БИОФИЗИКА, 2020, том 65, № 6, с. 1171-1183
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 577.3
КЛАССИФИКАЦИЯ АДАПТИВНЫХ ПРИЗНАКОВ И ВЫВОД КРИТЕРИЕВ
ИХ ЭВОЛЮЦИОННОЙ ОПТИМАЛЬНОСТИ
© 2020 г. Л.Л. Овсянников*, М.И. Шпитонков*, **
*Первый Московский государственный медицинский университет имени И.М. Сеченова МЗ РФ,
119991, Москва, Трубецкая ул., 8/2
E-mail: tatiana-chap@yandex.ru
**Вычислительный центр им. А.А. Дородницына Федерального исследовательского центра «Информатика
и управление» РАН, 119333, Москва, ул. Вавилова, 44/2
E-mail: mixash@bk.ru
Поступила в редакцию 01.07.2019 г.
После доработки 12.08.2020 г.
Принята к публикации 14.08.2020 г.
Предложена математическая модель динамики биологического сообщества, которая представляет со-
бой систему уравнений, описывающих динамику переменных экосистемы - плотность численности
популяции; вес тела половозрелых особей; плотность энергии, получаемой особью с пищей, - и учи-
тывает возрастную структуру популяции потребителя. С использованием результатов по конкурен-
ции, полученных для точечных моделей другими исследователями, получены алгоритмы определения
эволюционно оптимальных значений адаптивных признаков половозрелых особей популяции. Пока-
зано, что в ходе конкуренции за пищевой ресурс популяций, отличающихся между собой величинами
набора признаков, выживает только одна с набором признаков, обеспечивающим максимум функции
размножения, что соответствует общему критерию эволюционной оптимальности. Проведена класси-
фикация признаков по типам, показано, что каждому типу соответствует свой критерий эволюцион-
ной оптимальности. Предложены простые алгоритмы поиска эволюционно оптимальных значений
различных адаптивных признаков и веса тела, определено оптимальное распределение энергозатрат
на все виды жизнедеятельности организма. Численным экспериментом проиллюстрирована эволю-
ционная оптимальность этих признаков.
Ключевые слова: эволюционная оптимальность, устойчивость равновесного состояния, конкуренция
популяций, адаптивные признаки.
DOI: 10.31857/S0006302920060174
признаков являются оптимальными и в обычном
Действие естественного отбора на популяцию
смысле, а именно, представляют экстремум для
организмов проявляется посредством конкурен-
некоторых оценочных функций. Эти оценочные
ции популяций за ресурс. Для того чтобы орга-
низмы наилучшим образом приспособились к
функции составляют суть критериев эволюцион-
условиям обитания, природа снабдила их адап-
ной оптимальности адаптивных признаков. В об-
тивными признаками. Предполагается, что адап-
зорных работах [13, 15, 16] рассматривается мно-
тивные признаки имеют такие величины, кото-
жество моделей, в которых определяются опти-
рые способствуют популяции-носителю этих ве-
мальные величины различных адаптивных
личин признаков победе в конкуренции. Такие
признаков. Для этого авторы предлагали разные
значения признаков будем называть эволюцион-
критерии, смысл которых в том, что существуют
но оптимальными значениями (ЭОЗ). Среди наи-
целевые функции, которым оптимальные значе-
более известных публикаций, посвященных
ния адаптивных признаков доставляют экстрему-
определению оптимальных значений адаптивных
мы. При этом зачастую ошибочно утверждалось,
признаков в свете дарвиновской теории есте-
что найденные оптимальные значения являются
ственного отбора, можно отметить работы [1-13].
продуктом эволюции. Наиболее популярным
оказался энергетический критерий, суть которого
В работе [14] сформулирован принцип опти-
мальной конструкции, согласно которому опти-
в том, что адаптивный признак имеет такое ЭОЗ,
мальные в смысле естественного отбора значения
при котором энергозатраты на его создание и
функционирование минимальны. Среди наибо-
Сокращениe: ЭОЗ - эволюционно оптимальные значения.
лее известных публикаций, посвященных энерге-
1171
1172
ОВСЯННИКОВ, ШПИТОНКОВ
тическому критерию, можно выделить работы
n(i + 1) = A(f (i)) ⋅n(i),
(1.1)
[2-8, 12].
Согласно дарвиновской теории эволюции, а
где A(f (i)) - матрица Лесли, имеющая вид
также принципу конкурентного исключения Гау-
(i)
f f f
f
f
n
зе, величина адаптивных признаков является
0.0
1
продуктом естественного отбора, в процессе ко-
(i)
σ
0.0.0.0..0.0
1
n
2
торого выживает популяция носителей призна-
(i)
ков данной величины. Многие авторы при этом
0.σ .0.0.0..0.02
n
3
полагали, что эволюционные значения призна-
0.0..s.0.0..0.0
(i)
ков доставляют экстремум некоторым целевым
(i
)
n
4
A(f) =
; n
=
,
функциям [1-3, 5-13, 17].
0.0.0..s.0..0.0
(i)
n
5
Но многообразие различных критериев опти-
0.0.0.0
s
(i)
мальности, отсутствие связи между критериями,
n
6
а также отсутствие доказательств соответствия
s
0
0.0.0.0.0..
(i)
этих критериев теории естественного отбора
n
7
представляют существенный дефект теории эво-
где t0 = 2; T = 7; f - плодовитость особей поло-
люционной оптимальности признаков организма
возрелых когорт (одинакова для всех когорт),
и свидетельствует о незавершенности этого на-
правления.
nj(i) - численность j-й возрастной когорты (j =
Настоящая работа посвящена установлению
= t0 + 1, …, T) в i-й момент времени; σj - выжива-
связи между критериями оптимальности и соот-
емость ювенильных когорт. Общая выживае-
ветствия этих критериев теории эволюции (а
t0
именно, показано существование единого крите-
мость потомков выражается формулой w = ∏σi,
рия эволюционной оптимальности; установлена
1
связь частных критериев с единым; доказано со-
s
j - выживаемость особей взрослых когорт (упро-
ответствие этих критериев дарвиновской теории
щенно полагаем одинаковой (равной s) во всех
естественного отбора). Проведена классифика-
когортах.) Выживаемость s определяем выраже-
ция адаптивных признаков по типам. Было пока-
нием
зано, что каждому типу признаков соответствует
свой критерий эволюционной оптимальности.
s = exp(-dh),
(1.2)
Доказано, что эти критерии являются следствием
1
единого для всех, общего критерия эволюционной
где d - смертность взрослых когорт; [d] =
;
оптимальности, впервые сформулированного в
сутки
h - шаг интегрирования, (h = 1 сутки).
работах [4, 6, 8]. Предложен алгоритм поиска
ЭОЗ адаптивного признака вкупе с весом тела ор-
Рассмотрим подробно, от чего зависит смерт-
ганизма.
ность на примере приполярных животных. При-
полярные территории имеют низкие температу-
На адаптивных признаках конкретных попу-
ляций животных (например, толщине шерстного
ры окружающей среды. Это приводит к гибели
покрова лис Аляски и скорости плавания пелаги-
животных от переохлаждения. Для того чтобы
ческих рыб) показана справедливость изложен-
противодействовать этому, приполярные живот-
ной в работе теории.
ные (песцы, лисы, олени) имеют шерстный по-
кров. Чем толще покров, тем меньше вероятность
гибели. В результате зависимость смертности от
1. ДИНАМИКА ЭКОСИСТЕМЫ
толщины шерстного покрова α определим фор-
С ВОЗРАСТНОЙ СТРУКТУРОЙ
мулой
ПОПУЛЯЦИИ (МОДЕЛЬ ЛЕСЛИ)
Da
Рассмотрим простейший биоценоз, включаю-
da =
(1.3)
δ
α
щий популяцию организмов. Энергия в количе-
стве V поступает в биоценоз в виде корма. Пусть
Зависимость смертности от массы тела можно
t0 - возраст наступления половой зрелости, T -
представить аналогичным выражением:
средняя продолжительность жизни особей. Осо-
DP
би возраста τ, где τ ∈ [0; t0], назовем ювенильны-
dP =
,
(1.4)
γ
ми, особи возраста t, где t ∈ (t0; T], - взрослыми.
P
В результате имеем t0 ювенильных когорт и T - t0
где P - масса тела взрослой особи. Например, чем
взрослых.
больше масса тела, тем меньше вероятность гибе-
Динамику численности популяции опишем
ли от хищников. Da, δ, DP, γ - коэффициенты.
следующей системой:
Общую смертность определим формулой
БИОФИЗИКА том 65
№ 6
2020
КЛАССИФИКАЦИЯ АДАПТИВНЫХ ПРИЗНАКОВ
1173
D
D
a
dP
P
-
P
d = da + dP =
+ P.
(1.5)
i+1
i
ем
=
, где h - шаг интегрирования,
αδ Pγ
dt
h
В том случае, если одна смертность много
h = [d]-1 = 1 сутки; сb - удельная калорийность
больше другой, последней можно пренебречь.
тела особи. Упрощенно будем полагать, что Wb
Определим по формуле (1.1) плотности чис-
равна нулю (для взрослых когорт).
ленностей когорт на (i + 1)-м временном шаге
В итоге уравнение энергетического баланса
n(i+1).
примет вид
Энергетический баланс организма. Энергию,
B = W0 + Wf+ Wa+ Wb,
(1.13)
поступающую в организм с пищей, определим
выражением:
или в конкретном виде
V
B = χPβ + c f (i) + Kaαλ.
(1.14)
B =
,
(1.6)
N
Плотность численности взрослых особей попу-
где V - пищевая энергия, приходящая на единицу
ляции. Условно виды можно разделить на два ти-
площади биоценоза; N - плотность численности
па: 1-й тип - виды, у которых родители заботятся
взрослых особей, которую определяем формулой
о потомстве; 2-й тип - виды, у которых родители
не заботятся о потомстве. Для видов 1-го типа ха-
T
(
рактерна высокая выживаемость (w ≈ 1). При
N =
(1.7)
этом будем полагать, что ресурс потребляют толь-
t
+1
ко взрослые особи, а ювенильная часть популя-
Величина B пропорциональна плотности ре-
ции потребляет ресурс от родителей. В этом слу-
сурса R и определяется выражением
чае численность взрослых особей будет равна
B = mR,
(1.8)
V
N =
,
(1.15)
где m - коэффициент пропорциональности. За-
B
метим, что если адаптивный признак способству-
ет добыванию пищи, то m = αq, где α - величина
где V - плотность пищевой энергии, [ккал/(сутки · м2].
адаптивного признака; q - коэффициент. В ре-
Для видов 2-го типа характерна низкая выжи-
зультате формула (1.8) примет вид
ваемость (w ≈ 0). Взрослые особи потребляют
B = αqR.
(1.8a)
только некоторую часть пищевой энергии, дру-
гую часть потреблеют ювенильная доля популя-
Эта энергия расходуется на различные виды
ции. В этом случае численность взрослых особей
жизнедеятельности.
будет равна
W0 - затраты на основной обмен и передвиже-
Vϕ
ние. Определяются выражением
N =
,
(1.16)
B
W0 = χPβ.
(1.9)
где ϕ - доля пищевой энергетической плотности,
Wf - затраты на воспроизводство. Определя-
поглощаемой взрослыми особями популяции.
ются выражением
Динамика биоценоза. Пусть σj - выживаемость
ювенильных когорт (j = 1, …, t0); sj = s -выживае-
Wf = cf.
(1.10)
мость взрослых когорт, одинакова для всех ко-
Wa - затраты на функционирования адаптив-
горт; f - плодовитость особей, одинакова для всех
ного признака α. Определяются выражением
взрослых когорт.
Рассмотрим популяцию видов 2-го типа (ро-
(1.11)
Wa = Kaαλ.
дители заботятся о потомстве).
Wb - энергозатраты на привес. Определяются
Динамика численности возрастных когорт в
формулой
(i + 1)-й момент времени определяется выраже-
нием (1.1).
dP
Wb= сb
(1.12)
Находим численность взрослых особей попу-
dt
ляции:
Поскольку выражение (1.1) представляет дис-
(i+1)
n
dP
N(i+1) =
j
(1.17)
кретную модель, то производную
следует
dt
Определяем количество пищевой энергии, по-
представить в разностном виде. В результате име-
лучаемой взрослой особью:
БИОФИЗИКА том 65
№ 6
2020
1174
ОВСЯННИКОВ, ШПИТОНКОВ
2. ДИНАМИКА ЭКОСИСТЕМЫ БЕЗ УЧЕТА
V
B(i+1) =
(1.18)
ВОЗРАСТНОЙ СТРУКТУРЫ ПОПУЛЯЦИИ
(i+1)
N
(ТОЧЕЧНАЯ МОДЕЛЬ)
Из формулы (1.14) выразим плодовитость на
Динамику численности популяций предста-
(i + 1)-м временном шаге:
вим уравнением
dN
f (i+1) = (B(i+1) - a0Pβ - Kaαλ)/c.
(1.19)
= F(f,P,a)N.
(2.1)
dt
По известному значению f (i+1) определяем
Здесь N - численность популяции, F - коэф-
фициент размножения, определяемый формулой
матрицу Лесли A(f (i+1)) на (i+1)-м временном
шаге.
F = f -
d,
(2.2)
Далее по формуле (1.1) определяем численно-
где смертность d фиксирована и равна
f
, выра-
сти когорт на следующем временном шаге, затем
женной формулой (1.21).
повторяем операцию. При i →∝ экосистема при-
ходит в равновесное состояние. При этом
Поскольку выражение (1.1) представляет дис-
кретную модель, то в модельных расчетах произ-
n(i+1) = n(i) и выражение (1.1) станет
dN
водную
следует представить в разностном
n(i) = A(f (i)) ⋅ n(i),
(1.20)
dt
виде, а именно,
а равновесное значение плодовитости будет
dN
N
-
N
равно
i+1
i
=
,
(2.3)
dt
h
^
(1
-
s)
f
=
τ
,
(1.21)
0
где h - шаг интегрирования, h = [d]-1 = 1 сутки;
w(1
−
s
)
с
b - удельная калорийность тела особи.
Энергетический баланс особи. Количество пи-
a
D DP. ) - выживаемость
щевой энергии, приходящейся на одну особь, со-
где s = exp(-d) = exp(-
δ
ставляет
α
Pγ
t0
B = V/N.
(2.4)
взрослых когорт, w = ∏σi - общая выживае-
Уравнения энергетического баланса были
1
мость ювенильных когорт, τ0 = T - t0 - продол-
представлены выражениями (1.13) и (1.14). Пред-
ставим их заново:
жительность жизни взрослой особи.
B = W + Wf+ Wa+ Wb,
(2.5)
В приложении
1
представлен вывод
формулы (1.21) другим способом.
B = αPβ + cf + Kaαλ.
(2.6)
Рассмотрим важное упрощение формулы (1.21). В
В результате динамика экосистемы, представ-
том случае, если d достаточно мало то, отбросив
ленная точечной моделью, имеет вид:
sτ0 и разложив s в ряд (s ≈ 1 - d), получаем
N(i+1) = N(i) + N(i) (f(i) -
d).
(2.7)
f
≈
d/w.
(1.22)
где N(i+1) - численность популяции на (i+1)-м
Равновесное значение пищевой энергии, потреб-
временном шаге.
ляемой взрослой особью, составляет
^
B(i+1) = V/N(i+1).
(2.8)
B
= с
f
+ W0(P) + Wa(a).
(1.23)
Из уравнения (2.6) находим
Равновесное значение плотности численности
f (i+1) = (B(i+1) - a0Pβ - Kaαλ)/c.
(2.9)
взрослых особей определяется выражением
^
V
Подставив f(i+1) в уравнение (2.7), получаем
N = .
^
(1.24)
численность популяции на следующем вре-
B
менном шаге, затем повторяем операцию. В
Итак, система уравнений (1.1), (1.17)-(1.19)
результате при i → ∝ экосистема приходит к
описывает динамику экосистемы по модели Лес-
равновесному состоянию F =
f
-
d
= 0, где
ли, а система (1.21)-(1.24) - ее равновесное со-
стояние.
f
- равновесная плодовитость, соответству-
БИОФИЗИКА том 65
№ 6
2020
КЛАССИФИКАЦИЯ АДАПТИВНЫХ ПРИЗНАКОВ
1175
ющая формуле (1.26), т.е.
Здесь W0(j) - энергозатраты на основной об-
τ
(1
-
s)
мен плюс передвижение при добывании пищи.
f
=
=
(2.10)
d
τ
0
Они определяются следующим выражением [2]:
w(1
−
s
)
W0(j) = χ(Pj)0.75,
(3.5)
Отметим, что поскольку выживаемость s зависит
от веса P и адаптивного признака a, равновесная
где χ
- коэффициент пропорциональности,
плодовитость является функцией P и a (
f
f
= (P,a)).
[ккал/(кг0.75 сутки)].
f
Подставив (P,a) в уравнение (2.6), получаем рав-
Wf(j) - энергозатраты на воспроизводство,
новесное значение потребляемой пищевой энер-
определяемые формулой
гии:
Wf(j) = c fj,
(3.6)
B
f
= χPβ + (P,a) c + Ka aλ.
(2.11)
где c - затраты на одного потомка.
Равновесная численность популяции определя-
ется выражением
Wa(j) - энергозатраты на функционирование
адаптивного признака a:
N
= V/B.
(2.12)
Как видим, асимптотические решения обеих
Wa(j) = Ka aj.
(3.7)
моделей динамики экосистемы (модели Лесли и
Подставив выражения (3.5), (3.6) и (3.7) в вы-
точечной модели) идентичны. Поэтому в даль-
ражение (3.4), получаем
нейшем исследования по конкуренции популя-
ций будем проводить, используя более простую
B = χ(P(j))0.75 + c fj + Ka aj.
(3.8)
точечную модель.
Уравнения (3.1)-(3.8) описывают динамику
экосистемы, содержащей две конкурирующие
3. КОНКУРЕНЦИЯ ПОПУЛЯЦИЙ
популяции. Нетрудно показать, что эта система
НА ОСНОВЕ ТОЧЕЧНОЙ МОДЕЛИ
имеет два нетривиальных равновесных состоя-
ния: равновесие 1
Рассмотрим экосистему, в которой сосуще-
ствуют две популяции, отличающиеся величина-
(1)
N
>
0;
F
1
=
0
ми какого-либо адаптивного признака и массой
(3.9a)
тела. (Рассуждения без особого труда могут быть
(2)
N
=
0;
F
≠
0
2
обобщены на случай k популяций и m адаптивных
признаков.)
и равновесие 2
Динамику этих популяций опишем уравнени-
(1)
ями:
N
=
0;
F
1
≠
0.
(3.9a)
dNj
(2)
N
>
0;
F
=
0
2
= FjNj; j = 1, 2.
(3.1)
dt
Динамическая модель конкуренции представ-
ляет собой наглядную демонстрацию эволюции
1
-
s
j
адаптивных признаков, когда в ходе конкуренции
Здесь Fj = fj - gj; gj=
τ
; sj = exp(-dj);
0
w(1
−
s
)
выживает популяция-носитель наиболее опти-
j
мальных значений этих признаков.
dj = dja + djP, где dja = Da/aj - смертность, обуслов-
В работах [6, 8] было доказано, что из совокуп-
ленная адаптивным признаком a, djP = DP/Pj -
ности равновесных состояний асимптотически
смертность, обусловленная массой тела.
устойчивым будет то, которое удовлетворяет
условию:
Общая численность популяций равна
Fj =
max(F
,
F
)
=
0.
(3.10)
2
1
2
P,a
N =
Nj
(3.2)
Выражение (3.10) представляет собой общий
j
=1
критерий эволюционной оптимальности, суть ко-
Пищевая энергия, потребляемая одной осо-
торого в том, что в процессе конкуренции за ре-
бью, одинакова для обеих популяций и равна
сурс популяций, отличающихся величинами
адаптивных признаков и массы тела особей, вы-
B = V/N.
(3.3)
живает только одна популяция, особи которой
Энергетический баланс особи этих популяций
обладают такими значениями перечисленных
имеет вид:
адаптивных параметров, при которых коэффици-
ент размножения достигает своего максимума,
B = W0(j) + Wf(j) + Wa(j).
(3.4)
равного нулю в состоянии равновесия экосисте-
БИОФИЗИКА том 65
№ 6
2020
1176
ОВСЯННИКОВ, ШПИТОНКОВ
мы. Это соответствует широко известному в эко-
Если же адаптивный признак является «ней-
логии принципу конкурентного исключения
тральным» (первый тип), то для нахождения его
Гаузе.
эволюционно оптимального значения применя-
ем энергетический критерий. Суть его в том, что в
Из общего критерия эволюционной опти-
конкуренции побеждает та популяция, особи ко-
мальности следуют три частных. Это зависит от
торой потребляют наименьшее количество энер-
того, к какому из трех перечисленных типов при-
гии на создание и функционирование этого ней-
надлежит признак.
трального признака, т.е. справедливо выражение
Классификация адаптивных признаков. Услов-
но можно выделить следующие три типа адаптив-
(3.14)
Wa(j) = min(Wa(1), Wa(2)).
ных признаков. Первый тип - «нейтральные»
Энергетический критерий впервые был при-
адаптивные признаки. Они не влияют на смерт-
менен в работе [14] при расчете оптимальных па-
ность и на плодовитость, а также не способствуют
раметров кровеносной системы. В обзорных ра-
добыванию пищи. Но, тем не менее, без них су-
ботах [15, 16] а также в работе [13] приведена мас-
ществование организма невозможно (например,
са примеров применения этого критерия для
угол ветвления кровеносных сосудов). Второй
определения оптимальных значений различных
тип - адаптивные признаки, влияющие на смерт-
признаков организма. При этом зачастую оши-
ность и/или плодовитость (например, шерстный
бочно утверждалось, что эти признаки также яв-
покров у приполярных животных уменьшает их
ляются продуктами эволюции.
смертность от переохлаждения). Третий тип -
признаки, способствующие добыче пищи (на-
Резюмируя приведенные выводы, предложим
пример, длинная шея жирафа позволяет ему пи-
следующую схему связей адаптивных признаков
таться высоко растущими листьями).
и критериев оптимальности (рис. 1).
Если адаптивный признак принадлежит ко
второму типу (влияет на плодовитость или смерт-
4. АДЕКВАТНОСТЬ МОДЕЛЕЙ
ность), то условие (3.10) эквивалентно следующе-
ЭКОСИСТЕМЫ (МОДЕЛИ ЛЕСЛИ
му условию (доказательство в приложении 2):
И ТОЧЕЧНОЙ МОДЕЛИ)
B
Покажем адекватность моделей динамики
j
=
min(
1
B,B
2
).
(3.11)
P,a
экосистемы с учетом возрастной структуры (мо-
дель Лесли) и без ее учета (точечная модель).
Выражение (3.11) представляет собой полный
энергетический критерий эволюционной опти-
Рассмотрим гипотетическую экосистему с по-
мальности, суть которого в том, что в процессе
пуляцией пустынных животных, у которых адап-
тивным признаком являются большие уши (на-
конкуренции за ресурс популяций, отличающих-
ся величинами адаптивных признаков и массы
пример лисица фенек). С помощью ушей у них
тела особей, выживает только одна популяция,
происходит понижение температуры тела, что
та, особи которой обладают такими значениями
уменьшает смертность от перегрева. Как видим,
перечисленных параметров, при которых равно-
этот адаптивный признак относится ко второму
типу. Зависимость смертности от величины α
весное количество пищевой энергии, потребляе-
мое особью, минимально.
определяется формулой
В том случае, если адаптивный признак при-
dα = Dα/α,
(4.1)
надлежит третьему типу (способствует добыче
где α - площадь ушей особи в дм2. Кроме того,
пищи), выражение (3.10) эквивалентно выраже-
пусть существует хищник, питающийся этими
нию
ушастыми особями, причем чем выше масса, тем
R
j
=
min(
1
R,R
2
).
(3.12)
меньше смертность от хищников. Тогда зависи-
P,a
мость смертности от массы тела определим выра-
Доказательство аналогично доказательству
жением
выражения (3.11). Отсюда из выражения (1.8a) по-
dP = DP/P
(4.2)
лучаем
B
B
(здесь δ = γ =1 из формулы (1.5)).
1
R
2
R
=
;
2
=
(3.13)
Зададим конкретные значения коэффициен-
qα
1
qα
2
= 2 временные единицы
там модели. Пусть t0
Выражение (3.12) назовем критерием миниму-
(в.е.); T = 7 в.е., Dα = 0.223 дм2/(в.е.), DP =
ма плотности ресурса. Этот критерий созвучен
высказыванию Мак-Артура [17], что естествен-
= 0.8 кг/(в.е.), σ1 = 0.4 (в.е.)-1, σ2 = 0.5 (в.е.)-1,
ный отбор способствует выживанию такого хищ-
c =200 ккал/потомок, χ = 100 ккал/(кг0.75⋅(в.е.)),
ника, который питается наименьшим количе-
ством жертв.
Кα = 300 ккал/(дм2⋅(в.е.)), V = 14 ккал/(м2⋅(в.е.)).
БИОФИЗИКА том 65
№ 6
2020
КЛАССИФИКАЦИЯ АДАПТИВНЫХ ПРИЗНАКОВ
1177
Рис. 1. Схема связей адаптивных признаков и критериев оптимальности.
Рассмотрим популяцию, взрослые особи кото-
На рис. 2 и 3 представлена динамика числен-
ности популяций по точечной модели и по моде-
рой имеют следующие параметры: α = 0.7 дм2,
ли Лесли.
P = 4 кг.
Вычислим значения промежуточных парамет-
Расчеты показали, что при t → ∝ f → 2.19,
ров:
B → 930.2, N → 0.01505. Таким образом, числен-
ный эксперимент показал, что в обеих моделях
dα = 0.32, dP = 0.2, d = 0.52, s = 0.595, w = 0.2
экосистема приходит к одному и тому же состоя-
нию равновесия. Эти же значения получаем из
f
и равновесные значения переменных:
=
d
=
решения алгебраической системы уравнений рав-
новесного состояния экосистемы (2.10)-(2.12).
B
= 2.19, Wf = 437.9, W0 = 282.8, Wα = 210,
=
Алгоритмы поиска эволюционно оптимальных
N
= 930.2,
= 0.01505 особей/м2.
значений. В результате получаем простой алго-
Теперь рассмотрим результаты численного
ритм определения эволюционно оптимальных
моделирования динамики экосистемы (с теми же
значений массы тела и адаптивного признака,
параметрами) по точечной модели и по модели
суть которого состоит в определении точки
Лесли.
(P*, a*). Для этого в формуле (2.11) найдем точку
Рис. 2. Динамика численности популяции по точеч-
Рис. 3. Динамика численности популяции по модели
ной модели при α = 0.7 дм2; P = 4 кг.
Лесли при α1 = α2 = 0.7 дм2; P1 = P2 = 4 кг.
БИОФИЗИКА том 65
№ 6
2020
1178
ОВСЯННИКОВ, ШПИТОНКОВ
первая популяция, поскольку ее особи потребля-
ют наименьшее количество пищевой энергии.
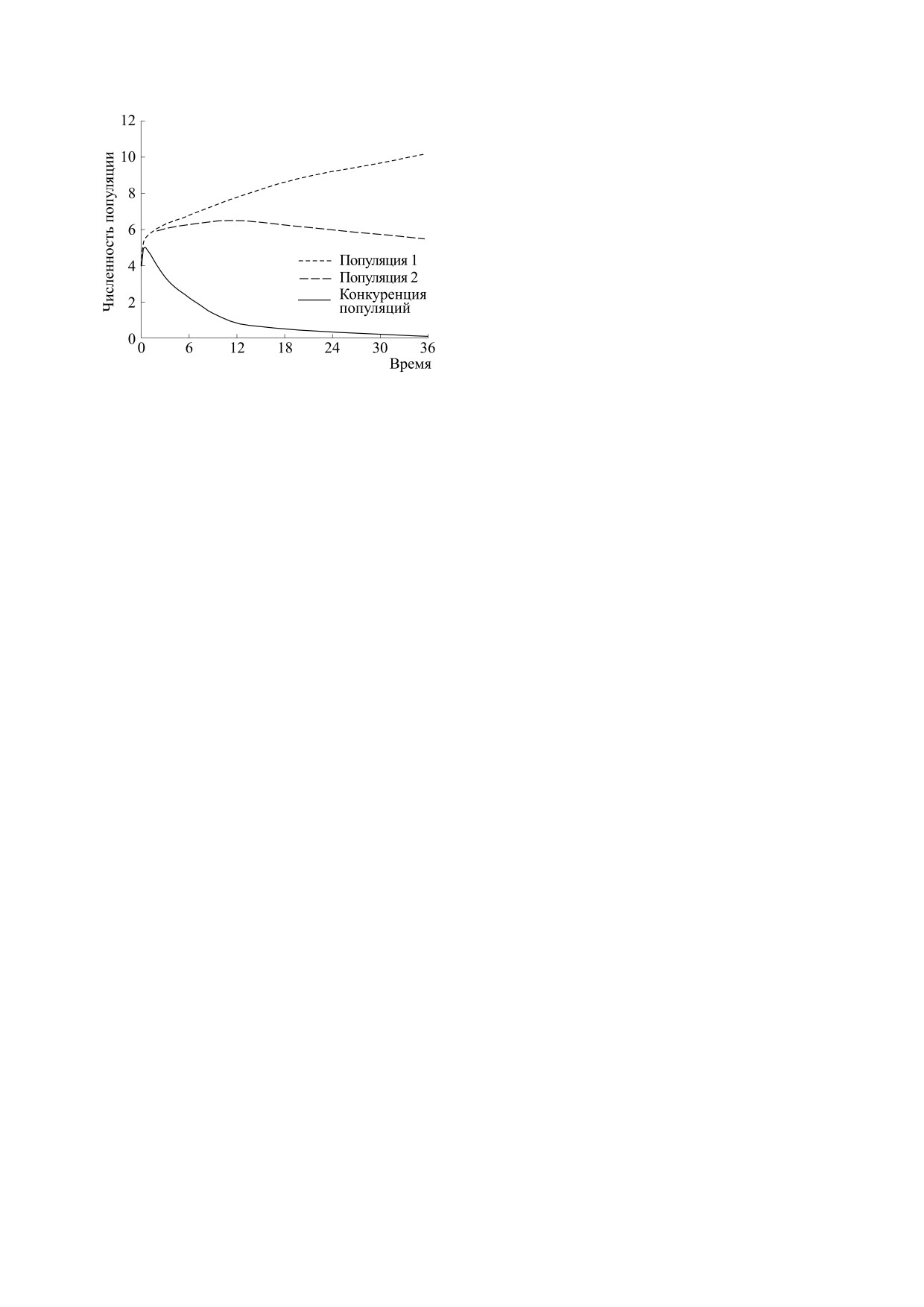
Из рис. 4 видно, что действительно побеждает
первая популяция. Расчеты показали, что ее чис-
ленность N1 → N* = 15.65, а численности N2 → 0
и N3 → 0.
Таким образом, для нахождения ЭОЗ призна-
ка мы можем не решать систему дифференциаль-
ных уравнений конкуренции, а применить один
из перечисленных критериев эволюционной оп-
тимальности (в зависимости от типа адаптивного
признака).
5. ПРИМЕРЫ ЭВОЛЮЦИОННО
ОПТИМАЛЬНЫХ ЗНАЧЕНИЙ
Рис. 4. Динамика численности конкурирующих по-
АДАПТИВНЫХ ПРИЗНАКОВ КОНКРЕТНЫХ
пуляций.
ЖИВОТНЫХ
В этом разделе вначале рассмотрим примеры
B
ошибочного применения энергетического крите-
(P*, a*), доставляющую минимум значению
рия для определения эволюционно оптимальных
Таким образом, задача свелась к поиску экстре-
значений некоторых адаптивных признаков.
мума функции двух переменных при конкретных
значениях параметров.
В работе [2] в качестве адаптивного признака
рассматривался шерстный покров арктических
Можем с помощью модели конкуренции убе-
лис. Для определения эволюционно оптималь-
диться в справедливости полного энергетического
ной величины шерстного покрова авторы приме-
критерия как на точечной модели, так и на моде-
нили энергетический критерий, ошибочно пола-
ли Лесли. Продемонстрируем это на примере на-
гая, что полученное значение будет также эволю-
шей экосистемы. C помощью программы Mathcad
ционным. Так, на стр.
206 авторы пишут:
находим оптимальные α* = 0,6 и P* = 2.5, доставля-
«Естественно предположить, что в ходе эволюции
ющие минимум B. Соответствующая оптимальная
селективные преимущества имеют особи, у кото-
плодовитость будет: f* = 2.58 (в.е.)-1. Подставив
рых параметры термоизоляционного слоя обес-
найденные значения в формулы (3.5), (3.6) и
печивают минимальный расход энергии…». Та-
(3.7), находим оптимальное распределение энер-
ким образом, энергетический критерий опти-
гии, полученной особью с пищей по всем видам
мальности термоизоляции имеет вид:
жизнедеятельности: W0* = 198.8, Wf = 515.5,
W = WT + Wh = min,
(5.1)
Wα* = 180. Просуммировав эти значения, опреде-
где WT - теплоотдача особи, которая определяет-
ляем полное количество энергии, получаемое
ся выражением
особью с пищей: B* = 894.3 ккал/(в.е.). Плотность
численности популяции будет равна N* = 15.65
WT = (ε/h) S(Ts - Te),
(5.2)
особей на гектар.
где ε - коэффициент теплопроводности, S - пло-
Убедимся в том, что найденные значения дей-
щадь поверхности тела особи, Ts - температура
ствительно будут оптимальными. Для этого вы-
поверхности тела, Te - средняя зимняя темпера-
числим величину B в окрестности точки (α*, P*).
тура окружающей среды. Обозначим
Пусть точками окрестности будут: α2 = 0.5, P2 = 2
(5.3)
и α3 = 0.7, P3 = 4. Значения переменной B для этих
KT = εS(Ts - Te).
точек будут B2 = B(0.5, 2) = 897,5, B3 = (0.7, 4) =
Подставив (5.3) в выражение (5.2), получаем:
= 930.2, т.е. оба значения больше B*. Итак, B* =
WT = KT/h,
(5.4)
= min(B1, B2, B3), где B1 = B*.
где h - толщина шерстного покрова. Wh - энерго-
Покажем, что значения α1 = α* = 0.6 и P1 =
затраты на генерацию шерсти, их определяем вы-
P* = 2.5 будут также оптимальными в эволюцион-
ражением
ном смысле. Для этого проведем численный экс-
Wh = Khh.
(5.5)
перимент. Рассчитаем на компьютере модель
Подставив выражения (5.4) и (5.5) в (5.1), по-
конкуренции всех трех популяций. Согласно пол-
лучаем:
ному энергетическому критерию эволюционной
оптимальности, победить в конкуренции должна
W = KT/h + Khh = min.
(5.6)
БИОФИЗИКА том 65
№ 6
2020
КЛАССИФИКАЦИЯ АДАПТИВНЫХ ПРИЗНАКОВ
1179
Продифференцировав уравнение (5.6) по h и
популяция-носитель значения h* выиграет кон-
приравняв производную к нулю, получаем опти-
куренцию у популяций с другими значениями h.
мальное значение:
Пусть коэффициенты экосистемы, содержа-
щей популяцию рыжей лисы Аляски, имеют сле-
K
T
дующие значения (отметим, что эти значения
h0 =
(5.7)
K
взяты из работы [2], стр. 207): ε =1.05 ккал/(мг⋅
h
Однако найденное значение не является про-
сутки), Ts = 38°C, Te = -20°C, S = 0.22 м2, P = 5 кг,
дуктом эволюции, поскольку данный признак
w = 0.7, Dh =0.00023 м/сутки, c = 20000 ккал/дете-
влияет на смертность и, следовательно, относит-
ныш, Kh = 5290 ккал/(мг⋅сутки), χ = 70 ккал/
ся не к первому, а ко второму типу. Поэтому для
(кг0.75 сутки), t0 = 2 года, T = 7 лет.
нахождения ЭОЗ этого признака применим соот-
ветствующий полный энергетический критерий.
По формулам
(5.3) и
(5.12a) вычисляем:
Запишем его в виде
KT=13.4 ккал/сутки, Kf = 6.57 ккал/сутки. Затем
B = W0 + Wf + WT + Wh = min,
(5.8)
по формулам (5.7) и (5.14) вычисляем: h0 = 0.05 м,
где B - полные энергозатраты организма на все
h*
= 0.061 м. По формуле
(5.11a) находим
виды жизнедеятельности, W0 - энергозатраты на
f* = 0.0053 сут-1. Полные энергозатраты организ-
основной обмен WA плюс механическое передви-
ма, согласно формуле (5.13), будут составлять
B* = 1048 ккал/сутки.
жение WM; WA = χP0.75, WM = θWA, χ = 70 ккал/
Численный эксперимент по конкуренции. Рас-
(кг0.75 сутки), θ = 0.7 ([2]) . В результате имеем
смотрим три популяции, отличающиеся толщи-
W0 = WA + WM = (1 + θ) WA.
(5.8a)
ной шерстного покрова. Эти популяции обитают
Wf - энергозатраты на воспроизводство, кото-
в одной экологической нише.
рые определяются выражением
Пусть h1 = h0 = 0.05, h2 = h* = 0.061, h3 = 0.07.
Wf = c ,
f
(5.9)
Соответствующие им равновесные значения B
будут равны B1 = 1062, B2 = 1048, B3
= 1054.
где
f
- равновесное значение плодовитости, ко-
торое мы будем определять формулой (1.26a).
Как видим, значению h* соответствует мини-
мальное значение B. На рис. 5 представлены ре-
f = dh/w,
(5.10)
зультаты конкуренции этих трех популяций
где w - выживаемость ювенильных особей, dh -
Из рис. 5 видно, что в конкуренции побеждает,
смертность взрослых особей от переохлаждения.
как и ожидалось, популяция с ЭОЗ величины h.
Определяем ее выражением
Численности популяций с другими значениями
dh = Dh/h.
(5.11)
признака стремятся к нулю.
В итоге
Эволюция адаптивного признака при изменении
Dh
внешних условий. Рассмотрим процесс эволюции
f =
(5.11a)
признака при изменении внешней среды. Пусть
hw
экосистема находится в равновесном состоянии
Подставив выражения (5.11) и (5.10) в (5.9), по-
(f1* = 0.0053сут-1, B1* = 1048 ккал/(особь сутки)
лучаем:
(5.12)
Wf = cDh/wh = Kf /h,
где
Kf = cDh/w.
(5.12a)
В результате выражение (5.8) представим в виде
B = W0 + Kf /h + KT/h + Khh = min.
(5.13)
Для нахождения эволюционно оптимального
значения h продифференцируем выражение
(5.13). Приравняв производную нулю, получаем
ЭОЗ
K
+
K
f
T
h* =
,
(5.14)
K
h
которое отличается от h0 (формула (5.7)).
Убедимся в том, что полученное значение дей-
ствительно эволюционно оптимальное. Для это-
го, используя модель конкуренции, проведем
Рис. 5. Динамика численности популяций (модель
численный эксперимент, в котором покажем, что
конкуренции) с разными значениями признака h.
БИОФИЗИКА том 65
№ 6
2020
1180
ОВСЯННИКОВ, ШПИТОНКОВ
6. АЛГОРИТМ ОПРЕДЕЛЕНИЯ
ЭВОЛЮЦИОННО ОПТИМАЛЬНЫХ
ЗНАЧЕНИЙ ПРИЗНАКА ТРЕТЬЕГО ТИПА
В работе [18] в качестве адаптивного признака
была рассмотрена скорость плавания пелагиче-
ских рыб. Для определения оптимальной скоро-
сти плавания авторы ошибочно применили энер-
гетический критерий. Однако этот признак отно-
сится к третьему типу (способствует добыче
ресурса), поэтому для определения его ЭОЗ необ-
ходимо использовать критерий минимума плотно-
сти ресурса.
Рассмотрим водоем, в котором находится фи-
топланктон плотностью R. Количество пищевого
ресурса (фитопланктона), потребляемого рыбой
за сутки, определяется формулой
Рис. 6. Динамика численности популяций (модель
конкуренции) с разными значениями адаптивного
M = ηvR,
(6.1)
- после).
признака (h1 - до мутации; hМ
где η - площадь забора ресурса, v - средняя ско-
рость плавания рыбы (м/сутки).
Динамика численности популяции рыб опреде-
(при V = 0.0002 ккал/(м2⋅сутки), N1* = 0.019 осо-
ляется выражением
бей/км2), соответствующем значению h1* = 0.061 м
dN
при Tе = -20°C.
= (fw -
d)N,
(6.2)
dt
В какой-то момент произошло потепление, в
где N - плотность численности взрослых особей;
результате которого средняя температура окружа-
f - плодовитость (количество икринок, произве-
ющей среды повысилась до -10°С. При этом ко-
денных самкой за нерест деленное на промежуток
эффициент смертности Dα понизился с 0.00023 до
времени между соседними нерестами); w - выжи-
ваемость (доля икринок, доживших до начала
0.00018. В результате экосистема выйдет из состо-
яния равновесия. Новое ЭОЗ станет равным
взрослого возраста особей);
d
- смертность
h2* = 0.055 м. Ему соответствуют равновесные
взрослых особей.
Энергетический баланс взрослой особи предста-
значения f2* = 0.0066, B2* = 1003.2, N2* = 0.02.
вим выражением
Через некоторое время произошла мутация, в
W = W0 + Wf + WA,
(6.3)
результате которой в популяции появилось не-
где W - полное количество энергии, получаемое
сколько особей с толщиной покрова hM = 0.058 м.
с пищей, которое затем расходуется на различные
Этой толщине соответствуют следующие равно-
виды жизнедеятельности. Определяется выраже-
весные значения: fM
= 0.00575; BM
= 985;
нием
NM = 0.0208. Таким образом, в экосистеме появи-
W = cbM = λvR,
(6.4)
лась популяция-конкурент с меньшим равновес-
где λ = cbη, cb - удельная калорийность ресурса,
ным значением полной потребляемой энергии. В
соответствии с полным энергетическим критерием
W0 - основной обмен, Wf - затраты энергии на
эта популяция должна вытеснить прежнюю из
воспроизводство, которые определяются выра-
экосистемы. В итоге величина адаптивного при-
жением
знака особей станет hM = 0.058 м. На рис. 6 приве-
Wf = сf,
(6.5)
ден результат этой мутации.
в котором c - калорийность одной икринки.
Видно, что численность популяции-мутанта
WA - мощность, расходуемая при плавании.
стремится к NM = 0.0208, в то время как числен-
Определяется выражением
ность первой популяции стремится к нулю. При
WA = Fсv,
(6.6)
дальнейших мутациях в экосистеме сохранится
только популяция с величиной признака
где Fс = Kvv - сила сопротивления при плавании.
h2* = 0.055 м, доставляющей глобальный мини-
В результате имеем
мум равновесному B = B2* = 984.
WA = Kvv2.
(6.7)
БИОФИЗИКА том 65
№ 6
2020
КЛАССИФИКАЦИЯ АДАПТИВНЫХ ПРИЗНАКОВ
1181
Для определения ЭОЗ скорости плавания при-
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
меним критерий минимума плотности ресурса.
Для более детального описания динамики эко-
Подставив формулу (6.7) в выражение (6.3), полу-
системы была применена широко известная в
чаем для равновесного состояния:
экологии модель Лесли. Оказалось, что в равно-
R
весном состоянии экосистемы значения пере-
λv
= W0 + с
f
+ Kvv2,
(6.8)
менных этой модели и значения переменных то-
чечной (без учета возрастной структуры) модели
R
где
- равновесная плотность ресурса,
f
=
d
/w -
совпадают.
равновесная плодовитость. Из выражения (6.8)
Для точечных моделей рядом авторов ([6, 8])
R
выразим
:
была разработана теория (Общий критерий эво-
1W
+
c
f
люционной оптимальности), позволяющая опре-
R
0
=
+
K
v
(6.9)
v
делять эволюционно оптимальные значения
λ
v
адаптивных признаков. В работе показано, что
Определим оптимальное значение v, доставля-
эту теорию можно применить, используя модель,
учитывающую возрастную структуру популяции
ющее минимум R. Для этого продифференциру-
(модель Лесли), что позволяет получить более
ем выражение (6.9) по v. Приравняв производную
адекватные результаты.
нулю, получаем
Оказалось, что адаптивные признаки можно
W
+
c f
классифицировать по типам и для каждого типа
0
v* =
(6.10)
признаков существует свой критерий эволюци-
K
v
онной оптимальности, который является след-
По данным наблюдений в работе [19] за популя-
ствием общего критерия.
цией северо-байкальского омуля компоненты
Показано, что, применяя к определенному
этой формулы имеют следующие значения:
признаку соответствующий критерий оптималь-
W0 = 1.9 ккал/сутки, Wf = c = 0.076 ккал/сутки,
f
ности, можно определить эволюционно опти-
мальное значение этого признака.
KV = 12.24 (ккал/сутки)⋅(с2/м2). Подставив эти
значения в формулу (6.10), получаем ЭОЗ скоро-
Предложенная классификация позволяет из-
сти плавания северо-байкальского омуля
бежать не вполне корректного определения ЭОЗ
признака на основе только одного критерия, кри-
v* = 0.4 м/с, что совпадает с данными наблюде-
терия минимума энергозатрат на функциониро-
ний за природными популяциями.
вание признака. Так ЭОЗ толщины шерстного
Соответствующее минимальное значение Rбудет
покрова лис Аляски [2] и скорости плавания рыб
равно
[18] должны определяться с помощью критерия
минимума потребляемой энергии и критерия ми-
2
нимума плотности ресурса.
R* = min R(v) =
(6.11)
(
Kv(
W +c
f
).
λ
0
Этот результат был обобщен на случай сово-
Подставив выражение (6.10) в (6.7), получаем
купности признаков, в том числе, когда в каче-
формулу активного обмена:
стве признака рассматривался вес тела особи.
Следует отметить, что полученные авторами
f
(6.12)
WA* = W0 + c .
теоретические результаты подтверждаются чис-
ленными экспериментами по определению ЭОЗ
Для рыб энергозатраты на воспроизводство
признаков особей популяции непосредственным
обычно не превышают 4%. Поэтому справедливо
моделированием конкуренции популяций с раз-
следующее выражение:
ными значениями адаптивных признаков.
(6.13)
Отметим так же любопытный факт. Оказалось
WA* ≈ W0.
что при эволюционно оптимальной скорости
Иначе говоря, активный обмен примерно ра-
плавания энергозатраты на плавание равны ос-
вен основному обмену. Этот факт был обнаружен
новному обмену. Этот факт был обнаружен эмпи-
эмпирически в работе [20].
рически в работе [20].
Итак, мы видим, что для определения ЭОЗ
Был проведен численный эксперимент по мо-
адаптивных признаков нам не надо решать трудо-
делированию эволюции адаптивного признака
емкую имитационную модель конкуренции, а до-
(толщины шерстного покрова лис Аляски) при
статочно определить, к какому типу относится
изменении значений параметров среды обита-
данный признак, и применить соответствующий
ния. Были получены удовлетворительные резуль-
критерий эволюционной оптимальности.
таты.
БИОФИЗИКА том 65
№ 6
2020
1182
ОВСЯННИКОВ, ШПИТОНКОВ
ВЫВОДЫ
ПРИЛОЖЕНИЕ 2
Предлагается математическая модель динами-
Пусть функция
ки биологического сообщества. Модель пред-
z = ϕ(x,y)
(п2.1)
ставляет собой систему уравнений, описываю-
щих динамику переменных экосистемы: плот-
задана уравнением:
ность численности популяции; вес тела
F(x,y,z) = 0,
(п2.2)
половозрелых особей; плотность энергии, полу-
причем F(x,y,z) - дифференцируемая функция
чаемой особью с пищей, и учитывает возрастную
структуру популяции потребителя.
переменных x, y, z.
Утверждение. Если
Используя результаты по конкуренции, полу-
ченные для точечных моделей другими исследо-
F(x*,y*,z) =
maxF(x,y,z)
(п2.3)
вателями, получены алгоритмы определения ЭОЗ
x,
y
адаптивных признаков половозрелых особей по-
и
пуляции.
Было показано, что в ходе конкуренции за пи-
Δ > 0, A > 0
щевой ресурс популяций, отличающихся между
2
2
∂
ϕ(x*,y*)
∂
ϕ(x*,y*)
собой величинами набора признаков, выживает
(A
=
,
B
=
,
(п2.4)
2
только одна, та, чей набор обеспечивает макси-
∂x
∂ ∂y
2
мум функции размножения, что соответствует
∂
ϕ(x*,y
2
C =
*),
Δ =
AC -B
),
общему критерию эволюционной оптимальности
2
∂y
(Ханина - Семевского) ([6, 8]).
Проведена классификация прìизнаков по ти-
то
пам. Показано, что каждому типу соответствует
z* = ϕ(x*,y*) = min ϕ (x,y),
(п2.5)
свой критерий эволюционной оптимальности,
x,y
который является следствием общего критерии.
Доказательство. Из условия (п2.3) следует, что
В результате были предложены простые алго-
∂F x*,y*)
∂F ∂ϕ x*,y*)
ритмы поиска ЭОЗ различных адаптивных при-
=
=
0,
знаков и веса тела. А также определено оптималь-
∂x
∂z
∂y
ное распределение энергозатрат на все виды жиз-
недеятельности организма.
∂F x*,y*)
∂F ∂ϕ x*,y*)
=
=
0.
∂y
∂z
∂y
ПРИЛОЖЕНИЕ 1
дF
∂ϕ(x*,y*)
∂ϕ(x*,y*)
Если
≠ 0 , то
=
0
и
=
0
∂
x
∂y
Равновесное значение плотности численности
дz
взрослых когорт составляет
Отсюда следует, что точка (х*,y*) является по-
n
n
ws
j
; j = t0 +1, …, T.
(п1.1)
дозрительной на экстремум для функции
j
=
z = ϕ(x,y).
Прирост численности популяции за один вре-
Но поскольку выполнено достаточное условие
менной шаг равен
экстремума функции (п2.4), то выражение (п2.5)
T-t
0
T−t
0
справедливо. Таким образом, утверждение дока-
n
w
n
n
w1−
s
N+ =
1
j
f
=
1
f
(п1.2)
зано.
1
-
s
1
Убыль численности популяции за один вре-
КОНФЛИКТ ИНТЕРЕСОВ
менной шаг составляет
Авторы заявляют об отсутствии конфликта
n1
N- =
(п1.3)
интересов.
В равновесном состоянии прирост равен убыли.
Приравняв правые части уравнений (п1.2) и
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
(п1.3), получаем:
Настоящая работа не содержит описания ис-
T -t
0
следований с использованием людей и животных
1
−
s
n
w
n
(п1.4)
1
f
=
1
в качестве объектов.
1
-
s
Отсюда
СПИСОК ЛИТЕРАТУРЫ
(1
-
s)
f =
(п1.5)
1. Н. В. Волосова, Математические структуры и
τ
0
w
(1
−
s
)
моделирование 2 (38), 27 (2016).
БИОФИЗИКА том 65
№ 6
2020
КЛАССИФИКАЦИЯ АДАПТИВНЫХ ПРИЗНАКОВ
1183
2. И. Ф. Образцов и М. А. Ханин. Оптимальные био-
11. R. E. Ulanowicz, Ecol. Modelling 220 (16), 1886
механические системы (Медицина, М., 1989).
(2009).
3. Л. Л. Овсянников, Журн. общ. биологии 53 (1), 92
12. Л. Л. Овсянников, Экстремальные принципы в био-
(1992).
логии (Моск. гуманитарный ун-т, М., 2017).
4. Л. Л. Овсянников и В. П. Пасеков. Журн. общ.
13. А.П. Левич, Искусство и метод в моделировании си-
биологии 51 (5),709 (1990).
стем (Москва-Ижевск, 2012).
5. Р. Розен, Принцип оптимальности в биологии (Мир,
14. Н. Рашевски, в сб. Теоретическая и математиче-
М., 1969).
ская биология (Мир, М.,1968), сс. 48-68.
6. Ф. Н. Семевский и С. М. Семенов, Математиче-
ское моделирование экологических процессов (Гидро-
15. П. В. Фурсова, Л. П. Левич и И. Л. Алексеев, Успе-
метеоиздат, Л., 1982).
хи соврем. биологии 123 (2), 115 (2003).
7. А. Т. Терехин и Е. В. Будилова, Журн. общ. биоло-
16. И. В. Волвенко, Биофизика 57 (3), 476 (2012).
гии 62 (1), 286 (2001).
17. Р. Мак-Артур, в сб. Теоретическая и математиче-
8. М. А Ханин, Н. Л. Дорфман, И. Б. Бухаров и
ская биология (Мир, Москва, 1968), сс. 420-430.
В. Г. Левадный, Экстремальные принципы в биоло-
18. А. А. Яржомбек, А. Б. Клянторин и С. Г. Подржан-
гии и физиологии (Наука, М., 1978).
ская, Экология 23 (1), 65 (1977).
9. J. Kozlovski and J. Weiner, Amer. Nat. 149 (1), 423
(1997).
19. И. Б. Волерман и В. В. Конторин, Биологические
сообщества рыб и нерпы в Байкале (Иркутск, 1983)
10. E. L. Charnov, Lafe History Invariants. Some Explora-
tion of Symmetry in Evolutionary Ecology (Univ. Press,
20. Г. Г. Винберг, Журн. общ. биологии 37 (1), 56
Oxford, 1993).
(1967).
Classification of Adaptive Traits and Criteria for Optimality in Adaptive Evolution
L.L. Ovsyannikov* and M.I. Shpitonkov*, **
*Sechenov First Moscow State Medical University, Ministry of Health of the Russian Federation,
Trubetskaya ul. 8/2, Moscow, 119991 Russia
**Dorodnicyn Computing Centre, Federal Research Center «Computer Science and Control»
of the Russian Academy of Sciences, ul. Vavilova 44/2, Moscow, 119333 Russia
This study proposes a mathematical model of biological community dynamics which is represented by a sys-
tem of equations that describe the dynamics of ecosystem variables - population density; body weight of ma-
ture individuals; the density of energy received by an individual with food - and takes into account the age
structure of the consumer's population. Using the results on competition obtained for point models by other
researchers, algorithms for determining the evolutionarily optimal values of adaptive traits of mature individ-
uals of the population were obtained. It has been shown that during food resource competition between pop-
ulations each harboring a different set of traits, selection favored only the population which will survive and
have larger reproductive function, that corresponds to the general criterion for optimality in adaptive evolu-
tion. The classification of traits by types is carried out, it is shown that each type corresponds to its own cri-
terion for optimality in adaptive evolution. Simple algorithms for a search for evolutionarily optimal values of
various adaptive traits and body weight are proposed, the optimal distribution of energy consumption for all
types of vital functions of the organism is determined. Optimality in adaptive evolution of these traits is illus-
trated by a numerical experiment.
Keywords: optimality in adaptive evolution, stability of equilibrium state, competition between populations, adap-
tive traits
БИОФИЗИКА том 65
№ 6
2020