БИОФИЗИКА, 2021, том 66, № 1, с. 49-70
МОЛЕКУЛЯРНАЯ БИОФИЗИКА
УДК 577.218
НЕЧЕТКОЕ МОДЕЛИРОВАНИЕ РЕГУЛЯЦИИ ГЕНОВ СЕГМЕНТАЦИИ
ДРОЗОФИЛЫ НА ЯЗЫКЕ ЛИНГВИСТИЧЕСКИХ ПРАВИЛ
© 2021 г. А.А. Макашов*, Е.М. Мясникова*, А.В. Спиров*, **
*Санкт-Петербургский политехнический университет Петра Великого,
195251, Санкт-Петербург, Политехническая ул., 29
**Институт эволюционной физиологии и биохимии им. И.М. Сеченова РАН,
194223, Санкт-Петербург, просп. Тореза, 44
***Институт научной информации по общественным наукам РАН,
117997, Москва, ул. Кржижановского, 15/2
E-mail: alexander.spirov@gmail.com
Поступила в редакцию 26.12.2019 г.
После доработки 12.08.2020 г.
Принята к публикации 21.09.2020 г.
Понимание молекулярных механизмов регуляции генов - одна из фундаментальных проблем, на
которые нацелена системная биология. В принятых подходах к моделированию генной регуляции
зачастую невозможно отделить свойства модели, адекватно отображающие характеристики биоло-
гического объекта, от обусловленных природой модели. В данной работе развит логический подход
к моделированию регуляции экспрессии генов, позволяющий в перспективе максимально исполь-
зовать все имеющиеся эмпирические данные и минимизировать использование априорных гипо-
тез. При этом правила регуляции генов формулируются в виде лингвистических (сформулирован-
ных на языке близком человеческому) условий. Авторы используют более точно отображающий ре-
альность нечеткий подход, известный как нечеткое лингвистическое моделирование. Впервые
лингвистическое моделирование использовано для разработки моделей активности цис-регулятор-
ных модулей и применено к проблемам анализа экспрессии генов биохимической разметки эмбри-
онов. Такой подход позволил прояснить существенные детали механизмов «считывания» первич-
ных морфогенетических градиентов двумя или более цис-регуляторными модулями одного гена в
их взаимодействии.
Ключевые слова: регуляция генов, паттерны экспрессии, энхансеры, грамматика энхансеров, логическое
моделирование, нечеткое моделирование.
DOI: 10.31857/S0006302921010063
В настоящее время несколько разных подхо-
Понимание генной регуляции в целом и пове-
дов используются для изучения работы гена по-
дения генных регуляторных сетей (ГРС), в част-
средством моделирования [2]. Основная методо-
ности, это важнейшие фундаментальные пробле-
логическая проблема в этой области заключается
мы, на которые нацелена современная системная
в том, что зачастую трудно или даже невозможно
биология. Для этих задач применяется подход,
различить свойства модели, адекватно отобража-
основанный на данных (data-driven approach), в
ющие некоторые характеристики исследуемого
котором имеющиеся данные используются для
биологического объекта, от тех свойств модели,
разработки модели, а затем выводы, полученные
которые обусловлены природой подхода к моде-
путем компьютерных тестов, проверяются новы-
лированию. Прежде всего это относится к про-
блеме выбора априорных гипотез при моделиро-
ми экспериментами, и эти новые результаты ис-
вании. Например, модели генной регуляции,
пользуются для улучшения модели [1]. Поэтому в
исторически развиваемые в парадигме математи-
системно-биологической парадигме равно важ-
ческой физики (прежде всего, модели реакции-
ны как эксперименты, так и количественный
диффузии), характеризуются рядом черт, обу-
анализ и моделирование полученных данных.
словленных природой подхода, достаточно дале-
кого от биологических реалий.
Сокращения: ГРС - генные регуляторные сети, ЦРМ -
цис-регуляторные модули, ТФ - транскрипционные фак-
Одной из самых развитых областей системно-
торы, НЛМ - нечеткое лингвистическое моделирование.
биологического анализа генной регуляции явля-
49
50
МАКАШОВ и др.
ется моделирование динамики паттернов (биохи-
ДНК-матрицы и механизма транскрипции (см.
мической разметки) эмбриона дрозофилы. Здесь
работы [4, 15]). Это прежде всего обусловлено су-
наиболее популярными подходами являются
щественной ограниченностью наших знаний о
нейросетевое (коннекционное, connectionist) и
природе и молекулярных механизмах этой инте-
термодинамическое моделирование [3, 4]. Кон-
грации [17-20].
некционистские модели (или модели генных
Все эти представления побуждают исследова-
схем, gene circuits) относятся к классу «огрублен-
телей рассматривать другие подходы к моделиро-
ных» подходов, тогда как термодинамическое мо-
ванию поведения ГРС, более ориентированные
делирование можно рассматривать как более де-
на данные и позволяющие непосредственно
тальный уровень моделирования [5]. Помимо
включать в модель весь доступный эмпирический
этих подходов следует упомянуть упрощенные
материал и делать это простым, естественным пу-
логические модели, опубликованные в начале
тем. Мы считаем, что наилучшим способом сде-
2000-х годов [6, 7], а также комбинированные мо-
лать это является дальнейшее развитие семейства
дели последних лет, в которых объединяются
разнообразных подходов, обычно называемых
транскрипционная модель и дифференциальные
нечетким лингвистическим моделированием
уравнения [8-10].
(НЛМ) [2]. Эти подходы, в частности, используют
Коннекционистские модели рассматривают
конструкции «если ..., в таком случае..., еще ...»
ГРС как простой однослойный перцептрон (ис-
(«if ..., then..., else...») для словесного описания
кусственная нейронная сеть). Подход описывает
разнообразных взаимоотношений между регули-
ГРС системой уравнений математической физи-
рующими входными сигналами и путями их ин-
ки (обычно уравнениями реакции-диффузии), по
теграции в выходные сигналы. Это позволяет
одному уравнению для скорости экспрессии каж-
сформулировать с помощью простых правил из-
дого гена, при этом члены реакции основаны на
вестные детали разнообразной и иерархической
матрице межгенных регуляторных взаимодей-
регуляторной организации генов.
ствий (gene-gene action) [3, 5]. Метод неявно ос-
В этой работе мы развиваем подход НЛМ к ре-
нован на ожидании того, что организация связей
гуляции экспрессии генов с точностью до отдель-
у хорошо обученного перцептрона будет соответ-
ных ЦРМов. Конкретно мы сосредоточимся на
ствовать функциональной структуре изучаемой
примере регуляций генов сегментации эмбриона
ГРС [11]. Эти ожидания очень старые, и такие мо-
дрозофилы, как одном из самых исследованных
дели, как оказалось, хороши для моделирования
модельных объектов.
поведения ГРС в норме (дикого типа), но подоб-
ное моделирование не всегда подходит для того,
Нечеткие логические модели регуляции генов.
чтобы предсказать последствия мутаций, возму-
Поведение биологических систем по своей при-
щений или шума (низкая прогностическая сила
роде нечеткое. Несмотря на то что новые высоко-
[12]). Нам известны только работы по моделиро-
производительные технологии предоставляют
ванию мутантов по генам Kr [13] и bcd [14].
биологам огромное количество ценных данных,
необходимы методы, которые включают в моде-
Термодинамическое моделирование основано
лирование погрешность (неточность, изменчи-
на экспериментальных или биоинформационных
вость), позволяя иметь дело со многими генами с
знаниях об организации регуляторных областей
невыясненной функцией и с «зашумленными»
(энхансеров или цис-регуляторных модулей,
экспериментальными данными. Для получения
ЦРМ) данного гена [2, 4, 15]. Это количественное
биологически значимых результатов информа-
моделирование для прогнозирования экспрессии
ция должна объединяться из разных источников
генов исходя из анализа их нуклеотидной после-
для конструирования моделей ГРС. Такие нечет-
довательности, и оно основано на подходах ста-
кие экспертные знания включают базы данных о
тистической термодинамики. Статистическая
генах и их продуктах, а также информацию о вза-
термодинамика характеризует специфическое
имодействиях между генами.
связывание транскрипционных факторов (ТФ) с
сайтами связывания на ДНК в равновесных усло-
Нечеткая логика, как фундаментальная со-
виях [16]. Таким образом, подход явно включает
ставляющая нечеткого метода, представляет со-
наши знания о сайтах связывания для ТФ, их си-
бой комбинацию различных математических
ле, ориентации и точной позиции. Это дает лож-
принципов представления знаний об изучаемом
ное впечатление, что подход рассматривает точ-
явлении в нечетком, не бинарном виде (в отличие
ную экспериментально выявленную регулятор-
от Булева подхода). Метод нечеткой логики со-
ную организацию энхансеров. Но на самом деле
стоит из ряда процедур, которые используются
это не так: подход обычно игнорирует многие де-
для сопоставления определенного входящего и
тали иерархической организации регуляторных
ему соответствующего выходящего сигнала: про-
взаимодействий. Он обычно игнорирует неадди-
цесс, называемый «нечетким выводом». Двумя
тивный, синергетический характер интеграции
наиболее известными методами вывода являются
регуляторного входящего сигнала при помощи
те, которые были связаны с именами E. Mamdani
БИОФИЗИКА том 66
№ 1
2021
НЕЧЕТКОЕ МОДЕЛИРОВАНИЕ РЕГУЛЯЦИИ ГЕНОВ
51
и M. Sugeno [21, 22]. Эти модели зависят от ис-
циональные генно-регуляторные модули, ЦРМ
пользования простого языка правил «if-then»,
[32-36]. Это можно назвать грамматическими
описывающего ответ системы как функции не-
правилами для энхансера или грамматикой цис-
скольких лингвистических переменных.
регуляторного модуля [30, 31, 37]. Это способ пе-
реформулировать экспериментальные выводы,
Лингвистические описания (или описания на
выраженные на языке экспериментаторов, на
«естественных языках», в отличие от, например,
язык, используемый для разработки моделей. В
формализма математической физики) наблюдае-
свою очередь, область грамматики генно-регуля-
мых процессов могут быть легко переформулиро-
торных элементов естественно отнести к более
ваны в предложения типа «if-then», которые от-
общей области проблем на стыке молекулярной
ражают поведение системы, без необходимости
биологии и лингвистики [38, 39].
использовать математические функции. Как пра-
вило, такие описания знаний на основе «есте-
Несмотря на относительную развитость ряда
ственного языка» явно или косвенно включают
направлений НЛМ на уровне грубого моделиро-
дискретизированные описания объектов или от-
вания, попытки развить подходы нечеткого моде-
носятся к ним (например, низкие, средние и вы-
лирования на уровне детального (fine-grained)
сокие концентрации).
описания находятся еще в самом зачаточном со-
стоянии (см. работу [40]). В этой статье мы пред-
Таким образом, представляется весьма пер-
лагаем нашу версию НЛМ на уровне детального
спективным развивать компьютерный подход,
моделирования. Мы соединяем подходы нечет-
позволяющий прямое преобразование каче-
кого моделирования с подходами, исходящими
ственных знаний и описаний в создаваемую ра-
из формулировки правил грамматики цис-регу-
бочую модель. При этом первым шагом будет
ляторных элементов и подходами к нахождению
найти подходящее формальное представление
логик генной регуляции при действии ТФ.
дискретных, неточных естественных языковых
терминов, которые описывают состояния объек-
Примечательно, что именно подход к модели-
тов. Второй шаг - найти подходящее формальное
рованию динамики генных сетей, основанный на
представление функций, которое позволит непо-
нечеткой логике [41], оказался наиболее эффек-
средственно объединить качественные знания на
тивным для тестовых задач обратной инженерии
основе естественного языка и тем самым облег-
функциональной организации ГРС по сравне-
чить интерпретацию функциональности.
нию с рядом других подходов ([42], проект «The
Развиваемый подход имеет отношение к логи-
#!Synapse:syn3049712/wiki/74631)). Это побуждает
ческому моделированию генной регуляции -
нас развить аналогичный подход для моделирова-
подходам к нахождению логик генной регуляции
ния паттернов экспрессии генов, с точностью до
(gene-regulatory logic inferring [23]). Логические
отдельных (полу)автономных ЦРМов.
вентили (logic gates), которые используют более
одного входа, уже давно применяются для описа-
Мы намереваемся интегрировать данные экс-
ния сложных взаимодействий между ТФ, специ-
прессии генов и информацию о связывании ТФ с
фически связывающимися со своими сайтами на
ДНК с конечной целью расшифровки логики ре-
генно-регуляторном элементе. Регуляторная
гуляции экспрессии гена. Такой подход может
функция, которой описываются цис-регулятор-
количественно охарактеризовать закономерно-
ные взаимодействия между ТФ, связанными со
сти взаимодействия между ТФ, объединяя цис-
своими сайтами, и управляющими экспрессией
регуляторные логики и кинетику транскрипции в
их гена-мишени, может быть представлена в виде
одной модели. Полученные логики цис-регуля-
Булевых логик [21, 24]. Соответственно, переход
ций могут затем использоваться для установле-
от булевой логики к нечеткой представляется
ния функциональной организации ГРС. Мы ил-
естественным обобщением.
люстрируем такой подход на тестовом примере
одного из наиболее изученных наборов ЦРМов -
В недавние годы опубликованы серии работ,
энхансеров гена hunchback, hb [43; 44; 45; 46; 47;
реализующих моделирование на основе нечеткой
48; 49; 50; 51]. Он служит экспериментальной мо-
логики (преимущественно, гибридные подходы)
делью для изучения многих других цис-регуля-
на уровне грубого моделирования ГРС (нечеткие
торных модулей в биологии развития и, в
модели генных сетей, fuzzy gene-circuit models)
более общем плане, в эволюционной биологии
[25-29]. Вместе с тем в системной биологии про-
(evo-devo).
должаются разработки моделей детального уров-
ня, включающих формулирование генно-регуля-
торной логики (с целью расшифровать логику
НАШ ПОДХОД
действия ТФ на генно-регуляторные элементы
при контроле транскрипции) [30, 31]. Параллель-
Мы развиваем наш подход НЛМ к моделям
но продолжаются работы по выяснению правил
генной экспрессии на уровне детального модели-
организации сайтов связывания для ТФ в функ-
рования (уровень кластеров сайтов связывания
БИОФИЗИКА том 66
№ 1
2021
52
МАКАШОВ и др.
для ТФ). Этот уровень основан на детальных зна-
Морфогенетический градиент фактора Bicoid
ниях о функциональной организации генных ре-
(Bcd) является одной из наиболее изученных мо-
гуляторных элементов и наиболее приближен к
делей эволюционной биологии развития [48, 59].
биологическим реалиям, в отличие от упрощен-
Фактор Bcd активирует набор генов сегментации
ных подходов грубого уровня.
дрозофилы через набор энхансеров, управляемых
Bcd. Было описано несколько десятков активиру-
Мы используем анализ грамматики энхансе-
ющихся при помощи Bcd энхансеров, действую-
ров (правила размещения сайтов связывания друг
щих в начале развития дрозофилы, и некоторые
относительно друга, включая расстояния между
из них (прежде всего проксимальный элемент hb
ними), принадлежащих к одному семейству, для
и элемент 2-й полосы гена even-skipped) были по-
формулировки генно-регуляторной логики в ви-
дробно изучены [15, 45, 47]. Функциональным яд-
де наборов регуляторных функций [28, 29]. Зна-
ром таких элементов является кластер сайтов свя-
ние этой грамматики позволяет в итоге сформу-
зывания фактора Bcd (рис. 1а,б). Bcd обычно тре-
лировать регуляторные функции для описания
бует несколько сайтов связывания для активации
поведения ГРС. Эти правила могут быть исполь-
экспрессии генов, и эта активация обычно явля-
зованы для развития НЛМ детального уровня.
ется кооперативной.
Самое общее преимущество НЛМ по сравне-
Энхансеры, активируемые Bcd, как и многие
нию с другими подходами к моделированию по-
другие ЦРМ, интегрируют множество регулятор-
ведения генных сетей заключается в том, что:
ных входных сигналов от активаторов и ко-акти-
1) подход не использует априорных моделей
ваторов и репрессоров, и ко-репрессоров для
механизмов генной регуляции и устроен как
обеспечения воспроизводимости и устойчивости
«черный ящик» (на входе регулирующие парамет-
раннего эмбрионального паттерна (рис. 1). Регу-
ры, на выходе уровень экспрессии гена), так что
лирование является иерархическим и включает в
зависимость выхода от входных параметров опре-
себя как минимум три уровня.
деляется системой нечеткого логического вы-
Первый уровень - это уровень «раскрытия»
вода,
хроматина, а ключевым фактором, участвующим
2) нечеткий логический вывод в силу простоты
в открытии, является Zelda, Zld (рис. 1б). Zld
организации находит конкретное частное реше-
представляет собой сайт-специфический TF, ко-
ние простыми формальными процедурами и ра-
торый также помогает связывать другие факторы,
ботает существенно быстрее по сравнению, на-
изменяя локальную доступность хроматина, тем
пример, с моделированием системами диффе-
самым играя главную роль в раскрытии конден-
ренциальных уравнений, что многократно
сированного хроматина. Исследователи называ-
ускоряет поиск при оптимизации эволюционны-
ют Zld-подобные факторы факторами-«пионера-
ми методами.
ми», поскольку они контролируют активацию ге-
нома зиготы через их способность раскрывать
Развиваемый здесь подход, восходящий к
конденсированный хроматин [60, 61]].
классическим публикациям по нечеткому моде-
лированию [52], можно трактовать как развитие
Второй уровень - типичное кооперативное
подхода кусочно-линейных аппроксимаций не-
связывание ДНК, например, между факторами
линейных функций в приложении к разработке и
Bcd, Hb и другими ключевыми активаторами, и
анализу нелинейных динамических моделей ген-
коактиваторами. Этот эффект увеличивает общее
ных и клеточных регуляторных сетей (см. работы
сродство связывания, что приводит к согласован-
[53, 54]).
ной «загрузке» сайтов при более низком общем
уровне концентрации ТФ. Как результат, это
Наш объект изучения. Почти 100 охарактеризо-
обеспечивает и резкий регуляторный «скачок» в
ванных энхансеров контролируют формирование
ответ на относительно небольшое увеличение
паттерна раннего эмбриона дрозофилы, вероят-
концентрации активаторов. Далее, обсуждаемые
но, наиболее изученный процесс эмбрионально-
энхансеры всегда содержат сайты связывания для
го развития ([45, 55-57]). Эти энхансеры регули-
факторов-репрессоров, как правило, вблизи сай-
руют экспрессию порядка 50 генов, контролиру-
тов для активаторов (рис. 1). Репрессоры обычно
ющих формирование передне-заднего и дорзо-
характеризуются как короткодействующие: они
вентрального паттернов на этапах сегментации и
действуют репрессивно на соседние с ними свя-
гаструляции. Такой массив экспериментальных и
занные активаторы.
биоинформационных данных дает беспрецедент-
ную возможность анализа регуляторной грамма-
Третий уровень представляет собой тран-
тики этих энхансеров, контролируемых пример-
скрипционную синергию, в которой многочис-
но 30 ТФ. Результаты геномного секвенирования
ленные специфически связанные ТФ взаимодей-
целого ряда различных видов дрозофилы делает
ствуют с компонентами транскрипционной ма-
возможным анализ эволюции этих энхансеров
шины, либо с базальными субъединицами, либо с
[58].
общими ко-активаторами транскрипции
[62].
БИОФИЗИКА том 66
№ 1
2021
НЕЧЕТКОЕ МОДЕЛИРОВАНИЕ РЕГУЛЯЦИИ ГЕНОВ
53
Рис. 1. Детали функциональной организации трех наиболее изученных энхансеров гена hunchback (hb): проксималь-
ный энхансер (а), дистальный («теневой», distal anterior enhancer, DAE) энхансер (б) и энхансер «полос» (stripe-ele-
ment) (в). Указаны позиции сайтов связывания ключевых транскрипционных факторов (найденных эксперименталь-
но или биоинформационными методами). Хорошо видна тенденция сайтов связывания находиться в кластерах. Сай-
ты активаторов показаны над элементом, а сайты репрессии - под ним: Bcd - Bicoid, Kr - Kruppel, Kni - Knirps, Gt-
Giant, Zd - Zelda (согласно [47] и [64], с изменениями.
Конкретно, например, показано, что Bcd активи-
Проксимальный элемент (рис. 1а) безусловно
рует синергическую транскрипцию, взаимодей-
относится к одним из самых изученных. Тем не
ствуя с TBP-ассоциированными факторами
менее многие критические детали его функцио-
TAFII60 и TAFII110 [17, 18]. Если эти факторы от-
нирования по сию пору неясны. Наиболее изу-
сутствуют, то активации транскрипции не проис-
ченная роль этого элемента - резко активировать
ходит.
ген hb при концентрации бикоида выше критиче-
ской [47, 48, 67-73]. Сходным образом работает
Цис-регуляторные модули гена hunchback. Со-
целый ряд ЦРМ ранних генов эмбриона дрозо-
гласно относительно недавним (и совсем недав-
филы, и этот энхансер считается для них прото-
ним) публикациям, ранний паттерн экспрессии
типическим. Элемент имеет как несколько экс-
гена hb находится под контролем трех ЦРМ:
периментально охарактеризованных сайтов для
проксимального, теневого и энхансера полос [46,
фактора Bcd, так и ряд сайтов, обнаруживаемых
47, 63, 64], что иллюстрировано схемами на рис. 1
биоинформационными методами. Резкий функ-
(данные сведены на нашем веб-ресурсе HOX pro
циональный отклик (почти по типу «все или ни-
[65, 66]). При этом функции проксимального
чего») объясняется хорошо исследованной ко-
и теневого элементов весьма схожи (что ставит
оперативностью связывания (и действия) факто-
вопрос о роли такой функциональной избыточ-
ра Bcd с этим элементом [48, 74, 75]. Помимо
ности) [46, 47]. Все три элемента работают на ин-
этого, в проксимальном энхансере обнаружены
тересующей нас здесь стадии раннего эмбриоге-
(и частично охарактеризованы эксперименталь-
неза - четырнадцатом цикле. Согласно недавним
но) сайты для Hb и Kr (рис. 1а), как и некоторых
исследованиям, вклад этих элементов в суммар-
других ТФ (не приведены на рисунке). Такая из-
ный уровень экспрессии гена hb можно тракто-
быточность сайтов как для активации, так и для
вать как близкий к аддитивному [63].
репрессии нуждается в дальнейшем анализе. Ос-
БИОФИЗИКА том 66
№ 1
2021
54
МАКАШОВ и др.
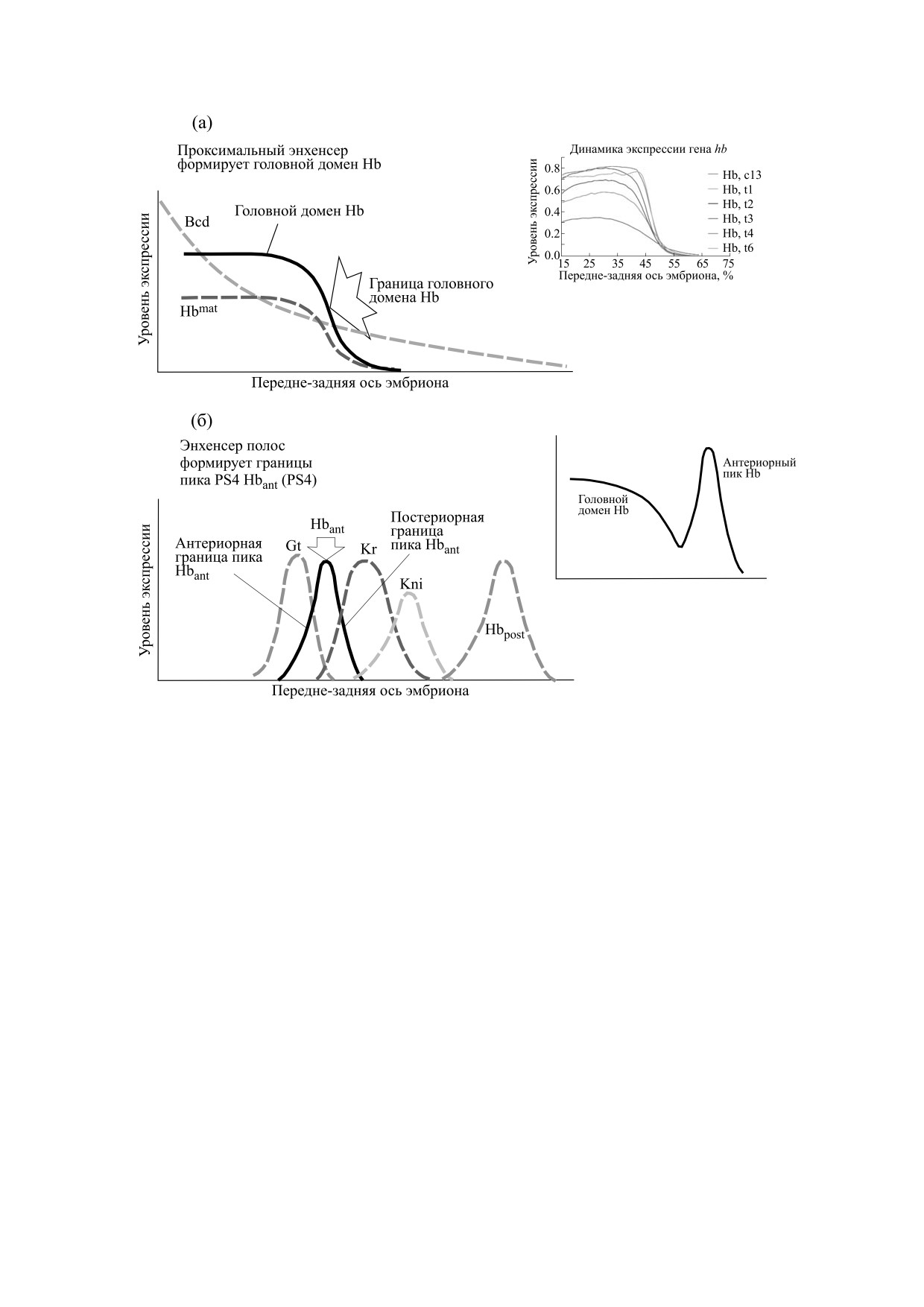
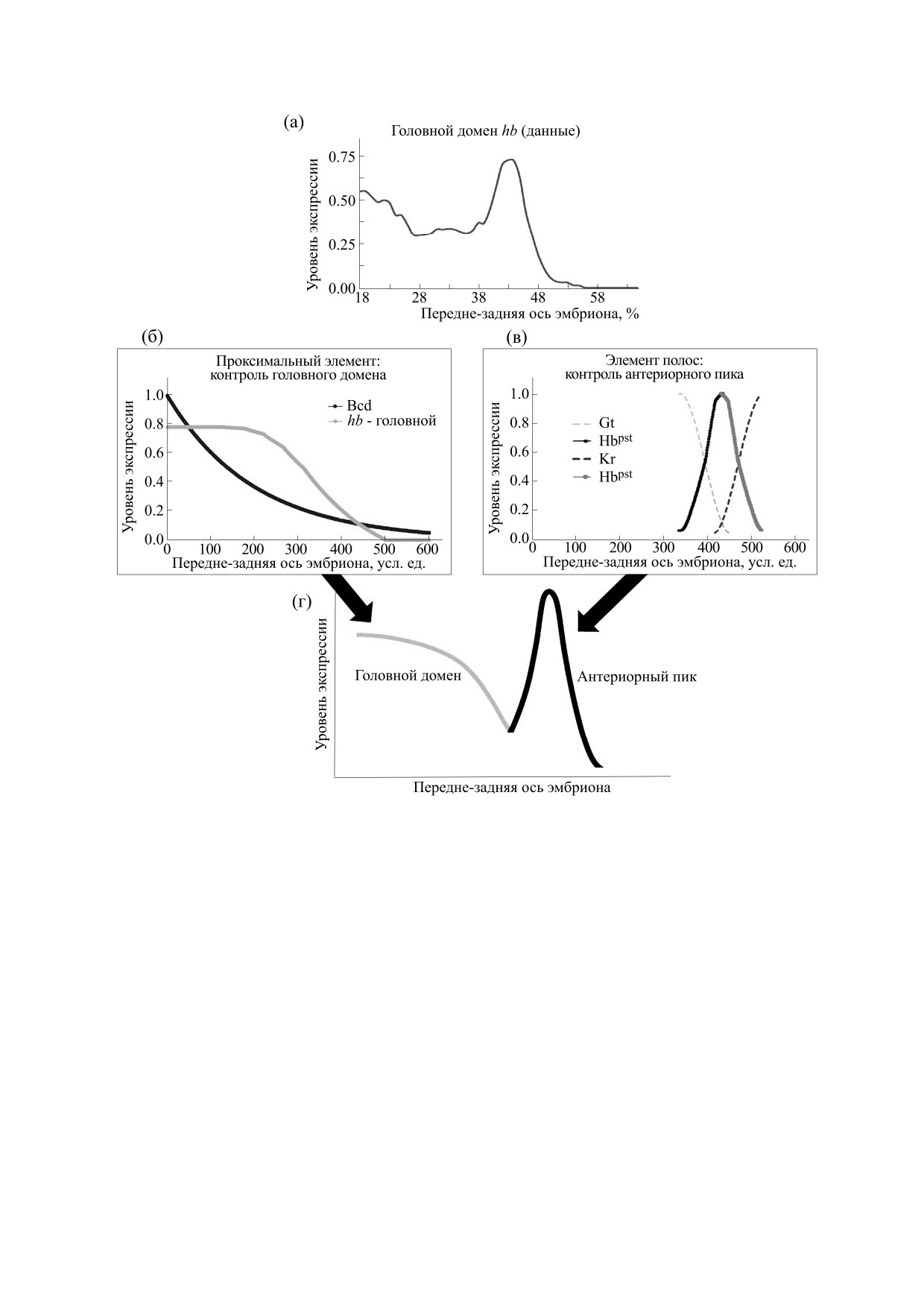
Рис. 2. Профиль экспрессии гена hb в головной половине раннего эмбриона дрозофилы включает головной домен и
антериорный пик (Hbant). Их формирование контролируется тремя (полу)автономными энхансерами. (а) - Роль
проксимального энхансера. В самом раннем развитии и до начала четырнадцатого цикла ген hb находится под
преимущественным контролем активатора Bicoid (Bcd) через проксимальный (и теневой) энхансеры. Это определяет
появление, рост и формирование головного домена hb со все более крутой (постериорной) границей (склоном) этого
домена. Приведен профиль материнского фактора Hb, также контролирующего головной домен. На врезке показано,
как меняется геометрия головного домена в раннем эмбриогенезе: склон становится все более крутым. (б) - Роль
энхансера полос. С начала четырнадцатого цикла контроль над hb переходит к элементу полос (см текст). Энхансер
контролирует формирование антериорного и постериорного (Hbpost) пиков (полос). На врезке иллюстрируются
вклады проксимального энхансера и энхансера полос в итоговую картину экспрессии гена hb в четырнадцатом цикле
(головной домен + антериорный пик). Экспрессия измеряется в относительных условных единицах от 0 до 1.
новная проблема заключается в том, что крутизна
задней частях эмбриона (антериорный и постери-
отклика гена hb в целом слишком велика, чтобы
орный пики или полосы, Hbant и Hbpost, как на
объяснить ее только кооперативностью действия
рис. 2б). (Hbant известен также под именем пик
Bcd (врезка на рис. 2а иллюстрирует этот рост
PS4).
крутизны границы домена hb). Вопрос о том, как
и какие именно другие ТФ участвуют в этих про-
Полагают, что для элемента полос ген hb акти-
цессах, дающих в итоге отклик «все или ничего»,
вируется какими-то малоисследованными общи-
требует дальнейшего изучения и объяснения.
ми активаторами, а роль этого элемента состоит
именно в репрессии hb всюду, за исключением
Энхансер полос включает в себя сайты связы-
двух позиций, где и формируются пики экспрес-
вания для факторов Kr, Kni, Hb [47, 76] (рис. 1в).
сии. При этом, в частности, склоны антериорно-
Под контролем энхансера полос ген-мишень дает
го пика «справа» определяются доменом репрес-
два четких острых пика экспрессии в передней и
сора Kr, а «слева» (как полагают) - доменом
БИОФИЗИКА том 66
№ 1
2021
НЕЧЕТКОЕ МОДЕЛИРОВАНИЕ РЕГУЛЯЦИИ ГЕНОВ
55
Рис. 3. Диаграмма системы нечеткой логики в приложении к проблеме моделирования экспрессии генного
регуляторного элемента (энхансера). Машина вывода (inference engine) использует нечеткие функции генной
регуляции, которые формулируются на основе анализа грамматики энхансера (см. текст).
головного Hb (саморепрессия), как иллюстриру-
и простой процедурой дефаззификации по высо-
ется рис. 2б.
те (height defuzzification) [78].
Детали и механизмы действия энхансера полос
Для нашего набора энхансеров (тест-объек-
все еще мало исследованы. В частности, неясны
тов) мы формулируем набор грамматических пра-
механизмы репрессии и степень их кооператив-
вил, позволяющий определять вид и характери-
ности.
стики регуляторных функций как кусочно-ли-
Моделирование активности цис-регуляторных
нейных средствами НЛМ (см. работы [52-54]).
модулей с использованием нечеткой логики. Мы
Конкретно мы делаем такую аппроксимацию для
используем стандартное общее представление
функций активации и репрессии гена-мишени
для НЛМ, приведенное на рис. 3. Специфика на-
транскрипционным фактором-эффектором.
шего подхода заключается в приложении общей
Мы исходим из того, что каждый ген, контро-
стратегии НЛМ к конкретике деталей функцио-
лирующий формирование эмбрионального пат-
нирования анализируемого семейства энхансе-
терна, состоит из нескольких (полу-) автономных
ров.
регуляторных элементов (ЦРМ), так что каждый
Взаимодействия между биологическими объ-
такой элемент описывается своей нечеткой моде-
ектами представляют собой процессы, которые
лью. На выходе такой модели уровень экспрессии
влияют на будущее состояние объектов-мишеней
гена, определяемый рассматриваемым ЦРМ, то-
исходя из текущего состояния объектов-эффек-
гда как на входе - уровни концентраций сайт-
торов. Компьютерные модели имитируют взаи-
специфических ТФ, способных специфически
модействия при помощи функций, которые опе-
связываться с рассматриваемым ЦРМ и действо-
рируют вычислительными представлениями о со-
вать на ген через него.
стояниях. Эти функции отображают текущие
Нечеткие функции генной регуляции. Даже са-
состояния эффекторов (входящие данные) в но-
мый поверхностный анализ динамики паттернов
вые состояния или изменения состояния мише-
экспрессии гена hb показывает, что кинетика
ней (выходные данные), как на рис. 3.
действия и активаторов и репрессоров нелиней-
В общем случае механизм логического вывода
на. Мы ниже приходим к заключению, что для
включает фаззификацию, нечеткий вывод (fuzzy
наших целей достаточно использовать признан-
inference), и дефаззификацию (рис. 3). На входе
ную и достаточно простую модель регуляции рас-
нашей НЛМ находится набор уровней экспрес-
сматриваемого гена данным фактором (через
сии регуляторных факторов, контролирующих
данный ЦРМ) - модель Хилла. Модель Хилла
экспрессию гена-мишени через анализируемый
описывает или активацию или репрессию. Мы,
нами энхансер этого гена (сравни рис. 1 и 2), на
естественно, используем нечеткую формулиров-
выходе - уровень экспрессии (т. е. уровень про-
ку этого закона. НЛМ могут быть сформулирова-
дукции мРНК) нашего гена-мишени. (Для еди-
ны таким образом, чтобы они аппроксимировали
нообразия экспрессия генов приведена и в экспе-
сигмоидные профили функций Хилла, описыва-
риментальных данных, и на выходе эксперимента
ющие активирующие и ингибирующие эффекты,
к относительным условным единицам от 0 до 1.)
что иллюстрируется на рис. 4.
Процедуру фаззификации в нашем случае
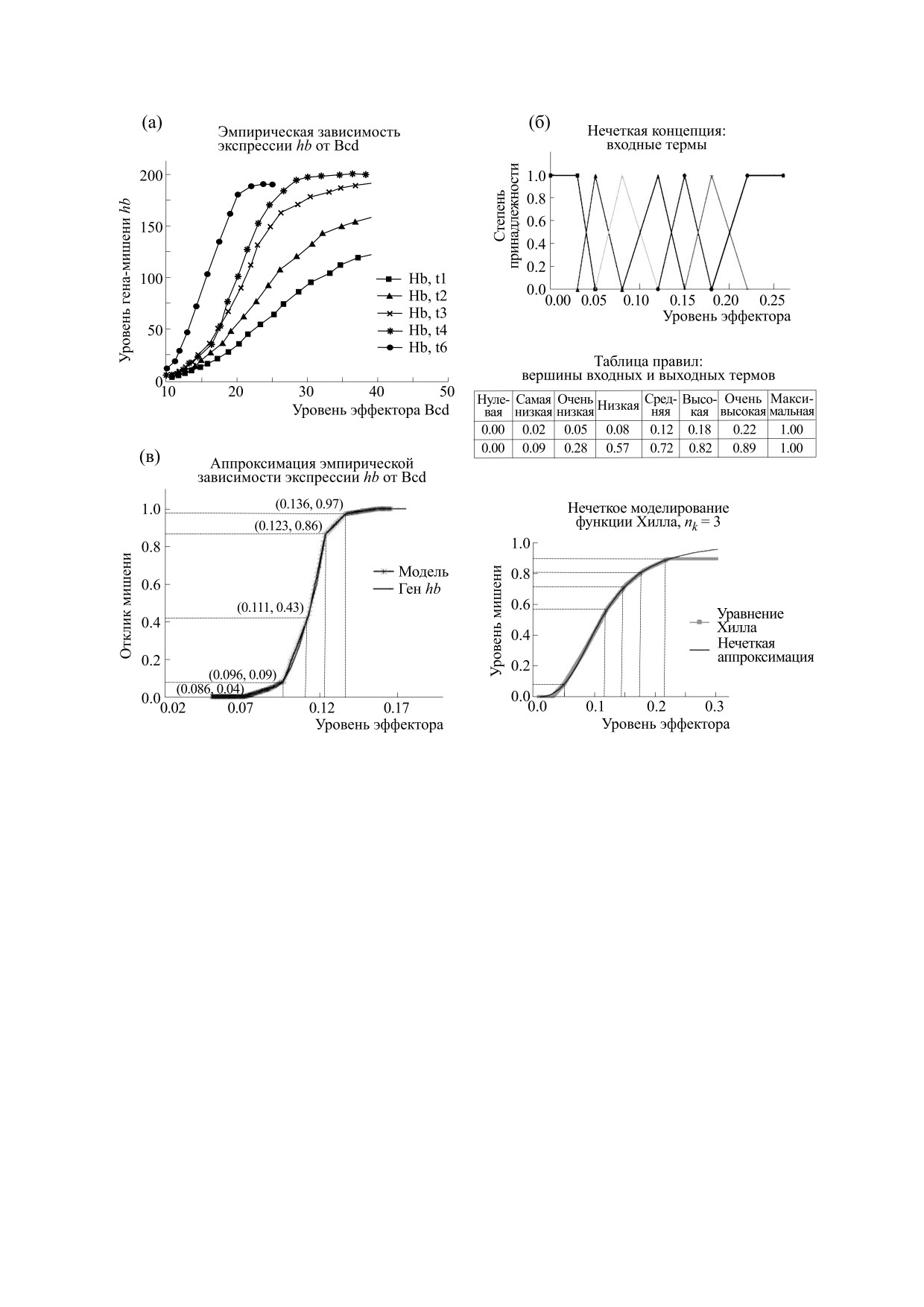
Достаточно четырех нечетких множеств и на-
естественно выполнять исходя из треугольных
бора простых правил для создания кусочно-ли-
термов с включением краевых трапециевидных
нейных аппроксимаций функций Хилла для ак-
термов [77] (см. рис. 4б). Для стадии дефаззифи-
тивации. Качество аппроксимации зависит от ко-
кации мы воспользуемся широко употребляемой
личества нечетких множеств, но оно уже
БИОФИЗИКА том 66
№ 1
2021
56
МАКАШОВ и др.
Рис. 4. Нечеткое представление функции Хилла для задач моделирования кооперативного действия активатора на ген-
мишень. (а) - Эмпирические зависимости экспрессии гена hb от уровней кооперативного активатора Bcd на ранних
стадиях эмбриогенеза дрозофилы (по материалам веб-ресурса FlyEx [91]). (б) - Детали подхода к нечеткому
представлению функции Хилла (коэффициент Хилла nk = 3) пятью термами: два крайних, имеющих форму трапеции,
и три средних - треугольные; представлены входные термы нечеткой концепции, таблица правил и результат
кусочно-линейной аппроксимации этой функции. (в) - Пример конкретного разбиения эмпирической зависимости
hb от Bcd (для временного класса t1 четырнадцатого цикла; сравни с панелью (а)) на пять термов. Эмпирический
график аппроксимирован отрезками прямых. Отмечены координаты точек перелома аппроксимации. Координаты
границ отрезков (по оси эффектор) соответствуют центрам входных термов, тогда как их координаты по оси «ген-
мишень» определяют значение центров выходных термов (эти пары значений приведены в скобках; см текст).
Экспрессия измеряется в относительных условных единицах от 0 до 1.
удовлетворительно, если даже используются
раметров самой функции Хилла, должен
только четыре нечетких множества (ср. [79]). Мы
решаться оптимизацией при адаптации модели.
в нашем моделировании использовали пять или
Наши алгоритмы процедур нечеткой логики. В
шесть термов для аппроксимации функций Хил-
описании подхода мы будем пользоваться терми-
ла (рис. 4).
нами эффектор и ген-мишень. Под эффектором
Используя только три нечетких множества,
типично понимается сайт-специфический ТФ и
можно задать удовлетворительную кусочно-ли-
предполагается, что в простейшем случае он дей-
нейную аппроксимацию функций Хилла для ко-
ствует на ген-мишень (через ЦРМ этого гена) или
оперативной репрессии [79]. В общем случае во-
как активатор, или как репрессор. Ниже мы часто
прос параметров фаззификации, как и вопрос па-
будем использовать понятие терм для лингвисти-
БИОФИЗИКА том 66
№ 1
2021
НЕЧЕТКОЕ МОДЕЛИРОВАНИЕ РЕГУЛЯЦИИ ГЕНОВ
57
ческой переменной, определяющей уровни экс-
экспрессии гена-мишени в правом конце этого
прессии эффектора и мишени.
участка.
Входные (и выходные) термы. Входные и выход-
Наши алгоритмы для модели двух эффекторов
ные термы определяются (строятся) по такому
одной мишени. Здесь мы рассмотрим реализацию
алгоритму.
нашего подхода на примере активации генной
экспрессии для случая, когда на ген-мишень дей-
Строим эмпирический график зависимости
ствует не один (как выше), а два эффектора. На-
уровня экспрессии гена-мишени от уровня эф-
пример, у нас имеются два фактора-активатора,
фектора (на примере активатора, как на рис. 4б).
А1 и А2, значения каждого из которых находятся
Делим профиль зависимости гена-мишени от эф-
в интервале [0, 1], определяющие вместе уровень
фектора на области, в которых профиль близок к
экспрессии мишени (который тоже определяется
линейному. Таким образом, профиль аппрокси-
на интервале [0, 1]). Пусть для простоты уровень
мируется ломаной. Области не должны пересе-
фактор А1 описывается двумя термами («отсут-
каться и в совокупности давать весь отрезок (0, 1).
ствует» и «присутствует»), тогда как фактора А2 -
Количество областей равно количеству входных
тремя: «низкий», «средний» или «высокий».
термов.
Далее мы определяем нечеткое множество для
Строим треугольные термы, вершины которых
гена-мишени как три терма для скорости тран-
совпадают со значениями уровня гена-активато-
скрипции: «нулевая», «низкая» и «высокая». Со-
ра в звеньях ломаной на графике (рис. 4б, внизу).
ответственно, система нечетких правил (П1-П6)
Координаты вершин занесены в верхнюю строку
для нашей модели может быть определена как на-
таблицы на рис. 4б. Пусть всего имеется n термов
бор правил общего вида «if-then» («если-тогда»):
(в примере n = 5). Тогда элементы таблицы в об-
щем виде заданы [0, В1, В2, …, Вn, 1], где Вi - ко-
П1: ЕСЛИ x1 = отсутствует И x2 = низкий, ТО-
ордината вершины i-го терма. Построение тер-
ГДА y = нулевая;
мов проиллюстрировано на рис. (4б, вверху). Тре-
П2: ЕСЛИ x1 = отсутствует И x2 = средний,
угольный терм имеет вершину в точке Вi со
ТОГДА y = нулевая;
значением 1 и вершины в точках Вi-1 и Вi+1 со
П3: ЕСЛИ x1 = отсутствует И x2 = высокий,
значением 0. Два крайних терма имеют вид трапе-
ТОГДА y = нулевая;
ции, т.е. в вершинах с координатами 0 и В1 у пер-
П4: ЕСЛИ x1 = присутствует И x2 = низкий,
вого терма и с координатами Вn и 1 у последнего
ТОГДА y = нулевая;
терма имеют значения, равные единице.
П5: ЕСЛИ x1 = присутствует И x2 = средний,
Определяем значения уровней гена-мишени в
ТОГДА y = медленная;
звеньях ломаной (рис. 4б, внизу). Эти значения,
являющиеся центрами выходных термов, занесе-
П6: ЕСЛИ x1 = присутствует И x2 = высокий,
ны во вторую строку таблицы на рис. 4б. По этим
ТОГДА y = быстрая.
координатам строятся выходные термы, следуя
Такой набор правил нагляднее представить
тем же правилам, что и входные.
таблицей (табл. 1).
Генерация термов в виде треугольников или
Рассмотрим в заключение более общий слу-
трапеций на основе их центров - в общем случае
чай, когда оба эффектора имеют по три уровня
является непростой задачей, так как необходимо
(рассмотрим на примере активации). Скорости
учитывать расстояния между центрами и следить
транскрипции теперь характеризуются пятью
за тем, чтобы центр масс фигуры совпадал с ис-
термами. Поскольку запись базой правил стано-
пользуемым центром.
вится слишком громоздкой, то приведем только
База правил: После определения термов кон-
табличный вид (табл. 2).
струируется база правил. Для этого на каждом ли-
Сходные построения используются для случая
нейном участке ломаной, начиная с левого, ста-
пары репрессоров, R1 и R2.
вится соответствие между номером входного тер-
ма, соответствующего участку, и номером
Пример базы правил для трех термов приведен
выходного терма, чей центр совпадает с уровнем
в табл. 3.
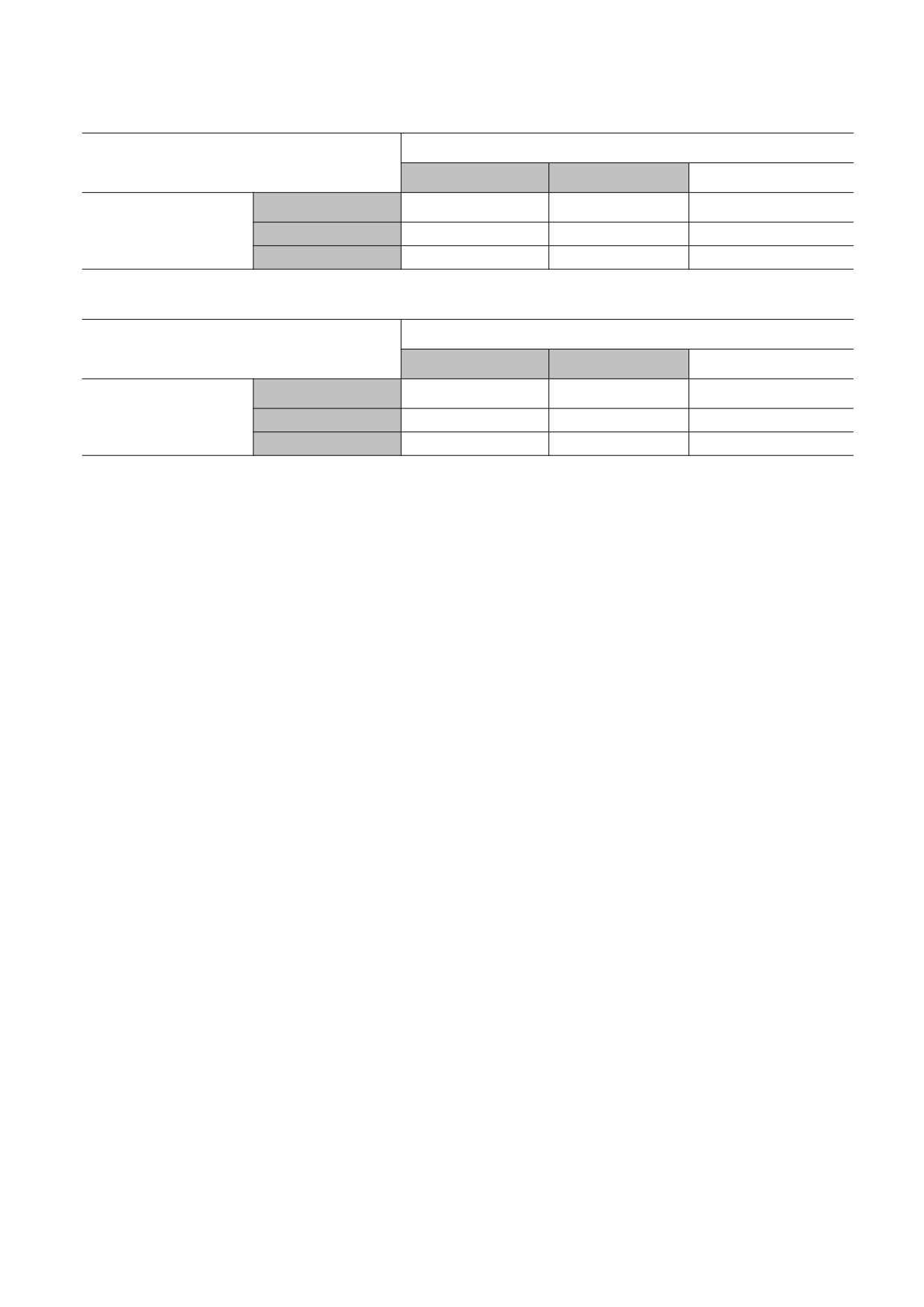
Таблица 1. Скорости транскрипции при разных значениях активаторов А1 и А2 (2 × 3)
Уровень фактора А1
Низкий
Средний
Высокий
Отсутствует
Нулевая
Нулевая
Нулевая
Уровень фактора А2
Присутствует
Нулевая
Медленная
Быстрая
БИОФИЗИКА том 66
№ 1
2021
58
МАКАШОВ и др.
Таблица 2. Скорости транскрипции при разных значениях активаторов А1 и А2 (3 × 3)
Уровень фактора А1
Низкий
Средний
Высокий
Уровень
Низкий
Нулевая
Самая медленная
Медленная
фактора А2
Средний
Самая медленная
Медленная
Быстрая
Высокий
Медленная
Быстрая
Самая быстрая
Таблица 3. Скорости транскрипции при разных значениях активаторов R1 и R2 (3 × 3)
Уровень фактора R1
Низкий
Средний
Высокий
Уровень
Низкий
Самая быстрая
Быстрая
Медленная
фактора R2
Средний
Быстрая
Медленная
Самая медленная
Высокий
Медленная
Самая медленная
Нулевая
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
4) Энхансер включает в себя несколько сайтов
связывания для близкодействующих репрессо-
Известно, что Bcd имеет тенденцию действо-
ров, и каждый сайт активации должен находиться
вать как активатор кооперативно, вместе с други-
поблизости (> 100 п.н.) от сайта репрессора.
ми (ко)-активаторами и кооперативность диктует
В противном случае энхансер будет работать в
свои правила для организации энхансеров. Мы
областях эмбриона, где он должен быть репресси-
можем назвать это грамматическими правилами
рован.
[32, 33, 37]. В наиболее изученных случаях можно
сформулировать грамматические правила, в част-
Полное детальное моделирование действия
ности, для силы, порядка и длины спейсеров для
энхансера должно в явном виде описывать три
комплекса кластеров сайтов связывания (см. ра-
иерархических уровня энхансера, управляемого
боты [30, 35, 45]).
морфогеном, с помощью пионерных факторов
(i), кооперативного связывания с матрицей акти-
Энхансеры, управляемые морфогенами. Для
ваторов (ii) и синергии с выходным механизмом
лингвистического описания функционирования
транскрипции (iii). В этой статье мы сосредото-
энхансера было бы разумно определить функцио-
чимся на нечетком моделировании только уровня
нальную организацию таких ЦРМ. Для этого не-
ii в сравнении кооперативной активации с коопе-
обходимо сформулировать грамматические пра-
ративной репрессией.
вила, описывающие энхансер.
Нечеткие правила грамматики энхансеров. Как
Главные правила (на примере энхансеров,
неоднократно отмечалось, в основе одного из
управляемых Bcd, как на рис. 1 (ср. с рис. 2):
наиболее изученных ЦРМ - проксимального эн-
1) Типичный энхансер, управляемый Bcd,
хансера - кластер сайтов связывания фактора
включает несколько сайтов связывания (рассеян-
Bcd (как иллюстрирует рис. 1а). Более того, функ-
ных по последовательностям энхансера) для пио-
ционально близкий (и функционально избыточ-
нерных факторов, таких как Zld или другие ко-
ный) теневой элемент также имеет в своей основе
факторы, способные «открывать» хроматин. В
кластер сайтов связывания Bcd (рис. 1б).
противном случае энхансер будет слишком слаб,
Кооперативность активации гена hb фактором
чтобы активироваться Bcd.
Bcd вообще и через проксимальный элемент в
2) Ядро энхансера состоит из кластера сайтов
частности многократно обсуждалась в литературе
связывания Bcd. Если два или более соседних
(см. работу [48] и ссылки в ней). Поэтому у нас
сайта расположены слишком близко друг к другу
есть все основания сравнивать нашу нечеткую
или слишком далеко друг от друга, то они не дей-
модель активации гена hb фактором Bcd (через
ствуют кооперативно.
проксимальный элемент) с моделью Хилла для
активации.
3) Энхансер включает в себя несколько сайтов
связывания для других активаторов, и эти актива-
С целью реализовать НЛМ обсуждаемых в ста-
торы имеют тенденцию действовать кооператив-
тье энхансеров мы выполняем анализ граммати-
но с Bcd.
ческих правил их конструирования из сайтов свя-
БИОФИЗИКА том 66
№ 1
2021
НЕЧЕТКОЕ МОДЕЛИРОВАНИЕ РЕГУЛЯЦИИ ГЕНОВ
59
зывания активаторов и репрессоров (см. рис. 1). В
казывают, что во второй половине четырнадцато-
итоге мы пришли к таким заключениям.
го цикла эмпирическая зависимость экспрессии
hb от концентрации Bcd достигает nk = 12 (как ил-
Во-первых, ядро энхансера состоит из класте-
люстрируется графиком на рис. 4а), что не может
ра сайтов связывания фактора Bcd. Эти сайты,
быть объяснено кооперативностью действия Bcd
как правило, разделяются расстояниями, равны-
ми или пропорциональными шагу спирали ДНК
[48]. В качестве альтернативных гипотез резонно
рассматривать (кооперативное) действие других
(10-11 п.н.) [44, 45, 80]]. Это то, что называют пе-
активаторов (см., например, работу [81]). Еще од-
риодичностью в распределении сайтов связыва-
но популярное альтернативное объяснение - это
ния на ДНК энхансера. Именно такое располо-
самоактивация гена hb кодируемым им фактором
жение делает возможным кооперативную актива-
Hb [48, 75, 82].
цию таких энхансеров
[80]. Специфическое
связывание с матрицей ДНК первой молекулы
Наконец, еще одно возможное объяснение -
Bcd облегчает связывание второй, эти две связан-
что высокая крутизна склона домена экспрессии
ные молекулы облегчают связывание третьей мо-
hb достигается не кооперативной активацией, а
лекулы Bcd и так далее. Такие процессы опреде-
является результатом кооперативной репрессии
ляют как кооперативное связывание с матрицей
также крутых склонов соседствующих доменов
ДНК (template binding cooperativity). Более того,
репрессоров (прежде всего Kr и Kni) [47, 83, 84].
анализируемый нами класс энхансеров включает
Вторая проблема состоит в том, что до сих пор
сайты связывания для некоторых других актива-
неясно, как взаимодействуют обсуждаемые эле-
торов, и эти сайты также имеют тенденцию рас-
менты с еще одним ЦРМ этого гена - энхансером
полагаться относительно сайтов Bcd согласно
полос (будет рассмотрен в следующем разделе).
правилу периодичности [45, 80].
Здесь следует отметить, что склон переднего
Во-вторых, энхансер включает в себя несколь-
домена экспрессии hb является, в свою очередь,
ко сайтов связывания для «близкодействующих»
вторичным морфогенетическим градиентом ре-
репрессоров, действующих посредством квен-
прессивного действия и, как полагают, контроли-
чинга (quenching), и каждый сайт активации дол-
рует позиции границ доменов минимум восьми
жен находиться поблизости (> 100 п.н.) от такого
генов-мишеней [44]. Поэтому исчерпывающие
сайта репрессора. Типичный энхансер имеет тен-
знания и строгие заключения о природе и меха-
денцию отвечать на связывание репрессора с его
низмах контроля крутизны и точности позиции
сайтом (в этом энхансере) неаддитивно. (Полага-
этого склона экспрессии hb имеют существенное
ют, что при этом задействуется вся молекулярная
биологическое значение.
машина транскрипции, включающая общие и
Соответственно, в этом разделе мы рассмот-
специфические факторы.) Такую неаддитивность
рим средствами НЛМ эти проблемы гена hb на
(в терминах химической кинетики) описывают
примере проксимального элемента.
как кооперативную или синергетическую репрес-
Кооперативная активация фактором Bcd. Мы
сию. У нас есть основания (по аналогии с коопе-
начнем наш анализ с простейших нечетких моде-
ративными активаторами) связать эту коопера-
лей действия единственного активатора на ген-
тивность с кластеризацией сайтов связывания
мишень (имея в виду кооперативное действие Bcd
этих ТФ.
на ген hb). Мы зададим уровни экспрессии и эф-
Модель проксимального элемента ген hb. Два
фектора (активатора e) и гена-мишени (t) тремя
весьма схожих по характеристикам и функциям
одинаковыми треугольными термами каждый
элемента гена hb - проксимальный и теневой эн-
(см. раздел «Наш подход»). Сила активатора (т.е.
хансеры - относятся к одним из самых изучен-
нелинейность, кооперативность его действия на
ных, как обсуждалось выше. Принято считать,
мишень) заданы тремя возможными вариантами
что первичная функция проксимального элемен-
таблицы нечетких правил, как показано на рис. 5
та — это обеспечивать активацию гена hb морфо-
для слабого, умеренного и сильного активатора.
геном Bcd. Тем не менее обширные исследования
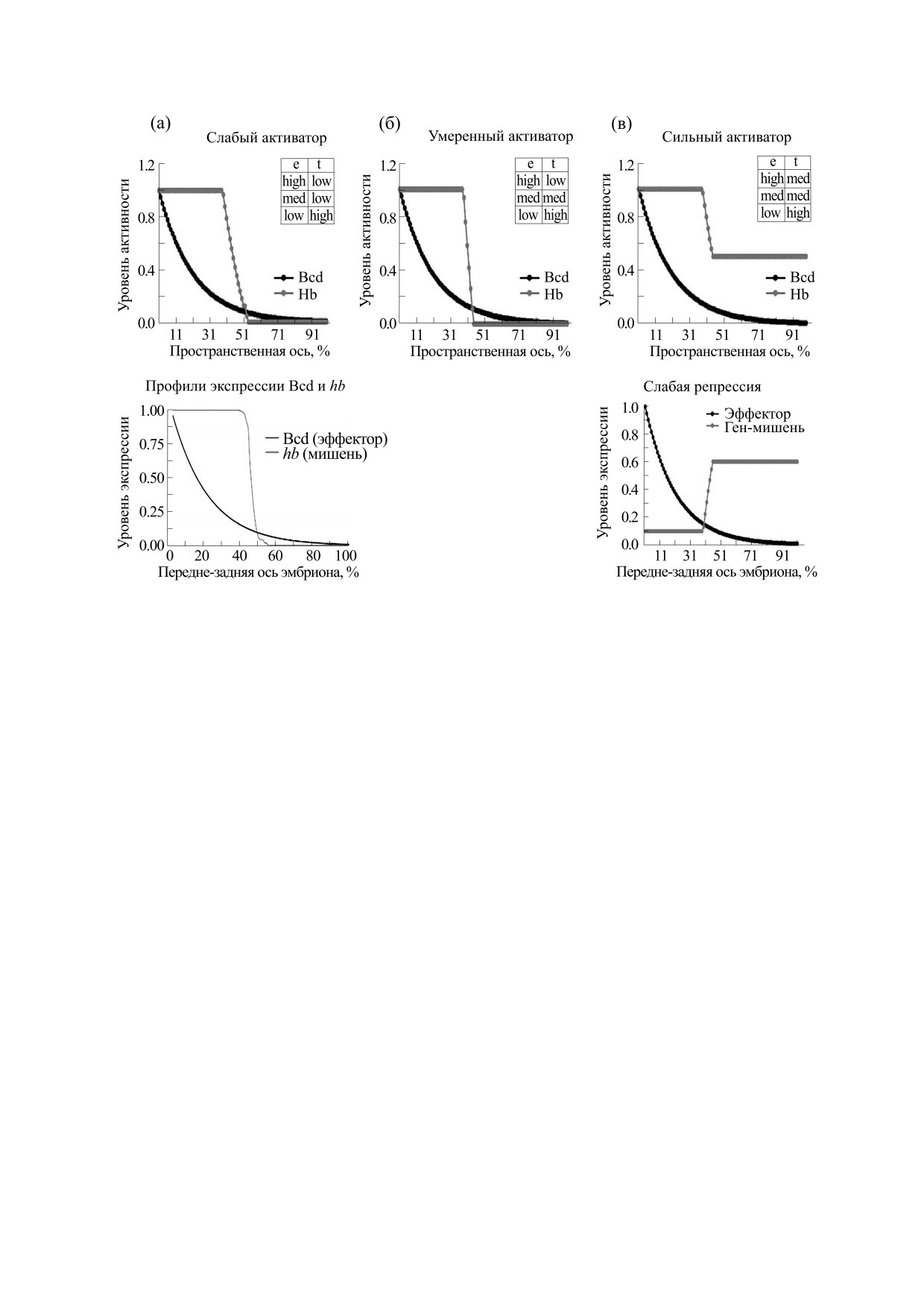
Рис. 2а демонстрирует, как выглядят в дина-
до сих пор не принесли ясности и однозначности
мике профили экспрессии hb вместе с профилем
в сложную картину экспериментальных наблюде-
Bcd. Сравнение результатов простой нечеткой
ний и выводов. Первая проблема заключается в
модели и реальных данных приводит к заключе-
том, что крутизна границы домена гена-мишени
нию, что модели слабой и средней (но не силь-
(hb) во второй половине четырнадцатого цикла
ной) активации ближе всего к эксперименталь-
слишком велика (см. рис. 2а), чтобы ее можно
ным данным. Более детальный анализ с большим
было объяснить только кооперативным действи-
числом термов и более детальными таблицами
ем Bcd [47, 48, 70-72]. В биохимических экспери-
правил показывает, что такая модель слишком
ментах показано, что кооперативность действия
проста (сверхупрощение) и требуются более де-
Bcd соответствует коэффициенту Хилла nk = 3
тальные нечеткие модели. При этом модель мож-
[68, 70], тогда как эмпирические наблюдения по-
но развивать просто увеличением числа одинако-
БИОФИЗИКА том 66
№ 1
2021
60
МАКАШОВ и др.
Рис. 5. Зависимость отклика (уровня экспрессии) простейшей нечеткой модели проксимального энхансера гена hb
(здесь t - ген-мишень) от меры кооперативности эффектора-активатора (определяемой таблицей нечетких правил).
Фактор-эффектором (е) выступает Bcd. Таблицы зависимости t от e соответствуют слабой (а), умеренной (б) и
сильной (в) модели активации (и для эффектора и для мишени задано по три уровня - high, med, low). Модель
сопоставляется с экспериментальными данными: на панели (а) внизу приведены эмпирические профили экспрессии
фактора Bcd и его мишени гена hb для эмбриона дрозофилы на стадии середины четырнадцатого цикла (по
материалам веб-ресурса FlyEx [91]). Можно заключить, что эмпирический профиль hb схож и с моделью (а), и с
моделью (б), но не с моделью (в). На панели (в) внизу приведен вид зависимости экспрессии гена-мишени при
альтернативной модели, когда эффектор является не активатором, а репрессором. Это соответствует таблице правил,
отличающейся от таковой на (в) тем, что столбец t перевернут (сверху вниз: high, med, med).
вых термов и тщательно подобранных бóльших
добранной так, чтобы склон домена экспрессии
по размеру таблиц правил. А можно использовать
находился (примерно) на одном месте при тести-
относительно небольшое (пять-шесть) число тер-
руемых значениях nk. Использовали шесть тер-
мов и для эффектора, и для мишени и простую
мов для эффектора и для мишени (см. рис. 6).
таблицу правил. Но при этом нужно подбирать
размеры каждого терма индивидуально (вручную
Вместе с нечеткой моделью Хилла в координа-
или автоматизированно, как рассматривается в
тах «доза-эффект» («активатор-мишень») ре-
разделе «Наш подход»). Мы ниже воспользуемся
зультаты представлены также в виде профилей в
последним подходом к НЛМ.
зависимости от кривой активатора, экспоненци-
Рассмотрим теперь нечеткое моделирование
альной вдоль пространственной оси (т. е. про-
кооперативного действия активатора на ген-ми-
филь активатора аппроксимировали отрезком
шень в рамках модели Хилла и с неодинаковыми
экспоненты). Это позволяло сравнить эмпириче-
термами. Это проиллюстрирует принцип разви-
ские профили паттернов экспрессии гена hb
ваемого подхода и позволит сравнить эмпириче-
вдоль главной (передне-задней) оси раннего эм-
ские профили гена-мишени (hb) с результатами
бриона дрозофилы. На рис. 6 модельные профи-
нечеткого моделирования такой моделью.
ли сопоставляются с экспериментальным профи-
Нечеткое моделирование функции Хилла вы-
лем гена hb на той самой ранней стадии четырна-
полняли, как описано в разделе «Наш подход».
дцатого цикла, когда, как полагают (см. работу
Моделирование выполняли для серии величин nk
[48]), экспрессия гена-мишени hb находится под
(= 1, 2, 3, 4) и константой «диссоциации» (Kd) по-
преимущественным контролем Bcd.
БИОФИЗИКА том 66
№ 1
2021
НЕЧЕТКОЕ МОДЕЛИРОВАНИЕ РЕГУЛЯЦИИ ГЕНОВ
61
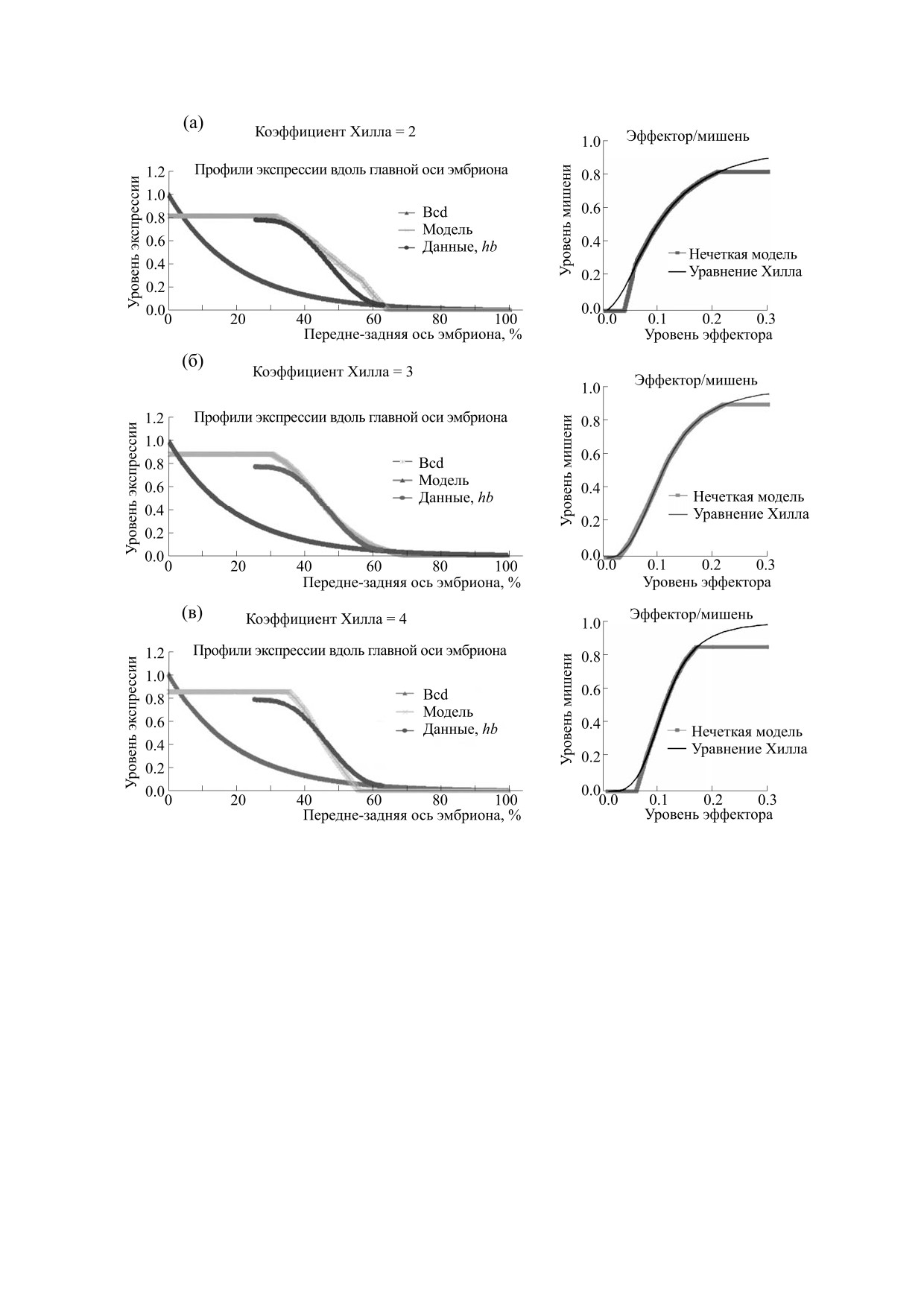
Рис. 6. Нечеткое моделирование действия активатора Bcd на проксимальный элемент гена hb демонстрирует, что
активация гена hb морфогеном Bcd в начале четырнадцатого цикла может быть аппроксимирована законом Хилла при
nk = 3. (а)-(в) - Модель профиля экспрессии гена-мишени вдоль передне-задней оси эмбриона под контролем
экспоненциального пространственного градиента Bcd при разных уровнях кооперативности действия фактора Bcd
(величина коэффициента Хилла nk =2, 3, 4) и соответствующая кривая зависимости уровня экспрессии гена-мишени
от уровня активатора (на врезках справа). Теоретические модели профиля экспрессии гена-мишени сопоставляются
с экспериментальным профилем гена hb в начале четырнадцатого цикла.
Рис. 6 демонстрирует, что эмпирический про-
тырнадцатого цикла. Это сопоставимо с заключе-
филь экспрессии гена hb (самое начало четырна-
ниями других авторов [68, 69]. Вместе с тем суще-
дцатого цикла) хорошо аппроксимируется нечет-
ственный рост крутизны границы домена hb в хо-
кой моделью функции Хилла с параметрами
де четырнадцатого цикла требует альтернативных
nk = 3 и Kd = 0.0013 (рис. 6б). На основании наших
гипотез для своего объяснения.
результатов мы приходим к заключению, что про-
стой гипотезы кооперативного действия ТФ Bcd
Кооперативная активация фактором Hbmat. Как
на проксимальный энхансер достаточно, чтобы
отмечалось выше, основная проблема с количе-
объяснить геометрию профиля Hb в начале че-
ственным моделированием динамики профиля
БИОФИЗИКА том 66
№ 1
2021
62
МАКАШОВ и др.
Hb заключается в том, что крутизна границы до-
мишень особенно нагляден, поскольку набор
мена экспрессии hb (т.е. именно то, что мы здесь
правил представим таблицей (раздел «Наш под-
моделируем) очень быстро, в ходе четырнадцато-
ход», табл. 1-3).
го цикла, нарастает и становится столь крутой,
В общем же совокупное действие может вы-
что в рамках модели Хилла это соответствует
числяться из отдельных акций (активации и ре-
nk = 12. Столь высокая кооперативность актива-
прессии) согласно операторам AND, OR или
ции сомнительна из общих соображений кинети-
MEAN (см. работу [41]).
ки и молекулярных механизмов действия актива-
В развитие вышеприведенных моделей нам ре-
тора Bcd. Тщательные количественные исследо-
зонно рассмотреть НЛМ с двумя активаторами,
вания кооперативности кинетики активации
Bcd и Hbmat. Bcd мы моделируем двумя или тремя
кластеров сайтов связывания Bcd в биохимиче-
термами (как фактор А2 в табл. 1 и 2), тогда как
ских экспериментах также ставят под сомнение
столь высокую кооперативность.
Hbmat - тремя термами (фактор А1). Результат ра-
боты такой НЛМ приведен на рис. 7в. Наше моде-
Возможное и широко обсуждаемое объясне-
лирование в рамках табл. 1 и 2 дало сходные ре-
ние состоит в том, что проксимальный элемент
зультаты.
интегрирует активирующее действие не только от
Таким образом, (неаддитивное) действие пары
Bcd, но и от других ТФ-активаторов (например
Hb [81]). Прежде всего, обращает на себя внима-
активаторов (Bcd и Hbmat) более реалистично мо-
ние наличие в проксимальном элементе как
делирует активацию гена мишени. Поэтому мы
предполагаемых, так и экспериментально под-
можем предположить, что коактивация этих двух
твержденных сайтов связывания фактора Hb.
факторов существенна для объяснения крутизны
Вместе с тем, как хорошо известно, фактор Hb
профиля hb в более позднем четырнадцатом
поставляется в яйцеклетку-зиготы материнским
цикле.
организмом еще до активации эмбрионального
Самоактивация гена. Наконец, рассмотрим по-
гена hb [85], так что он формирует домен в перед-
следнюю из простых и распространенных альтер-
ней части раннего эмбриона с пологой границей
нативных объяснений динамики склона домена
(напоминая более поздний домен эмбрионально-
экспрессии hb. А именно, исследуем поведение
го Hb). Важность этого материнского домена в
такой НЛМ, в которой исходный, начальный
предопределении раннего развития эмбриона хо-
профиль экспрессии Hb (в результате экспрессии
рошо известна [86]. Поэтому мы выполнили здесь
гена hb), полученный, например, кооперативной
альтернативное нечеткое моделирование, приняв
активацией фактором Hbmat (nk = 3), на втором
в качестве активатора материнский Hbmat.
шаге моделирования сам становится активатором
Результаты НЛМ для кооперативной актива-
для экспрессии своего гена, как на рис. 7г. В
ции фактором Hbmat (с коэффициентами Хилла
представленной серии тестов мы считаем меха-
nk = 2, 3, 4) приведены на рис. 7а. Рисунок нагляд-
низм самоактивации кооперативным (nk = 3). В
результате такой серии шагов самоактивации
но иллюстрирует, что действие пространственно
более крутого (чем экспонента фактора Bcd) ак-
профиль экспрессии очень быстро становится
тиватора может объяснить бóльшую крутизну
очень крутым.
склона домена экспрессии зиготного гена hb. Там
Следовательно, в результате тестов с нашими
же приведены реальный профиль экспрессии ге-
НЛМ мы приходим к заключению, что (коопера-
на hb для середины четырнадцатого цикла. Вид-
тивная) самоактивация гена фактором, который
но, что кооперативное действие Hbmat может ча-
этот ген кодирует, - это самое простое объясне-
стично объяснить наблюдаемую более высокую
ние крутизны домена экспрессии гена hb. Эти ре-
крутизну склона hb (чем модель для Bcd, см.
зультаты согласуются с заключениями некоторых
рис. 7б). (Заметим также, что больший, чем в эм-
предыдущих публикаций [48, 75, 82]. Тем не ме-
пирических данных, сдвиг границы домена экс-
нее проблема еще далека от своего окончательно-
прессии hb в модели объясним отсутствием дегра-
го разрешения, поскольку самоактивация гена hb
дации в нашей простой НЛМ.) Однако крайне
не исследована во всех деталях и у исследователей
высокая и все нарастающая крутизна склона до-
нет единого мнения по этому вопросу (см. работу
мена экспрессии hb во второй половине четырна-
[47]).
дцатого цикла по-прежнему нуждается в дальней-
Возможное альтернативное объяснение суще-
шем исследовании.
ственного нарастания крутизны границы домена -
это действие элемента полос, рассмотренное ниже.
Коактивация двумя факторами. В развитие под-
хода к НЛМ генно-регуляторных элементов не-
обходимо уметь включать в модель действие на
Модель элемента полос гена hb. Опубликован-
данный элемент нескольких регулирующих ТФ.
ные результаты анализа активности элемента по-
Случай нечеткого действия двух эффекторов на
лос свидетельствуют, что его функция - это ре-
БИОФИЗИКА том 66
№ 1
2021
НЕЧЕТКОЕ МОДЕЛИРОВАНИЕ РЕГУЛЯЦИИ ГЕНОВ
63
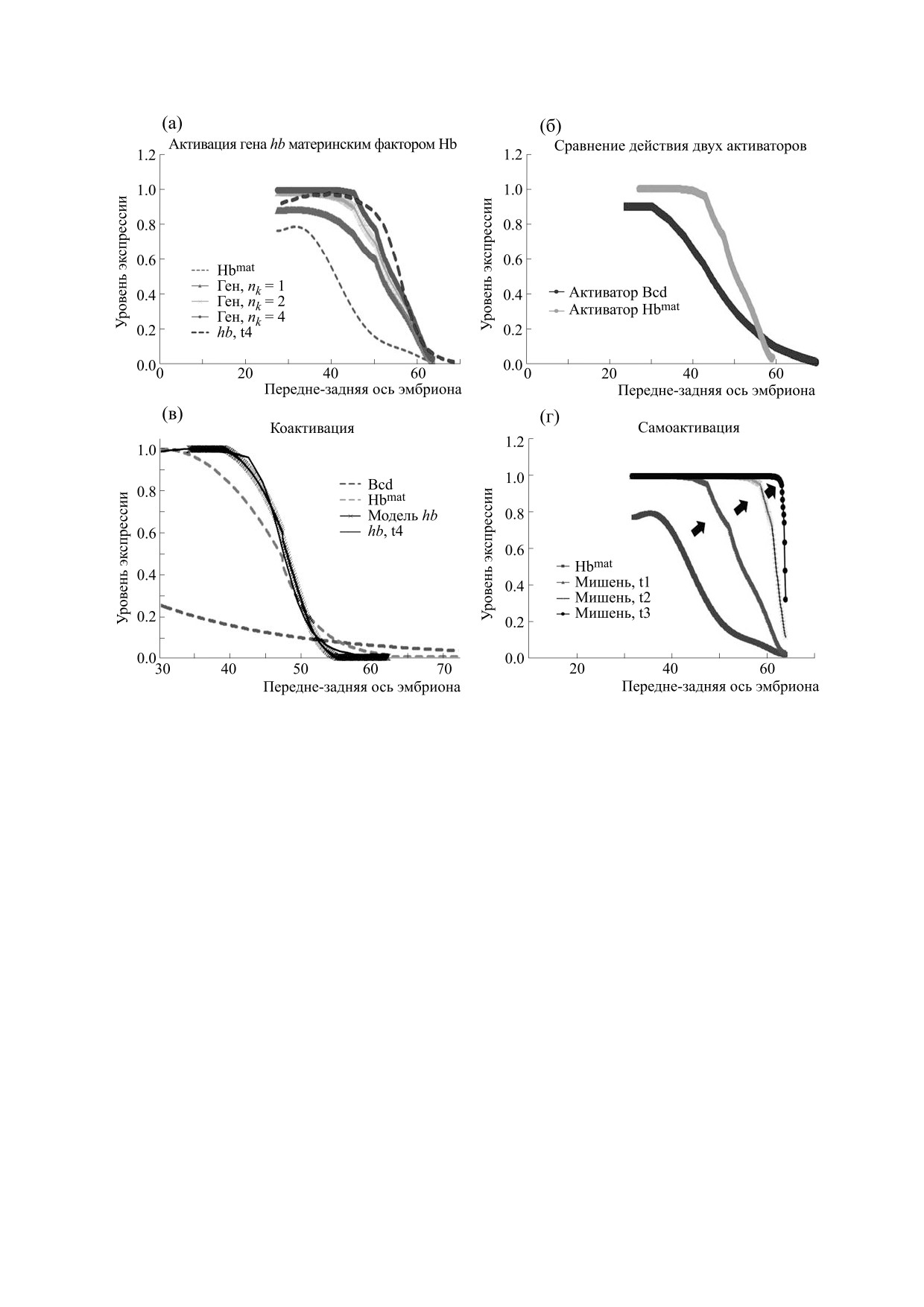
Рис. 7. Возможные более общие случаи механизмов действия проксимального энхансера гена hb. (а) и (б) - Нечетко-
логическое моделирование действия активатора Hbmat в рамках модели Хилла с коэффициентами Хилла nk = 2, 3, 4.
(а) - Профили экспрессии гена hb как результат действия активатора Hbmat в рамках модели Хилла с nk = 2, 3, 4 в
сравнении с эмпирическим профилем экспрессии hb в середине четырнадцатого цикла (t4). (б) - Сопоставление
результата нечеткого моделирования отдельно для активатора Bcd и для активатора Hbmat (при одном и том же
значении коэффициента Хилла nk = 3). Видно, что действие материнского Hb способно объяснить бóльшую крутизну
склона домена экспрессии гена hb. (в) - Нечеткое моделирование совместного действия пары активаторов
(коактивация) Bcd и Hbmat. Также приведен профиль экспрессии в середине четырнадцатого цикла (t4). Видно, что
совместное действие двух активаторов в рамках нечеткой модели дает более крутой склон домена экспрессии гена hb,
чем действие каждого из факторов по отдельности. (г) - Динамика профиля экспрессии гена hb для случая нечеткой
модели самоактивации. Исходный профиль материнского фактора Hbmat действует как активатор на зиготный ген hb,
давая профиль экспрессии hb к моменту t1 (мишень, t4). Далее, с момента t1 этот профиль зиготного фактора Hb
действует как активатор (само-активация), давая, в свою очередь, профиль экспрессии hb к моменту t2 (мишень, t2).
Наконец, с момента t2 этот профиль зиготного фактора Hb действует как активатор, давая в итоге очень крутой
профиль экспрессии hb к моменту t3 (мишень, t3).
прессирование гена-мишени на всем протяже-
активация фактором Bcd» и на рис. 5. Рис. 5в(1)
нии главной оси раннего эмбриона, за
иллюстрирует то общее соображение, что для
исключением двух небольших областей, как на
крутых склонов доменов экспрессии требуется
рис. 2б [47]. Это и дает в итоге два четких пика
как активация, так и репрессия по неаддитивно-
экспрессии.
му (кооперативному) механизму.
Мы начнем наш анализ кооперативной ре-
Продолжим изучение средствами НЛМ коопе-
прессии с общих результатов, аналогично тако-
ративного действия репрессоров на ген-мишень в
вым, описанным в параграфе «Кооперативная
рамках модели Хилла с достаточным числом тер-
БИОФИЗИКА том 66
№ 1
2021
64
МАКАШОВ и др.
мов и разными уровнями кооперативности (с раз-
преимущественным контролем
«S-образного»
личными значениями коэффициента nk). Мы со-
профиля фактора-репрессора Gt [88]. Результаты
средоточимся на анализе поведения домена PS4
нашего нечеткого моделирования контроля эф-
фектором Gt переднего склона пика PS4 в сопо-
(антериорный пик, Hbant, см рис. 2б). Этот домен
ставлении с профилем модели Хилла с nk = -3, -4
представляет собой четкий и крутой пик локаль-
демонстрируют, что эмпирическая кривая весьма
ной экспрессии. Его задний склон, как полагают,
сходна с результатами нечеткого моделирования
находится под преимущественным контролем «S-
(не показано). Отметим здесь, что знак действия
образного» профиля фактора Kr (а также фактора
Gt на ген hb (активатор или репрессор) все еще
Kni) [50, 87]. Передний склон домена PS4, как по-
обсуждается [64]. В качестве альтернативы неко-
лагают, находится под преимущественным кон-
торые авторы рассматривают механизм саморе-
тролем профиля фактора-репрессора Gt [88].
прессии hb через энхансер полос [47]. Поэтому
Есть также экспериментальные наблюдения, что
вопрос нуждается в дальнейшем эксперимен-
этот склон находится под контролем собственно
тальном и теоретическом анализе.
фактора Hb, кодируемого геном hb [47], что труд-
нее представить с точки зрения кинетики.
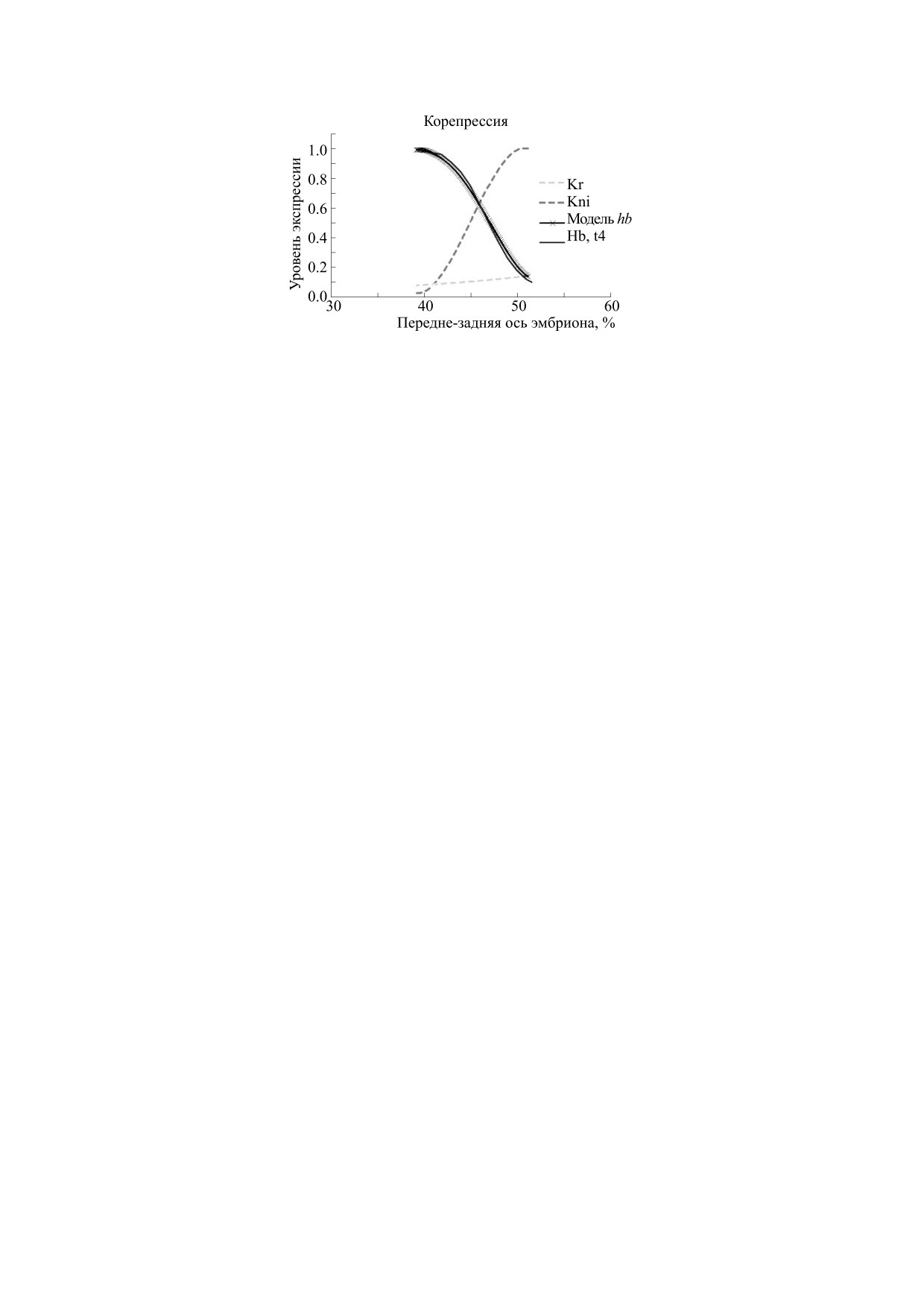
Корепрессия двумя факторами. Альтернативная
гипотеза состоит в том, что сайты связывания Kr
Сначала рассмотрим результаты моделирова-
оказывают репрессивное действие кооперативно
ния кооперативности репрессии гена-мишени
с сайтами связывания другого репрессора - Kni,
репрессором Kr, исходя из простейшей гипотезы,
которых охарактеризовано тоже три, так что по-
что именно профиль Kr отвечает за геометрию
лучается плотный маленький кластер из трех сай-
профиля пика экспрессии hb (его постериорный
тов связывания в одной половине энхансера и
склон), как иллюстрируется на рис. 8а-в. Моде-
трех - в другой (рис. 1в). В целом этот вопрос тре-
лирование выполняли для ряда величин nk (= -1,
бует дальнейших исследований.
-2, -3, -4) и константой «диссоциации» подо-
С целью проанализировать возможности ко-
бранной так, чтобы склон домена экспрессии на-
оперативной репрессии двумя факторами (Kr и
ходился (примерно) на одном месте при тестиру-
Kni) мы выполнили соответствующее нечеткое
емых значениях nk. Профили репрессоров для
моделирование, как объяснено в разделе «Наш
ранней экспрессии генов сегментации естествен-
подход», табл. 3. Результаты приведены на рис. 9.
но аппроксимировали отрезком S-образной (сиг-
Видно, что совместная кооперативность двух ре-
моидной) функции (см. рис. 2). Использовано
прессоров объясняет крутизну профиля домена.
пять или шесть термов для эффектора и для ми-
Так что такая модель лучше подходит для интер-
шени (рис. 8).
претации грамматики элемента полос.
Результаты численного решения уравнения
На основании наших результатов мы прихо-
Хилла (относительно участка сигмоидного про-
дим к заключению, что простой гипотезы коопе-
филя репрессора) сопоставляются с результатами
ративного действия эффекторов-репрессоров на
нечеткого моделирования и с эмпирическим про-
энхансер полос достаточно, чтобы объяснить гео-
филем экспрессии гена hb (на уровне мРНК) в се-
метрию склонов антериорного (PS4) пика hb в че-
редине четырнадцатого цикла, как представлено
тырнадцатом цикле. При этом каждый склон пи-
на рис. 8а-в. Рис. 8 демонстрирует, что эмпири-
ка PS4 независимо контролируется своим ре-
ческий профиль экспрессии hb примерно соот-
прессором (или репрессорами). Примечательно,
ветствует решению Хилла со значением nk между
что кооперативность репрессии достаточно высо-
-3 и -4.
ка и сравнима с кооперативностью активации
Найденная нами существенная кооператив-
этого гена в том же четырнадцатом цикле. Мы не
ность действия Kr на hb находится в некотором
знаем публикаций, где бы обсуждалась коопера-
несогласии с известным количеством и распреде-
тивность репрессии для генов сегментации.
лением экспериментально подтвержденных сай-
Аддитивность действия регуляторных элементов
тов связывания для Kr на энхансере полос (пара в
данного гена. В заключение отметим, что при ана-
одной половине энхансера и всего один - в дру-
лизе генной регуляции для распространенных
гой, как на рис. 1в). В этой связи можно предло-
случаев нескольких (автономных и полуавтоном-
жить, что не все сайты для Kr найдены. Здесь су-
ных) регуляторных элементов возникает общая
щественно, что биоинформационный анализ де-
проблема: как интегрируется действие этих эле-
монстрирует наличие большего числа сайтов для
ментов, если они активны одновременно (и в
Kr, расположенных плотными кластерами, как в
данном клеточном массиве).
случае теневого элемента (см. рис. 1б). Альтерна-
Недавние исследования, проведенные, в част-
тивная модель кооперативной корепрессии рас-
ности, и на рассматриваемых здесь регуляторных
смотрена в следующем подразделе.
элементах, показали целый спектр суммирования
Далее, аналогичным образом, антериорный
действия двух элементов одного гена: суперадди-
склон домена PS4, как полагают, находится под
тивность, аддитивность и субаддитивность [89].
БИОФИЗИКА том 66
№ 1
2021
НЕЧЕТКОЕ МОДЕЛИРОВАНИЕ РЕГУЛЯЦИИ ГЕНОВ
65
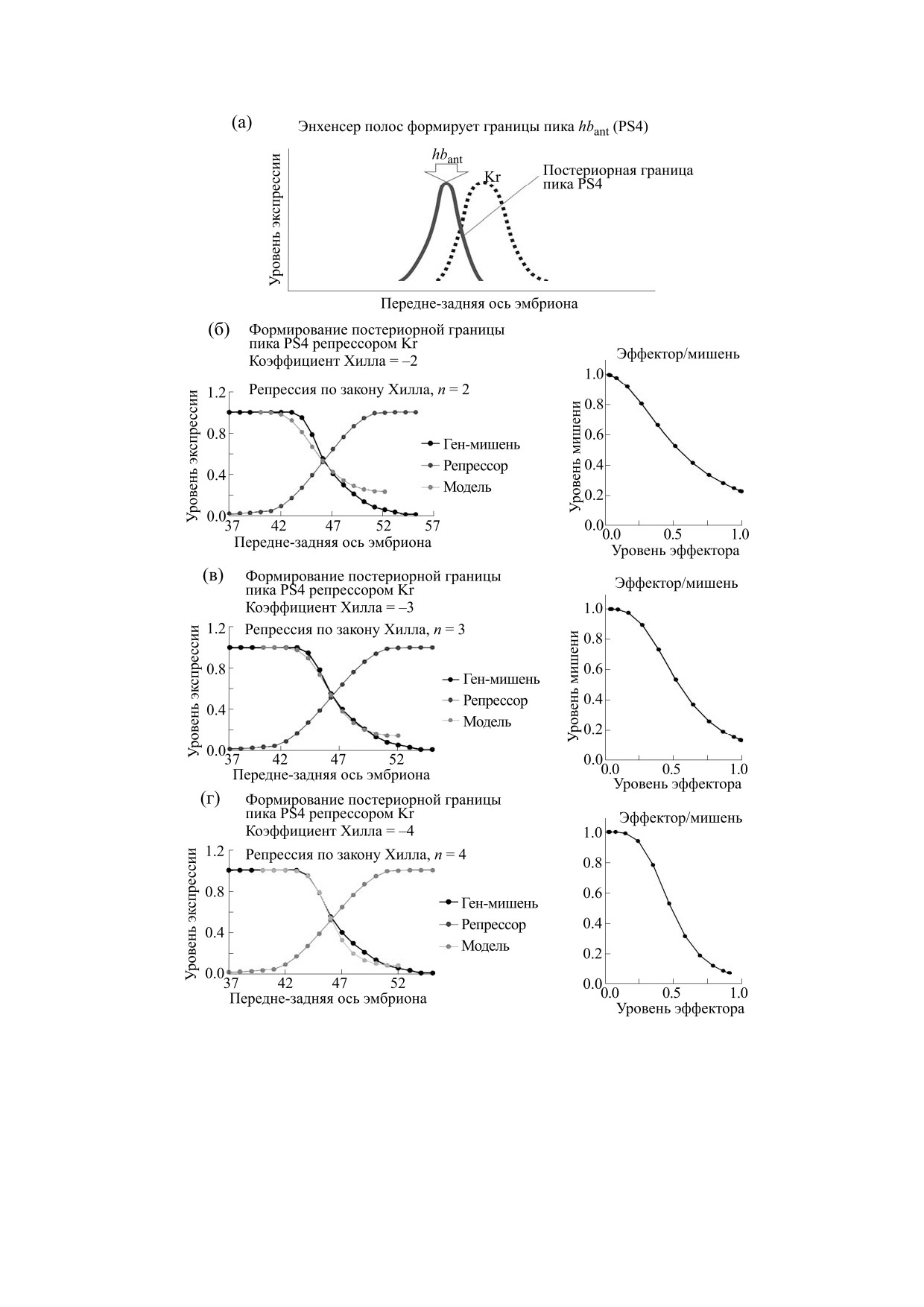
Рис. 8. Репрессия гена hb фактором Kr через элемент полос подчиняется закону Хилла. (а) - Крутой склон домена
экспрессии репрессора Kr формирует постериорную границу пика PS4 (Hbant). (б)-(г) - Модель профиля экспрессии
гена-мишени вдоль участка передне-задней оси эмбриона под контролем «S-образного» профиля репрессора Kr при
трех разных уровнях кооперативности (величина коэффициента Хилла, nk = -2, -3, -4) и соответствующая кривая
зависимости уровня экспрессии гена-мишени hb от уровня репрессора Kr (на врезках справа). Теоретические модели
профиля экспрессии гена-мишени сопоставляются с экспериментальным профилем гена hb в середине
четырнадцатого цикла.
БИОФИЗИКА том 66
№ 1
2021
66
МАКАШОВ и др.
Рис. 9. Нечеткое моделирование действия пары репрессоров (Kr и Kni, корепрессия) через элемент полос на крутизну
постериорного склона пика PS4. Совместное действие двух репрессоров в рамках нечеткой модели дает более крутой
склон домена экспрессии гена hb, чем действие каждого из факторов по отдельности (сравни с рис. 7в).
Исходя из простой гипотезы аддитивности
гуляции генов, вовлеченных в процессы сегмен-
действия элемента полос и проксимального эле-
тации.
мента, мы наблюдаем, что суммарная картина
экспрессии гена-мишени hb хорошо аппрокси-
ВЫВОДЫ
мирует эмпирические профили мРНК hb к сере-
дине четырнадцатого цикла, как показано на
Нами впервые нечеткое моделирование при-
рис. 10.
меняется для разработки моделей активности
цис-регуляторных модулей и впервые применя-
Полученные здесь результаты являются суще-
ется к проблемам экспрессии генов биохимиче-
ственным дополнением к нашим результатам де-
ской разметки ранних эмбрионов. Подход пред-
тального детерминистического моделирования
ставлен в приложении к анализу конкретной про-
экспрессии гена hb (системой детальных уравне-
блемы перехода от самых ранних механизмов
ний в частных производных) [51], где акцент был
паттерна эмбриональной сегментации к более
сделан на наличии двух промотеров у этого гена.
поздним. По такому сценарию охарактеризована
Для получения исчерпывающей картины деталей
экспрессия целого ряда генов эмбрионального
динамики этого гена требуется свести воедино
паттерна, но наиболее изучена она для гена hb.
оба подхода.
Сценарий заключается в том, что самый ранний
Сходно организованные гены. Тщательный ана-
паттерн формируется как активация гена-мише-
лиз результатов исследования поведения ЦРМ
ни (например, ранний hb) морфогенетическими
градиентами (прежде всего, Bcd) в форме широ-
генов сегментации эмбриона дрозофилы показы-
вает, что схема динамики паттерна, анализируе-
ких доменов экспрессии. На этом этапе действу-
мая здесь на примере hb, может быть характерна
ют свои, ранние механизмы контроля границы
для целого ряда генов. А именно, имеются ЦРМ,
экспрессии (типично - кооперативная активация
которые контролируют формирование достаточ-
гена-мишени морфогеном). Примечательно, что
эти процессы находятся под контролем соответ-
но обширных доменов экспрессии (аналогично
проксимальному элементу гена hb). И имеются
ствующих ранних полуавтономных регуляторных
ЦРМ, которые контролируют формирование бо-
элементов (например, проксимальный элемент
лее коротких острых и четких пиков экспрессии в
hb). На следующем этапе (на фоне этих широких
пределах начального широкого домена, как это
доменов) быстро формируются четкие и узкие
полосы (пики) экспрессии. Они контролируются
имеет место в случае элемента полос гена hb.
Прежде всего это относится к таким генам семей-
своими регуляторными механизмами (например,
ства gap (к которым относится и hb), как gt, Kr и
контролем соседствующими доменами факто-
tailless. Эти ЦРМ были исследованы в ряде публи-
ров-репрессоров). Эти процессы регулируются
каций [55-57, 90]. Их функциональная близость к
своими полуавтономными регуляторными эле-
ментами (например, элемент полос гена hb). Ре-
hb делает такие наблюдения по деталям их регуля-
ции (сходство с hb), в достаточной мере ожидае-
зультирующий паттерн экспрессии анализируе-
мым. Все эти факты, вместе взятые, свидетель-
мых генов высоко динамичен и определяется на-
ствуют о том, что моделируемые в этой статье де-
ложением двух процессов контроля паттерна
тали динамики экспрессии под контролем пар
экспрессии.
ЦРМ с весьма различающейся стратегией кон-
С помощью подхода НЛМ проанализирована
троля паттерна могут быть общим принципом ре-
регуляция проксимальным энхансером крутизны
БИОФИЗИКА том 66
№ 1
2021
НЕЧЕТКОЕ МОДЕЛИРОВАНИЕ РЕГУЛЯЦИИ ГЕНОВ
67
Рис. 10. Аддитивность действия двух основных регуляторных элементов гена hb в четырнадцатом цикле. Сумма
профилей нечетких моделей действия проксимального элемента и элемента полос хорошо аппроксимируют
эмпирический профиль экспрессии гена hb. (а) - Экспериментально наблюдаемый профиль экспрессии гена hb в
начале четырнадцатого цикла (на уровне мРНК; график получен исходя из данных, доступных на веб-ресурсе BID
BDTNP [92]). (б) - Профиль головного домена экспрессии hb как результат кооперативной ко-активации Bcd и
другими активаторами через проксимальный элемент. (в) - Антериорный пик экспрессии hbant (домен PS4) как
результат кооперативной корепрессии пар (или большего числа) репрессоров (независимо для переднего и заднего
склона пика hbant). (г) - Суммарный профиль головной экспрессии, полученный нечетким моделированием и хорошо
аппроксимирующий экспериментальный профиль (а).
постериорного склона переднего домена экс-
высокую крутизну исследуемого склона, необъ-
прессии hb. Показано, что рост этой крутизны
яснимую только кооперативной активацией мор-
может быть объяснен кооперативной коактива-
фогеном Bcd.
цией двух активаторов (Bcd и Hbmat), а также са-
Подходом НЛМ также проанализирована ре-
моактивацией фактором Hb его гена. Это соот-
гуляция энхансером крутизны постериорного
ветствует количеству, силе и размещению сайтов
склона антериорного пика экспрессии гена hb
для ТФ этого энхансера (грамматика ЦРМ) и, по
(домен PS4). Показано, что крутизна этого скло-
крайней мере отчасти, объясняет парадоксально
на может быть объяснена (кооперативной) коре-
БИОФИЗИКА том 66
№ 1
2021
68
МАКАШОВ и др.
прессией минимум двух факторов (Kr и Kni). Это
man, and H. Iba (Wiley Interscience, 2015), pp. 240-
соответствует грамматике энхансера полос.
268.
Следует подчеркнуть, что кооперативные ме-
6. L. Sanchez and D. Thieffry, J. Theor. Biol. 211 (2), 115
(2001).
ханизмы действия ЦРМ абсолютно необходимы в
становлении эмбрионального паттерна для полу-
7. D. Thieffry and L. Sanchez, Curr. Opin. Genet. Dev.
чения резкого ответа по типу «вкл/выкл» при
13 (4), 326 (2003).
формировании резкой границы экспрессии вдоль
8. K. Kozlov, V. Gursky, I. Kulakovskiy, and M. Samson-
морфогенетических градиентов.
ova, BMC Genomics 15 (12), S6 (2014)
Мы уверены, что нечеткое лингвистическое
9. K. Kozlov, V. V. Gursky, I. V. Kulakovskiy, et al., BMC
моделирование поведения ГРС способно есте-
Genomics 16 (13), S7 (2015).
ственным образом охарактеризовать очерченные
10. V. V. Gursky, K. N. Kozlov, I. V. Kulakovskiy, et al.
признаки типичных энхансеров. Таким образом,
PloS One 12 (9), e0184657 (2017).
мы можем включить все уровни синергии энхан-
11. M. A. Gibson and E. Mjolsness, in Computational Mod-
серов: кооперативное действие пионерного фак-
eling of Genetic and Biochemical Networks, Ed. by
тора (типа Zld), кооперативное связывание ТФ с
J. M. Bower and H. Bolouri (MIT Press, Cambridge,
матрицей ДНК и синергические взаимодействия
MA, 2001), pp. 1-48.
связанных ТФ с машиной транскрипции. Мы
12. E. Myasnikova and A. Spirov, J. Bioinform. Comput.
убеждены, что модели активности генных сетей,
Biol. 16 (2), 1840008 (2018).
основанные на натуральных языках и нечеткой
логике, имеют внутреннюю функциональную ор-
13. K. Kozlov, S. Surkova, E. Myasnikova, et al., PLoS
ганизацию, более близкую к молекулярной ма-
Comput. Biol. 8 (8), e1002635 (2012).
шинерии генно-регуляторных механизмов, чем
14. С. А. Андреев, М. Г. Самсонова и В. В. Гурский,
модели в парадигме математической физики. А
Биофизика 60 (2), 225 (2015).
это должно давать нечетким моделям бóльшую
15. H. Janssens, S. Hou, J. Jaeger, et al., Nat Genet. 38
предсказательную силу.
(10), 1159 (2006).
16. N. E. Buchler, U. Gerland, and T. Hwa, Proc. Natl.
Acad. Sci. USA 100, 5136 (2003).
ИСТОЧНИКИ ФИНАНСИРОВАНИЯ
17. F. Sauer, S. K. Hansen, and R. Tjian, Science 270, 1825
Результаты раздела «Наш подход» были полу-
(1995).
чены в рамках государственного задания Мини-
18. F. Sauer, D. A. Wassarman, G. M. Rubin, and R. Tjian,
стерства образования и науки Российской Феде-
Cell 87, 1271 (1996).
рации (№ 16.8549.2017 / 8.9). Остальные результа-
19. K. Gupta, D. Sari-Ak, M. Haffke, et al., J. Mol. Biol.
ты получены в рамках гранта Российского
428 (12), 2581 (2016).
научного фонда (проект № 17-18-01536).
20. E. Nogales, R. K. Louder, and Y. He, Annu. Rev. Bio-
phys. 46, 59 (2017).
КОНФЛИКТ ИНТЕРЕСОВ
21. E. H. Mamdani and S. Assilian, Int. J. Man-Machine
Автор заявляет об отсутствии конфликта инте-
Studies 7 (1), 1 (1975).
ресов.
22. M. Sugeno, Industrial applications of fuzzy control (Else-
vier Science Pub. Co, 1985).
23. S. Istrail and E. H. Davidson, Proc. Natl. Acad. Sci.
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
USA 102, 4954 (2005).
Настоящая работа не содержит описания ис-
24. R. D. Zeigler, J. Gertz, and B. A. Cohen, BMC Bioin-
следований с использованием людей и животных
formatics 8, 272 (2007).
в качестве объектов.
25. C.-P. Lee, Y. Leu, W.-N. Yang, Applied Soft Comput-
ing 12 (3), 1115 (2012).
СПИСОК ЛИТЕРАТУРЫ
26. J. Bordon, M. Moskon, N. Zimic, and M. Miha,
IEEE/ACM Trans. Comput. Biol. Bioinform. 12 (5),
1. J. Linde, S. Schulze, S. G. Henkel, and R. Guthke,
1199 (2015).
EXCLI J. 14, 346 (2015).
27. B. H. Wang, J. W. Li, and J. S. Lim, Genetics Mol. Res.
2. H. de Jong, J. Comput. Biol. 9 (1) 67 (2002).
15 (3), gmr.15039002 (2016).
3. J. Jaeger, Manu, and J. Reinitz, Curr. Opin. Genet.
28. J. Liu, Y. Chi, C.Zhu, and Y. Jin, BMC Bioinformatics
Dev. 22, 533 (2012).
18 (1), 241 (2017).
4. M. A. H. Samee, B. Lim, N. Samper, et al., Cell Sys-
29. K. Wu and J. Liu, IEEE Transactions on Fuzzy Systems
tems 1 (6) 396 (2015).
25 (6) 1546 (2017).
5. A. Spirov, and D. Holloway, in Evolutionary Algorithms
30. S. Ben-Tabou de-Leon and E. H. Davidson, Dev Biol.
in Gene Regulatory Network Research, Ed. by N. No-
325, 317 (2009).
БИОФИЗИКА том 66
№ 1
2021
НЕЧЕТКОЕ МОДЕЛИРОВАНИЕ РЕГУЛЯЦИИ ГЕНОВ
69
31. B. Yan, D. Guan, C. Wang, et al., Nat. Commun. 8 (1),
59. D. Lebrecht, M. Foehr, E. Smith, et al., Proc. Natl.
1044 (2017).
Acad. Sci. USA 102 (37), 13176 (2005).
32. S. Rastegar, I. Hess, T. Dickmeis, et al., Dev Biol. 318,
60. Z. Xu, H. Chen, J. Ling, et al., Genes & Development
366 (2008).
28 (6), 608 (2014).
33. K. J. Won, A. Sandelin, T. T. Marstrand, and A. Krogh,
61. S. M. Foo, Y. Sun, B. Lim, et al., Curr. Biol. 24, 1341
Bioinformatics 24, 1669 (2008).
(2014).
34. J. Gertz, E. D. Siggia, and B. A. Cohen, Nature 457,
62. C. P. Verrijzer, and R. Tjian, Trends Biochem. Sci. 21,
215 (2009).
338 (1996).
35. L. Li and Z Wunderlich, Front. Genet. 8, 63 (2017).
63. M. V. Staller, B. J. Vincent, M. D. J. Bragdon, et al.,
Proc. Natl. Acad. Sci. USA 112 (3), 785 (2015).
36. J. O. Yáñez-Cuna, E. Z. Kvon, and A. Stark, Trends
Genet. 29, 11 (2013).
64. X.-Y. Li, and M. B. Eisen, BioRhiv
2018-07-
3010.1101/3808571 (2018).
37. J. Grice, B. Noyvert, L. Doglio, and G. Elgar, PloS
One 10 (7), e0130413 (2015).
65. A. V. Spirov, T. Bowler, and J. Reinitz, Nucl. Acids Res.
28, 337 (2000).
38. С. Т. Золян и Р. И. Жданов, Философия науки и
техники 23 (1), 88 (2018).
66. A. V. Spirov, M. Borovsky, and O.A. Spirova, Nucl. Ac-
ids Res. 30, 351 (2002).
39. S. T. Zolyan, and R. I. Zhdanov, Semiotica 2018 (225),
1 (2018). DOI: 10.1515/sem-2016-0214
67. W. Driever, G. Thoma, and C. Nüsslein-Volhard, Na-
40. X. Li, Y. Li, Y. Liu, and L. Wang, Int. J. Pattern Recog-
ture 340, 363 (1989).
nit. Artif. Intell. 31 (10), 1 (2017).
68. X. G. Ma, D. Yuan, K. Diepold, et al., Development
41. R. Küffner, T. Petri, L. Windhager, and R. Zimmer
122, 1195 (1996).
PloS One 5 (9), e12807 (2010).
69. D. S. Burz, R. Rivera-Pomar, H. Jäckle, and S. D. Ha-
42. D. Marbach, T. Schaffter, C. Mattiussi, and D. Florea-
nes, EMBO J. 17, 5998 (1998).
no, J. Comput. Biol. 16 (2), 229 (2009).
70. T. Gregor, D.W. Tank, E. W. Wieschaus, and W. Bialek,
43. D.M. Holloway, J. Reinitz, A. Spirov, and C. E. Vanar-
Cell 130, 153 (2007).
io-Alonso, Trends Genet. 18, 385 (2002).
71. F. He, Y. Wen, D. Cheung, et al., BMC Dev. Biol. 10,
44. D. Yu and S. Small, Curr. Biol. 18, 868 (2008).
80 (2010).
45. D. Papatsenko, Y. Goltsev, and M. Levine, Nucl. Acids
72. F. He, J. Ren, W. Wang, J. Ma, PLoS One 6 (4), e19122
Res. 37 (17), 5665 (2009).
(2011).
46. M. W. Perry, A. N. Boettiger, and M. Levine, Proc Nat
73. D. M. Holloway, F. J. P. Lopes, L. da Fontoura Costa,
Acad Sci USA 108, 13570 (2011).
et al., PLoS Comput. Biol. 7 (2), e1001069 (2011).
47. M. W. Perry, J. P. Bothma, R. D. Luu, and M. Levine,
74. G. Struhl, K. Struhl, and P.M. Macdonald, Cell 57 (7),
Curr. Biol. 22, 2247 (2012).
1259 (1989).
48. F. J. P. Lopes, A. V. Spirov, and P. M. Bisch, Dev. Biol.
75. F. J. P. Lopes, F. M. C. Vieira, D. M. Holloway, et al.,
370 (2), 165 (2012).
PLoS Comput. Biol. 4 (9), e1000184 (2008).
49. E. A. Zagrijchuck, M. A. Sabirov, D. M. Holloway, and
76. J. S. Margolis, M. L. Borowsky, E. Steingrímsson,
A. V. Spirov, J. Bioinform. Comput. Biol. 12, 1441009
et al., Development 121 (9), 3067 (1995).
(2014).
77. J. M. Mendel, Uncertain Rule-Based Fuzzy Logic Sys-
50. D. M. Holloway and A. V. Spirov, PloS One 10 (3),
tems: Introduction and New Directions (Prentice-Hall,
e0118450 (2015).
Upper Saddle River, NJ, 2001).
78. Ю. И. Митюшин, Б. И. Мокин и А. П. Ротштейн,
51. A. V. Spirov, E. M. Myasnikova, and D. M. Holloway,
Soft Computing: идентификация закономерностей
J. Bioinform. Comput. Biol. 14 (2), 1 (2016).
нечеткими базами знаний (Унiверсум-Вiнниця,
52. T. Takagi and M. Sugeno, IEEE Trans. System Man
2002).
Cybernet. 15, 116 (1985).
79. L. Windhager, Ph.D. Dis. (der Fakultat fur Mathema-
53. M. Storace and O. De Feo, IEEE Trans. Circuits and
tik, Informatik und Statistik, der Ludwig-Maximilians-
Systems I: Regular Papers 51 (4), 830 (2004).
Universitat, Munchen, 2013).
54. Y. Park, K. M. Shaw, H. J. Chiel, and P. J. Thomas,
80. V. J. Makeev, A. P. Lifanov, A. G. Nazina, and
D. A. Papatsenko, Nucl. Acids Res. 31, 6016 (2003).
Eur. J. Appl. Math. 29 (5), 905 (2016).
81. M. Simpson-Brose, J. Treisman, and C. Desplan, Cell
55. M. D. Schroeder, M. Pearce, J. Fak, et al., PLoS Biol.
78 (5), 855 (1994).
2, E271 (2004).
82. E. M. Myasnikova, and A.V. Spirov, BioSystems 166,
56. E. Segal, T. Raveh-Sadka, M. Schroeder, et al., Nature
50 (2018).
451 (7178), 535 (2008).
83. D. E. Clyde, M. S. Corado, X. Wu, et al., Nature 426
57. M. Kazemian, C. Blatti, A. Richards, et al., PloS Biol.
(6968), 849 (2003).
8 (8), e1000456 (2010).
84. X. Y. Li, S. MacArthur, R. Bourgon, et al., PLoS Biol.
58. M. J. Bronski, C. C. Martinez, H. A. Weld, and
6 (2), e27 (2008).
M. B. Eisen, Genes, Genomes, Genetics
10,
85. J. S. Margolis, M. Borowsky, C. W. Shim, and J. W. Po-
g3.400959 (2019). DOI: 10.1534/g3.119.400959
sakony, Dev. Biol. 163, 381 (1994).
БИОФИЗИКА том 66
№ 1
2021
70
МАКАШОВ и др.
86. G. Struhl, P. Johnston, and P. A. Lawrence, Cell 69
90. 90. E. Z. Kvon, T. Kazmar, G. Stampfel, et al., Nature
(2), 237 (1992).
512 (7512), 91 (2014).
87. J. Jaeger, Cell. Mol. Life Sci. 68 (2), 243 (2011).
91. A. Pisarev, E. Poustelnikova, M. Samsonova, and
88. R. Kraut and M. Levine, Development 111 (2), 611
J. Reinitz, Nucl. Acids Res. 37 (Database issue), D560-
(1991).
6 (2009).
89. J. P. Bothma, H. G. Garcia, S. Ng, et al., Elife 4,
92. A. C. Fowlkes, C. L. Luengo Hendriks, S. V. E. Ke-
e07956 (2015).
ränen, et al., Cell 133, 364 (2008).
Fuzzy Linguistic Modelling of Regulation of Drosophila Segmentation Genes
A.A. Makashov*, E.M. Myasnikova*, and A.V. Spirov**, ***
*Peter the Great St. Petersburg Polytechnic University,
ul. Polytechnicheskaya 29, St. Petersburg, 195251 Russia
**Sechenov Institute of Evolutionary Physiology and Biochemistry, Russian Academy of Sciences,
prosp. Toreza 44, St. Petersburg, 194223 Russia
*** Institute of Scientific Information for Social Sciences, Russian Academy of Sciences,
ul. Krzhizhanovskogo 15/2, Moscow, 117997 Russia
Understanding molecular mechanisms involved in gene regulation is one of the underlying issues in systems
biology. Often, common approaches used to model gene regulation fail to distinguish between the model
properties that adequately reflect the characteristics of a biological object and those associated with the na-
ture of the model. This study concerns the development of a logical approach for modeling the regulation of
gene expression thereby enabling further maximum use of all the available empirical data and minimum use
of a priori hypotheses. Moreover, the rules for gene regulation are formulated in the form of linguistic mod-
elling, namely, restated in the language of approach developers instead of the language of researchers. We use
a fuzzy approach known as fuzzy linguistic modelling which reflects reality more accurately. In our study, for
the first time, we used linguistic modelling for modeling the activity of the cis-regulatory modules and applied
it to the problems of analysis of gene expression for biochemical labeling of embryos. This approach made it
possible to clarify the essential details of the mechanisms of “reading out” primary morphogen gradients by
two or more cis-regulatory modules of the same gene in combination.
Keywords: gene regulation, expression patterns, enhancers, enhancer grammar, logical modelling, fuzzy modelling
БИОФИЗИКА том 66
№ 1
2021