БИОФИЗИКА, 2020, том 65, № 5, с. 986-993
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 57.01+573+575+576.7+611-013+611.018
ПРЕДСТАВЛЕНИЕ ТРЕХМЕРНОЙ СТРУКТУРЫ ЭПИТЕЛИЯ
КАК РЕГУЛЯРНОЙ КЛЕТОЧНОЙ СЕТИ
(НА ПРИМЕРЕ ГЛАЗА ДРОЗОФИЛЫ)
© 2020 г. Г.А. Савостьянов
Институт эволюционной физиологии и биохимии им. И.М. Сеченова РАН,
194223, Санкт-Петербург, просп. Тореза, 44
E-mail: genasav38@mail.ru
Поступила в редакцию 19.03.2020 г.
После доработки 23.05.2020 г.
Принята к публикации 25.05.2020 г.
На примере анализа пространственной организации сетчатки дрозофилы показано, что структуру
клеточных пластов можно представлять трехмерными клеточными решетками, имеющими вид эта-
жерок. Такие решетки отражают топологию клеточных пластов и являются новым предметом ги-
стологии. Для их исследования предложен специальный подход, заключающийся в синтезе семей-
ства моделей пространственной организации тканей, сравнении тканевых срезов с сечениями мо-
делей и выбору той из них, которая соответствует реальности. Этот подход существенно облегчает
реконструкцию и открывает путь к построению трехмерной вычислительной гистологии.
Ключевые слова: топология тканей, клеточная решетка, коннектом, компьютерное моделирование,
трехмерная гистология, глаз дрозофилы.
DOI: 10.31857/S0006302920050166
Современная гистология в значительной мере
влияет на работу практических гистологов. Таким
остается плоскостной. Это - следствие того, что
образом, трехмерная гистология пока не создана,
основным методом исследования в ней служит
а закономерности эволюционного становления
изучение тонких срезов, не отражающих морфо-
пространственной организации тканей остаются
логические и функциональные взаимосвязи кле-
неизвестными. Это сдерживает понимание зако-
ток в тканевом пространстве. Несмотря на появ-
номерностей трансформации тканей в патологии
ление новых методов микроскопии, компьютер-
и, в частности, в морфогенезе опухолей.
ных технологий трехмерной реконструкции, а
Ранее нами были предприняты шаги к ликви-
также количественной гистологии [1] знания о
дации этого пробела путем дальнейшего развития
трехмерной структуре тканей явно недостаточны.
теории и пополнения семейства тканевых моде-
Конечным результатом исследования до сих пор
лей [9-12]. Она основана на представлениях о
служат микрофотографии (или рисунки) плос-
том, что ткани состоят не из клеток как таковых,
костных картин, которые видны в микроскоп, их
а из клеточных групп, возникающих в результате
качественное описание или морфометрическая
разделения функций между клетками. Такие
характеристика. Более того, в современной опи-
группы получили название гистионов. Полиме-
сательной гистологии не выделены главные пара-
ризация гистионов дает клеточные сети различ-
метры, которые следует изучать при реконструк-
ной размерности (1D, 2D и 3D), состава и струк-
ции пространственной организации тканей, и ос-
туры. Такое представление позволяет сформули-
новные усилия направляются на выяснение
ровать более адекватный набор аксиом и строить
геометрии клеток и формы ткани как континуу-
расширенные семейства топологических и гео-
ма, но не на характер клеточных взаимосвязей и
метрических моделей гистоархитектуры. Предла-
образуемых ими сетей. Разработка моделей для
гаются также два варианта компьютерного пред-
исследования этих вопросов проводится в рамках
ставления моделей: в виде полиэдров, отражаю-
математической теории строения биологических
щих геометрию клеток, и в виде клеточных
тканей [2-8]. Однако пока она основана на из-
решеток, отражающих топологию клеточных се-
лишне жесткой аксиоматике и предлагает огра-
тей. Для их анализа в реальных тканях был разра-
ниченный набор моделей тканевой структуры,
ботан специальный подход, который заключается
поэтому в существующем виде эта теория слабо
в сравнении тканевых срезов с сечениями моде-
986
ПРЕДСТАВЛЕНИЕ ТРЕХМЕРНОЙ СТРУКТУРЫ ЭПИТЕЛИЯ
987
лей и выбору той из них, которая соответствует
этих сечениях единый двенадцатиугольный ком-
реальности. Этот подход показал свою эффектив-
плекс из светопреломляющих, ретинальных и
ность при трехмерной реконструкции ряда по-
первичных пигментных клеток обозначен буквой
кровных и сенсорных эпителиев и позволил
А, гексагональные вторичные и тетрагональные
определить строение их клеточных сетей [9-13].
третичные пигментные клетки - буквами B и С
соответственно, гексагональные механорецепто-
Целью настоящей работы было изучение с по-
ры - буквой D. Внутреннее строение этих клеток
мощью разработанного подхода пространствен-
также не учитывается. Другими словами, в нашем
ной организации сетчатки дрозофилы. Первый
рассмотрении элементы моделей считались «не-
шаг, направленный на исследование геометрии
прозрачными» и характеризовались лишь внеш-
ее клеток, был сделан нами ранее [14]. В настоя-
ней формой, имеющей вид полиэдров с выров-
щей работе приводятся результаты изучения то-
ненными гранями. Наконец, не учитывалось на-
пологии ее клеточных сетей.
личие синапсов и нервных волокон. Такая
упрощенная схема строения сетчатки на продоль-
МАТЕРИАЛЫ И МЕТОДЫ
ном разрезе, с отмеченными уровнями a—c, пред-
ставлена в нижней части на рис. 1а.
Глаз мухи Drosophila melanogaster отличается
высокой упорядоченностью расположения омма-
Построенные схемы служили основой для со-
тидиев, что делает его удобным объектом для ис-
здания трехмерных моделей сетчатки, которое
следования. Особенно это относится к глазу заро-
проводили с использованием разработанной на-
дыша мухи, имеющему более простую геомет-
ми компьютерной программы «Гистоарх» [20].
рию. Изучению структуры глаза у зародышей и
Для этого в качестве исходной принимали двух-
взрослых особей посвящено множество работ, из
мерную мозаику b на рис. 1б, которая приближа-
которых мы используем лишь несколько [15-19].
ется к геометрически правильной форме. С помо-
В соответствии с ними в каждом омматидии раз-
щью «Гистоарха» проводили непрерывное преоб-
личают три основные части: преломляющую или
разование этой мозаики и получали серию
диоптрическую (хрусталик и кристаллический
геометрических вариантов и топологических
конус), воспринимающую или рецепторную (ре-
трансформаций мозаик, показанных на рис. 1в.
тинальные клетки) и изолирующую, состоящую
Эти мозаики отражали геометрию и смежность
из пигментных клеток. При этом хрусталик с кри-
клеток на различных уровнях сетчатки, от апи-
сталлическим конусом, все ретинальные и две
кального до базального, Полученные двумерные
первичных пигментных клетки совместно обра-
мозаики превращали в решетки, показанные на
зуют единый комплекс [19]. В дальнейшем его
рис. 1г. В соответствии с работой [3] это делалось
внутреннее строение мы детализировать не бу-
следующим образом: центры полигонов обозна-
дем. Такие комплексы изолированы друг от друга
чали кружками, а связи между ними (непосред-
вторичными и третичными пигментными клетка-
ственное соседство) - отрезками. Такие решетки
ми, а также механорецепторами. Их взаиморас-
отражают без геометрических деталей состав (с
положение на разных уровнях сетчатки в настоя-
помощью оттенков цвета) и топологию клеточ-
щее время представляется в виде различных двух-
ных сетей на соответствующих уровнях пласта.
мерных схем. При этом трехмерная форма
На основании полученной серии мозаик с по-
составляющих глаз клеток, их численные соотно-
мощью «Гистоарха» строили трехмерные модели в
шения, смежность, а также клеточная сеть, отра-
полиэдрах, отражающие геометрию отдельных
жающая топологию глаза, на схемах не отражены
клеток сетчатки, а также их объединения в масси-
и остаются неизвестными. Выяснение этих во-
просов с помощью моделей и составляло нашу за-
вы. Кроме того, на основании полученных реше-
ток, соединяя их одноцветные кружки стержнями
дачу.
(отрезками того же цвета), строили трехмерную
Исходным материалом для построения трех-
модель сети в виде «этажерки», которая отражала
мерных моделей сетчатки служили обобщенные
топологию ткани в трехмерном пространстве. Оба
результаты морфологических исследований нор-
варианта моделей имели векторную природу. Эти
мального глаза дрозофилы, представленные в ви-
модели сравнивали с имеющимися данными о
де двухмерных схем различной полноты [16, 18].
строении реальной сетчатки. Для этого использо-
Эти схемы были сведены нами в единую картину
вали предложенный ранее [10] комплекс призна-
(рис. 1).
ков: клеточный состав, численные соотношения
В верхней части рис. 1а показано существую-
клеток, их смежность и микроокружение. Для
щее представление о строении омматидия заро-
большей информативности построенные модели и
дыша дрозофилы на продольном разрезе. На
их сечения группировали в наглядные композиции
рис. 1б дается его апикальная поверхность
с помощью программы Adobe Photoshop®. По-
(по [15]), и картины поперечных сечений на уров-
скольку в нашем случае высокой регулярности сет-
нях a—c, имеющих вид двухмерных мозаик. На
чатки и ее моделей эти признаки являются неслу-
БИОФИЗИКА том 65
№ 5
2020
988
САВОСТЬЯНОВ
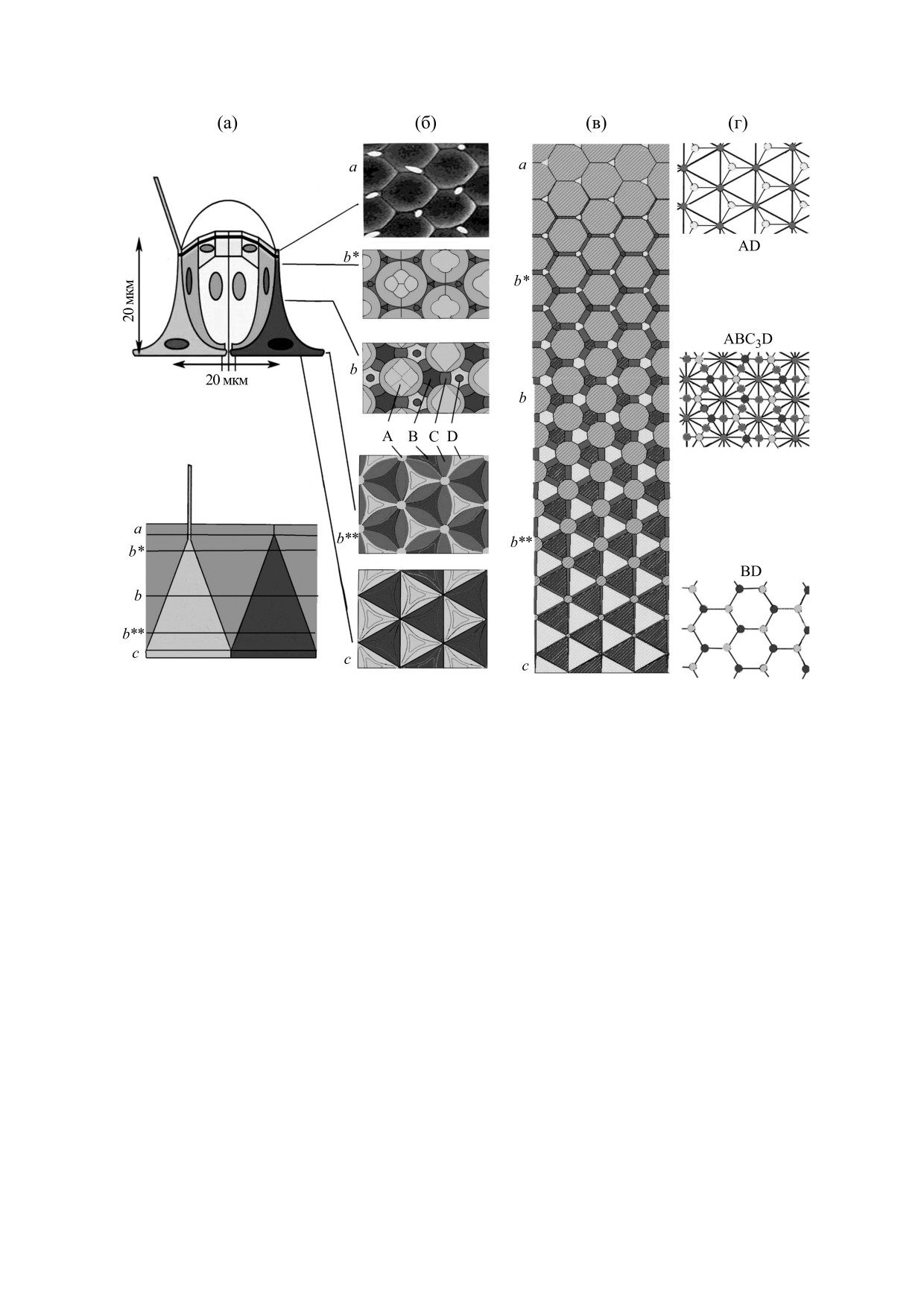
Рис. 1. Существующие двухмерные представления о строении омматидия зародыша дрозофилы. (а) - Вверху показано
существующее представление о строение омматидия зародыша дрозофилы на продольном срезе. Внизу дана его упро-
щенная схема, дополненная верхним и нижним уровнями. Третичная пигментная клетка на срез не попала. (б) - Вид
поперечных сечений омматидия на уровнях а-с (буквы b со звездочками обозначают мозаики с той же топологией, что
и мозаика b, но отличающиеся от них геометрически); A - «непрозрачный» комплекс первичных пигментных и рети-
нальных клеток («радужка»), B и C - вторичные и третичные пигментные клетки, D - механорецептор. (в) - Аппрок-
симация поперечных сечений омматидия двумерными мозаиками на уровнях а-с. (г) - Представление топологии мо-
заик регулярными клеточными решетками с указанием их клеточного состава и смежности.
чайными, статистическая обработка результатов
ментных клеток C. Наконец, на базальном уровне
сравнения не требовалась.
c мозаика представлена треугольными профилями
клеток B и D. Эти мозаики аппроксимируют с точ-
ностью до топологии сечения омматидиев, схема-
РЕЗУЛЬТАТЫ
тически показанных на рис. 1б. Таким образом,
несмотря на обилие геометрических модификаций
Результаты трансформации исходной мозаики,
мозаик на уровнях b, b* и b**, их общий набор
полученные с помощью «Гистоарха», представле-
включает в себя только три топологических вари-
ны на рис. 1в набором регулярных двумерных мо-
заик. На апикальном уровне a они состоят из девя-
анта, обозначенных буквами a, b и c.
тиугольных профилей комплексов А и треуголь-
Клеточные решетки, отражающие состав и то-
ных профилей щетинок механорецепторов D.
пологию этих мозаик без геометрических дета-
Расположенная ниже основная часть мозаик
лей, представлены на рис. 1г. Поскольку они ха-
(уровни b, b*, b**) состоит из двенадцатиугольных
профилей комплексов А, гексагональных профи-
рактеризуются высокой регулярностью строения
лей механорецепторов D и вторичных пигментных
(трансляционной симметрией), можно выделить
клеток B, а также тетрагональных третичных пиг-
их элементарные ячейки и определить их состав,
БИОФИЗИКА том 65
№ 5
2020
ПРЕДСТАВЛЕНИЕ ТРЕХМЕРНОЙ СТРУКТУРЫ ЭПИТЕЛИЯ
989
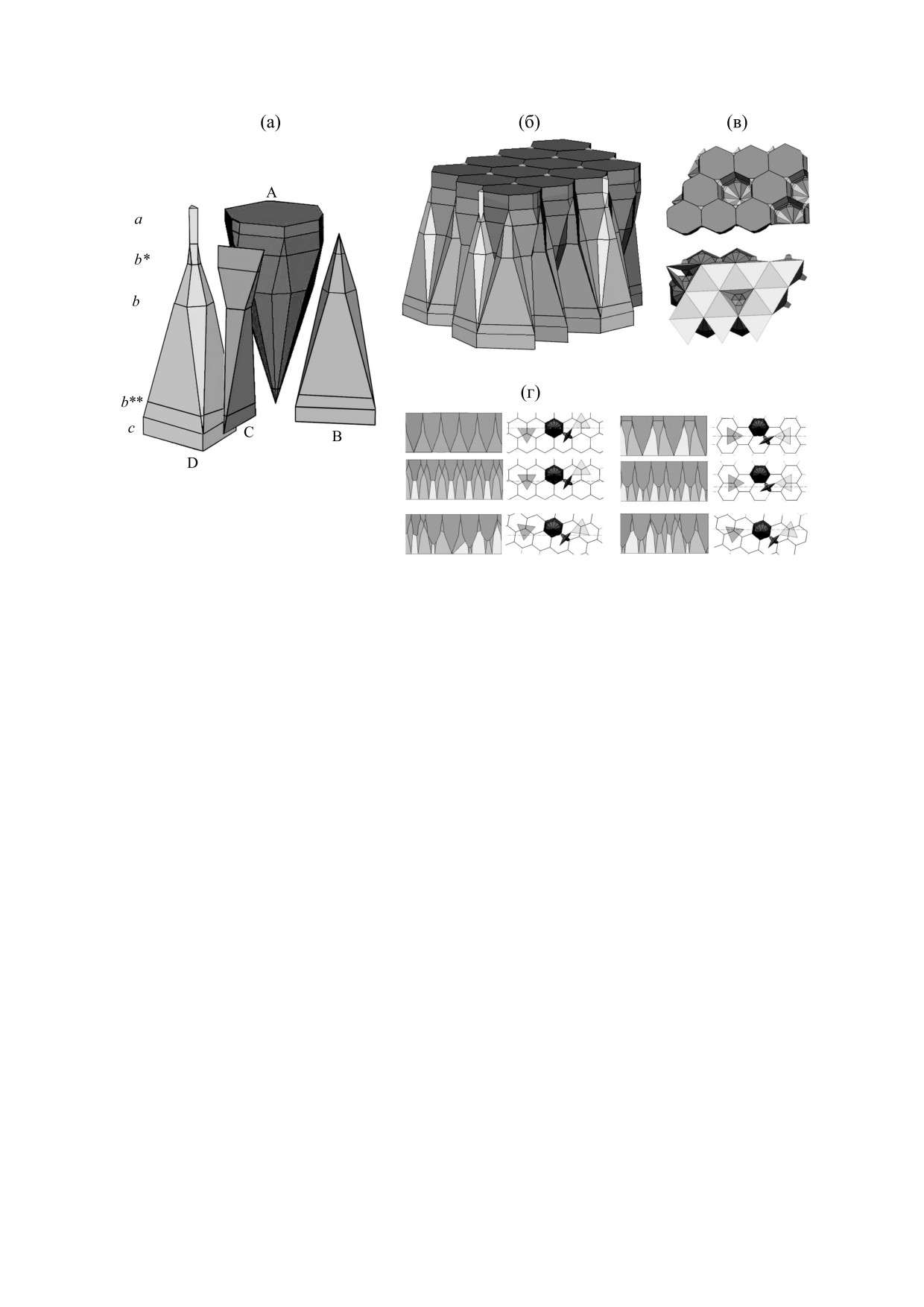
Рис. 2. Трехмерные модели пространственной организации сетчатки зародыша дрозофилы состава АВС3D. (а) -
Геометрия клеток и их взаиморасположение. Буквенное обозначение клеток повторяется. (б) - Внешний вид
клеточного массива. (в) - Строение внутриэпителиальных клеточных ниш. Вверху - ниши омматидиев, внизу -
вторичной пигментной клетки. (г) - Базально-апикальные сечения массива, проведенные в различных направлениях.
Можно видеть, как сильно зависит получаемая картина от направления и глубины сечения.
как это описано ранее [10]. Для мозаик a, b и c он
ня b*. Наконец, на поверхности глаза находится
будет равен АD, ABC3D и BD соответственно.
мозаика a (рис. 1, уровень а), состоящая из топо-
логических девятиугольников (комплексы А) и
Описанные мозаики (рис. 1в) представляют со-
треугольников (механорецепторные сенсиллы) в
бой упрощенные поперечные сечения сетчатки.
соотношении
1/1. Эта мозаика соответствует
На основании их набора с помощью компьютер-
уровню a модели.
ной программы «Гистоарх» была построена трех-
мерная модель сетчатки, представленная в левой
Нетрудно видеть, что по составу, численным
части рис. 2а. Она показывает геометрию клеток
соотношениям (АD, ABC3D и BD), геометрии и
различных типов в виде полиэдров, число их ребер
и граней, а также клеточное взаиморасположение.
смежности клеток, характеру микроокружения
Видно, что только механорецепторы D выходят и
каждой клетки, а также регулярности строения
на апикальную, и на базальную поверхности. Ком-
(трансляционной симметрии) все мозаики на се-
плексы А достигают только апикальной, а вторич-
чениях омматидиев и модели полностью совпада-
ные пигментные клетки B - только базальной по-
ют. Это позволяет заключить, что модель отража-
верхности. Третичная пигментная клетка С не до-
ет трехмерное строение сетчатки дрозофилы. В
стигает ни одной из этих поверхностей и находится
частности, модель впервые позволяет точно опи-
в глубине пласта. На построенной модели отмече-
сать микроокружение каждого элемента сетчатки
ны также уровни сечений, соответствующие раз-
в трехмерном пространстве. Так, каждый «непро-
личным мозаикам сетчатки. Например, нижнее ее
зрачный» комплекс A связан с тремя вторичными
сечение с на рис. 1 соответствует сечению c трех-
пигментными клетками B, тремя механорецепто-
мерной модели.
рами D и с шестью третичными пигментными
Следующие сечения омматидия соответствуют
клетками C. Каждая вторичная пигментная клет-
уровням b** и b модели. Последовательность гео-
метрических изменений профилей элементов
ка и механорецептор связаны с тремя комплекса-
сетчатки на более высоких уровнях также отража-
ми A и тремя третичными пигментными клетка-
ется моделью, что можно видеть на примере уров-
ми. Последние связаны с двумя комплексами A,
БИОФИЗИКА том 65
№ 5
2020
990
САВОСТЬЯНОВ
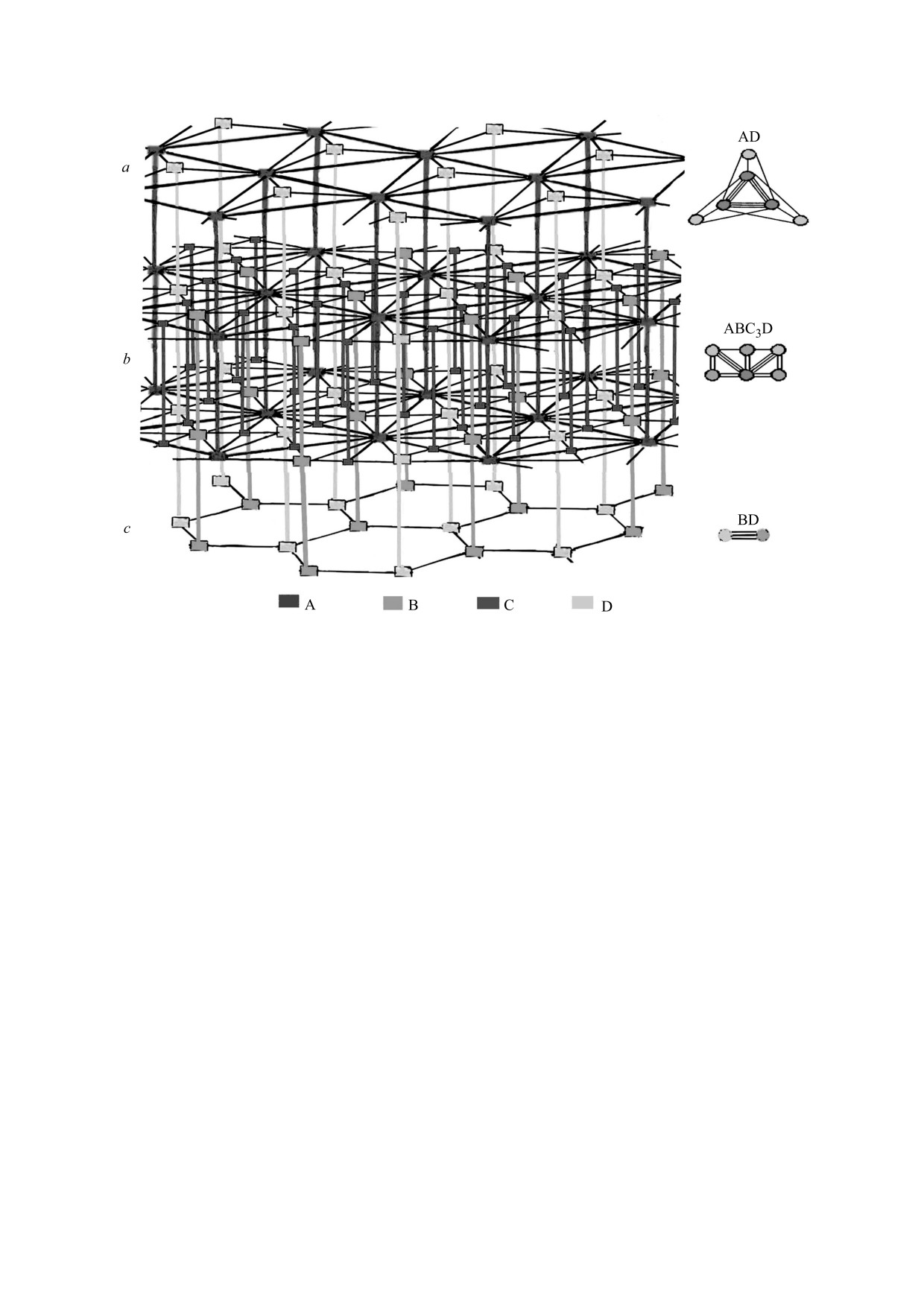
Рис. 3. Топологическая характеристика пространственной организации сетчатки. Слева - клеточная решетка
(коннектом) в виде «этажерки», отражающей топологию ткани без геометрических деталей на уровнях a-c. Для
наглядности средний уровень b растянут и заключен между двумя идентичными двумерными решетками. Справа
показаны гистионы, отражающие состав и смежность клеток в двумерных решетках каждого уровня «этажерки».
Гистион среднего уровня b является репрезентативным, поскольку он наиболее полно отражает клеточный состав
решетки в целом.
вторичной пигментной клеткой и механорецеп-
С помощью программы «Гистоарх» выполня-
тором D. Модель описывает и чередование раз-
ются сечения клеточного массива в различных
личных контактов. Такая точность описания до-
направлениях и оценивается их информатив-
стигнута впервые. Кроме того, модель предсказы-
ность. Например, показанные на рис. 1в аппрок-
симирующие мозаики отражают косое сечение
вает то, каким образом должно меняться
модели. Сечения, получаемые в апикально-ба-
строение сетчатки на различных уровнях.
зальном направлении, показаны на рис. 2г. По-
Из описанных клеток с помощью компьютер-
скольку пласт характеризуется анизотропией, ре-
ной программы «Гистоарх» можно построить
зультат таких сечений очень сильно зависит от их
клеточный массив и исследовать его поверхность.
направления и глубины и дает различные карти-
Общий вид такого массива показан на рис. 2б.
ны. Таким образом, компьютерная реализация
Удаляя из него отдельные клетки, можно рас-
модели показала, что определить геометрию кле-
ток и строение клеточной решетки путем иссле-
смотреть строение внутриэпителиальных клеточ-
дования базально-апикальных и плохо ориенти-
ных ниш, которые обеспечивают вышеописанное
рованных срезов массива весьма сложно.
микроокружение удаленных клеток. Эти ниши
для комплексов A и механорецептора D показаны
Построенная модель не только отражает гео-
на рис. 2в. Отметим, что ниша клеток B практиче-
метрию клеток, но и содержит в себе строение
ски совпадает с нишей механорецептора. Иссле-
трехмерной клеточной сети сетчатки. Набор
довать строение ниш впервые оказалось возмож-
двухмерных решеток, показанных на рис. 1г, поз-
ным благодаря применению нашего подхода.
воляет построить модель клеточной сети сетчатки
БИОФИЗИКА том 65
№ 5
2020
ПРЕДСТАВЛЕНИЕ ТРЕХМЕРНОЙ СТРУКТУРЫ ЭПИТЕЛИЯ
991
в явном виде. Для этого, как было сказано выше,
ние в различных тканях должно быть включено в
кружки двумерных решеток, соответствующие
число главных задач создающейся трехмерной
одним и тем же клеткам, нужно объединить с по-
гистологии.
мощью вертикальных отрезков в виде стержней.
При кажущейся сложности «этажерок» полез-
Возникающая в итоге трехмерная решетка приоб-
ную вспомогательную роль для характеристики
ретает вид «этажерки», которая отражает тополо-
коннектома пласта могут играть гистионы. Бу-
гию ткани в трехмерном пространстве (рис. 3).
дучи более простыми, они дают почти всю ин-
На рис. 3 стержни - это различные клетки сет-
формацию о каждом уровне «этажерки». Но одно
чатки, простирающиеся на различные уровни, го-
ее важное свойство они не отражают. Это свой-
ризонтальные отрезки между стержнями отража-
ство - ориентационный порядок решеток, т.е.
ют клеточную смежность на различных уровнях. В
направления межклеточных связей и углы, под
построенной таким путем «этажерке» выделяются
которыми эти связи осуществляются. Таким об-
три уровня: a, b и c с решетками различного соста-
разом, гистион, оставаясь полезным понятием,
ва и топологии. Решетка каждого уровня характе-
не может полностью заменить клеточную решет-
ризуется своим гистионом (показаны справа от
ку и «этажерку».
«этажерки»). Гистионы - это элементарные струк-
Кроме топологического описания сетчатки,
турные единицы двумерных решеток. Они ком-
была продолжена характеристика формы ее кле-
пактно отражают состав, численные соотношения
ток, начатая ранее [14]. Наряду с уточнением их
и смежность (микроокружение) клеток в решет-
геометрии, с помощью «Гистоарха» был показан
ках. Например, легко видеть, что репрезентатив-
общий вид клеточного массива и его поверхно-
ный (наиболее полный по составу) гистион «эта-
стей. Также впервые показано строение в нем
жерки» имеет состав ABC3D, а смежность его кле-
внутриэпителиальных клеточных ниш и дана их
количественная характеристика (т.е. форма и
ток показана числом отходящих от них отрезков.
число граней различных клеток, формирующих
В целом можно сказать, что трехмерная модель
каждую нишу, а также порядок их чередования).
из полиэдров, построенная на основании сече-
Важно также то, что с помощью компьютерных
ний омматидиев, совместно с клеточной решет-
моделей была проведена оценка информативно-
кой в виде «этажерки» дают полное описание гео-
сти сечений массива, сделанных в различных на-
метрии клеток сетчатки и топологии ее клеточ-
правлениях. Эксперименты с моделями показа-
ной сети. Отметим, что глаз взрослой дрозофилы
ли, как разнообразно и порой обманчиво может
выглядеть ткань на различных срезах в зависимо-
имеет ту же топологию и отличается лишь геомет-
сти от их ориентации. Опираясь на такие срезы,
рической модификацией его клеток. Отметим
трудно судить о клеточном составе и простран-
также, что при специальном исследовании глаза
ственной организации пласта. В итоге найдено,
модель можно сделать более подробной и учесть
что наиболее информативными являются танген-
внутреннее строение комплекса А, т.е. диоптри-
циальные или слегка скошенные срезы, а менее
ческой и фоторецепторной частей омматидия.
информативными - срезы в базально-апикаль-
ном направлении. Для описания строения таких
срезов ранее был предложен комплекс новых ин-
ОБСУЖДЕНИЕ
формативных признаков [10].
Главный смысл данной работы состоит в том,
Построенные модели можно надстроить свер-
что в ней впервые в явной форме и точно установ-
ху так, чтобы щетинки исчезли, и тогда апикаль-
лена топология сетчатки, представленная моде-
ная мозаика будет состоять только из гексаго-
лью в виде трехмерной клеточной решетки. Такая
нальных профилей комплекса А. Надстроенные
решетка имеет вид «этажерки». Эта модель пред-
модели будут давать полное описание возможной
ставляет без геометрических деталей наиболее су-
гистоархитектуры сетчатки. Они позволят судить
щественную черту строения сетчатки, а именно:
о ее мозаиках даже на тех уровнях, которые не бы-
ее клеточную сеть, т.е. все взаимосвязи клеток в
ли исследованы экспериментально (благодаря
этой способности модели позволяют проводить
трехмерном пространстве пласта. Поэтому мы
реконструкцию реальной ткани при меньшем ко-
назвали такую решетку тканевым коннектомом,
личестве ее срезов). В то же время в реальных тка-
расширив первоначальный смысл этого термина
нях в отдельные периоды (в развитии, при повре-
(взаимосвязи не только нейронов мозга, но и кле-
ждении) описанная гистоархитектура может реа-
ток в различных тканях). Современная плоскост-
лизовываться и не полностью. Это имеет место и
ная гистология пока упускает этот аспект, что яв-
в нашем случае: в отличие от модели в реальной
ляется важным пробелом. Этот пробел заполняет
сетчатке две клетки доходят до апикальной по-
предложенная модель коннектома, который яв-
верхности, и все клетки доходят до базальной по-
ляется новым предметом гистологии. Его изуче-
верхности. Это означает, что в данном случае
БИОФИЗИКА том 65
№ 5
2020
992
САВОСТЬЯНОВ
верхняя и нижняя части модели в сетчатке не ре-
ФИНАНСИРОВАНИЕ РАБОТЫ
ализованы. Однако реальный пласт может до-
Работа выполнена в рамках Государственного
страиваться как сверху, так и снизу. Тогда модель
задания № 075-00776-19-02.
сможет прогнозировать то, как будет изменяться
строение сетчатки при более полной ее реализа-
ции. Еще одним различием модели и реальной
КОНФЛИКТ ИНТЕРЕСОВ
сетчатки является то, что все ее клетки обычно
Авторы заявляют об отсутствии конфликта
подвергаются геометрической модификации.
интересов.
Так, в верхней части пласта комплексы А являют-
ся выпуклыми, а вторичные пигментные клетки
В, механорецепторы D и смежные с ними третич-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
ные клетки С - вогнутыми. В нижней части (на
уровне b**) клетки С становятся уже выпуклыми.
Настоящая работа не содержит описания ка-
Возможность геометрической модификации кле-
ких-либо исследований с использованием людей
ток при сохранении топологии пласта уже обсуж-
и животных в качестве объектов.
далась ранее [10]. Такая модификация не затраги-
вает коннектом, является переменчивой и может
СПИСОК ЛИТЕРАТУРЫ
отражать физиологические особенности клеток в
различные периоды их жизнедеятельности.
1. А. Г. Никоненко, Введение в количественную ги-
стологию (Книга-плюс, Киев, 2013).
В целом полученные результаты ранжируют
2. F. T. Lewis, Amer. Sci. 34 (3), 359 (1946).
по важности задачи, связанные с изучением про-
странственной организацией клеточных пластов
3. В. В. Смолянинов, Математические модели биоло-
любого состава. Прежде всего следует определять
гических тканей (Наука, М., 1980).
топологию тканей (их коннектомы) как наиболее
4. K. J. Dormer, Fundamental tissue geometry for biologist
существенной и постоянной их характеристики.
(Cambridge Univ. Press, London, 1980).
Затем с помощью морфометрии можно прово-
5. В. М. Маресин, Пространственная организация эм-
дить изучение особенностей геометрической ва-
бриогенеза (Наука, М., 1990).
риабельности клеток в различных условиях.
6. R. Nagpal, A. Patel, and M. C. Gibson, BioEssays 30
(3), 260 (2008).
ЗАКЛЮЧЕНИЕ
7. F. Graner and D. Riveline, Development 144, 4226
(2017).
Впервые установлена топология сетчатки гла-
8. K. Goodwin and C. M. Nelson, Exp. Cell Res. 358, 45
за дрозофилы, т.е. структура ее клеточной сети.
(2017).
Такая сеть представляет собой регулярную трех-
9. Г. А. Савостьянов, Биофизика 46 (3), 512 (2001).
мерную решетку и имеет вид «этажерки». Она вы-
10. Г. А. Савостьянов, Основы структурной гистоло-
ступает как новый способ характеристики про-
гии. Пространственная организация эпителиев.
странственной организации тканей и количе-
(Наука, СПб., 2005).
ственного описания их коннектомов. Кроме того,
описана геометрия клеток сетчатки и строение
11. Г. А. Савостьянов, Изв. РАН, сер. биол., № 2, 164
внутриэпителиальных клеточных ниш. Это стало
(2012).
возможным благодаря применению нового под-
12. Г. А. Савостьянов, Цитология 58 (8), 577 (2016).
хода к реконструкции пространственной органи-
13. Е. Г. Магницкая, Н. М. Грефнер, Т. Б. Голубева и
зации тканей, основанного на компьютерном мо-
др., Сенсорные системы 23 (4), 334 (2009).
делировании. Этот подход радикально улучшает
14. Г. А. Савостьянов, Сенсорные системы 15 (2), 121
результаты реконструкции пространственной ор-
(2001).
ганизации сетчатки и исследования топологии ее
клеточных сетей. Он способен также прогнозиро-
15. D. F. Ready, T. E. Hanson, and S. Benzer, Develop.
вать направления тканевого развития. Уже гово-
Biol. 53 (2), 217 (1976).
рилось, что его применение к реконструкции
16. T. Wolff and D. F. Ready, Development 113 (3), 825
трехмерного строения ряда покровных и сенсор-
(1991).
ных эпителиев также оказалось результативным.
17. D. T. Miller and R. L. Cagan, Development 125 (12),
Можно надеяться, что применение этого подхода
2327 (1998).
к изучению и других тканей будет способствовать
18. S. Hilgenfeldt, S. Erisken, and R. W. Carthew, Proc.
пониманию их перестроек в нормальном разви-
Natl. Acad. Sci. USA 105 (3), 907 (2008).
тии и патологии. Развитие исследований в этом
направлении может в конечном счете привести
19. R. Cagan, Curr. Top. Dev. Biol. 89, 115 (2009)
к построению трехмерной эволюционной гисто-
20. Е. Г. Савостьянова, А. В. Воробьев, Н. М. Грефнер
логии.
и др. Морфология 131 (4), 8 (2007)
БИОФИЗИКА том 65
№ 5
2020
ПРЕДСТАВЛЕНИЕ ТРЕХМЕРНОЙ СТРУКТУРЫ ЭПИТЕЛИЯ
993
Representation of a Three-Dimensional Epithelial Structure as a Cell Regulatory Network
in a Drozophila Eye Model
G.A. Savostyanov
Sechenov Institute of Evolutionary Physiology and Biochemistry, Russian Academy of Sciences,
prosp. Toreza 44, St. Petersburg, 194223 Russia
Analysis of the spatial arrangement of the Drosophila retina has shown that a structure of cell layers can be
represented by three-dimensional cell lattices, they resemble a stack. Such lattices simulate the topology of
cell layers and are a new subject of histology. The proposed approach to study these lattices is to perform syn-
thesis of a series of models of the spatial arrangement of tissues, compare biological tissue sections and model
sections and choose the one that has the most natural structure. This approach greatly eases the reconstruc-
tion and opens the way for the development of the computational three-dimensional histology.
Keywords: tissue topology, cell lattice, connectom, computer modeling, reconstruction, three-dimensional histolo-
gy, Drosophila eye
БИОФИЗИКА том 65
№ 5
2020