БИОФИЗИКА, 2021, том 66, № 1, с. 168-175
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 577.3
МЕТОДЫ ПОДОБИЯ И ТРЕХМЕРНОЕ МОДЕЛИРОВАНИЕ
ЭЛЕКТРИЧЕСКОЙ АКТИВНОСТИ СИНОАТРИАЛЬНОГО УЗЛА СЕРДЦА
КРОЛИКА И СОПРЯЖЕННОГО С НИМ МИОКАРДА ПРЕДСЕРДИЯ
© 2020 г. В.В. Галанин
Медицинский университет «Реавиз», 443001, Самара, ул. Чапаевская, 227
E-mail: galanin_v@hotmail.com
Поступила в редакцию 09.04.2020 г.
После доработки 10.07.2020 г.
Принята к публикации 14.07.2020 г.
Показана возможность применения методов теории подобия для трехмерного моделирования элек-
трической активности cиноатриального узла сердца кролика и сопряженного с ним сократительно-
го миокарда предсердия. Предлагаемая математическая модель представляет собой модель иерар-
хического типа, в которой на верхнем уровне воспроизводится процесс взаимодействия cиноатри-
ального узла и сопряженного с ним сократительного миокарда предсердия; на нижнем уровне
модели отражена динамика ионных процессов на мембранах кардиомиоцитов. Приведены приме-
ры использования данной трехмерной модели для описания: а) генерации ритма в cиноатриальном
узле сердца; б) явления удвоения периода следования потенциалов действия на участке сократи-
тельного миокарда предсердия с пониженной электропроводимостью межклеточных контактов; в)
модулирующего влияния электрического взаимодействия фибробластов и кардиомиоцитов на
электрическую активность синоатриального узла.
Ключевые слова: синоатриальный узел, кардиомиоцит, фибробласт, математическое моделирование,
теория подобия, сложные системы.
DOI: 10.31857/S000630292006018X
сложной системы уравнений нельзя свести к про-
Импульс электрического возбуждения сердца
стой сумме решений образующих ее частей.
формируется в ткани cиноатриального узла
(САУ) в результате синхронизации спонтанной
Вместе с тем, как было показано в предыдущей
электрической активности огромного числа
нашей работе [5], применение методов теории
пейсмекерных клеток. В основе внутрипейсме-
подобия для моделирования САУ позволяет су-
керной синхронизации лежит электротоническое
щественно уменьшить число рассматриваемых
взаимодействие самовозбудимых клеток по-
клеток, не теряя важную информацию о системе
средством щелевых контактов. Такой механизм
в целом. При этом новая редуцированная система
обеспечивает как синхронный переход возбужде-
уравнений на практике оказывается более удоб-
ния с САУ на окружающий сократительный мио-
ной, поскольку теория подобия позволяет не
кард предсердия, так и слаженность работы по-
только уменьшить число используемых диффе-
следнего.
ренциальных уравнений, но и находить парамет-
Вышеописанный процесс моделируется с по-
ры системы как единого целого.
мощью системы, которая может включать более
Более того, настоящий подход позволяет луч-
тысячи взаимосвязанных дифференциальных
ше понять принцип организации данной слож-
уравнений (отметим здесь модели неоднородного
ной биологической системы: в результате взаи-
САУ сердца кролика [1-4] на основе детального
модействия компоненты системы оказываются
описания ионных процессов на мембране кар-
определенным образом подобными всей системе
диомиоцитов). При этом связь между уравнения-
в целом. Здесь по-прежнему под подобием мы по-
ми системы достаточно глубокая, так как измене-
нимаем пропорциональность сходных величин и
ние параметров в уравнении для одной клетки
параметров сопоставляемых объектов.
обязательно влечет изменения в решении для
остальных клеток. Как следствие, решение такой
Целью работы является применение методов
теории подобия для трехмерного моделирования
Сокращениe: САУ - cиноатриальный узел.
электрической активности САУ сердца кролика и
168
МЕТОДЫ ПОДОБИЯ И ТРЕХМЕРНОЕ МОДЕЛИРОВАНИЕ
169
сопряженного с ним сократительного миокарда
предсердия.
ТРЕХМЕРНАЯ МОДЕЛЬ
СИНОАТРИАЛЬНОГО УЗЛА
И СОПРЯЖЕННОГО С НИМ
СОКРАТИТЕЛЬНОГО МИОКАРДА
ПРЕДСЕРДИЯ
У кролика САУ располагается в межвенной
области правого предсердия вдоль пограничного
Рис. 1. Схема трехмерной клеточной сети САУ (Sa) и
гребешка (crista terminalis) между эндокардом и
окружающего его сократительного миокарда пред-
сердия (Atr); c - клетки центра узла, p - клетки его
эпикардом, занимая практически всю толщу
периферии.
стенки [6, 7]. Расстояние от центра узла (области
первичного водителя ритма, leading pacemaker
site) до его периферии в направлении погранич-
тральную часть схемы заполняют пейсмекерные
ного гребешка составляет 2.0-2.5 мм, толщина
клетки узла (затемненная область на схеме), во-
стенки около 0.25-0.75 мм, длина пейсмекерных
круг которого расположены сократительные
клеток САУ > 25 мкм [8]. Тогда можно принять,
клетки предсердия (светлая область на схеме).
что расстояние от центра узла до гребешка превы-
Каждая клетка контактирует с соседними клетка-
шает длину клетки приблизительно в 80 раз, а
ми посредством щелевых контактов.
толщина стенки узла - в 30 раз. С учетом этих
данных представим САУ и окружающий его со-
Колебания мембранного потенциала клеток
кратительный миокард предсердия в виде трех-
САУ и сократительного миокарда предсердия
мерной прямоугольной клеточной сети (рис. 1)
описываются следующей системой дифференци-
размером Mx × My × Mz = 160 × 170 × 30. Всю цен-
альных уравнений:
dV
ijk
ion
x
x
C
ijk
=-I
ijk
+
g
i−1 jk
(
V
i−1 jk
−V
ijk
)
+
g
ijk
(
V
i+1 jk
−V
ijk
)
+
d
t
y
y
+g
(
V
−V
)
+
g
(
V
−V
)
+
(1)
ij−1k ij−1
k
ijk
ijk
ij+1k
ijk
z
z
+g
ijk
−1
(
V
ijk−1
−V
ijk
)
+
g
ijk
(
V
ijk+1
−V
ijk
)
,
где индексы i = 1, …, Mx, j = 1, …, My, k = 1, …, Mz
клетками по направлению осей x, y и z соответ-
ственно.
задают положение клетки; Cijk - электроемкость
Будем считать, что ткань узла по своим био-
мембраны клетки с номером i, j, k; Vijk - ее
электрическим свойствам неоднородна только в
ion
мембранный потенциал;
- сила тока через
I
радиальном направлении [1, 2, 8, 9]. Поэтому вос-
ijk
ионные каналы мембраны клетки (соответствую-
пользуемся следующими формулами для элек-
троемкостей клеток и электропроводимостей
щие формулы для токов приведены в работе [5]);
межклеточных контактов, которые выражают из-
x,y,z
g
- электропроводимости щелевых контак-
ijk
менение этих величин при переходе от центра уз-
тов, соединяющих данную клетку с соседними
ла к его периферии [1, 2, 10]:
C
c
−
C
p
C
=
C
+
,
(2)
ijk
c
2
2
1
+
exp
−a
(
i
−
M
cx
)
+
(
j
−
M
cy
)
−
r
)
(
x,y,z
x,y,z
g
−
g
x,y,z
x,y,z
c
p
g
ijk
=
g
c
+
,
2
2
1
+
exp
−a
(i
−
M
)
+
(
j
−
M
)
−
r
(3)
(
cx
cy
)
i
=
M
p
1x
,...,M
p2
x
,
j
=
M
p
1y
,...,M
p
2
y
,
k
=
1,...,
M
z
,
БИОФИЗИКА том 66
№ 1
2021
170
ГАЛАНИН
x,y,z
где Cс и
- электроемкость мембраны и элек-
g
мость межклеточных контактов на периферии уз-
с
ла; a и r - параметры модели [2, 10]. Кроме того,
тропроводимость межклеточных контактов в
допустим, что максимальная электропроводи-
x,y,z
центре узла (Mcx, Mcy, k), k = 1, …, Mz; Cp и
g
p
-
мость ионных каналов пейсмекерных клеток ли-
электроемкость мембраны и электропроводи- нейно зависит от электроемкости:
ion
ion
ion
ion
g
c
-
g
p
g
ijk
=
g
c
+
(
C
ijk
−C
c
)
(4)
C
c
−
C
p
Электроемкость всех сократительных клеток
потенциала оставшихся клеток подберем мас-
предсердия и электропроводимость их межкле-
штабные коэффициенты
K
x,y,z
электропроводи-
g
ijk
точных контактов примем одинаковыми для всех
мостей клеточных контактов так, чтобы компен-
таких клеток.
сировать увеличение падения потенциала вдоль
Исходная трехмерная модель САУ содержит
ткани узла, возникшее вследствие изъятия части
сотни тысяч клеток и, соответственно, на поря-
слоев из трехмерной клеточной системы. При
этом для масштабных коэффициентов контакт-
док большее количество дифференциальных
уравнений. С другой стороны, число элементов
ных электропроводимостей
K
x,y,z
и масштабных
g
ijk
системы можно существенно сократить, если ис-
коэффициентов для величины падения потенци-
пользовать подход, предлагаемый теорией подо-
ала между соседними элементами
K
,
(Vi-1 jk
-V
ijk )
бия. Для этого по аналогии с предыдущей нашей
K
,
K
,
K
, входящих в систе-
работой [5] оставим в клеточной сети, например,
(Vi+1jk
-V
ijk )
(
V
ij −
−
1k ijk
V
)
(
V
ij +
−
1k ijk
V
)
только каждый десятый слой, отстоящий от цен-
му уравнений (1) и определяющих связь между
тра узла на одинаковом расстоянии. При этом в
соседними клетками, должны выполняться усло-
дифференциальных уравнениях для мембранного
вия:
K
x
⋅
K
=
1,
K
x
⋅
K
=
1,
g
(
V
i −
1jk
−V
ijk
)
g
(
V
i +
1jk
−V
ijk
)
i −1 jk
ijk
(5)
K
y
⋅
K
=
1,
K
y
⋅
K
=
1.
g
ij −1k
(
V
ij −
1
−
k ijk
V
)
g
ijk
(
V
ij +
1k
−V
ijk
)
Использование методов подобия для модели-
отношению к остальной части сложной системы,
рования сложных систем позволяет представить
является правильное воспроизведение связей
отдельные ее подсистемы, не раскрывая полно-
между величинами на границах [11]. Примени-
стью их внутреннего строения, т.е. данный под-
тельно к нашему случаю под связями на границах
ход отражает иерархический тип организации
мы подразумеваем соотношение мембранных по-
сложных биологических систем. При этом одним
тенциалов взаимодействующих клеток.
из важных моментов, обеспечивающим адекват-
На рис. 2 представлены графики решений ре-
ное отражение какой-либо части подсистемы по дуцированной системы дифференциальных урав-
нений (1) при следующих значениях параметров
модели: размер сети Mx × My × Mz = 16 × 17 × 3
(243 пейсмекерные клетки и 573 сократительные
клетки); координаты центральных клеток (Mcx,
Mcy, k) = (8, 9, k); крайних периферических -
(Mp1x, Mp1y, k) = (4, 5, k) и (Mp2x, Mp2y, k) = (12, 13,
1
k), k = 1, …, 3 (рис. 1);
K-
x y,z
=
35; a = 2; r = 1.9.
g
ijk
Выбор масштабных коэффициентов в трех-
мерной модели определялся также следующими
требованиями: а) в окружающем узел сократи-
тельном миокарде возбуждались потенциалы
действия; б) расчетный период следования по-
тенциалов действия T = 320 мс соответствовал
экспериментально наблюдаемому значению ин-
Рис. 2. Колебания мембранного потенциала пейсме-
тактного САУ 348 ± 50 мс [12]; в) наблюдалось по-
керных клеток САУ и сократительного миокарда
предсердия, расположенных вдоль медианной оси
степенное изменение формы потенциала дей-
клеточной сети.
ствия при переходе от центра узла к периферии,
БИОФИЗИКА том 66
№ 1
2021
МЕТОДЫ ПОДОБИЯ И ТРЕХМЕРНОЕ МОДЕЛИРОВАНИЕ
171
характерное для интактного САУ [8, 9] (макси-
мальный диастолический потенциал становился
более негативным, скорость нарастания потен-
циала действия увеличивалась). В нашей трех-
мерной модели максимальный диастолический
потенциал уменьшается от -70 мВ до -77 мВ; ве-
личина скорости нарастания переднего фронта
потенциала действия увеличивается от 19 В/с в
центральной части до 31 В/с на периферии. В од-
номерных моделях данное изменение формы по-
тенциала действия более выражено. В модели [2],
содержащей цепочку из 50 пейсмекерных клеток
Рис. 3. Схема трехмерной клеточной сети редуциро-
и 150 сократительных клеток предсердия, ско-
ванного пейсмекера (Sa) и сопряженного с ним со-
рость нарастания переднего фронта увеличива-
кратительного миокарда предсердия (Atr).
лась от 7 В/с в центре до ≈ 60 В/с на периферии. В
одномерной неоднородной модели (non-uniform
model) [1] из 15 пейсмекерных клеток централь-
дели оказался в два раза меньше, чем в модели
ного типа и 15 клеток периферического типа для
изолированной клетки предсердия [13].
данной величины были получены следующие
Для характеристики эффективности передачи
значения: 8.4 В/с (центр) и 60.7 В/с (периферия).
генерируемого в САУ потенциала действия на со-
Можно предположить, что различие в степени
пряженный с ним сократительный миокард пред-
трансформации формы потенциалов действия
сердия был проведен расчет так называемого
клеток при переходе от центра САУ к периферии
фактора надежности SF (safety factor). Фактор на-
в настоящей трехмерной модели менее выражено
дежности определяется как отношение суммы за-
по сравнению с одномерными цепочками вслед-
рядов Qc, произведенного данной клеткой в фазу
ствие более значительного взаимного электрото-
деполяризации, и Qout, отданного следующей
нического влияния, которое возникает между
клетками в трехмерных сетях. Как представляет-
клетке посредством щелевых контактов, к заряду
ся, по этой же причине овершут потенциала дей-
Qin, полученному ею от предыдущей деполяризо-
ствия типичных клеток предсердия в нашей мо-
ванной клетки:
SF
=
(Q
c
+Q
out
)/Q
in
=
I
c
dt
+
I
out
dt
I
out
dt
,
(6)
A
A
A
где Ic = CdV/dt - емкостный ток, обусловленный
потенциала
K
V
, контактной электропроводимо-
ijk
изменением плотности ионов на поверхности
K
x,y,z
сти
g
, максимальной электропроводимости
мембраны клетки, Iin и Iout - входящий и выходя-
ilk
соответствующих ионных каналов
K
ion
и посто-
щий через щелевые контакты ионные токи, A -
g
ilk
интервал времени от момента, когда скорость
янных времени кинетических переменных
ion
Kτ
ilk
возрастания потенциала действия достигает 1%
удалось заменить весь узел небольшим пейсмеке-
от максимального значения, до момента, когда
ром, состоящим из трех клеток периферического
мембранный потенциал достигает максимума
типа (рис. 3). Этот редуцированный пейсмекер
[14, 15]. Проще говоря, фактор надежности пока-
будет генерировать потенциалы действия
зывает, во сколько раз пороговый уровень, необ-
(рис. 4а) в окружающем сократительном миокар-
ходимый для деполяризации данной клетки, пре-
де аналогично тому, как это происходит в случае
вышает порог деполяризации соседней с ней
узла, размер которого составлял 9 × 9 × 3 (рис. 4б).
клетки, которая ее активизировала. Эффективная
Общее число и порядок расположения сократи-
передача потенциала действия с клетки на клетку
тельных клеток остался прежним.
означает, что SF > 1. В нашей модели вычисления,
выполненные для пейсмекерных клеток, распо-
Процедура подбора масштабных множителей
состояла в следующем. Непосредственная замена
ложенных на границе с сократительными клетка-
пейсмекера тремя клетками периферического ти-
ми, дают величину фактора надежности SF = 1.91.
па приводит к значительному изменению всего
Далее, развивая наш подход, с помощью под-
паттерна электрической активности как в клетках
бора масштабных множителей для мембранного самого узла, так и в клетках сократительного мио-
БИОФИЗИКА том 66
№ 1
2021
172
ГАЛАНИН
Рис. 4. Сравнение потенциалов действия, возбуждаемых редуцированным (а) и многоэлементным (б) пейсмекером в
сократительных клетках, расположенных вдоль медианной оси клеточной сети. Потенциалы действия редуцирован-
ной модели (а) рассчитаны при следующих значениях масштабных коэффициентов: KV = 1.07; KgNa = 0.6; KgCa = 0.4;
KgK = 0.6; KgfNa = 0.1; KτCa = 0.2; KτK = 2.8; Kτf = 4.0; Kg
x,y,z
= 10-4.
c
карда: за счет гиперполяризующего влияния
МОДЕЛИРОВАНИЕ ЭЛЕКТРИЧЕСКОЙ
большого числа сократительных клеток (в нашей
АКТИВНОСТИ СОКРАТИТЕЛЬНОГО
модели их число равно 573) на оставшиеся три
МИОКАРДА ПРЕДСЕРДИЯ ПРИ НАЛИЧИИ
В НЕМ НЕОДНОРОДНОСТИ ПО
пейсмекерные клетки в новой редуцированной
КОНТАКТНОЙ ЭЛЕКТРОПРОВОДИМОСТИ
клеточной системе потенциалы действия не воз-
буждаются. Поэтому, чтобы существенно умень-
Смоделируем участок, обладающий понижен-
шить данное влияние и создать условия для гене-
ной электропроводимостью межклеточных кон-
рации потенциала действия, в уравнениях для
тактов в области сократительного миокарда пред-
мембранного потенциала оставшихся пейсмекер-
сердия (наиболее темный участок на рис. 5, обо-
ных клеток был введен масштабный множитель
значенный «block»). В расчетах использовалась
описанная выше модель размером 16 × 17 × 3. При
контактной электропроводимости
K
x,y,z
<<
1. За-
gc
снижении контактной электропроводимости на
тем с помощью выбора масштабных множителей,
данном участке до 5.05% от проводимости между
соответствующих мембранному потенциалу и
остальными сократительными клетками в нем
максимальной проводимости ионных каналов,
возникают потенциалы действия с удвоенным
достигается равенство амплитуды потенциала
периодом (рис. 6). Данный феномен можно объ-
яснить тем, что при увеличении межклеточной
действия редуцированной и многоэлементной
проводимости происходит фазовый сдвиг мем-
моделей. Наконец, для совпадения периодов воз-
бранного потенциала. В результате импульс од-
буждаемых потенциалов действия в уравнениях
ной из клеток попадает в рефрактерный период
для кинетических переменных ионных каналов
соседней с ней и потенциал действия для нее не
были подобраны соответствующие значения мас-
возникает. Эффекты, подобные тем, что описаны
штабных множителей постоянных времени.
выше, отмечались и в работе [16].
Рис. 5. Схема трехмерной клеточной сети САУ (SA) и
окружающего его сократительного миокарда
Рис.
6. Изменение формы потенциала действия
предсердия (Atr) с участком (Block), обладающим
сократительных клеток предсердия, расположенных
пониженной электропроводимостью межклеточных
вдоль правой границы клеточной сети (i =16, j = 1...9,
контактов. Участок block электрически изолирован
k
= 2). Участку block соответствуют клетки с
от клеток предсердия, расположенных слева от него.
номерами i =16, j = 1...3, k = 2,
БИОФИЗИКА том 66
№ 1
2021
МЕТОДЫ ПОДОБИЯ И ТРЕХМЕРНОЕ МОДЕЛИРОВАНИЕ
173
УЧЕТ НАЛИЧИЯ СЕРДЕЧНЫХ
ФИБРОБЛАСТОВ В ТКАНИ
СИНОАТРИАЛЬНОГО УЗЛА.
МОДУЛИРУЮЩЕЕ ВЛИЯНИЕ
ЭЛЕКТРИЧЕСКОГО ВЗАИМОДЕЙСТВИЯ
ФИБРОБЛАСТОВ И КАРДИОМИОЦИТОВ
НА ЭЛЕКТРИЧЕСКУЮ АКТИВНОСТЬ
СИНОАТРИАЛЬНОГО УЗЛА
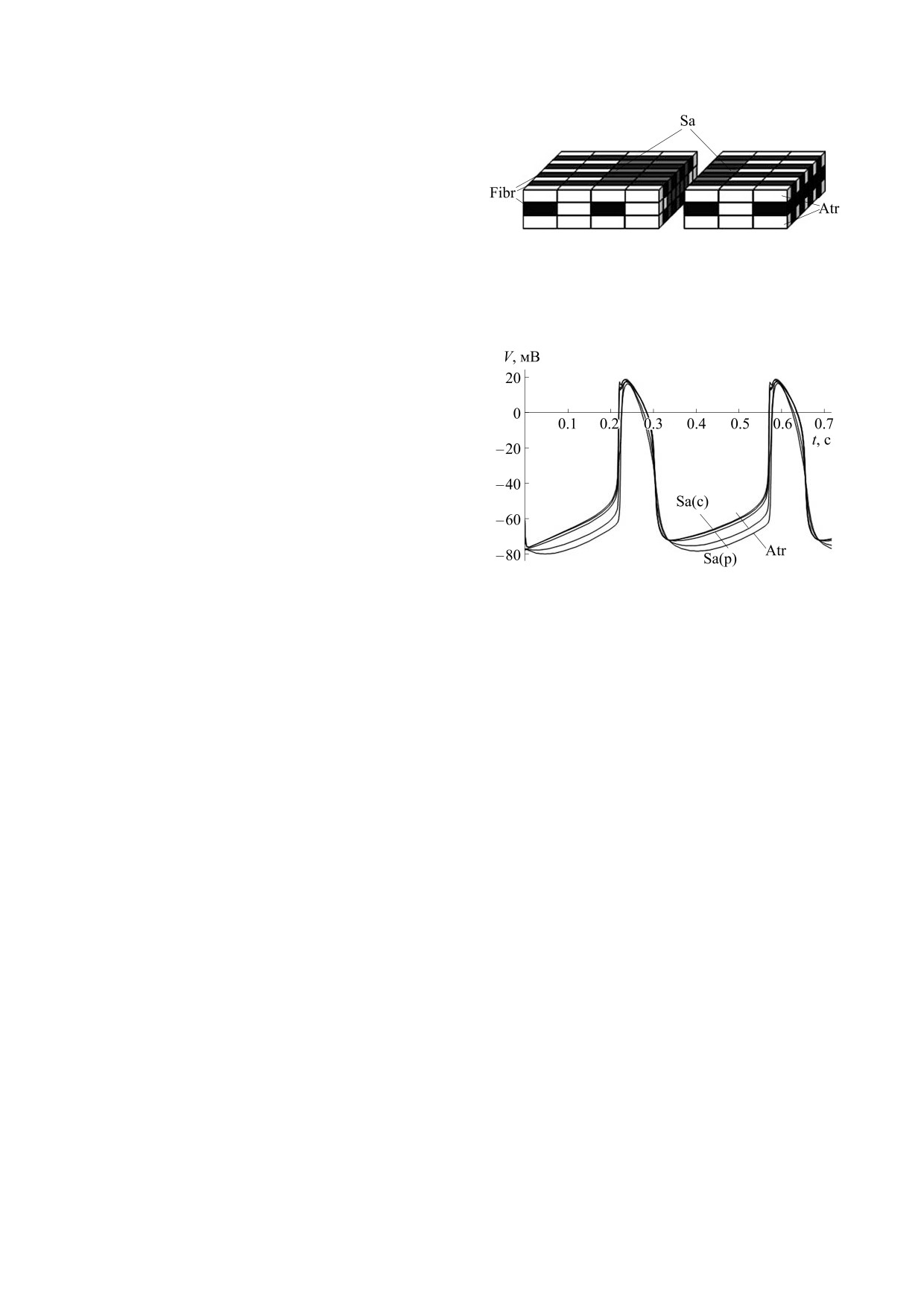
Рис. 7. Схема трехмерной сети клеточных элементов
Ткань САУ пронизана сетью сердечных фиб-
САУ (Sa) и окружающего его сократительного
робластов (их доля составляет более 50% от обще-
миокарда предсердия (Atr) с включением сети
го количества клеток узла), являющихся электро-
сердечных фибробластов (Fibr).
невозбудимыми клетками и взаимодействующи-
ми друг с другом через щелевые контакты [17, 18].
Кроме того, между фибробластами и кардиомио-
цитами также имеются щелевые контакты, и ион-
ные токи через мембраны фибробластов напря-
мую влияют на электрическую активность близ-
лежащих кардиомиоцитов [18, 19].
Воспользуемся нашей трехмерной прямо-
угольной клеточной схемой САУ и сократитель-
ного миокарда, включив в нее сеть фибробластов
в виде многоуровневой решетки (рис. 7).
Дифференциальные уравнения для мембран-
ного потенциала фибробластов имеют вид, ана-
логичный уравнениям (1). Электроемкость мем-
браны фибробласта 6.3 пФ, электропроводимость
щелевых контактов, соединяющих фибробласты
Рис. 8. Колебания мембранного потенциала клеток
САУ и окружающего миокарда предсердия, взаимо-
с соседними кардиомиоцитами, примем равной
действующих с сетью сердечных фибробластов.
ion
2.5 нСм [20, 21]. Выражения для силы тока
че-
I
ijk
рез ионные каналы мембраны фибробластов, а
Далее рассмотрим влияние, оказываемое сжа-
также величины остальных параметров ионных
тыми фибробластами, на электрическую актив-
каналов соответствуют модели, предложенной в
ность кардиомиоцитов САУ и окружающего со-
работе [20].
кратительного миокарда с помощью следующего
Фибробласты отвечают на механическое воз-
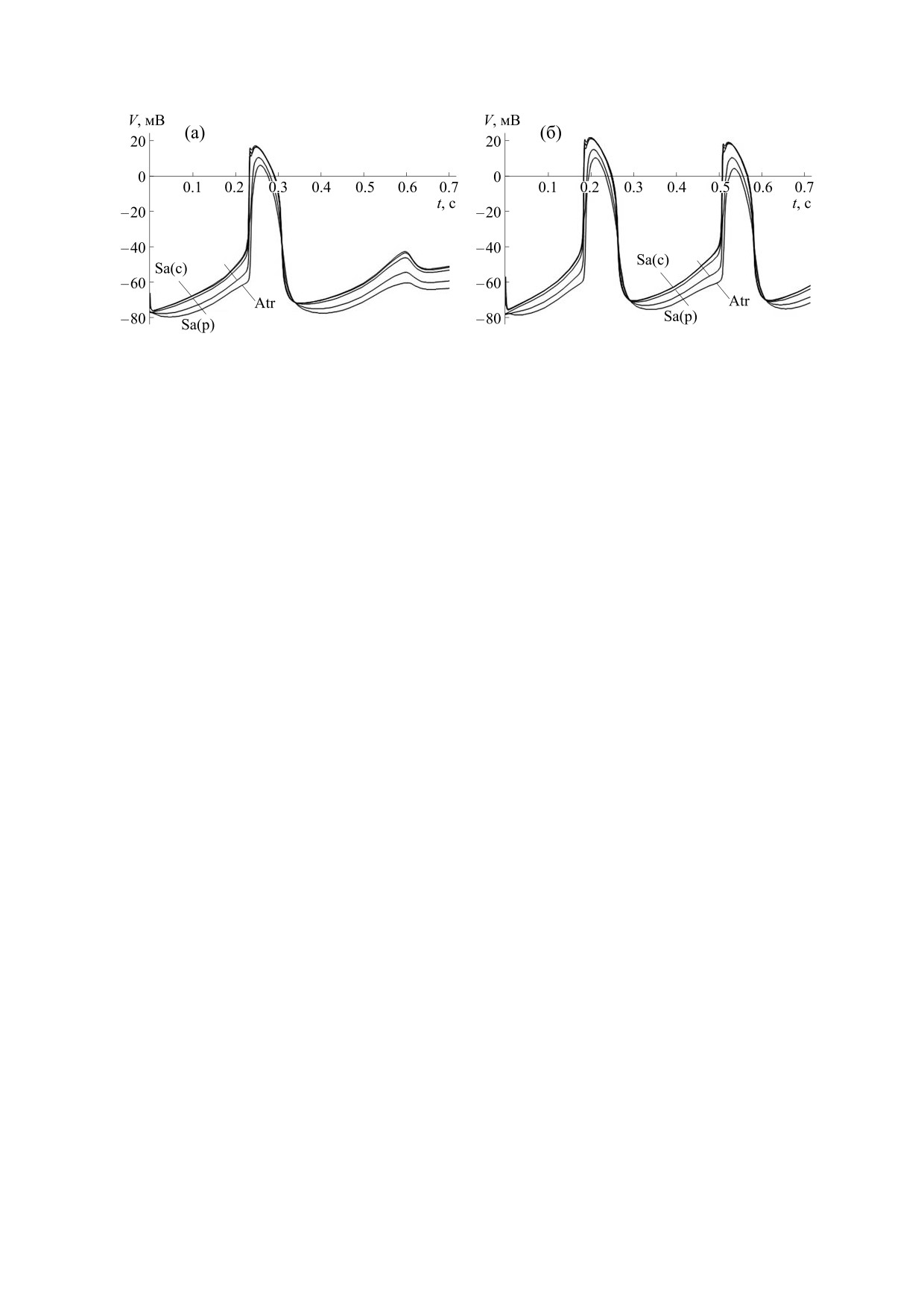
численного эксперимента. Уменьшим электро-
действие генерацией механоиндуцированного
проводимость контактов между кардиомиоцита-
потенциала [22]. Его появление обусловлено на-
ми настолько, чтобы в ткани САУ не могли воз-
личием в мембране фибробластов механочув-
никнуть повторные потенциалы действия
ствительных ионных каналов (stretch-activated
(рис. 9а). Этого можно достигнуть, если умень-
channel). Поэтому дополним используемую элек-
шить контактную электропроводимость в нашей
трофизиологическую модель мембраны фиброб-
модели в 1.23 раза.
ластов слагаемым, описывающим ионный ток че-
рез механочувствительные ионные каналы:
Затем создадим условие, соответствующее
Isac = gsac (V - Erev), где gsac - электропроводи-
сжатию фибробластов ткани синусного узла.
Вследствие активации механочувствительных
мость механочувствительных каналов, Erev - по-
ионных каналов на мембране фибробластов по-
тенциал реверсии.
является деполяризующий механоиндуцирован-
Рис. 8 иллюстрирует решение системы диффе-
ный потенциал. Например, сжатие фибробласта
ренциальных уравнений для мембранного потен-
на величину 4 мкм приводит к возникновению
циала, соответствующего узловым и сократитель-
электропроводимости механочувствительных ка-
ным клеткам трехмерной модели с номерами
налов и потенциала реверсии, равных
i = 1, …, 9, j = 9, k = 2, в отсутствие деформации
gsac = 5.6 нСм и Erev = 16.0 мВ [22]. При этом, как
мембраны фибробластов (gsac = 0). Результатом
показали результаты проведенных расчетов, сме-
взаимодействия фибробластов и кардиомиоци-
щение мембранного потенциала фибробластов в
тов является замедление ритма электрической ак-
сторону деполяризации создало условие для воз-
тивности САУ. Период колебаний мембранного
никновения повторных потенциалов действия в
потенциала клеток увеличился с 320 мс в отсут-
ткани САУ и сопряженного с ним сократительно-
ствие фибробластов до 340 мс при их наличии.
го миокарда (рис. 9б). Таким образом, сеть фиб-
БИОФИЗИКА том 66
№ 1
2021
174
ГАЛАНИН
Рис. 9. Мембранные потенциалы клеток САУ и окружающего миокарда предсердия в сети с фибробластами при низ-
кой контактной электропроводимости в отсутствие (а) и при наличии (б) механического сжатия фибробластов.
робластов оказывает модулирующее действие на
БЛАГОДАРНОСТИ
электрическую активность кардиомиоцитов тка-
Автор сердечно благодарит к.б.н. А.И. Сироту
ни САУ.
за неустанное внимание, поддержку и полезные
обсуждения ряда вопросов, затрагиваемых в
статье.
ЗАКЛЮЧЕНИЕ
Представленная в настоящей работе математи-
КОНФЛИКТ ИНТЕРЕСОВ
ческая модель ткани САУ сердца кролика показа-
Авторы заявляют об отсутствии конфликта
ла, что использование подхода теории подобия -
интересов.
это действенный метод для моделирования данной
сложной биологической системы, позволяющий
значительно уменьшить объем вычислений при
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
сохранении наиболее существенных свойств и па-
Настоящая работа не содержит описания ис-
раметров модели. При таком подходе появляется
следований с использованием людей и животных
больше возможностей для предсказания новых
в качестве объектов.
свойств моделируемой системы, что способствует
лучшему пониманию механизмов, лежащих в ос-
нове изучаемых процессов.
СПИСОК ЛИТЕРАТУРЫ
1. R. V. Oren and C. E. Clancy, PLoS Comput. Biol. 6
Результаты, приведенные в работе, показыва-
(12), e1001041
(2010). DOI:
10.1371/journal.pcbi.
ют, что данная модель воспроизводит наблюдае-
1001041
мые процессы генерации ритма в САУ сердца, яв-
2. S. Inada, H. Zhang, J. O. Tellez, et al., PLoS One 9 (4),
ление удвоения периода следования потенциалов
e94565 (2014). DOI: 10.1371/journal.pone.0094565
действия на участке сократительного миокарда
3. C. Ly and S. H. Weinbergb, J. Theor. Biol. 439, 35
предсердия с пониженной электропроводимо-
(2018).
стью межклеточных контактов, модулирующее
4. Р. А. Сюняев и Р. Р. Алиев, Биофизика 54 (1), 77
влияние электрического взаимодействия фиб-
(2009).
робластов и кардиомиоцитов на электрическую
5. В. В. Галанин, Биофизика 64 (3), 578 (2019).
активность САУ. Кроме того, используемый в ра-
боте подход можно будет применить к моделям,
6. T. Opthof, Cardiovasc. Drug. Ther. 1, 573 (1988).
предназначенным для изучения разнообразных
7. Д. В. Абрамочкин, Г. С. Сухова и Л. В. Розенштра-
эффектов электромеханического сопряжения в
ух, Успехи физиол. наук 40 (4), 21 (2009).
миокарде, а также для исследования возможных
8. W. K. Bleeker, A. J. C. Mackaay, M. Masson-Pevet,
нарушений сердечного ритма.
et al., Circ. Res. 46, 11 (1980).
9. M. R. Boyett, H. Honjo, M. Yamamoto, et al., Am. J.
Несмотря на то что в настоящей трехмерной
Physiol. 276, H686 (1999).
модели использовались данные для сердца кро-
10. H. G. Zhang, A. V. Holden, I. Kodama, et al., Am. J.
лика, наш метод можно обобщить для моделиро-
Physiol. Heart Circ. Physiol. 279, 397 (2000).
вания процессов электрической активности в
11. В. А. Веников и Г. В. Веников, Теория подобия и мо-
предсердиях и других видов теплокровных жи-
делирования (применительно к задачам электро-
вотных.
энергетики) (УРСС, М., 2014).
БИОФИЗИКА том 66
№ 1
2021
МЕТОДЫ ПОДОБИЯ И ТРЕХМЕРНОЕ МОДЕЛИРОВАНИЕ
175
12. C. J. H. J. Kirchhof, F. I. M. Bonke, M. A. Allesie, and
18. P. Kohl, P. Camelliti, F. L. Burton, and G. L. Smith, J.
W. J. E. P. Lammers, Pflugers Arch. 410, 198 (1987).
Electrocardiol. 38, 45 (2005).
13. B. Wohlfart and G. Ohlen, Clin. Physiol. 19, 11 (1999).
19. А. С. Толстокоров, Р. А. Сюняев и Р. Р. Алиев,
14. R. M. Shaw and Y. Rudy, Circ. Res. 81 (5), 727 (1997).
Биофизика 60 (2), 322 (2015).
15. Y. Wang and Y. Rudy, Am. J. Physiol. Heart Circ.
20. K. A. Mac Cannell, H. Bazzazi, L. Chilton, et al., Bio-
Physiol. 278, H1019 (2000).
phys. J. 92, 4121 (2007).
16. P. Ostborn, B. Wohlfart, and G. Ohlen, J. Theor. Biol.,
21. P. Kohl, A. G. Kamkin, I. S. Kiseleva, and D. Noble,
211, 201 (2001).
Exp. Physiol. 79, 943 (1994).
17. P. Camelliti, G. R. Green, I. Le Grice, and P. Kohl,
22. A. G. Kamkin, I. S. Kiseleva, and G. Isenberg, Cardio-
Circ. Res. 98 (6), 828 (2004).
vasc. Res. 57, 793 (2003).
Similitude Methods and Three-Dimensional Simulation of the Electrical Activity
of the Rabbit Sinoatrial Node and Adjacent Atrial Myocardium
V.V. Galanin
Medical University «Reaviz», Chapayevskaya ul. 227, Samara, 443001 Russia
This study focuses on possible applications of similitude design methods for three-dimensional simulation of
the electrical activity of the sinoatrial node of the rabbit heart and adjacent atrial myocardium. The proposed
mathematical model is hierarchy-shaped where the relationship between the sinoatrial node and adjacent
atrial myocardium is at the top and the dynamics of ion processes on the cardiomyocytes membranes is at the
lower level of the model. Examples of using this three-dimensional model are given for the description of: a)
rhythm generation in the sinoatrial node of the heart; b) the effect of doubling the action potential duration
in the site of the atrial myocardium with reduced electrical intercellular conductivity; c) the modulating effect
of the electrical fibroblast-cardiomyocyte relationship on the pacemaker activity.
Keywords: sinoatrial node, cardiomyocyte, fibroblast, mathematical simulation, similitude design method, com-
plex systems
БИОФИЗИКА том 66
№ 1
2021