовом пространстве (рис. 1).
20 кг) за счет высокой теплоемкости внутренняя
температура будет держаться длительное время на
Для моделирования прохождения воздуха че-
рез клуб введены воздушные цилиндры (по два на
низкой отметке даже при положительных значе-
улочку) с изменяющейся высотой цилиндра. Эти
ниях температуры наружного воздуха. В этом слу-
цилиндры по своему объему будут эквивалентны
чае важно произвести подогрев кормовых запасов
общему объему воздуха в улочке, занимаемой
заранее с помощью системы электроподогрева.
пчелами. Высота воздушных цилиндров изменя-
Может возникать и другой случай, когда днем до-
ется по линейному закону в функции темпера-
вольно долго держалась высокая температура, а
туры.
ночью пошло понижение, и возникает вопрос -
Так как в зимний период плотность клуба из-
есть ли необходимость сразу включать электро-
меняется, то будет изменяться и радиус цилин-
подогрев. Таким образом, существует проблема
дров клуба в зависимости от температуры окру-
обоснованности использования в такие периоды
жающего воздуха. Объем клуба пропорционален
системы подогрева ульев. Если такая система бу-
квадрату радиуса, следовательно, при максималь-
дет работать, основываясь только на сигналах
ной плотности пчел радиус клуба уменьшится в
0,71 раз от первоначального значения. В соответ-
датчиков температуры, и не будет учитывать теп-
ствии с принятыми геометрическими значения-
лоемкости тел, находящихся в улье, то это может
ми радиусы отдельных улочек будут описываться
приводить к повышенному расходу электроэнер-
уравнениями:
гии, задержке пчел на вылет из ульев или, наобо-
рот, к провокации на выход, гибели пчел с пони-
R
1 bee
= 1,07 ·Т
+ 110 мм
женным запасом корма.
R
= 0,93 ·Т
+95 мм
2 bee
,
(1)
Следовательно, необходимо разработать мо-
R
= 0,7 ·Т
+70 мм
3 bee
дель весенней агрегации пчел и провести модели-
R4 bee =
0,3 ·
Т
+35 мм
рование состояния внутреннего микроклимата
улья с применением системы электрообогрева.
где Т - температура окружающего воздуха.
БИОФИЗИКА том 65
№ 5
2020
980
ОСЬКИН, ОВСЯННИКОВ
Два воздушных цилиндра в улочке имеют вы-
Полученные уравнения для определения гео-
соту dair = 1.25 мм (при температуре наружного
метрических и физических параметров вводи-
воздуха 0°С). При снижении температуры будет
ли в соответствующие блоки программы. Ранее
происходить уменьшение этого параметра по ли-
анализ проводили только для стационарного
нейному закону, что можно представить следую-
режима [1, 5]. Теперь моделирование проходи-
щим образом:
ло для нестационарного режима, и в системы
уравнений были добавлены производные по
dair = 1.25 + 0.025Т (мм).
(2)
времени. Так, обобщенная математическая мо-
Моделирование физических процессов,
дель, описывающая тепловые процессы в улье,
происходящих в улье, проводили в Comsol 5.4. имеет вид:
∂T
∂T
∂T
∂T
ρ
⋅ с
⋅
+
ρ
⋅ с
⋅
+
ρ
⋅ с
⋅
+
ρ
⋅ с
⋅
+
ρ
⋅ с
⋅ u
⋅∇T
+
wood wood
hc
hc
empty h c empty h c
bee
bee
air1
air1
air1
∂t
∂t
∂t
∂t
+ρair2
⋅ с
air2
⋅ u
air2
⋅∇T +∇q
air1
+∇
q
air2
+∇q
wood
+∇q
hc
+∇q
empty h c
+∇q
bee
Nu = Q
bee
λ
bee
=
0,0076
−
0,0017
⋅Т
0
;ρ
пч
=
243
−
8
⋅Т
0
,
(3)
2
Q
bee
=
3,2
⋅Т
0
−
20
⋅Т
0
+
922
q
=-λΔT
i
i
где ρair1 и ρair2 - соответственно плотности воз-
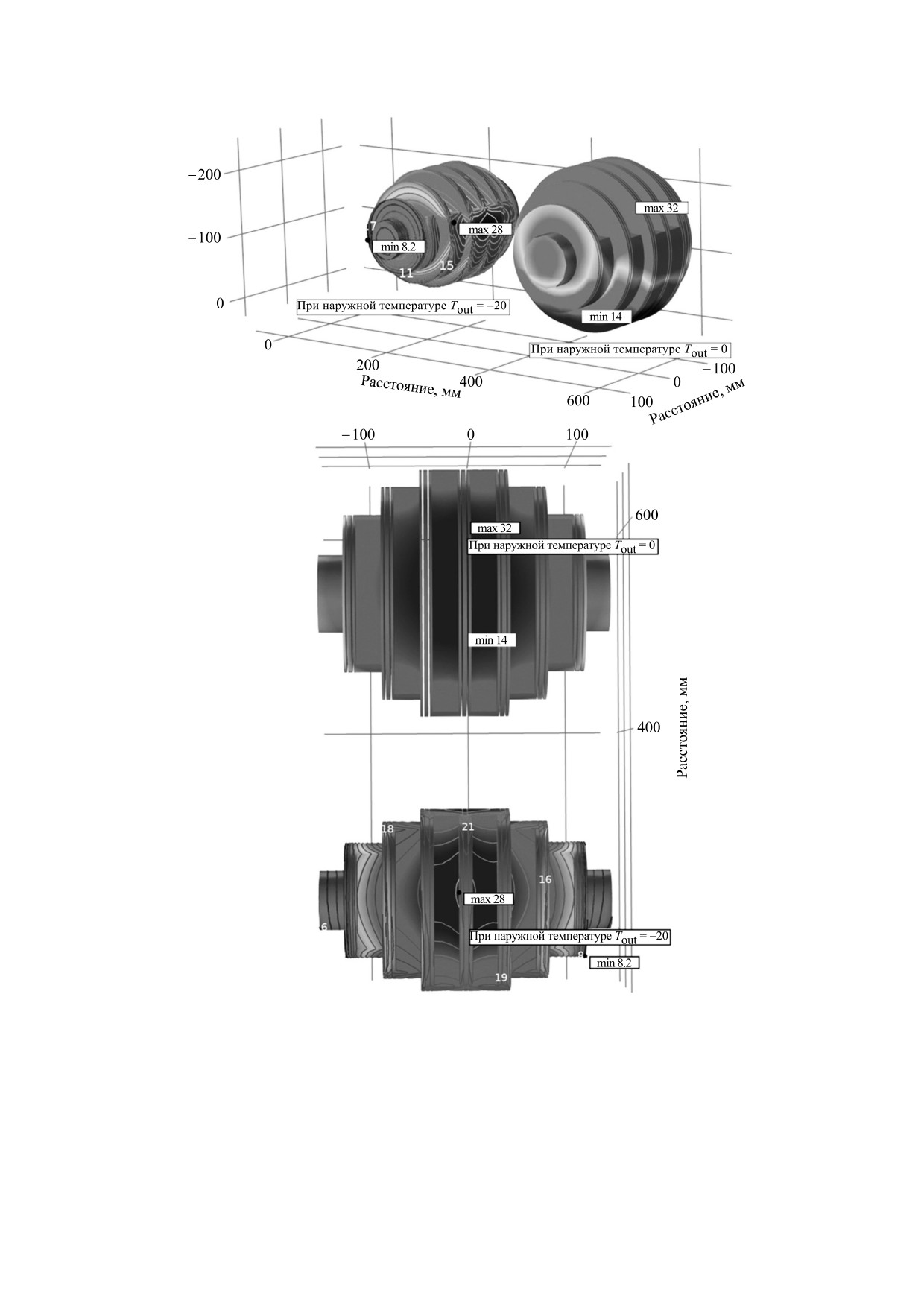
видно, что при температуре наружного воздуха -
духа, поступающего снаружи и проходящего че-
20°С клуб пчел сжимается, при этом минималь-
рез пчелиный клуб; cwood, chc, cempty hc, cbee - со-
ная температура зафиксирована на периферии
ответственно теплоемкости деревянных элемен-
клуба и равна 8.2°С, а максимальное значение
тов, медовых сот, пустых сот, пчелиного клуба;
температуры характерно для центра клуба (28°С).
cair1 и cair2 - соответственно теплоемкости воз-
При температуре наружного воздуха 0°С агрега-
духа первого и второго воздушных блоков (внут-
ция пчел увеличивает свои размеры, и минималь-
реннего воздуха и воздуха внутри клуба); uair1 и
ная температура становится равной 14°С, а мак-
uair2 - соответственно поля скоростей первого и
симальная - 32°С.
второго воздушных блоков, м/с; qair1, qair2, qwood,
В результате анализа температурных измене-
qhc, qempty hc, qbee - соответственно плотности
ний отдельных элементов улья и агрегации пчел
теряемых тепловых потоков путем теплопровод-
установлено, что при изменении наружной тем-
ности, воздушных блоков 1 и 2, деревянных эле-
пературы от -30°С до 13°С температурные поля
ментов, медовых сот, пустых сот, пчелиного
соответствуют реально возможным для данной
клуба, Вт/м2; Nu - число Нуссельта.
конструкции улья и конкретного количества
Аналогично были добавлены производные по
пчел. Пример распределения температуры внутри
времени в модели движения воздуха и влажности,
улья при наружном значении этого параметра,
представленные в литературе [5].
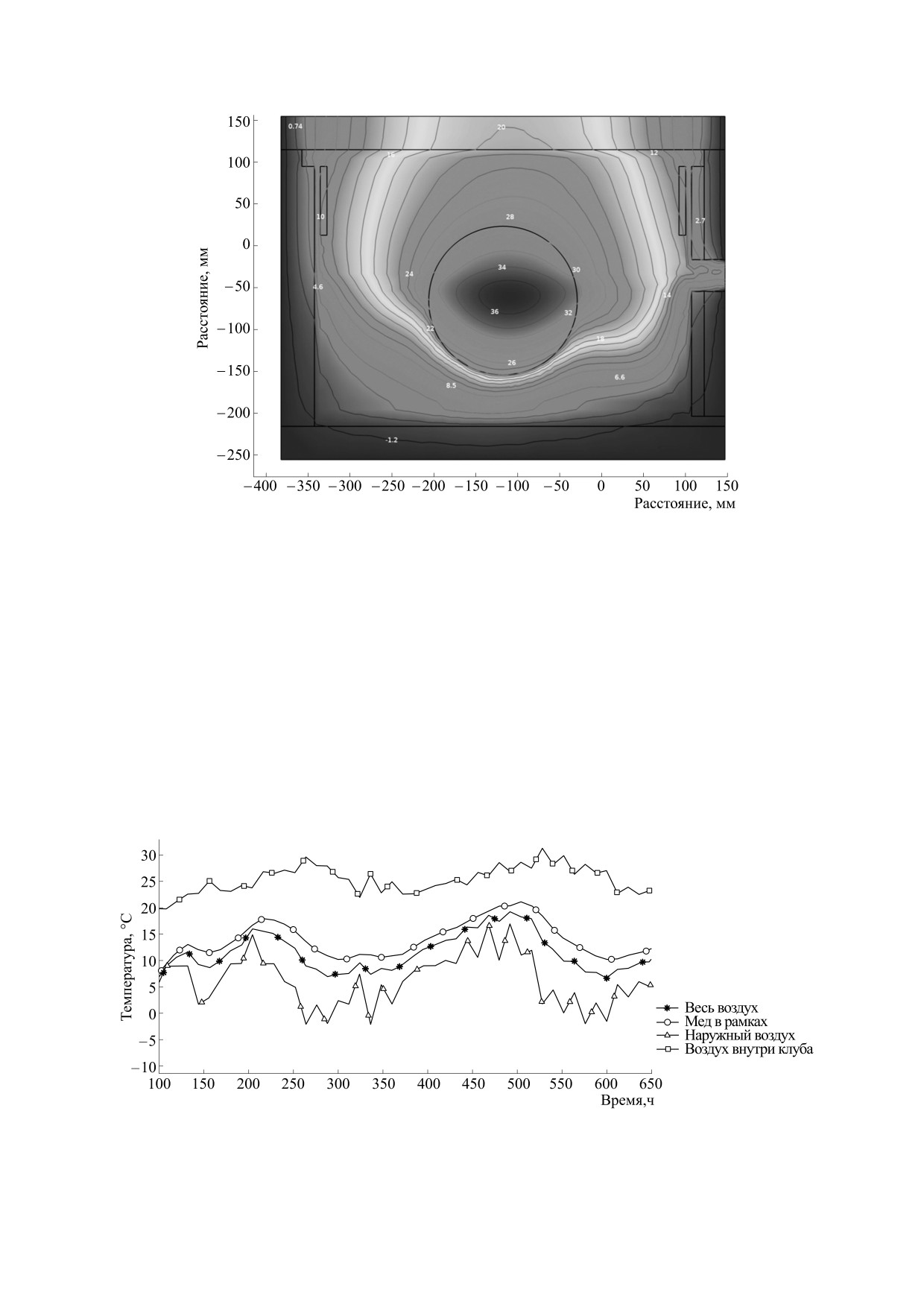
равном -3°С, приведен на рис. 3. На данном ри-
сунке представлен разрез через центр улья, и вид-
но, что ближе к стенкам улья температура нахо-
РЕЗУЛЬТАТЫ
дится на уровне -1°С, а максимальная температу-
ра зафиксирована внутри клуба со значением
Первоначально были моделированы темпера-
36°С. Также можно наблюдать высокие значения
турные поля внутри улья при разных наружных
температуры в верхней части улья - от 12°С до
температурах. При этом проводилось сопоставле-
20°С, что связано с хорошей теплоизоляцией по-
ние с реально возможными значениями темпера-
толка улья и тепловой конвекцией. Можно видеть
туры воздуха внутри клуба и внутри улья. Анали-
и выход теплого воздуха через верхний леток (в
зировалось изменение геометрии клуба вместе с
правой части разреза).
максимальными и минимальными значениями
температуры в отдельных точках агрегации пчел.
Моделирование транспорта влаги внутри улья
На рис. 2 представлены геометрические конфигу-
и движения воздуха во внутриульевом простран-
рации видов агрегации пчел при разных темпера-
стве показало совпадение практически всех про-
турах окружающего улей воздуха. Из рисунка
цессов с вариантом стационарного анализа [5].
БИОФИЗИКА том 65
№ 5
2020
МОДЕЛИРОВАНИЕ ВЕСЕННЕЙ АГРЕГАЦИИ ПЧЕЛ В УЛЬЕ
981
Рис. 2. Геометрические конфигурации видов агрегации пчел при разных температурах наружного воздуха.
Таким образом, при длительном сохранении тем-
ний внутри объекта исследований. При наступле-
пературы окружающего улей воздуха учет в моде-
нии конца зимы и начала весны расширяется
лях переходных процессов особого влияния не
диапазон суточных колебаний, и картины темпе-
оказывает на расчеты температурных распределе-
ратурных полей внутри улья начинают отличать-
БИОФИЗИКА том 65
№ 5
2020
982
ОСЬКИН, ОВСЯННИКОВ
Рис. 3. Температурное поле улья в разрезе через центральный леток и при температуре наружного воздуха минус 3°С.
ся от стационарных режимов. Такие температур-
весенний период. Для этого были взяты значения
ные перепады в течение суток характерны для
температуры воздуха в отдельные дни в Красно-
южных регионов России. В этих частых темпера-
дарском крае (Лабинском районе) в 2017 г. Темпе-
турных переходах начинает оказывать влияние на
ратурный диапазон был зафиксирован через каж-
температуру воздуха внутри улья теплоемкость
дые 12 ч с середины февраля до середины марта и
введен в модель.
элементов улья.
Следующим этапом моделирования было
ОБСУЖДЕНИЕ
определение диапазонов колебаний температуры
отдельными элементами объекта исследований
После осуществления моделирования неста-
при реальных изменениях температуры в зимне- ционарных режимов в улье и получения соответ-
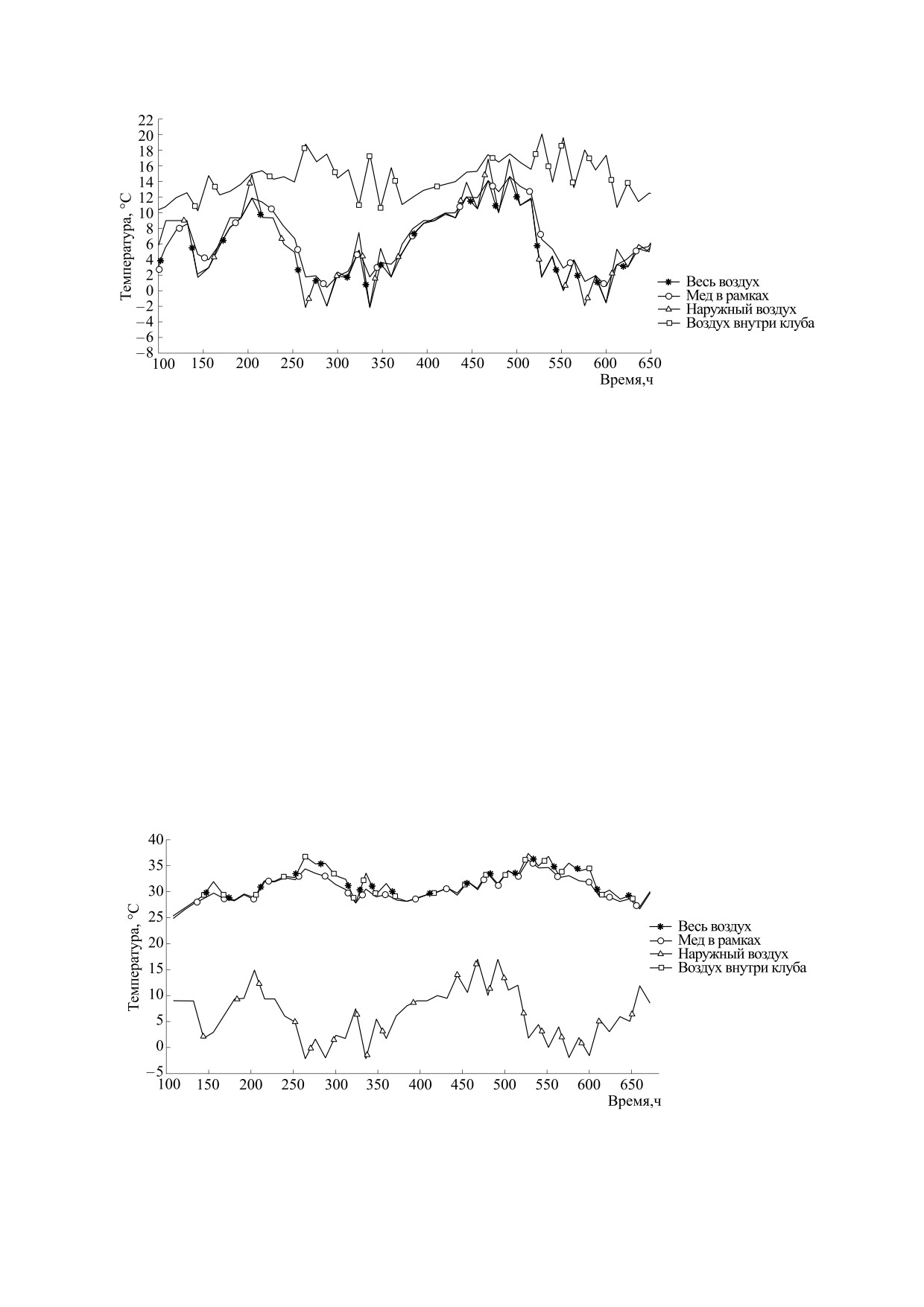
Рис. 4. Графики изменения средней температуры отдельных элементов улья.
БИОФИЗИКА том 65
№ 5
2020
МОДЕЛИРОВАНИЕ ВЕСЕННЕЙ АГРЕГАЦИИ ПЧЕЛ В УЛЬЕ
983
Рис. 5. Графики изменения минимальной температуры отдельных элементов улья.
ствующих данных был проведен анализ получен-
лоемкостью данного вещества. Также на темпера-
ных результатов. В первую очередь было необхо-
турном графике меда можно видеть более плав-
димо определить, как влияет теплоемкость
ные переходы от одной фиксированной точки к
материалов на температурные графики внутрен-
другой. Диапазон изменения температуры внут-
него воздуха в улье. На рис. 4 представлены гра-
реннего воздуха клуба пчел составил от 20°С до
фики изменения температуры окружающего улей
32°С (ΔT = 12°С), но переходы от одной точки к
воздуха (наружный воздух) и средних значений
другой более резкие, что связано с малой тепло-
температур: всего внутреннего воздуха в улье
емкостью воздуха и реакцией пчел на изменения
(весь воздух), медовых запасов в улье (мед в рам-
температуры. Температура воздуха внутри улья в
ках), внутреннего воздуха в агрегации пчел (воз-
среднем колебалась от 7°С до 19°С (ΔT = 12°С) с
дух внутри клуба).
умеренными колебаниями температуры между
соседними временными данными.
При анализе полученных графиков установле-
но следующее. За рассматриваемый период диа-
Представляют интерес графики изменения
пазон изменения наружной температуры воздуха
минимальной температуры в отдельных материа-
составил от -3°С до 17°С (ΔT = 20°С). Изменения
лах объекта в зависимости от времени (рис. 5). Из
температуры меда были явно меньше - от 8°С до
рисунка видно, что минимальная температура
21°С (ΔT = 13°С), что объясняется высокой теп-
воздуха внутри клуба пчел не опускается ниже
Рис. 6. Графики изменения максимальной температуры отдельных элементов улья.
БИОФИЗИКА том 65
№ 5
2020
984
ОСЬКИН, ОВСЯННИКОВ
плюс 11°С, что соответствует реальным значени-
при весенних колебаниях температуры теплоем-
ям, установленным различными авторами [2, 3,
кость медовых запасов значительно влияет на
7-10]. Минимальная температура всего внутрен-
внутренний температурный режим улья. Запасы
него воздуха доходила до температуры за предела-
тепловой энергии в меде сглаживают температур-
ми улья (-2°С), конечно, такая температура была
ные колебания воздуха внутри улья. Если темпе-
только на периферии улья - за крайними рамка-
ратура воздуха за ульем меняется на 10°С в тече-
ми. При этом график минимальной температуры
ние 12 ч, то не следует включать нагреватели из-за
в точках минимума практически совпадает с гра-
наличия запаса тепловой энергии в меде. Если
фиком наружной температуры. В точках с высо-
температура воздуха установилась низкая и такой
кими значениями наружной температуры график
режим длится более суток, то нужно включать на-
наружного воздуха значительно ниже, что объяс-
греватели. При этом нагреватели лучше секцио-
няется запасом тепловой энергии в меде. В этих
нировать на три части, и весной следует вести по-
точках наблюдается совпадение значений темпе-
догрев только кормовых запасов, т.е. крайних ра-
ратур внутреннего воздуха и меда. Также можно
мок. Такой режим позволит не провоцировать
отметить минимальное значение температуры
пчелиную матку на раннюю закладку яиц и, в то-
меда, которое находится на уровне 1°С, и эти зна-
же время, даст возможность пчелам забирать мед
чения присущи крайним рамкам с медом.
даже с периферии улья.
На рис. 6 показаны графики максимальных
температур в отдельных элементах улья. Здесь
КОНФЛИКТ ИНТЕРЕСОВ
видно, что графики максимальной температуры
Авторы заявляют об отсутствии конфликта
воздуха в улье и внутри клуба полностью совпада-
интересов.
ют. Это связано с тем, что такие температуры
пространственно расположены близко друг к
другу - при выходе воздуха из клуба. К этим гра-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
фикам близок и график максимальной темпера-
туры меда, так как это тоже температура воздуха
Настоящая работа не содержит описания ка-
внутри клуба рядом с нижним краем меда в цен-
ких-либо исследований с использованием людей
тральных рамках.
и животных в качестве объектов.
ВЫВОДЫ
СПИСОК ЛИТЕРАТУРЫ
1. С. В. Оськин и Д. А. Овсянников, Биофизика 65
Теплоемкость меда значительно влияет на
(2),1 (2020).
микроклимат пчелиного улья в переходные тем-
пературные периоды. Наиболее значительные
2. Е. К. Еськов и В. А. Тобоев, Биофизика 54 (1),114
колебания температуры окружающего воздуха от-
(2009).
мечаются весной на юге России, в частности в
3. В. А. Тобоев и М. С. Толстов, в сб. Мат-лы Всерос.
Краснодарском крае. Эти периоды особенно тя-
науч. конф-ии «Физические процессы в биологических
жело проходят пчелиные семьи: при высоких
системах» (Казань, 2014), сс. 97-102.
температурах пчелиный клуб расширяется,
4. В. А. Тобоев и М. С. Толстов, Межотраслевой ин-
вплоть до распада, а при похолоданиях - наобо-
ститут «Наука и образование». Ежемесячный науч-
рот, пчелы уменьшают размеры своей агрегации.
ный журнал, № 3, 116 (2014).
При высоких температурах пчелы могут перехо-
5. С. В. Оськин и Д. А. Овсянников, Биофизика 64
дить уже в активный режим с повышенным по-
(1),153 (2019).
треблением меда и начинают выращивать рас-
6. В. А. Тобоев, Пчеловодство, № 1, 20 (2007).
плод. В это время, если наступает похолодание,
7. А. И. Касьянов, Пчеловодство, № 2, 16 (2003).
пчелы собираются вокруг расплода и интенсивно
его обогревают, и могут возникнуть проблемы с
8. А. Д. Трифонов, Пчеловодство, № 11, 21 (1990).
невозможностью взять мед на дальних рамках. В
9. А. Ф. Рыбочкин и И. С. Захаров, Компьютерные си-
такие периоды пчеловоды стараются применять
стемы в пчеловодстве, 2-изд. (Курский гос. техн.
электрообогрев с автоматическим включением
ун-т., Курск, 2004).
нагревателей. Однако управление электрообогре-
10. С. В. Оськин и Д. А. Овсянников, Электротехноло-
вом по датчикам температуры может привести к
гические способы и оборудование для повышения
значительному энергопотреблению и провока-
производительности труда в медотоварном пчело-
ции пчел на раннюю закладку расплода. Полу-
водстве Северного Кавказа (Изд-во ООО «Крон»,
ченные модели показали, что, действительно,
Краснодар, 2015).
БИОФИЗИКА том 65
№ 5
2020
МОДЕЛИРОВАНИЕ ВЕСЕННЕЙ АГРЕГАЦИИ ПЧЕЛ В УЛЬЕ
985
Simulation of Bees’ Aggregation in the Hive in Spring when Daily Outdoor Air
Temperature Fluctuates a Great Deal
S.V. Oskin and D.A. Ovsyannikov
Kuban State Agrarian University named after I.T. Trubilin, ul. Kalinina 13, Krasnodar, 350044 Russia
There is a lot of research reporting on bee wintering process that shows that the state of bee family during win-
tering is yet to be understood. However, the most critical time for bees is late winter and spring. There are
large fluctuations of temperature in the air during this period stimulating bees to occasionally become active
and switch to the ectothermic state and vice versa. This phenomenon has mostly been observed in the apiar-
ies, which cultivate the bees in the North Caucasus region. With springtime strong positive air temperature
tendency, bees fly from the hive, and not all bees return to the hive by the end of the day. Low temperatures
may lead to aggregation of bees in awkward places where there is neither food nor direct access to the honey
reserves. This study focuses on bee behavior under these natural conditions. The main physical effects were
simulated using the Comsol® software. The components associated with transient processes were added to
the previously developed models, which include the heat capacity of the beehive elements. The models also
addressed the change in the volume of bees’ winter aggregation depending on the ambient temperature. Our
simulation results suggest that with spring air temperature fluctuations, the heat capacity of honey reserves
has a significant effect on the temperature regime inside the hive. Honey-produced energy in the form of heat
mitigates temperature fluctuations inside the hive. This is an important factor in honey bee health.
Keywords: bee aggregation, hive, temperature, electric heating, simulation
БИОФИЗИКА том 65
№ 5
2020