БИОФИЗИКА, 2021, том 66, № 1, с. 13-22
МОЛЕКУЛЯРНАЯ БИОФИЗИКА
УДК 577.3
ПЛЮСЫ И МИНУСЫ КОЛЬЦЕВЫХ РНК
В КАЧЕСТВЕ ГУБОК ДЛЯ микроРНК
© 2021 г. М.А. Дук*, М.Г. Самсонова**
*Физико-технический институт им. А.Ф. Иоффе, 194021, Санкт-Петербург, Политехническая ул., 26
**Санкт-Петербургский политехнический университет Петра Великого,
195251, Санкт-Петербург, Политехническая ул., 29
E-mail: duk@mail.ioffe.ru
Поступила в редакцию 21.08.2020 г.
После доработки 21.08.2020 г.
Принята к публикации 04.09.2020 г.
Роли некодирующих РНК различных типов в клетке активно изучаются в последние годы. Одним
из типов некодирующих РНК являются кольцевые РНК, которые ранее рассматривались как
отходы процесса сплайсинга, однако на нынешний день у многих из них были обнаружены важные
регуляторные функции, в том числе и работа в качестве губки для миРНК. Нами проведено
сравнение работы такой губки для миРНК с работой классической ветви регуляции с помощью
транскрипционного фактора с точки зрения эффективности, времени работы и шума. Показано,
что кольцевая РНК с множеством сайтов связывания для миРНК оказывается значительно
эффективнее, чем транскрипционный фактор, и такой контур работает быстрее, однако это верно
только в условиях неполного насыщения сайтов связывания молекулами миРНК. Кроме того,
шумовые характеристики контура значительно ухудшаются с ростом количества сайтов
связывания. Показано также, что кольцевая РНК с одним сайтом связывания малоэффективна в
качестве губки для миРНК. Предполагается, что такой тип регуляции в клетке является скорее
некоторым специфическим инструментом.
Ключевые слова: кольцевые РНК, миРНК, губки для миРНК, некодирующие РНК, шум.
DOI: 10.31857/S0006302921010026
которые бы могли быстро связывать миРНК и
В последние годы некодирующие РНК при-
убирать их из системы.
влекают все большее внимание исследователей.
Помимо активно изучающихся микроРНК, или
Обнаружение все новых типов кольцевых РНК
миРНК, большой интерес стали представлять
привело к пониманию, что кольцевые РНК - это
кольцевые РНК, которые встречаются повсе-
множество типов РНК с различным происхожде-
местно [1-3] и имеют различные функции и про-
нием и функциями. Так, кольцевые РНК могут
исхождение.
образовываться в процессе сплайсинга из выре-
Изначально обнаруживаемые в клетке кольце-
занных интронов, но гораздо чаще - в результате
вые РНК принимали за продукт сплайсинга ин-
неканонического обратного сплайсинга или
тронов, ошибочно соединившийся в кольцо и не
бэксплайсинга, происходящего во время генера-
играющий никакой значимой роли [4]. Однако
ции пре-мРНК или уже на готовой пре-мРНК [1,
вскоре стало понятно, что кольцевые РНК игра-
3, 7]. Процесс бэксплайсинга идет иначе, чем ка-
ют существенную роль в регуляции в клетке. У
нонический сплайсинг: в процессе бэксплайсин-
кольцевых РНК некоторых типов были обнару-
га нижестоящий донор сплайсинга присоединя-
жены сайты связывания для миРНК [3, 5, 6], в не-
ется к предшествующему акцептору сплайсинга,
которых случаях количество сайтов связывания
таким образом получается обратный порядок эк-
было более 70, как для ciRS-7, связывающей miR-
зонов, см. рис. 1. Предполагается, что бэксплай-
7 [5], а миРНК, которые связывались с этими сай-
синг мешает процессу линейного сплайсинга, и
тами кольцевых РНК, играют важнейшие регуля-
таким образом производство кольцевых РНК мо-
торные роли. Благодаря этому было выдвинуто
жет подавлять производство линейной мРНК [1,
предположение, что кольцевые РНК работают в
8, 9].
качестве «губок» для миРНК (miRNA sponging),
Тем не менее бэксплайсинг происходит на-
Сокращениe: ТФ - транскрипционный фактор.
много реже, чем линейный сплайсинг [4, 10], и
13
14
ДУК, САМСОНОВА
вая РНК работает значительно быстрее, чем ТФ,
внутренний шум у такого контура оказывается
значительно больше.
МЕТОДЫ
Для оценки эффективности работы контура с
кольцевой РНК для удаления миРНК из клетки
сравнивается такая ветвь с классической ветвью
регуляции, в которой ТФ репрессирует миРНК.
Рис. 1. Схематическое представление различия ли-
Допустим, в системе производится пре-мРНК γ1,
нейного сплайсинга и обратного сплайсинга (бэкс-
из нее после сплайсинга получается мРНК r, на
плайсинга). В процессе линейного сплайсинга из
пре-мРНК вырезаются интроны и итоговым продук-
основе этой мРНК строится белок - транскрип-
том является зрелая мРНК. В процессе обратного
ционный фактор q, который подавляет выработку
сплайсинга экзоны поменяли порядок и пре-мРНК
миРНК m1 в системе. Считаем, что мРНК ис-
превратилась в кольцевую РНК.
пользуется несколько раз, т.е. не разрушается по-
сле производства белка на ее основе. Таким обра-
количество кольцевых РНК одного типа в клетке
зом, в системе происходят реакции, показанные в
табл. 1. Здесь скорость производства миРНК под
значительно меньше, чем количество мРНК, по-
действием транскрипционного фактора задается
лученных с этого же гена.
Функции кольцевых РНК тоже различны, так,
функцией Хилла: km(q) = (kmH2)/(H2 + q2).
кроме функционирования в качестве губок для
Уравнения для такой ветви регуляции будут
миРНК, называется участие в развитии некото-
иметь вид
рых болезней [1, 11], участие в противовирусном
иммунитете [8], а также возможность кодировать
dγ1/dt = kγ - gγγ1 - krγ1,
небольшие белки [8].Тем не менее большое коли-
dr/dt = krγ1 - grr,
чество обнаруженных кольцевых РНК, содержа-
щих только один или два сайта связывания
dq/dt = kqr - gqq,
(1)
миРНК [1, 7], наводят на мысль, что функциони-
рование кольцевых РНК в качестве губок для
dm1/dt = kmH2/(H2 + q2) - gmm.
миРНК переоценено в литературе.
В данной работе представляется сравнение
Рассмотрим теперь ветвь регуляции, представ-
между контуром, в котором производство миРНК
ляющую собой связывание миРНК с помощью
подавляется на транскрипционном уровне за счет
кольцевой РНК (miRNA sponging). В данном слу-
репрессии белком-транскрипционным факто-
чае вырабатывается пре-мРНК γ2, на основе нее в
ром, и контуром, в котором кольцевая РНК рабо-
результате обратного сплайсинга образуется
тает в качестве губки для миРНК. Показывается,
кольцевая РНК c0, которая связывает миРНК m2
что только кольцевая РНК с множественными
в системе. Индексы 1 и 2 здесь даны пре-мРНК и
сайтами связывания способна работать сопоста-
миРНК исключительно для отличия рассматри-
вимо или эффективнее, чем транскрипционный
ваемых контуров друг от друга, характеристики
фактор (ТФ), и только в условиях избыточности
же, такие как скорость производства и распада,
сайтов связывания.
полагаем в двух контурах одинаковыми для воз-
Кроме того, были проанализированы шумо-
можности сравнения контуров между собой. Ре-
вые характеристики, и показано, что хоть кольце-
акции показаны в табл. 2. Здесь n - число сайтов
Таблица 1. Реакции в системе при регуляции с помощью транскрипционного фактора
Производство
Распад
kγ
g
γ
Пре-мРНК
∅→
γ
1
γ1→∅
kr
g
r
мРНК
γ
→
r
1
r→∅
kq
gq
ТФ
r→r +q
q→∅
m
k q)
m
g
миРНК
∅ →
m
1
m
1
→∅
БИОФИЗИКА том 66
№ 1
2021
ПЛЮСЫ И МИНУСЫ КОЛЬЦЕВЫХ РНК
15
Таблица 2. Реакции в системе при регуляции с помощью кольцевой РНК
Производство/связывание
Распад
Расщепление комплексов
kγ
gγ
Пре-мРНК
∅→
γ
γ
→∅
1
2
k
m
g
m
мРНК
∅→
m
2
m
2
→∅
k
c
g
c
Кольцевая РНК
γ
2
→
c
0
c
0
→ ∅
k
cm
g
c
g
cm
Комплекс-1
c
+m
→c
c
→∅
c
→c
+m
0
2
1
1
1
0
2
k
cm
g
c
g
cm
Комплекс-2
c
1
+m
2
→c
2
c
2
→∅
c
2
→c
1
+m
2
…
Комплекс-n
k
cm
g
c
g
cm
c
n
−1
+
m
2
→
c
n
c
n
→∅
c
n
→c
n
−1
+
m
2
связывания, а индекс у c отражает количество уже
Предполагаем, что скорость собственного рас-
занятых сайтов связывания.
пада пре-мРНК в п.о. такая же, как у мРНК, ис-
ходя из разницы длины мРНК и пре-мРНК, по-
Уравнения для такой ветви регуляции будут
лучаем скорость распада в молекулах в минуту:
иметь следующий вид:
g
dγ2/dt = kγ - gγγ2 - kсγ2,
γ = 0.0005 мин-1.
Средняя длина белка около
500 a.a.
dс0/dt = kсγ2 - gсc0 - kсmc0m2 + gсmc1,
(BNID:108984), а скорость трансляции - 5 a.a./с
dс1/dt = kсmc0m2 - gсc1 - kсmc1m2 - gсmc1 + gсmc2,
(BNID:104598, [16]), таким образом, вычисляем,
что скорость производства белка kp = 0.6 мин-1.
dс2/dt = kсmc1m2 - gсc2 - kсmc2m2 - gсmc2 + gсmc3,
Скорости распада белков в клетках человека
…
(2)
очень различны, поэтому используем не среднее,
dсn/dt = kсmcn-1m2 - gmcn - gсmcn,
а медианное значение, приведенное в BNID:
112253 ([17]): gp
= 0.034 ч-1 ≈ 0.0006 мин-1.
n-1
n
dm
/
dt = k
−
g
m
−
k
m
c
+
g
c
И микроРНК, и кольцевые РНК считаются
2
m
m
2
cm
2
i
cm
j
i
=0
j
=1
очень стабильными [2, 10], поэтому полагаем, что
их скорость распада сопоставима со скоростью
Вычисление коэффициентов модели произво-
распада белков. Характерную скорость производ-
дилось с учетом данных из литературы.
ства миРНК тоже сложно оценить, так как
Средняя скорость работы полимеразы
миРНК может образовываться как из вырезан-
3.79 kb/мин [12], средняя длина одного интрона в
ных интронов пре-мРНК, так и транскрибиро-
пре-мРНК 3.4 kb и их среднее количество 8 [13],
ваться отдельным продуктом (около 80 нуклеоти-
таким образом, интроны занимают порядка
дов пре-миРНК переносится в цитоплазму и об-
27.2 kb, на кодирующую часть гена у эукариот
резается до длины примерно 22 нуклеотида [18] в
приходится порядка 1.3 kb [14], таким образом по-
процессе сплайсинга). Для нашей задачи ско-
лагаем, что средняя длина пре-мРНК 28.5 kb, от-
рость производства миРНК выбираем так, чтобы
куда получаем скорость производства пре-мРНК
количество молекул миРНК одного типа было
kγ = 0.13 мин-1.
порядка 103, что соответствует данным по разным
типам миРНК (BNID:101297-101299).
Среднее время распада мРНК у бактерий 5-
Скорость производства мРНК из пре-мРНК
10 мин [15], среднее время полураспада мРНК у
kr, скорость производства кольцевой РНК kc, а
млекопитающих
-
50 мин (BNID:112681),
также константы связывания и развязывания
комплексов молекул kсm и gсm также подбираем из
вычисляя получаем скорость распада в среднем
расчета, чтобы итоговое количество молекул бел-
gr = 0.01 мин-1.
ка ТФ, мРНК и миРНК в стационарном состоя-
БИОФИЗИКА том 66
№ 1
2021
16
ДУК, САМСОНОВА
Таблица 3. Коэффициенты, использованные в расчетах
Описание
Значение
Источник
kγ
Скорость производства пре-мРНК
0.13 мин-1
[12], [14]
kc
Скорость производства кольцевой РНК
0.00483 мин-1
*, [10]
kr
Скорость производства мРНК
0.8 мин-1
BNID:108248, 101729
km
Скорость производства миРНК
1 мин-1
BNID:101297-01299
kq
Скорость производства белка-ТФ
0.6 мин-1
BNID:108984 [16]
gγ
Скорость распада пре-мРНК
0.0005 мин-1
[13-15]
gc
Скорость распада кольцевой РНК
0.0006 мин-1
*
gr
Скорость распада мРНК
0.01 мин-1
[15], BNID:112681
gm
Скорость распада миРНК
0.0006 мин-1
*
gq
Скорость распада белка-ТФ
0.0006 мин-1
[17]
Связывание комплексов
kcm
1 мин-1
*
кольцевая+миРНК
Развязывание комплексов
gcm
1 мин-1
*
кольцевая+миРНК
H
Коэффициент в функции Хилла
6000
*
Примечание. * - Подобранные коэффициенты.
нии по порядку соответствовало бы данным из
процессом. Компоненты полученного вектора z
литературы. Всего мРНК в клетке порядка 15000
добавлялись к количеству молекул миРНК (m1
и их около 20000 типов (BNID:108248), полагаем,
или m2) в каждый i-й момент времени, после чего
мРНК одного типа в среднем будет порядка 100,
численно решалась система уравнений (1) или (2)
количество молекул белка одного типа порядка
для следующего момента времени. Таким обра-
103-104 (BNID:101729). Количество кольцевых
зом, был получен вектор количества миРНК в си-
РНК одного типа в клетке неизвестно, но бэкс-
стеме с внешним шумом mnoise. Количество моле-
плайсинг, приводящий к появлению кольцевой
кул миРНК в начальный момент времени для
РНК, случается значительно реже, чем прямой
mnoise (t = 0) = mstat.
линейный сплайсинг [10], поэтому скорость про-
изводства кольцевой РНК должна быть значи-
тельно меньше чем скорость производства
РЕЗУЛЬТАТЫ
мРНК, что также учитывалось при подборе коэф-
фициентов.
Скорость производства кольцевой РНК неиз-
Коэффициент диссоциации H в функции Хил-
вестна, известно лишь, что обратный сплайсинг
ла для контура с ТФ был выбран равным полови-
случается значительно реже, чем прямой сплай-
не характерного значения количества молекул
синг. Скорость производства была выбрана таким
белка ТФ.
образом, чтобы кольцевая РНК с несколькими
сайтами (в нашем случае для наглядности было
Использованные коэффициенты представ-
выбрано семь сайтов) приводила бы к тому же
стационарному значению количества мик-
лены в табл. 3.
роРНК, что и рассматриваемая схема с ТФ.
При исследовании внешнего шума рассматри-
На рис. 2 показано сравнение количества
валась величина t|mnoise - mstat|, где mstat - коли-
миРНК при различных способах регуляции -
чество миРНК в квазистационарном состоянии.
сравнение свободной выработки миРНК, регуля-
Вектор внешнего шума строился таким образом:
ции с помощью ТФ и регуляции с помощью коль-
z (i) = exp (-0.1) z (i -1)+
1
-
exp
(-0.2
) ξ,
где ξ -
цевой РНК с одним сайтом связывания; сравне-
стандартный белый шум, построенный процеду-
ние свободной выработки миРНК, регуляции с
рой MATLAB. Вектор z является дискретной мо-
помощью ТФ и регуляции с помощью кольцевой
делью процесса Орнштейна-Уленбека, являю-
РНК с семью сайтами связывания, а также коли-
щегося стационарным марковским гауссовым
чество связанных комплексов в системе регуля-
БИОФИЗИКА том 66
№ 1
2021
ПЛЮСЫ И МИНУСЫ КОЛЬЦЕВЫХ РНК
17
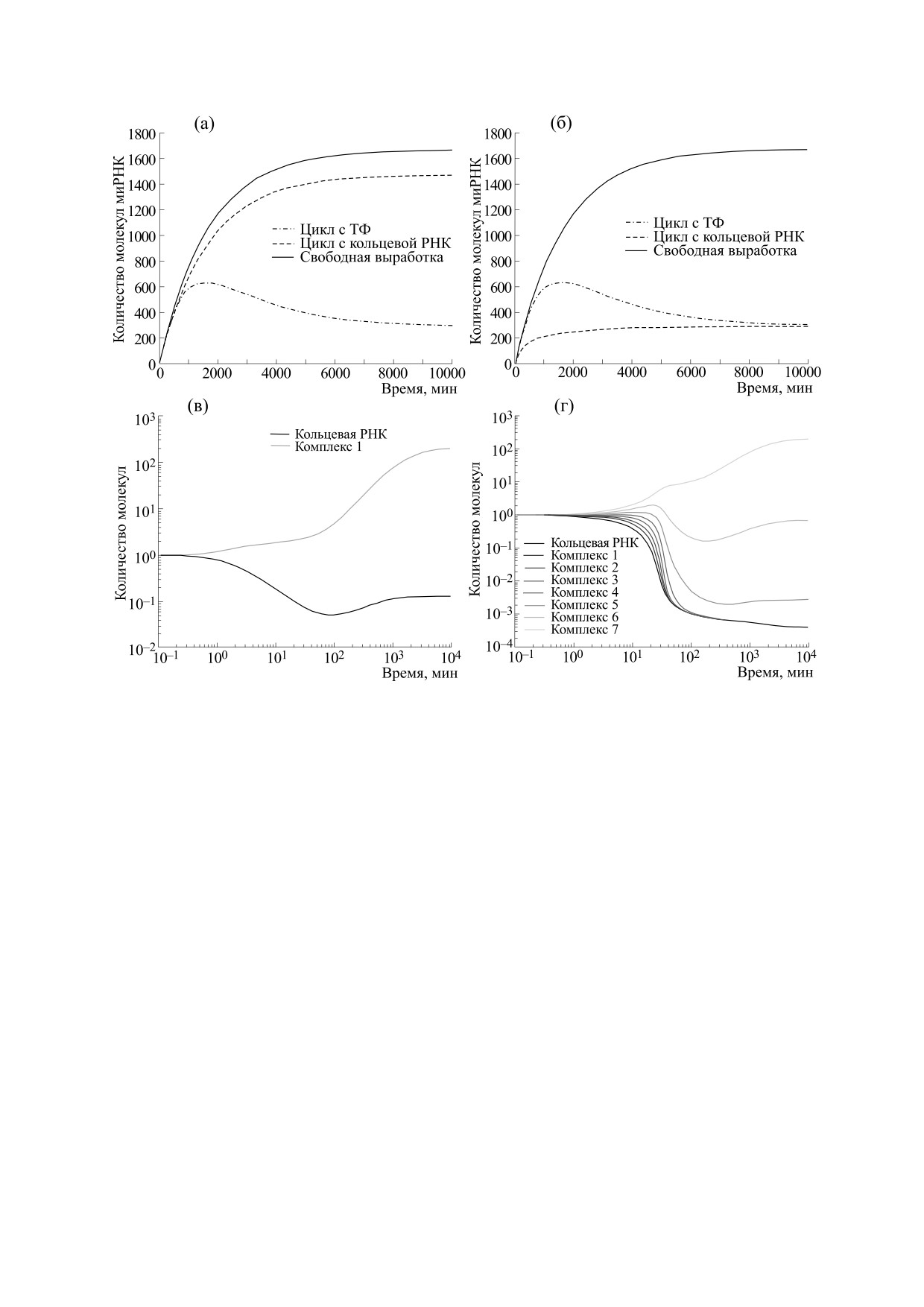
Рис. 2. (а) и (б) - Сравнение динамики количества микроРНК при свободной выработке, при регуляции с помощью
ТФ и с помощью кольцевых РНК с одним и семью сайтами связывания соответственно; (в) и (г) - динамика
количества кольцевой РНК и ее связанных комплексов во времени при наличии одного и семи сайтов связывания
соответственно.
ции с помощью кольцевой РНК с одним или се-
этой задержки на рис. 2б можно видеть максимум
мью сайтами связывания.
в количестве миРНК в контуре с транскрипцион-
ным фактором, связанный с тем, что производ-
Можно видеть, что одинаковая эффектив-
ство необходимого для регуляции количества
ность подавления миРНК в случае семи сайтов
белка ТФ требует больше времени, чем производ-
связывания достигается при примерно 200 моле-
ство РНК. Иными словами, в случае существова-
кулах кольцевой РНК против примерно 12000 мо-
ния в системе подходящей на роль губки для
лекул транскрипционного фактора. Таким обра-
миРНК кольцевой РНК, она удаляет миРНК из
зом, даже при очень низкой концентрации, кото-
системы быстро и эффективно в сравнении с
рая была обнаружена у большинства типов
классической ветвью регуляции транскрипцион-
кольцевых РНК [1, 2], кольцевые РНК могут ра-
ным фактором. Однако, несмотря на скорость ра-
ботать достаточно эффективно при условии на-
боты и эффективность, подобные губки для
личия множественных сайтов связывания, тогда
миРНК обладают некоторыми существенными
как кольцевая РНК с одним сайтом связывания
минусами, о которых будет написано ниже, и ко-
не обеспечивает достаточного уровня подавления
торые, возможно, и являются причинами не
своей мишени.
столь широкой их распространенности.
Также немаловажно, что контур с кольцевой
Тем не менее при сравнении ветвей регуляции
РНК значительно быстрее приходит к стационар-
важны также и шумовые характеристики этих
ным значениям, тогда как в контуре с транскрип-
контуров. На рис. 3 показаны результаты стоха-
ционным фактором наблюдается задержка. Из-за
стического моделирования контуров в квазиста-
БИОФИЗИКА том 66
№ 1
2021
18
ДУК, САМСОНОВА
ционарном состоянии с использованием метода
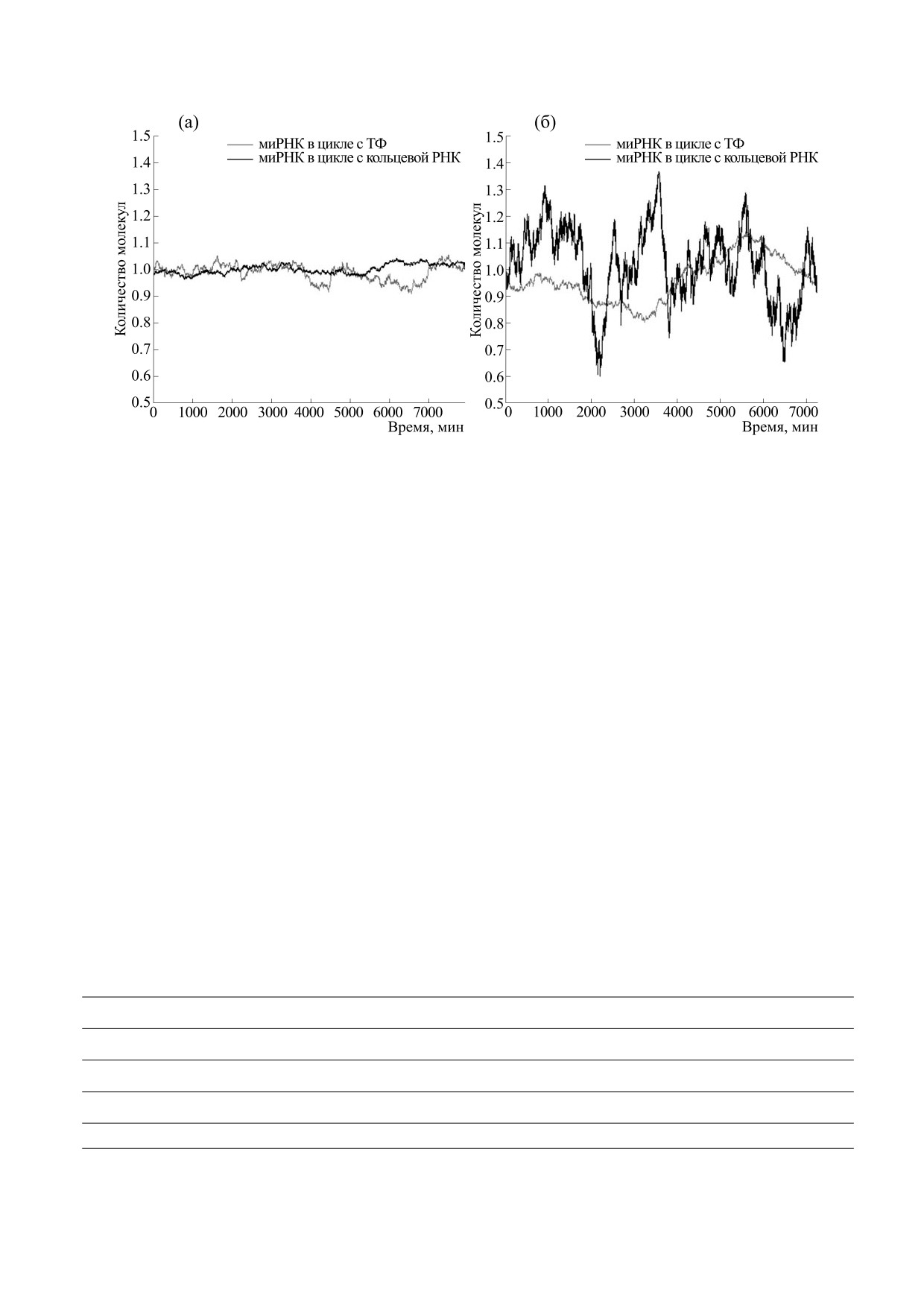
Гиллеспи, описанного в работе [19]. Так как при
увеличении числа сайтов вероятность связыва-
ния кольцевой РНК хотя бы с одной молекулой
миРНК значительно возрастает, на рис. 3в для
кольцевой РНК с семью сайтами связывания
практически не встречается свободная кольцевая
РНК, тогда как на рис. 3б с кольцевой РНК с од-
ним сайтом связывания свободная кольцевая
РНК встречается часто. Наличие такого количе-
ства свободной кольцевой РНК в контуре с одним
сайтом связывания косвенно говорит о низкой
эффективности такого регулятора в качестве губ-
ки для миРНК.
С другой стороны, при наличии множества
сайтов связывания, к которым миРНК может как
прикрепляться, так и отсоединяться, увеличива-
ется вариабельность в количестве свободных и
присоединенных к кольцевой молекуле молекул
миРНК, что означает увеличение внутреннего
шума, что можно наблюдать на рис. 3в в сравне-
нии с рис. 3б. Не очень большой уровень внутрен-
него шума, наблюдаемый на рис. 3а для контура с
ТФ, можно связать с меньшей чувствительностью
количества белка-ТФ к колебаниям в пре-мРНК
в сравнении с чувствительностью у количества
кольцевой РНК.
На рис. 4 показано относительное количество
миРНК в рассматриваемых контурах. Можно ви-
деть, что шум в контуре с кольцевой РНК с одним
сайтом связывания меньше, чем в контуре с ТФ,
однако в контуре с кольцевой РНК с семью сайта-
ми связывания шум гораздо больше, чем в конту-
ре с ТФ. Таким образом, внутренний шум, свя-
занный с прикреплением и отсоединением одной
молекулы миРНК (что происходит в контуре, ре-
гулируемом кольцевой РНК с одним сайтом свя-
зывания) не достигает уровней, которые получа-
ются из-за сложения шума от производства
мРНК, а потом белка при сложном создании ре-
гулятора в контуре с транскрипционным факто-
ром, что, в частности, можно было наблюдать,
например, в работах [20, 21], где сравнивалась ре-
гуляция белка-мишени с помощью ТФ и с помо-
щью миРНК и делался вывод о лучшей работе ре-
гулятора-РНК в сравнении с регулятором-бел-
ком. Однако увеличение числа сайтов
связывания значительно увеличило внутренний
шум в контуре с кольцевой РНК. Учитывая тот
факт, что подтвержденные в качестве губок для
миРНК кольцевые РНК могут иметь до 70 сайтов
связывания [5], внутренний шум при такой ветви
регуляции будет очень заметным.
Рис. 3. Стохастическое моделирование контуров:
(а) - регуляция с помощью транскрипционного фак-
Также была исследована способность конту-
тора; (б) - регуляция с помощью кольцевой РНК,
ров подавлять внешний шум, который вносили в
один сайт связывания; (в) - регуляция с помощью
кольцевой РНК, семь сайтов связывания. На рисун-
количество миРНК после прихода системы в ста-
ках (б) и (в) показана только концентрация свобод-
ционарное состояние, после чего рассматривали
ной кольцевой РНК, концентрации связанных ком-
величину |mnoise - mstat|, где mnoise - количество
плексов не показаны.
БИОФИЗИКА том 66
№ 1
2021
ПЛЮСЫ И МИНУСЫ КОЛЬЦЕВЫХ РНК
19
Рис. 4. Сравнение относительного количества миРНК m⁄(<m>) в системе с ТФ и с кольцевой РНК: (а) - с одним
сайтом связывания, (б) - с семью сайтами связывания.
стационарного количества миРНК в системе с
миРНК при наличии внешнего шума, а mstat -
транскрипционным фактором, а также в системе
стационарное количество миРНК. Вектор внеш-
с кольцевой РНК с одним или с семью сайтами
него шума строили так же, как и ранее в работе
связывания.
[21]. В данном случае контур с кольцевой РНК с
семью сайтами связывания немного лучше по-
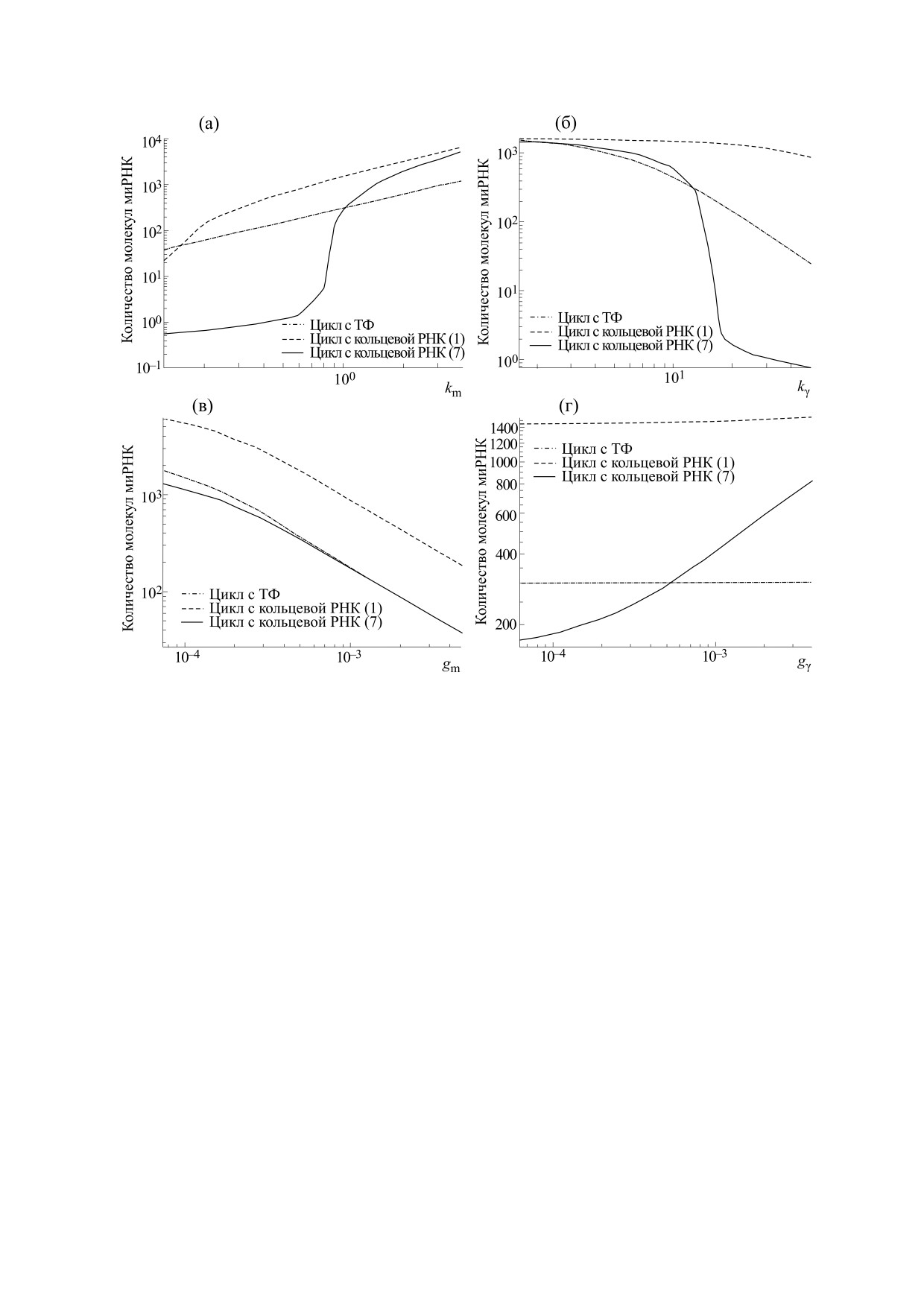
Можно видеть, что при различных скоростях
давлял внешний шум. Примеры результатов для
производства и распада микроРНК (km и gm) и
одного и того же вектора внешнего шума показа-
скоростях производства (kγ и gγ) пре-мРНК регу-
ны в табл. 4. Лучшее подавление внешнего шума
лятора (ТФ или кольцевой РНК) контур с ТФ по-
при отсутствии внутреннего шума обеспечивает-
давляет миРНК эффективнее, чем контур с коль-
ся за счет того, что контур с кольцевой РНК с се-
цевой РНК с одним сайтом связывания. Однако
мью сайтами работает быстрее, чем контур с
для кольцевой РНК с семью сайтами связывания
транскрипционным фактором, система быстрее
картина другая. РНК значительно эффективнее
приходит к стационарному состоянию после каж-
подавляет миРНК в условиях, когда не происхо-
дого единичного внесения шума, соответственно
дит полного насыщения молекулы кольцевой
суммарный итоговый шум в количестве миРНК
РНК молекулами миРНК, т.е. при малых скоро-
оказывается меньше. Тем не менее эффектив-
стях производства миРНК или больших скоро-
ность подавления внешнего шума теряется, если
стях производства пре-мРНК кольцевой РНК
рассматривать систему, содержащую и внешний
(см. рис. 5а,б). Когда концентрация миРНК в си-
и внутренний шум, о котором написано выше.
стеме оказывается достаточно большой, контур с
Интересно также посмотреть на эффектив-
кольцевой РНК с семью сайтами связывания на-
ность кольцевой РНК при различных значениях
чинает работать так же неэффективно, как контур
коэффициентов. На рис. 5 показаны значения
с кольцевой РНК с одним сайтом связывания.
Таблица 4. Величина Σ|mnoise - mstat| в контуре с транскрипционным фактором и в контуре с кольцевой РНК с
семью сайтами связывания при одних и тех же значениях внесенного в систему внешнего шума
Контур с транскрипционным фактором
Контур с кольцевой РНК
89.3375
86.9057
91.7398
89.2120
86.4638
83.6620
87.5369
84.2301
БИОФИЗИКА том 66
№ 1
2021
20
ДУК, САМСОНОВА
Рис. 5. Сравнение стационарного количества миРНК в системе с ТФ и с кольцевой РНК с одним и семью сайтами
связывания: (а) - при разных значениях скорости производства миРНК km, (б) - при разных значениях скорости
производства пре-мРНК регулятора kγ, (в) - при разных скоростях распада миРНК gm, (г) - при разных скоростях
распада пре-мРНК регулятора gγ.
Важно отметить, что переход от эффективного
ность такого контура, приближая ее к эффектив-
режима работы к неэффективному режиму ока-
ности контура с одним сайтом связывания (см.
зывается достаточно резким при изменении ско-
рис. 5г).
ростей производства субстрата или мишени.
Интересно отметить, что при уменьшении
ОБСУЖДЕНИЕ
скорости распада миРНК gm не наблюдается за-
В последние годы было найдено много кольце-
метных различий в количестве миРНК в контурах
вых РНК, содержащих малое количество сайтов
(см. рис. 5в). Это связано с тем, что стабильность
связывания для миРНК. Несмотря на то что не-
миРНК очень высока, т.е. характерная скорость
которые коэффициенты модели были подобраны
распада миРНК, используемая нами, очень мала,
исходя из общих представлений биохимии про-
и дальнейшее ее уменьшение слабо влияет на на-
цессов, в нашей статье можно видеть, что кольце-
блюдаемую картину. Однако скорость распада
вая РНК с одним сайтом связывания не может до-
пре-мРНК кольцевой РНК gγ заметно влияет на
стичь уровня эффективности классической регу-
контур с кольцевой РНК с семью сайтами связы-
ляции с помощью транскрипционного фактора.
вания, так как нехватка пре-мРНК для создания
Учитывая вероятность отсоединения миРНК от
кольцевой РНК увеличивает вероятность, что все
кольцевой РНК, при наличии только одного сай-
сайты связывания уже выработанных кольцевых
та связывания кольцевая РНК очень часто оказы-
РНК будут заняты, что уменьшает эффектив-
вается свободной, что можно увидеть при стоха-
БИОФИЗИКА том 66
№ 1
2021
ПЛЮСЫ И МИНУСЫ КОЛЬЦЕВЫХ РНК
21
стическом моделировании (см. рис. 3) и что кос-
сайтов связывания кольцевой РНК молекулами
венно указывает на то, что такая кольцевая РНК
миРНК. В случае же насыщения, которое проис-
неэффективна в качестве губки для миРНК.
ходит при увеличении количества миРНК или
уменьшении количества кольцевой РНК (напри-
Исходя из этого факта, можно предположить,
мер, в случае изменения скоростей производства
что скорее миРНК регулирует такую кольцевую
km и kγ), эффективность кольцевой РНК с множе-
РНК, а не наоборот, сама же кольцевая РНК с ма-
лым количеством сайтов связывания миРНК од-
ственными сайтами связывания оказывается
ного типа выполняет в клетке иные функции.
сравнимой с эффективностью кольцевой РНК с
одним сайтом связывания, причем переключение
Тем не менее кольцевая РНК с множествен-
с эффективного режима на неэффективный ре-
ными сайтами связывания значительно эффек-
жим происходит достаточно резко, см. рис. 5а,б.
тивнее понижает количество активных миРНК в
Также эффективность контура с кольцевой РНК
системе, чем транскрипционный фактор. В на-
оказывается чувствительной не только к скорости
шем случае для наглядного сравнения была вы-
производства пре-мРНК kγ, но и к ее скорости
брана кольцевая РНК только с семью сайтами
распада gγ, при этом эффективность контура с
связывания. Для уменьшения количества миРНК
до одинакового уровня понадобилось 200 моле-
транскрипционным фактором оказывается не на-
кул такой кольцевой РНК против 12000 молекул
столько чувствительной к скорости распада пре-
транскрипционного фактора, что говорит о высо-
мРНК, что, вероятно, связано с достаточной ста-
кой эффективности такой губки для миРНК.
бильностью молекул белка.
Учитывая тот факт, что у кольцевых РНК, под-
Подводя итог, можно сказать, что только коль-
твержденных в качестве губок для миРНК, обна-
цевые РНК с достаточно большим количеством
руживается 30-70 сайтов связывания, даже очень
сайтов связывания могут рассматриваться в каче-
низкие уровни выработки кольцевых РНК могут
стве губок для миРНК, однако работа таких губок
быть достаточными для эффективной регуляции.
для миРНК имеет свои особенности. С одной
Еще одним несомненным плюсом в регуляции
стороны, такие губки могут быть очень эффек-
миРНК с помощью кольцевой РНК со множе-
тивны, но только в некоторых пределах значений
ственными сайтами связывания, кроме ее эффек-
коэффициентов производства и распада компо-
тивности, является скорость работы такого кон-
нентов. В условиях, когда за счет изменения этих
тура. Если для понижения количества миРНК с
скоростей сайты связывания кольцевой РНК
помощью транскрипционного фактора требуется
оказываются занятыми, эффективность работы
время на сборку белка и на достижение необходи-
такого контура резко падает. Немаловажно, что
мого количества регулятора, кольцевая РНК со
губки для миРНК работают быстро, но скорость
множественными сайтами связывания начинает
их работы компенсируется наличием значитель-
работать сразу, не позволяя количеству миРНК
ного внутреннего шума, таким образом - губки
значительно возрасти.
для миРНК есть скорее некоторый специфиче-
ский инструмент в живых клетках, нежели наибо-
С другой стороны, шумовые характеристики у
лее эффективный способ уменьшить количество
контура с кольцевой РНК очень плохие по срав-
активных миРНК в системе.
нению с контуром с транскрипционным факто-
ром, и внутренний шум нарастает с увеличением
количества сайтов связывания, поэтому контуры,
БЛАГОДАРНОСТИ
регулируемые транскрипционным фактором, бо-
лее эффективны в деле тонкой настройки, тогда
Авторы выражают благодарность С.А.Руко-
как контуры с кольцевой РНК с множественны-
лайне за консультации.
ми сайтами связывания работают по принципу:
«быстро, но не точно». Несмотря на то что контур
ФИНАНСИРОВАНИЕ РАБОТЫ
с кольцевой РНК немного лучше подавляет
внешний шум, чем контур с транскрипционным
Исследование роли кольцевых РНК выполне-
фактором, этот эффект оказывается незаметным
но М.А. Дук в ФТИ им.А.Ф. Иоффе в рамках на-
при рассмотрении одновременно внешнего и
учной темы 0040-2019-0003 «Нелинейные про-
внутреннего шума за счет большой величины по-
цессы и механизмы переноса вещества в конден-
следнего. Большой внутренний шум, связанный с
сированных средах и биоструктурах», вери-
таким способом регуляции, возможно, является
фикация биологической значимости исследова-
одной из причин слабой его распространенности
ния выполнена М.Г. Самсоновой при финансо-
в сравнении с другими способами регуляции в
вой поддержке Минобрнауки России в рамках ре-
клетке.
ализации программы Научного центра мирового
Кроме того, кольцевая РНК с множественны-
уровня по направлению «Передовые цифровые
ми сайтами связывания очень эффективно рабо-
технологии» СПбПУ (соглашение
№ 075-15-
тает только в условиях неполного насыщения
2020-934).
БИОФИЗИКА том 66
№ 1
2021
22
ДУК, САМСОНОВА
КОНФЛИКТ ИНТЕРЕСОВ
8. M. Wang, F. Yu, W. Wu, et al., Int. J. Biol. Sci. 13, 1497
(2017). DOI: 10.7150/ijbs.22531
Авторы заявляют об отсутствии конфликта
9. R. Ashwal-Fluss, M. Meyer, N. R. Pamudurti, et al.,
интересов.
Mol. Cell 56 (1), 55 (2014).
10. L. L. Chen and L. Yang, RNA Biol. 12 (4), 381 (2015).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
11. Y. Dong, D. He, Z. Peng, et al., J. Hematol. Oncol. 10,
Art. 2 (2017). DOI: 10.1186/s13045-016-0370-2
Настоящая работа не содержит описания ис-
12. J. Singh and R. A. Padgett, Nat. Struct. Mol. Biol. 16
следований с использованием людей и животных
(11), 1128 (2009).
в качестве объектов.
13. J. Hnilicová and D. Stanek, Nucleus 2 (3), 182 (2011).
14. L. Xu, H. Chen, X. Hu, et al., Mol. Biol. Evol. 23 (6),
СПИСОК ЛИТЕРАТУРЫ
1107 (2006).
1.
J. Greene, A. M. Baird, L. Brady, et al., Front. Mol.
15. Y. Taniguchi, P. J. Choi, G. W. Li, et al., Science 329
Biosci. 4, 38 (2017).
(5991), 533 (2010).
16. S. O. Olofsson, K. Boström, P. Carlsson, et al., Am.
2.
S. Guil and M. Esteller, Trends Biochem. Sci. 40 (5),
Heart J. 113 (2, Pt 2), 446 (1987).
248 (2015).
17. M. K. Doherty, D. E. Hammond, M. J. Clague, et al.,
3.
L. M. Holdt, A. Kohlmaier, and D. Teupser, Cell. Mol.
J. Proteome Res. 8 (1), 104 (2009).
Life Sci. 75, 1071 (2018).
18. T. C. Chang and J. T. Mendell, Annu. Rev. Genomics
4.
J. Salzman, C. Gawad, P. L. Wang, et al., PLoS One 7
Hum. Genet. 8, 215 (2007).
(2), e30733 (2012).
19. D. T. Gillespie, Phys. Rev. E 54 (2), 2084 (1996).
5.
T. B. Hansen, J. Kjems, and C. K. Damgaard, Cancer
20. M. Osella, C. Bosia, D. Cora, and M. Caselle, PLoS
Res. 73, 5609 (2013).
Comput. Biol. 7 (3), e1001101 (2010).
6.
I. Jost, L. A. Shalamova, G. K. Gerresheim, et al.,
21. S. Tej, K. Gaurav, and S. Mukherji, Phys. Biol. 16 (4),
RNA Biol.
15
(8),
1032
(2018).
DOI:
046008 (2019). DOI: 10.1088/1478-3975/ab1563
10.1080/15476286.2018.1435248
22. M. A. Duk, M. G. Samsonova, and A. M. Samsonov,
7.
S. Memczak, M. Jens, A. Elefsinioti, et al., Nature 495,
BMC Genomics 15 (Suppl. 12), S9 (2014). DOI:
333, (2013). DOI: 10.1038/nature11928
10.1186/1471-2164-15-S12-S9
Pros and Cons of Circular RNAs as miRNA Sponges
M.A. Duk* and M.G. Samsonova**
*Ioffe Institute, ul. Polytekhnicheskaya 26, St. Petersburg, 194021 Russia
**Peter the Great St. Petersburg Polytechnic University, ul. Polytekhnicheskaya 29, St. Petersburg, 195251 Russia
In recent years, researchers have been challenged to study roles for non-coding RNA of various types in the
cell. Circular RNAs (or circRNAs) are a type of non-coding RNAs previously considered junk by-products
in splicing process. but to date, many of them are found to demonstrate important regulatory functions and
also act as miRNA sponge. This study involved the comparison between the properties of miRNA sponge and
the transcription factor associated with regulation of gene expression, assessing the efficiency, the time taken
by the loop and noise characteristics. It was shown that circRNA with multiple binding sites for miRNA is
much more effective than the transcription factor, and this loop works faster, however, this is only true when
the binding sites are incompletely saturated with miRNA molecules. Moreover, the noise characteristics of
the loop significantly are getting worse with an increase in the number of binding sites. It was also shown that
circRNA with one binding site acts inefficiently as a sponge of miRNA. The results suggest that this type of
regulation in the cell is rather a specific tool.
Keywords: circular RNAs, miRNA, miRNA sponge, non-coding RNA, noise
БИОФИЗИКА том 66
№ 1
2021