БИОФИЗИКА, 2021, том 66, № 2, с. 219-229
МОЛЕКУЛЯРНАЯ БИОФИЗИКА
УДК 535.8; 574.583; 578.4; 577.3
РОЛЬ ВИРУСОВ И ВИРУСНОГО ЛИЗИСА В ИЗМЕНЕНИИ ОПТИЧЕСКИХ
СВОЙСТВ ВОДНОЙ СРЕДЫ ИХ ОБИТАНИЯ
© 2021 г. С.А. Шоларь*, О.А. Степанова**
*Морской гидрофизический институт РАН, 299011, Севастополь, ул. Капитанская, 2
E-mail: sa.sholar@mail.ru
**Институт природно-технических систем РАН, 299011, Севастополь, ул. Ленина, 28
E-mail: solar-ua@ya.ru
Поступила в редакцию 30.03.2020 г.
После доработки 17.12.2020 г.
Принята к публикации 11.01.2021 г.
В обзоре представлены данные о роли вирусов гидросферы и вызываемого ими вирусного лизиса
одноклеточных хозяев в изменении оптических свойств воды. В ранних работах считалось, что
вирусы водоемов как представители наноразмерной фракции микропланктона не оказывают
существенного влияния на оптические свойства воды. Однако в дальнейших экспериментах
зарубежных коллег и собственных исследованиях авторов было выявлено, что вирусный лизис
представителей бактерио- и фитопланктона приводит к многочисленным последствиям, итогом
которых является изменение оптических характеристик воды как жидкой среды их обитания.
Предположено, что обнаруженное явление можно использовать в практических целях.
Ключевые слова: оптика моря, вирусы гидросферы и их хозяева, вирусный лизис, показатель ослабления
света.
DOI: 10.31857/S0006302921020022
тически значимых частиц (бактерий, представи-
Океаническая вода, как и вода любого водое-
телей фитопланктона), может быть рассмотрен
ма, представляет собой сложную физико-хими-
учеными и оценен как фактор, приводящий к из-
ко-биологическую систему, включающую рас-
менениям в составе и численности оптически ак-
творенные вещества, взвесь и множество разно-
тивных компонентов. Иными словами, вирусы
образных живых организмов. Основными
водоемов, как самые многочисленные и наиме-
факторами, определяющими ее оптические свой-
нее изученные, но играющие ключевую роль в
ства, как указано в работе К.С. Шифрина «Введе-
глобальной экосистеме [4, 5], могут быть значи-
ние в оптику океана» [1], являются оптические
мы и в физике водоемов, в том числе в оптике мо-
компоненты - чистая вода, растворенные веще-
ря и гидросферы в целом.
ства (неорганические и органические) и взвесь -
минеральная и органическая. Множество разно-
Влияние различных факторов биотического и
образных органических веществ в водной среде
абиотического происхождения на вирусы гидро-
сферы в настоящее время находится еще в стадии
находятся в двух формах - входят в состав частиц
активного исследования и осмысления. Логичное
взвеси, в частности, в планктон (микропланк-
тон), либо молекулярно растворены в океаниче-
предположение, что высокая численность виру-
ской воде [1]. Состав фитопланктонного сообще-
сов в водоемах и те процессы, за которые вирусы
ответственны, могут отражаться на изменениях
ства является важным источником оптической
некоторых физических параметров водной среды
изменчивости в океане [2]. При этом виропланк-
их обитания, пока еще не получило достаточных
тон, увеличивающаяся сезонная численность ко-
подтверждений в силу слабой изученности. Такая
торого отмечается в периоды «цветения» его хозя-
ситуация связана в первую очередь со сравни-
ев - представителей бактерио- и фитопланктона,
тельно недавней активизацией исследований в
также является составной частью микропланкто-
области молодой науки - морской (водной) виру-
на [3]. Следовательно, являясь частью планктона,
сологии, произошедшей лишь в конце XX столе-
виропланктон, вызывающий вирусный лизис оп-
тия, когда впервые для прямого счета вирусов в
Сокращения: РОВ - растворенное органическое вещество,
пробах воды использовали электронную микро-
ПОС - показатель ослабления света.
скопию [6].
219
220
ШОЛАРЬ, СТЕПАНОВА
Бурное развитие нового научного направле-
их обитания [21, 22]. Одной из причин слабой за-
ния - морской (водной) вирусологии - постави-
интересованности специалистов по физике и
ло перед учеными множество проблем, среди ко-
биофизике моря в изучении влияния вирусов и
торых можно отметить следующие:
вирусного лизиса на некоторые физические пара-
• контаминация гидросферы и ее обитателей
метры водной среды их обитания в лабораторных
вирусами суши с проистекающими последствия-
и естественных (природных) условиях может
ми вплоть до их адаптации к новым условиям сре-
быть недостаточная информированность в этом
ды и возникновение так называемых «новых» ви-
направлении (по этому вопросу). Проведенный
русов;
литературный поиск выявил ряд работ зарубеж-
ных коллег по подробному изучению вирусного
• значение морских вирусов в инфекционной
лизиса как морских бактерий и цианобактерий,
патологии гидробионтов и их возможная роль
так и микроводорослей в условиях эксперимента,
в патологии организмов суши, в том числе чело-
а также и его влияния на оптические свойства
века;
водной среды [2, 23-29].
• изучение сезонности, численности, морфо-
логии и размерного спектра сообществ виро-
Публикации отечественных исследователей
планктона и виробентоса как отражение эколо-
свидетельствуют о значении хозяев морских виру-
гии их хозяев;
сов, например представителей фитопланктона, в
• роль вирусов в круговороте органического
оптике моря [30-32], а также в изменении оптиче-
углерода, в биоразнообразии, в процессах функ-
ских характеристик в жидкой среде наращиваемых
ционирования пищевых цепей в водоемах, а так-
культур микроводорослей в лабораторных услови-
же влияние вирусов и вирусного лизиса на физи-
ях [33]. Есть сведения и о влиянии представителей
ческие параметры водной среды их обитания;
морской микробиоты, в частности бактерий, водо-
рослей и простейших одноклеточных, на «элек-
• изучение воздействий глобальных и регио-
трическую энергию» в гидросфере, что «проявля-
нальных экологических факторов на вирусную
ется на акватории макроскопически - в виде ло-
составляющую гидросферы, в том числе на от-
дельных представителей;
кального электрического эффекта» [34]». С учетом
недостаточной изученности и освещенности в пуб-
• поиск, изоляция и изучение отдельных пред-
ликациях проблемы влияния вирусов и вирусного
ставителей вирусов гидросферы и их геномов, в
лизиса на физические характеристики водной сре-
том числе альговирусов, ответственных за лизис
ды, в том числе значения черноморской микро-
первичных продуцентов органической материи,
биоты в этом явлении, нами были выполнены не-
диметилсульфида и кислорода в Мировом Океа-
которые исследования. Результаты наших первых
не, что влияет на атмосферу и климат нашей Пла-
экспериментальных работ с использованием чер-
неты;
номорской микробиоты (разные штаммы альгови-
• практическое применение вирусов гидро-
русов и культур разных видов микроводорослей)
сферы (в изучении сезонности и географии рас-
свидетельствовали об изменении электрической
пространения и распределения хозяев-микроор-
проводимости морской воды при вирусном лизисе
ганизмов, в определении видов неизвестных мик-
[35, 36]. Также наши исследования в лабораторных
роводорослей, в фармакологии, в биологических
экспериментах, как и работы зарубежных коллег,
способах борьбы с обрастателями и коррозией, а
выявили значение вирусного лизиса в изменении
также и в других направлениях нанотехнологий).
оптических свойств водной среды обитания виру-
По мере развития водной вирусологии круг
сов гидросферы [36-39].
проблем только расширяется. Определенную
лепту в решение проблем водной вирусологии
Цель представленной работы - проанализиро-
внесли результаты и наших исследований, как и
вать имеющуюся информацию о роли вирусов и
результаты других российских ученых-исследо-
вирусного лизиса в гидросфере в изменении оп-
вателей из Лимнологического института СО РАН
тических свойств воды, как среды их обитания,
(Иркутск) [7-13] и Института биологии внутрен-
на уровне экспериментальных и полевых иссле-
них вод им. И.Д. Папанина (пос. Борок Ярослав-
дований. При этом были поставлены следующие
ской области) [14-20].
задачи - выявить теоретические предпосылки
Анализ доступной литературы свидетельствует
значения вирусного лизиса в оптике гидросферы,
о слабой исследовательской заинтересованности
описать некоторые эксперименты по изучению
и активности отечественных ученых и ученых
вирусного лизиса и набор используемых при этом
стран бывшего СССР в получении знаний о веро-
материалов и методов, представить результаты
ятной роли вирусов гидросферы и вызываемого
собственных исследований на основе черномор-
ими лизиса одноклеточных хозяев (представите-
ской микробиоты, выдвинуть предположения о
лей бактерио- и фитопланктона) в изменениях
возможном использовании альговирусов и ви-
некоторых физических параметров водной среды
русного лизиса в практических целях.
БИОФИЗИКА том 66
№ 2
2021
РОЛЬ ВИРУСОВ И ВИРУСНОГО ЛИЗИСА
221
Первый этап в изучении роли вирусов и вирусно-
ды по данным зарубежных исследователей. В ран-
го лизиса в изменении оптических свойств воды, как
них работах по изучению оптических свойств
среды их обитания, в качестве теоретических пред-
вирусов, средний размер которых составляет
посылок. Основание в 1892 г. вирусологии как
0.07 мкм (70 нм), была определена их слабая роль
науки, принадлежащее нашему отечественному
в рассеивании и преломлении света по сравне-
ученому Д.И. Ивановскому, привело к тому, что
нию с такими оптически активными компонен-
уже к началу XX века (с 1926 г.) исследователи за-
тами, как их хозяева - представители бактерио- и
нимались выделением вирусов (бактериофагов)
фитопланктона [53]. К заключению о том, что
из морской воды к ряду патогенных микроорга-
водные вирусы, хотя и очень распространены в
низмов. В середине XX века наши отечественные
море, не являются основным источником обрат-
исследователи - гидробиологи А.Е. Крисс и
ного рассеяния, пришли и исследователи, ис-
Е.А. Рукина проводят изоляцию вирусов (бакте-
пользующие в экспериментах лазерный нефело-
риофагов) к морским бактериям из материала
метр [23, 24]. Тем не менее ими было показано,
(воды и ила), отобранного из экосистемы Черно-
что вирусная инфекция и лизис гетеротрофных
го моря [40, 41]. При этом используются методи-
бактерий резко изменяют форму индикатрисы
ки, позволяющие накопить морские вирусы в
рассеяния, делая ее более плоской как в прямом,
жидкой культуре бактерий, что в случае присут-
так и в обратном направлениях. Однако, несмот-
ствия вирусов наблюдалось в виде просветления
ря на высокую численность водных вирусов, уче-
и повышения прозрачности жидкой среды за счет
ными в ранних работах высказывались предполо-
контакта бактериофагов со своими хозяевами
жения, что морские вирусы играют второстепен-
(клетками культур бактерий) и последующего ви-
ную роль в оптике океана из-за их малого размера
русного лизиса. Иными словами, вирусный лизис
и рассеивающего поперечного сечения.
одноклеточных хозяев в жидкой среде наблюда-
В исследованиях отмечается, что фундамен-
ется в виде повышения прозрачности этой среды,
тальные изменения в рассеянии света связаны с
что давно известно специалистам в области виру-
заражением морских бактерий, а именно основ-
сологии и используется в ряде вирусологических
ное влияние вирусов на оптические свойства вод-
методик, в том числе и в авторских [42, 43].
ной среды связано с вирусным лизисом их хозяев
Высокая оценка учеными водных вирусов как
[24]. В этой работе эксперименты проводились в
главных игроков в глобальной экологии обуслов-
100-литровых автоклавируемых полиэтиленовых
лена их влиянием на функционирование водных
пакетах, при этом в схему эксперимента кроме
экосистем за счет активного участия в круговоро-
емкости с зараженными вирусом бактериями
те органического углерода, углекислого газа, кис-
входил и контроль (емкость с бактериями без ви-
лорода, диметилсульфида (биоактивный газ,
руса). Неполяризованное объемное рассеяние
участвующий в образовании облаков), в биораз-
света на длине волны 514 нм от бактериаль-
нообразии своих хозяев, в процессах функциони-
ных/вирусных суспензий измеряли с помощью
рования пищевых цепей и пр. [3-5, 44-48]. А од-
лазерного нефелометра (Dawn Wyatt Technologies,
но из важнейших и крайне актуальных значений
США). Источник света этого прибора представ-
вирусов гидросферы - их потенциальная роль в
ляет собой аргон-ионный лазер с вертикально по-
патологии организмов суши, в том числе и людей
ляризованным световым лучом. В работе впервые
[49-51].
количественно описано два оптических измене-
ния, связанных с инфицированием и лизисом
Однако для расширения знаний об экологии
морских бактерий бактериофагом: 1) быстрые,
водных вирусов необходимо также уделять вни-
сильные сдвиги в величине и форме индикатрисы
мание изучению вопросов, связанных с ролью
рассеяния света; 2) быстрое увеличение концен-
вирусов, как самых многочисленных гидробион-
трации окрашенного растворенного органиче-
тов, и вызываемого ими вирусного лизиса в изме-
ского материала («желтого вещества», именуемо-
нении оптических характеристик водной среды
го также РОВ). Высказано предположение, что
их обитания для дальнейшего учета этого явления
вызванные вирусом (вирусным лизисом) измене-
в теории и практике оптики моря и гидросферы в
ния обратного рассеяния будут обнаруживаться с
целом. Полученные при изучении значения ви-
помощью спутниковых или авиационных мето-
русного лизиса в оптике водной среды результаты
дов дистанционного зондирования, что позволит
помогут при объяснении наблюдаемых аномалий
определять локализацию наведенной вирусами
оптических свойств воды в разных океанических
гибели их хозяев. Так, согласно работе [54], в зо-
регионах и послужат оригинальным дополнени-
не, где произошел вирусный лизис, по спутнико-
ем к выдвигаемым разными учеными предполо-
вым данным отмечалась повышенная отража-
жениям и объяснениям [52].
тельная способность поверхности. Появление об-
Эксперименты и набор используемых материа-
ласти с высокой отражательной способностью в
лов и методов при изучении влияния вирусов и ви-
западном Ла-Манше, наблюдаемой со спутника,
русного лизиса на оптические свойства водной сре-
по мнению исследователей было вызвано прекра-
БИОФИЗИКА том 66
№ 2
2021
222
ШОЛАРЬ, СТЕПАНОВА
щением цветения микроводоросли Emiliania hux-
массовая клеточная смертность таких масштабов
leyi. В экспедиционных исследованиях на четы-
может быть обнаружена в природных системах с
рех станциях были взяты пробы воды, причем од-
помощью современных дистанционных датчи-
на станция была снаружи и три станции - в зоне
ков. В работе использовали метод трансмиссион-
высокой отражательной способности. Для мони-
ной электронной микроскопии, что позволяло
торинга численности фитопланктона, кокколи-
наблюдать повышенное количество вирусопо-
тов, бактерий и вирусов в этом исследовании ис-
добных частиц. В серии экспериментов проводи-
пользовали аналитическую проточную цитомет-
ли описание изменений оптических свойств жид-
рию. Было выявлено, что внутри зоны
кой среды при распаде клеток доминирующего
концентрация вирусов была выше, численность
вида в результате вирусного лизиса. Исследовате-
клеток Е. huxleyi ниже, число кокколитов и бакте-
ли высказали мнение, что после лизиса домини-
рий выше. Эти данные были интерпретированы
рующих видов собственные оптические свойства
учеными как вызванный вирусом лизис клеток
(поглощение и рассеяние) водоема резко изме-
E. huxleyi в период цветения, вызывающий отде-
нятся и наблюдаемые изменения в оптических
ление больших концентраций кокколитов, что
свойствах исследуемого объема будут обусловле-
приводит к высокой отражательной способности,
ны разрушением (лизисом) инфицированных
наблюдаемой на спутниковых снимках. Эта ин-
клеток, который вызывает высвобождение рас-
терпретация была подтверждена выделением двух
творимых и нерастворимых веществ в окружаю-
вирусов, Eh V84 и Eh V86, из области высокой от-
щую среду.
ражательной способности.
Таким образом, во время фазы вирусного ли-
Таким образом, ученые приходят к выводу о
зиса популяции хозяина, рассеяние и поглоще-
том, что вирусная инфекция оптически значимых
ние живыми клетками или их колониями в боль-
биологических частиц представляет собой еще
шинстве случаев изменяются на поглощение све-
один процесс, который может быстро трансфор-
та малыми частицами и РОВ. Кроме того, можно
мировать клетки микропланктона в новые типы
ожидать положительный отклик со стороны гете-
оптически значимого материала, т. е. окрашен-
ротрофных бактерий, инфузорий, нанофлагеллят
ное растворенное органическое вещество и об-
и других представителей микропланктона в ре-
ломки клеток. Следовательно, вирусы и вызывае-
зультате высвобождения свежего органического
мый ими процесс вирусного лизиса оказывают
материала, вызывающего дальнейшие измене-
значительное влияние на оптические свойства
ния, наблюдающиеся в окраске воды. Описание
вод океана.
обнаруженных в ходе работы оптических измене-
В работе [27] в лабораторных условиях изуча-
ний при массовом лизисе может, по мнению ис-
следователей, в будущем использоваться для
лись оптические изменения, сопровождающие
идентификации вирусолитических событий в
распад популяции нитчатых цианобактерий из
природе по данным оптического дистанционного
мелкого эвтрофного озера. В экспериментальных
зондирования, получаемого со спутников или
условиях было обнаружено резкое разрушение
доминирующих видов водорослей или цианобак-
датчиков на буях и судах. В связи с этим исследо-
терий, что, в свою очередь, по мнению авторов,
ватели основное внимание уделили описанию
прелизиса или пику цветения фитопланктона по-
может быть связано с вирусной активностью. В
сле лизиса. Полученные результаты свидетель-
течение двух недель непрерывного добавления
ствуют о самых больших оптических изменениях,
богатой питательными веществами среды отме-
чалось почти полное разрушение доминирующей
которые могут произойти в полевых условиях в
описываемые в работе периоды. Иными словами,
популяции за несколько дней. При этом разруше-
выполненные исследования и интерпретация по-
ние происходило неоднократно и характеризова-
лученных данных могут быть экстраполированы
лось заметным повышением прозрачности воды.
на природные условия, в гидросферу.
Рассеяние света уменьшилось на 80%, поглоще-
ние - на 20-80%. Наблюдалось большое сходство
В работе [25] описаны изменения в экспери-
в оптических изменениях между несколькими
ментах с течением времени концентраций одно-
экспериментами, проведенными в разные сезоны
клеточного хозяина и вируса, наряду с соответ-
года. Во время разрушения хозяев вирусов отме-
ствующими изменениями первичных гидроопти-
чали увеличение количества РОВ и частиц суб-
ческих характеристик, когда четыре штамма
микронного размера, спектр поглощения кото-
цианобактерий Synechococcus были заражены ви-
рых соответствовал поглощению хлорофилла а.
русами, выделенными и очищенными от бакте-
Отношение фикоцианина к хлорофиллу а и соот-
рий из воды залива Мэн. При этом отмечались
ношение феопигмента к хлорофиллу а оказались
сильные оптические сдвиги до и после лизиса на
хорошими показателями наблюдаемого разруше-
временных масштабах, соизмеримых с процессом
ния клеток. Спектры отражения, которые были
вирусной инфекции. В частности, обратное рас-
смоделированы с использованием индикатрисы
сеяние и поглощение (особенно в области види-
объемного рассеяния излучения, показали, что
мых длин волн) немного увеличились после зара-
БИОФИЗИКА том 66
№ 2
2021
РОЛЬ ВИРУСОВ И ВИРУСНОГО ЛИЗИСА
223
жения, но заметно уменьшились после вирусного
качестве входных данных при расчете рассеяния
лизиса. Наиболее быстрые изменения наблюда-
Ми, проводившегося для оценки эффектов дина-
лись во флуоресценции хлорофилла. Замечено
мики этих частиц в терминах относительных
было незначительное увеличение флуоресценции
вкладов различных классов размеров частиц в по-
в течение двух суток после первоначального зара-
казатели рассеяния и обратного рассеяния. Этот
жения. Такие изменения, вероятно, по мнению
анализ выявил значительное увеличение влияния
исследователей, отражают фундаментальные
агрегации частиц на рассеяние света в течение
сдвиги в фото физиологии хозяина после зараже-
вторых и третьих суток после заражения. Авторы
ния. После лизиса хозяина наблюдалось значи-
выполненного эксперимента приходят к выводу,
тельное снижение флуоресценции хлорофилла а,
что вирусный лизис бактерий и последующая на-
синхронно со значительным увеличением коли-
блюдаемая динамика частиц ведут к большим ко-
чества свободных вирусов на клетку хозяина. Эти
лебаниям их распределения в широком диапазо-
оптические изменения сопровождались и значи-
не размеров и в масштабе времени от нескольких
тельными изменениями в спектрах размеров суб-
часов до нескольких дней, и такие процессы при-
микронных частиц по мере лизиса клеток и вы-
водят к соответственно большим изменениям оп-
свобождающегося при этом их внутриклеточного
тических свойств исследуемой суспензии.
содержимого. В ходе работы для каждого образца
В одной из работ [26] изучали воздействие ви-
измеряли три оптических параметра: объемное
русного лизиса на культуру микроводоросли Mi-
рассеяние на 514 нм, флуоресценцию хлорофилла
cromonas pusilla в стадии роста. При этом опреде-
и спектральное поглощение, которое измеряли с
ляли влияния вирусного лизиса на концентра-
помощью спектрофотометра Camspec (США),
цию и состав растворенного органического
оборудованного интегрирующей сферой с одно-
углерода. Это оценивали по получению из лизата
сантиметровой кварцевой кюветой с жидкост-
прозрачных экзополимерных частиц и двух фрак-
ным образцом, размещенной в сфере и установ-
ций флуоресцентного РОВ: ароматических ами-
ленной на пути луча. Оптические эффекты вирус-
нокислот (возбуждение/эмиссия,
280/320 нм,
ной инфекции на оптическое рассеяние были
F(280/320)) и морского гуминового флуоресцент-
исследованы с помощью нефелометра Wyatt
ного РОВ (320/410 нм, F(320/410)). Концентрация
Technologies Dawn-F (США), оснащенного ар-
растворенного органического углерода увеличи-
гон-ионным лазером. Этот прибор измерял
валась в 4.5 раза быстрее, и конечная концентра-
функцию объемного рассеяния (при 514 нм) на
ция в инфицированной культуре достигала в
15 углах.
2.6 раза больших значений по сравнению с неин-
Результаты изучения временной динамики
фицированной. Производство F(280/320) и
распределения частиц по размерам и связанной с
F(320/410) было соответственно в 4.1 и 2.8 раза
этим оптической изменчивости, вызванной ви-
выше в зараженных культурах, а повышенное со-
русной инфекцией морских гетеротрофных бак-
отношение между F(280/320) и F(320/410) в лиза-
терий, в проведенном лабораторном эксперимен-
тах свидетельствовало о более высоком вкладе
те, описаны в работе [29]. Распределение частиц
лабильных (белковых) компонентов в продуци-
по размерам в диапазоне от < 50 нм до 200 нм из-
руемый вирусами РОВ, чем в водорослевых экс-
мерялось параллельно со спектральными показа-
судатах. Производство прозрачных экзополимер-
телями поглощения частиц и ослабления света,
ных частиц было в 1.8 раза быстрее и достигало
из которых был определен показатель рассеяния
уровня в 1.5 раза выше у инфицированных виру-
частицами - bp(λ).
сом клеток культуры M. pusilla в сравнении с неза-
раженными клетками. Измеренное увеличение
Так, в течение 12 ч после заражения бактери-
концентраций растворенного органического уг-
альная популяция хозяина разрушилась, числен-
лерода и прозрачных экзополимерных частиц
ность вирусов увеличилась, и при этом появились
свидетельствовало о том, что вирусный лизис
субмикронные частицы, поскольку бактерии бы-
имеет множественный и противоположный ха-
ли разрушены c высвобождением содержимого
рактер последствий для производственных и экс-
клетки, что привело к большому снижению bp(λ),
портных процессов в пелагическом океане. Та-
замеченному как почти полная очистка (просвет-
ким образом, влияя на микробную пищевую сеть
ление) смеси используемых суспензий частиц. На
путем увеличения производства как лабильного,
протяжении оставшейся части эксперимента зна-
так и рефрактерного растворенного органическо-
чительные изменения распределения частиц по
го углерода и окрашенного РОВ, вирусный лизис
размерам происходили главным образом в преде-
изменяет оптическую сигнатуру РОВ, а также
лах диапазона размеров относительно крупных
оказывает влияние на агрегацию частиц посред-
частиц (4 мм), вероятно, в результате агрегации
ством усиленного производства прозрачных эк-
частиц меньшего размера, происходящей при ли-
зополимерных частиц. Итоги вирусной активно-
зисе хозяина. Полученные результаты распреде-
сти многочисленны и имеют противоположные
ления частиц по размерам были использованы в
последствия для производства и экспортных про-
БИОФИЗИКА том 66
№ 2
2021
224
ШОЛАРЬ, СТЕПАНОВА
цессов в пелагическом (открытом) океане и по-
значимом вкладе процессов вирусного лизиса для
этому, по мнению авторов процитированной ра-
получения лучшего понимания механизмов
боты, очень важно расширять знания об относи-
функционирования океанических биогеохими-
тельно значимом вкладе этих процессов для
ческих циклов.
получения лучшего понимания механизмов
Первые результаты собственных исследований
функционирования океанических биогеохими-
по изучению роли вирусного лизиса в изменении оп-
ческих циклов.
тических характеристик черноморской воды
Таким образом, анализ информации, полу-
(2018-2019 гг.). Проведенные нами исследования
ченной при обзоре ряда публикаций зарубежных
были обусловлены отсутствием данных о роли
исследователей, посвященных изучению роли
вирусного лизиса в изменениях оптических
вирусов гидросферы в изменении оптических ха-
свойств черноморской воды при использовании
рактеристик главным образом в ходе экспери-
черноморской микробиоты (культуры микрово-
ментов в лабораторных условиях, выявил, что:
дорослей и штаммы альговирусов), при этом бы-
• вирусы, как самостоятельные частицы, явля-
ли учтены результаты, полученные в ходе иссле-
ясь самыми многочисленными среди гидробион-
дований данной проблемы зарубежными колле-
тов, тем не менее, не оказывают существенного
гами, применявшими микробиоту в основном из
влияния на оптику моря из-за их малого размера
тихоокеанского региона.
и поперечного сечения рассеяния;
Использованные в проведенных эксперимен-
• вирусный лизис представителей бактерио- и
тах культуры микроводорослей были получены из
фитопланктона приводит к многочисленным по-
коллекции живых культур микроводорослей от-
следствиям (обломки клеток, свободные кокко-
дела экологической физиологии водорослей Фе-
литы, РОВ, и т.д.), итогом которых является из-
дерального исследовательского центра «Институт
менение оптических характеристик жидкой сре-
биологии южных морей им. А.О. Ковалевского
ды их обитания, что было зафиксировано и в
РАН». Штаммы альговирусов были выделены из
экспериментах, и по спутниковым данным;
различных проб (вода, донные осадки, гидро-
• в составе материалов и методов, которые ис-
бионты), отобранных из экосистемы Черного мо-
пользуют зарубежные исследователи в экспери-
ря в акватории у побережья Севастополя. Изоля-
ментах по выявлению роли вирусов в изменении
цию альговирусов проводили при помощи запа-
оптических свойств водной среды, необходимо
тентованных авторских способов
[42,
43].
отметить применение в основном двух резервуа-
Свойства выделенных вирусов, в том числе и ана-
ров (колб, туб, полиэтиленовых мешков и других
лиз их секвенированных геномов, описаны в ра-
емкостей), заполняемых водой и биотическими
ботах [55, 56].
составляющими; один из резервуаров служит в
Первые эксперименты по изучению роли ви-
качестве контроля (рост культур бактерий, ци-
русной инфекции в изменении оптических ха-
анобактерий, микроводорослей без вирусного
рактеристик проводили с использованием уни-
лизиса), второй - в качестве опытного (с добавле-
версальной лабораторной установки, схематиче-
нием в культуру хозяина вирусной суспензии); в
ское изображение которой представлено на
ходе экспериментов используются методы и ком-
рис. 1а. Устройство созданной лабораторной
плекс приборов, необходимых для определения
установки, как и полученные при этом результа-
изменения в динамике численности вирусов и их
ты, подробно изложены в наших опубликован-
хозяев и оптических свойств исследуемой в экс-
ных работах
[35-37]. Было зафиксировано
периментах жидкой среды;
(рис. 1б) влияние вирусного лизиса (двух штам-
• изменения обратного рассеяния, обуслов-
мов черноморских альговирусов - TvV-S1 и PtV-
ленные вирусным лизисом, можно обнаружить с
S18 при контакте с культурами двух видов черно-
помощью спутниковых или авиационных мето-
морских микроводорослей - Tetraselmis viridis и
дов дистанционного зондирования, что позволит
Phaeodactylum tricornutum) на снижение значений
определять локализацию наведенной вирусами
показателя ослабления света (ПОС) по данным
гибели их хозяев;
малогабаритного спектрального измерителя по-
• вирусная инфекция оптически значимых
казателя ослабления направленного света (про-
биологических частиц бактерио- и фитопланкто-
зрачномера), разработанного в отделе оптики и
на представляет собой еще один, пока малоизу-
биофизики моря Морского гидрофизического
ченный процесс, который может быстро транс-
института РАН [57]. Однако в использованной
формировать клетки микропланктона в новые
лабораторной установке имелась лишь одна ем-
типы оптически значимого материала, в том чис-
кость, которая и служила в эксперименте в каче-
ле и в окрашенное растворенное органическое ве-
стве опыта, результаты которого представлены в
щество;
виде графика на рис. 1б, что не позволяло одно-
• в соответствии с мнением исследователей
временно получать сведения об изменениях ПОС
очень важно расширять знания об относительно
в контроле - роста и развития культур микрово-
БИОФИЗИКА том 66
№ 2
2021
РОЛЬ ВИРУСОВ И ВИРУСНОГО ЛИЗИСА
225
Рис. 1. (а) - Схема универсальной лабораторной установки контроля и мониторинга изучаемой жидкой среды [21];
(б) - изменение показателя ослабления света под действием вирусного лизиса (альговирусы TvV-S1и PtV-S18)
микроводорослей Tetraselmis viridis и Phaeodactylum tricornutum [22].
дорослей без вирусного лизиса. Для дальнейших
ны два идентичных пластиковых сосуда (диамет-
исследований и их оценки необходимо было про-
ром 28 см и высотой 30 см), в которых можно бы-
ведение экспериментов в двух емкостной уста-
ло размещать до
15 л изучаемых жидких
новке или стенде, где помимо опыта были бы
модельных биологических систем на основе мор-
условия и для контроля.
ской воды. Одна из емкостей служила для кон-
троля, другая для опыта. Апробирование создан-
В ходе экспериментов по изучению оптиче-
ских свойств водной среды (черноморской воды)
ного экспериментального лабораторного стенда
в присутствии черноморской микробиоты был
было проведено в нескольких исследованиях, в
разработан и адаптирован простой и доступный
которых в качестве модельных биологических си-
экспериментальный лабораторный стенд (рис. 2),
стем использовали как монокультуры черномор-
подробное описание которого представлено в ра-
ских микроводорослей, так и их смесь, которые
боте [38]. Для создания стенда были использова-
подвергали воздействию альговирусов в опыте,
БИОФИЗИКА том 66
№ 2
2021
226
ШОЛАРЬ, СТЕПАНОВА
Рис. 2. Схема экспериментального лабораторного
стенда для изучения изменения оптических свойств
водной среды под влиянием микробиоты и вирусного
лизиса [24]: 1 - источник искусственного освещения
(светодиодная лампа ECO A60, 11 Вт 4000 К); 2 - ра-
бочая зона, где размещается прозрачномер и датчик
температуры; 3 - контрольная емкость (модель био-
логической системы без альговирусов); 4 - емкость
для опыта (модель биологической системы с альго-
вирусом); 5 - аквариумные насосы для перемешива-
ния содержимого; 6 - светоотражающая поверх-
ность; 7 - блок регулировки интенсивности работы
насосов и передачи данных на персональный ком-
пьютер.
сравнивая показатели ПОС с контролем - без ви-
русного лизиса.
Выполненные эксперименты выявили влия-
ние черноморской микробиоты на оптические
свойства водной среды - наблюдалось повыше-
ние ПОС в результате роста культур микроводо-
рослей (в контроле) и его снижение вследствие
вирусного лизиса культур (в опыте), что пред-
ставлено на рис. 3 в виде графиков изменения ве-
личин ПОС в динамике. Результаты, полученные
на примере одной из черноморских культур под
воздействием альговируса в опыте и без вирусно-
го лизиса в контроле, подробно описаны в работе
[39]. Как можно видеть из графиков на рис. 3в, в
инфицированной культуре в первые сутки опыта
ПОС выше, что отчасти не противоречит и дан-
ным экспериментов, описанных в работе [25].
При этом между данными измерений ПОС на
длине волны 625 нм и численностью клеток куль-
туры микроводоросли, используемой в этом экс-
перименте, с учетом контроля и опыта была
Рис. 3. Изменение показателя ослабления света в смеси
выявлена тесная связь, описанная линейным
морской воды и культур микроводоросли Tetraselmis vir-
регрессионным уравнением с высоким коэффи-
idis (а), смеси микроводорослей Tetraselmis viridis и
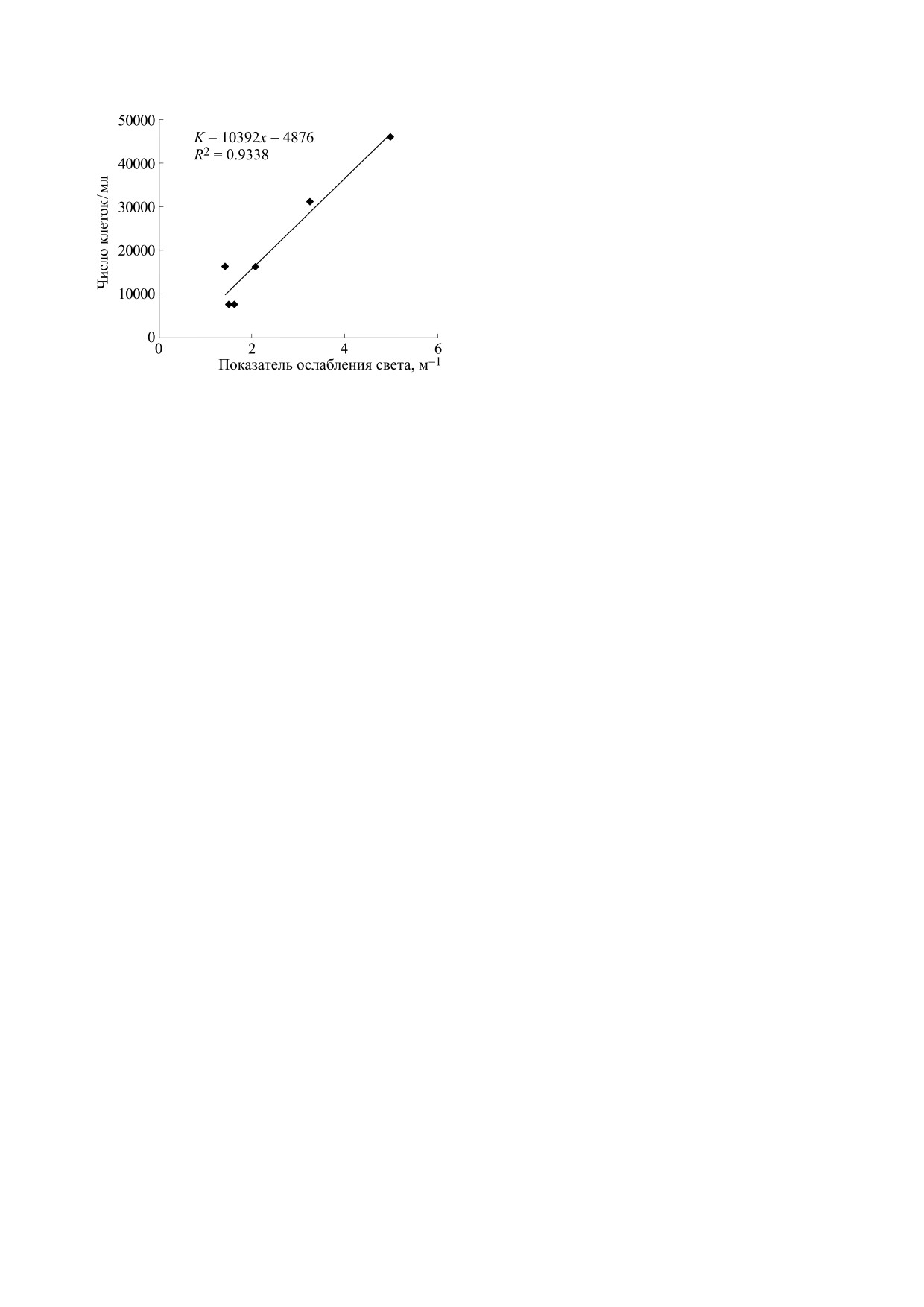
циентом детерминации (рис. 4).
Dunaliella tertiolecta (б), микроводоросли Dunaliella viridis
(в) с добавлением вирусной суспензии альговирусов
Таким образом, полученные результаты явля-
TvV-S1 (а, б) и DvV-SI1 (в) в опыте и в контроле. Экспе-
ются первыми в плане экспериментального изу-
рименты выполнены в 2019 г. [24].
чения роли черноморской микробиоты и вирус-
ного лизиса в изменении оптических характери-
стик черноморской воды по данным ПОС и с
Возможное практическое применение альговиру-
учетом изменения численности клеток хозяев
сов и вирусного лизиса. Известно, что вирусы ши-
черноморских альговирусов.
роко используются в практике нанотехнологий
БИОФИЗИКА том 66
№ 2
2021
РОЛЬ ВИРУСОВ И ВИРУСНОГО ЛИЗИСА
227
нов, а также и для снижения явления обрастания
в замкнутых местах их содержания (бассейнах).
Перспективно использование альговирусов
и вирусного лизиса для профилактики и/или
устранения явления «цветения воды» в различ-
ных водоемах (бассейнах, водохранилищах, кана-
лах, городских бухтах), что улучшит их эстетиче-
ский вид.
Однако все представленные предположения
для использования альговирусов и вызываемого
ими вирусного лизиса нуждаются в дополнитель-
ных исследованиях, экспериментах и практиче-
ских изучениях, что одновременно будет являться
и расширением знаний в области молодой науки -
морской (водной) вирусологии.
Рис. 4. Линейная зависимость значений ПОС и числен-
ности клеток культуры микроводоросли Dunaliella viridis
ЗАКЛЮЧЕНИЕ И ВЫВОДЫ
на длине волны 625 нм по данным контроля (без вируса)
и опыта (с вирусом DvV-SI1) [24].
Проведенный литературный обзор доступной
информации о роли вирусов гидросферы, самых
многочисленных и наименее изученных среди
[58]. Так, в одной из публикаций [59] приведены
гидробионтов, в изменении оптических свойств
современные данные о применении вирусов в ка-
воды водоемов, с учетом собственных результа-
честве эффективных инструментов в нанобиотех-
тов, полученных в ходе экспериментов с исполь-
нологии и наномедицине (биочипы, латексные ча-
зованием черноморской микробиоты, выявил
стицы, рекомбинантные вирусы-химеры, сорбен-
следующее.
ты вирусов на основе полианилина и наноалмазов
1. Самые многочисленные среди гидробион-
и др.). Описано их использование для фундамен-
тов вирусы водоемов как самостоятельные части-
тальных исследований и в различных сферах нано-
цы по мнению исследователей не оказывают су-
технологий, включая электронику.
щественного влияния на оптические свойства во-
ды, что объясняют их нано размером и малым
С учетом полученных данных как в экспери-
поперечным сечением рассеяния.
ментах зарубежных коллег, так и в наших иссле-
дованиях логично предположить, что альговиру-
2. По данным экспериментов зарубежных кол-
сы и вирусный лизис могут использоваться в про-
лег при использовании как опытных, так и кон-
филактике и/или в борьбе с такими широко
трольных емкостей вирусный лизис представите-
распространенными явлениями в гидросфере,
лей бактерио- и фитопланктона приводит к мно-
как биокоррозия и биообрастание, где основную
гочисленным последствиям, итогом которых
роль играют и микроводоросли.
является изменение оптических характеристик
жидкой среды. Эти изменения при экстраполя-
По нашему мнению, возможно применение
ции на природные водоемы было предложено
альговирусов и вызываемого ими вирусного ли-
определять дистанционно как показатели наве-
зиса в марихозяйствах в качестве «природного
денной (вызванной) вирусами гибели их одно-
фермента» для расщепления (трансформации)
клеточных хозяев.
живого органического вещества на легко усваива-
3. Вирусная инфекция оптически значимых
емый коктейль макро- и микромолекулярной ор-
биологических частиц бактерио- и фитопланкто-
ганической субстанции [60]. В таком виде РОВ
на представляет собой еще один пока малоизу-
поглощается и утилизируется различными гидро-
ченный процесс, который может быстро транс-
бионтами, что способствует их росту и развитию
формировать клетки микропланктона в новые
[3, 60-63]. Это свойство альговирусов может ис-
типы оптически значимого материала. Причем
пользоваться в технологиях для растворения по-
это свойство может быть использовано в практи-
средством лизиса культур микроводорослей с це-
ке, в том числе и в нанотехнологиях.
лью выделения из них белков, ферментов, вита-
4. По мнению ученых, назрела актуальность и
минов и др. биологически активных веществ без
необходимость расширения знания о вкладе про-
их повреждения.
цессов вирусного лизиса в физико-химико-био-
Альговирусы и вызываемый ими вирусный ли-
логическую составляющую гидросферы для по-
зис, с нашей точки зрения, можно применять в
лучения лучшего понимания механизмов функ-
методиках по обработке кожи (при обрастании
ционирования океанических биогеохимических
микроводорослями) обитаемых в неволе дельфи-
циклов.
БИОФИЗИКА том 66
№ 2
2021
228
ШОЛАРЬ, СТЕПАНОВА
5. Сведений, особенно в работах отечествен-
10. В. В. Дрюккер, Н. В. Дутова, А. С. Ковадло и
ных ученых, о роли вирусного лизиса в измене-
Т. Я. Косторнова, Изв. ИрГУ. Науки о Земле 1 (1),
189 (2008).
нии оптических характеристик водной среды в
гидросфере на уровне экспериментов или поле-
11. В. В. Дрюккер, Н. В. Дутова и А. С. Ковадло, Докл.
РАН 440 (2), 282 (2011).
вых исследований недостаточно, и они не учиты-
12. Н. В. Дутова и В. В. Дрюккер, Гидробиол. журн. 45
ваются в теории и практике оптики моря.
(4), 82 (2009).
Дальнейшие исследования по изучению роли
13. Н. В. Дутова и В. В. Дрюккер, Докл. РАН 450 (4),
черноморской микробиоты в изменении физиче-
468 (2013).
ских характеристик морской среды будут направ-
14. А. И. Копылов, Журн. Сиб. фед. ун-та. Биология 6
лены на использование в экспериментах более
(4), 354 (2013).
широкого спектра черноморских альговирусов (в
15. А. И. Копылов, Д. Б. Косолапов и Е. А. Заботкина,
авторской коллекции имеются альговирусы к се-
Микробиология 76 (6), 879 (2007).
ми видам микроводорослей), как и круга их хозя-
16. А. И. Копылов, Д. Б. Косолапов и Е. А. Заботкина,
ев. В опытах предполагается использовать влия-
Биол. внутренних вод, № 1, 49 (2008).
ние таких физических факторов, как магнитное и
17. А. И. Копылов, Д. Б. Косолапов, Е. А. Заботкина и
электрическое поля, ультрафиолетовое облуче-
В. Страшкрабова, Изв. РАН. Сер. биол., № 6, 661
ние и некоторые другие абиотические воздей-
(2010).
ствия.
18. А. И. Копылов, Д. Б. Косолапов и Е. А. Заботкина,
Микробиология 80 (2), 241 (2011).
19. А. И. Копылов, А. Ф. Сажин, Е. А. Заботкина и
ФИНАНСИРОВАНИЕ РАБОТЫ
Н. Д. Романова, Океанология 55 (4), 620 (2015).
Работа выполнена в рамках госбюджетной те-
20. А. И. Копылов, Д. Б. Косолапов и Е. А. Заботкина,
мы ФИЦ МГИ РАН (тема № 0827-2019-0001,
Поволжский экологич. журн., № 2, 128 (2017).
№ госрегистрации АААА-А18-118012690119-7) и
21. О. А. Степанова, Системы контроля окружающей
Государственного задания ИПТС (тема № 0012-
среды, № 12 (32), 99 (2018).
2019-0003,
№ госрегистрации АААА-А19-
22. С. А. Шоларь и М. Е. Ли, Системы контроля окру-
119040590054-4).
жающей среды, № 14 (34), 17 (2018).
23. W. M. Balch, J. M. Vaughn, J. F. Novotny, et al., Lim-
nol. Oceanogr. 45 (2), 492 (2000).
КОНФЛИКТ ИНТЕРЕСОВ
24. W. M. Balch, J. M. Vaughn, J. F. Novotny, et al., Lim-
Авторы заявляют об отсутствии конфликта
nol. Oceanogr. 47 (5), 1554 (2002).
интересов.
25. W. M. Balch, J. M. Vaughn, J. I. Goes, et al., Limnol.
Oceanogr. 52 (2), 727 (2007).
26. C. Lønborg, M. Middelboe, and C. P. D. Brussaard,
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
Biogeochemistry 116 (1-3), 231 (2013).
Настоящая работа не содержит описания ис-
27. S. G. H. Simis, M. Tijdens, H. L. Hoogveld, et al., J.
следований с использованием людей и животных
Plankton Res. 27 (9), 937 (2005).
в качестве объектов.
28. S. G. H. Simis, M. Tijdens, H. L. Hoogveld, et al.,
Limnol. Oceanogr. 52, 184 (2007).
29. J. Uitz, D. Stramski, A. C. Baudoux, et al., Limnol.
СПИСОК ЛИТЕРАТУРЫ
Oceanogr. 55, 2317 (2010).
1. К. С. Шифрин, Введение в оптику океана (Гидро-
30. В. И. Маньковский, Морской гидрофизич. журн.
метеоиздат, Л., 1983).
5, 78 (1999).
2. D. Stramski, A. Bricaud, and A. Morel, Appl. Optics 40
31. В. И. Маньковский, Морской гидрофизич. журн.
(18), 2929 (2001).
5, 14 (2011).
3. K. E. Wommack and R. R. Colwell, Microbiol. Mol.
32. V. I. Man’kovsky and E. V. Man’kovskaya, Phys.
Biol. R. 64 (1), 69 (2000).
Oceanography 1, 41 (2017).
4. M. Middelboe and C. Brussaard, Viruses, 9 (10), 302
33. Р. П. Тренкеншу, А. С. Лелеков, П. Е. Гаврилов
(2017).
и др., Актуальные вопросы биологической физики
5. C. A. Suttle, Nature Rev. Microbiol. 5, 801 (2007).
и химии 1 (1), 77 (2016).
6. J. McN. Sieburth, V. Smetacek, and J. Lenz, Limnol.
34. В. В. Александров, Электрофизика пресных вод
Oceanogr. 23 (6), 1256 (1978).
(Гидрометеоиздат, Л., 1985).
7. В. В. Дрюккер и Н. В. Дутова, Докл. РАН 410 (6),
35. О. А. Степанова и П. В. Гайский, Системы контро-
847 (2006).
ля окружающей среды 11 (31), 48 (2018).
8. В. В. Дрюккер и Н. В. Дутова, Докл. РАН 427 (2),
36. О. А. Степанова, П. В. Гайский и С. А. Шоларь,
277 (2009).
Системы контроля окружающей среды 13 (33), 19
9. В. В. Дрюккер, Н. В. Дутова, А. С. Ковадло и др., в
(2018).
сб. Проблемы устойчивого функционирования водных
37. С. А. Шоларь, П. В. Гайский и О. А. Степанова, в
и наземных экосистем (Изд-во РГУ, Ростов-на-До-
кн. Мезомасштабные и субмезомасштабные процес-
ну, 2006), сс. 118-121.
сы в гидросфере и атмосфере, под ред. Ю. В. Воро-
БИОФИЗИКА том 66
№ 2
2021
РОЛЬ ВИРУСОВ И ВИРУСНОГО ЛИЗИСА
229
бьева (Изд. Инст. океанол. им. П. П. Ширшова
49. O. A. Stepanova, Y. A. Solovyova, and A. V. Solovyov,
РАН, М., 2018), сс. 410-412.
Ukr. Bioorg. Acta 2, 53 (2011).
38. С. А. Шоларь, О. А. Степанова и Л. В. Стельмах,
50. M. S. Petro, I. V. Agarkova, and T. M. Petro, J. Neuro-
Системы контроля окружающей среды 2 (36), 13
immunol. 297, 46 (2016).
(2019).
51. R. H. Yolken, L. Jones-Brando, and D. D. Dunigan,
39. С. А. Шоларь, О. А. Степанова, Л. В. Стельмах
Proc. Natl. Acad. Sci. USA 111 (45), 16106 (2014).
и др., в кн. Современные проблемы оптики есте-
52. E. Organelli, H. Claustre, A. Bricaud, et al., Geophys.
ственных вод (Химиздат, Спб., 2019), сс. 156-160.
Res.-Oceans. 122 (5), 3542 (2017).
40. А. Е. Крисс, Морская микробиология (глубоковод-
53. D. Stramski and C. D. Mobley, Limnol. Oceanogr. 42,
ная) (АН СССР, М., 1959).
538 (1997).
41. А. Е. Крисс и Е. А. Рукина, ДАН СССР, 57 (8), 833
54. W. H. Wilson, G. A. Tarran, and D. Schroeder, J. Mar.
(1947).
Biol. Assoc. UK 82 (3), 369 (2002).
42. О. А. Степанова, Патент Украины на изобретение
55. O. A. Stepanova, A. L. Boĭko, and I. S. Shcherbatenko,
№ 2003065499, Бюл. № 4 (2003).
Mikrobiolohich. Zh. 75 (5), 76 (2013).
43. О. А. Степанова, Патент Украины на изобретение
56. O. A. Stepanova, Russ. J. Mar. Biol. 42 (2), 123 (2016).
№ 201003881, Бюл. № 2 (2010).
57. М. Е. Ли, Системы контроля окружающей среды
44. Е. В. Лихошвай, Наука из первых рук 70 (4), 88
17, 7 (2012).
(2016).
58. P. Singh, M. Gonzales, and M. Manchester, Drug De-
45. N. Guixa-Boixereu, C. Pedros-Alio, J. I. Calderón-
velop. Res. 67 (1), 23 (2006).
Paz, et al., in Ecology of Marine Viruses (CIESM Work-
59. Л. В. Урываев, С. В. Альховский, Е. И. Самохвалов
shop Monographs, Banyuls, 2003), pp. 41-44.
и др. Вопросы вирусологии 1, 52 (2012).
46. S. Wilhelm, M. Weinbauer and C. Suttle, Manual of
60. O. A. Stepanova, Russ. J. Mar. Biol. 43 (2), 127 (2017).
Aquatic Viral Ecology (ASLO, Waco, 2010).
61. E. Beretta and Y. Kuang, Math. Biosci. 149 (1), 57
47. Proposal for SCOR WG to investigate the role of virus-
(1998).
es in marine ecosystems, in Proc. Sci. Comm. Ocean.
62. I. Hewson, J. M. O’Neil, C. A. Heil, et al., Aquat. Mi-
Res. (Baltimore, 2005), v. 40, pp. 66-70 (Annex 4).
crob. Ecol. 25 (1), 1 (2001).
48. P. Hyman and T. Stephen, Viruses of Microorganisms
63. A. G. Murray and P. M. Eldridge, J. Plankton. Res. 16
(Caister Acad. Press, Ashland, 2018).
(6), 627 (1994).
The Role of Viruses and Viral Lysis in Alteration of the Optical Properties of Aquatic
Environment as the Habitat of Viruses
S.A. Sholar* and O.A. Stepanova**
*Marine Hydrophysical Institute, Russian Academy of Sciences, Каpitanskaya ul. 2, Sevastopol, 299011 Russia
**Institute of Natural and Technical Systems, Russian Academy of Sciences, ul. Lenina 28, Sevastopol, 299011 Russia
This review has its focus on the role of hydrosphere viruses and hydrosphere virus-induced viral lysis of sin-
gle-celled hosts in alteration of the optical properties of water. Our earlier research showed that viruses of res-
ervoirs as representatives of the nano-sized fraction of microplankton have no significant effect on the optical
properties of water. However, further experiments conducted by foreign researchers and our current study in-
clude reports that viral lysis of the representatives of bacterio- and phytoplankton leads to numerous effects
ultimately altering the optical characteristics of the aquatic environment as the habitat of viruses. It is as-
sumed that the discovered phenomenon can be used for practical purposes.
Keywords: sea optics, hydrosphere viruses and their hosts, viral lysis, light beam attenuation coefficient
БИОФИЗИКА том 66
№ 2
2021