БИОФИЗИКА, 2021, том 66, № 2, с. 248-258
МОЛЕКУЛЯРНАЯ БИОФИЗИКА
УДК 51-76; 577.214
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ ПОВЕДЕНИЯ
ТРАНСКРИПЦИОННОГО ПУЗЫРЯ В ПЛАЗМИДЕ pPF1
И ЕЕ МОДИФИКАЦИЯХ. СВЯЗЬ МЕЖДУ ЭНЕРГЕТИЧЕСКИМ
ПРОФИЛЕМ ПЛАЗМИДЫ И НАПРАВЛЕНИЕМ ТРАНСКРИПЦИИ
© 2021 г. А.А. Гриневич, И.С. Масулис, Л.В. Якушевич
Институт биофизики клетки РАН - обособленное подразделение ФИЦ «Пущинский научный центр биологических
исследований РАН», 142290, Пущино Московской области, Институтская ул., 3
E-mail: grin_aa@mail.ru
Поступила в редакцию 06.11.2020 г.
После доработки 26.01.2021 г.
Принята к публикации 29.01.2021 г.
C использованием нелинейного уравнения синус-Гордона и его решения - кинка - моделировали
и изучали динамику транскрипционного пузыря в плазмиде pPF1. На основе рассчитанного энер-
гетического профиля плазмиды pPF1 и ее модификаций было показано, что минимальная потенци-
альная энергия формирования кинка или нуклеации транскрипционного пузыря соответствует об-
ласти между генами белков Egfp и mCherry. Вставка в область между Egfp и mCherry однородных по-
следовательностей показала, что кинк с большей вероятностью активируется в polyT или polyC по
сравнению с polyA или polyG, что свидетельствует о зависимости нуклеации транскрипционного пу-
зыря от физической массы оснований. В случае вставок в область между Egfp и mCherry небольших
фрагментов природной последовательности Escherichia coli модель позволяет выявить нити ДНК с
наибольшей вероятностью нуклеации транскрипционного пузыря и соответственно определить на-
правление (в сторону гена Egfp или mCherry) транскрипции, что говорит о связи между направле-
нием транскрипции и энергетическим профилем плазмиды.
Ключевые слова: математическое моделирование, нелинейное уравнение синус-Гордона, кинк,
солитоны, плазмида pPF1, ДНК, транскрипционный пузырь.
DOI: 10.31857/S0006302921020058
Плазмиды представляют собой кольцевые мо-
Математическое моделирование с использова-
лекулы ДНК. Они являются удобными объектами
нием методов нелинейной математики и теорети-
для искусственных манипуляций с генетическим
ческой физики - один из наиболее простых и эф-
материалом. Векторная плазмида pPF1 - типич-
фективных инструментов изучения динамики
ный представитель этого класса биологических
сложных биологических систем. Постановка и
объектов (рис. 1). Она была получена из плазми-
реализация компьютерных экспериментов, заме-
ды pET-28b [1] и дополнена двумя генами, коди-
няющих и дополняющих эксперименты с реаль-
рующими зеленый и красный флуоресцентные
ными системами, все более широко и активно
белки Egfp и mCherry [2]. Данный вектор, как ре-
осуществляется исследователями из-за целого
портерная конструкция, позволяет одновремен-
ряда преимуществ: они дешевле, быстрее и обла-
но регистрировать транскрипцию в двух направ-
дают предсказательными возможностями. В на-
лениях, инициированную на участках последова-
стоящей работе методы математического модели-
тельности между генами флуоресцентных белков.
рования применяются для изучения особенно-
Начало процесса транскрипции определяется
стей динамики транскрипционного пузыря в
взаимодействием РНК-полимеразы с промотор-
плазмиде pPF1 и предсказания с их помощью - в
ными областями ДНК. В результате происходит
какой области плазмиды с наибольшей вероятно-
локальное расплетание двойной спирали ДНК,
стью произойдет нуклеация транскрипционного
формирование открытого транскрипционного
пузыря и в каком направлении пойдет процесс
комплекса (транскрипционного пузыря) протя-
транскрипции.
женностью ~ 13 пар оснований (п.о.) и последую-
щее движение этого пузыря вдоль молекулы ДНК
Сокращениe: п.о. - пары оснований.
в процессе элонгации транскрипции [3].
248
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ
249
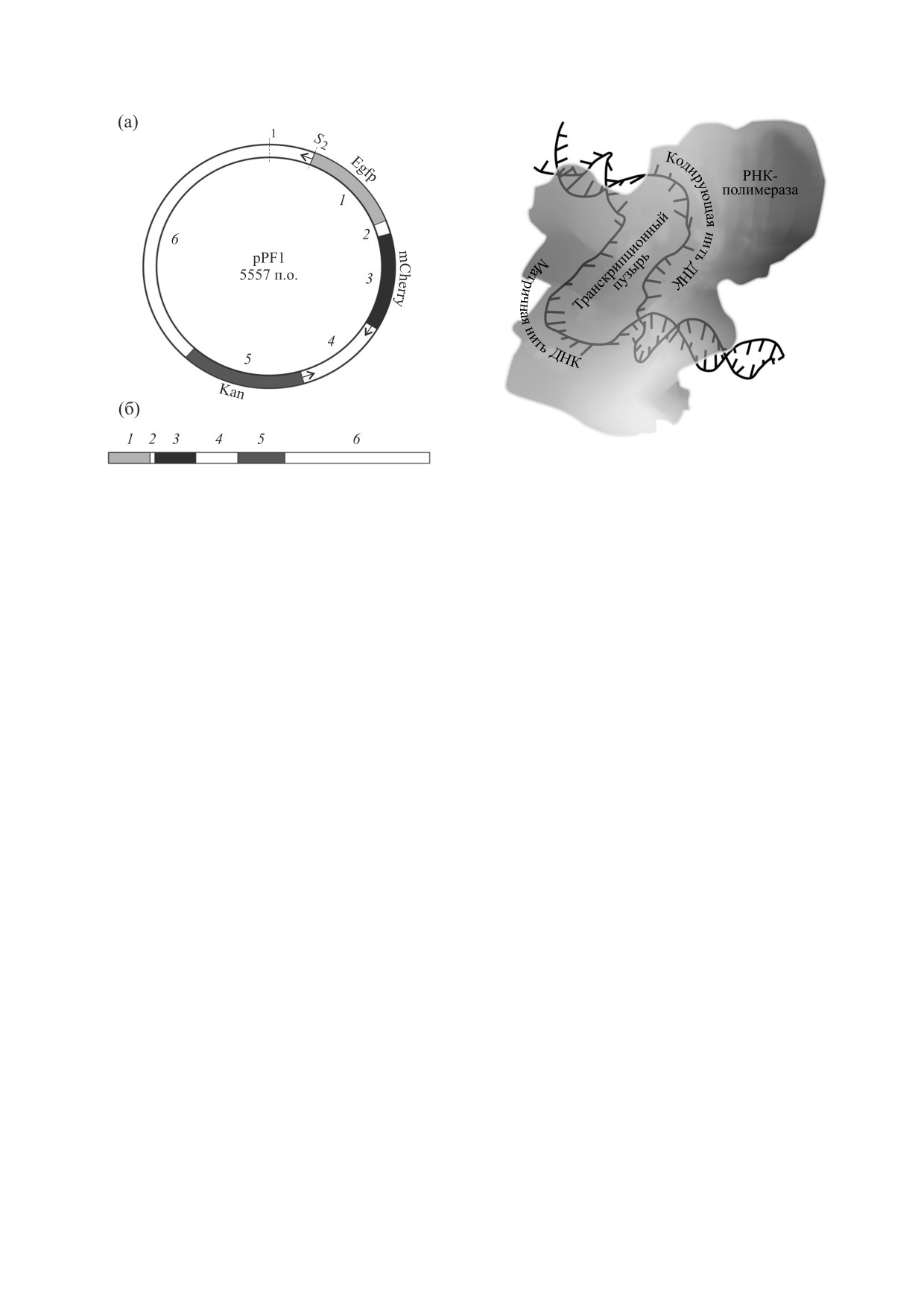
S
Рис. 2. Транскрипционный пузырь.
Рис. 1. Кольцевая (a) и линейная (б) схемы плазмиды
pPF1. Ген зеленого белка Egfp (участок 1) и ген устой-
чивости к канамицину Kan (участок 5) транскрибиру-
структуры двойной спирали с учетом механохи-
ются по комплементарной нити. Ген красного белка
мических свойств.
mCherry (участок 3) транскрибируется по основной
нити. S1 - точка, указывающая начало последова-
Одним из инструментов, используемых для
тельности; S2 - точка, указывающая начало первого
математического моделирования конформаци-
участка.
онной подвижности ДНК, является аппарат не-
линейной математики и, в частности, свойства
нелинейного уравнения синус-Гордона и его од-
Направление синтеза РНК задается конфигу-
носолитонных решений - кинков [10, 11]. Движе-
рацией транскрипционного пузыря и ориентаци-
ние кинков можно рассматривать как движение
ей матричной и нематричной нитей в полости
квазичастиц в потенциальном поле молекулы.
РНК-полимеразы (рис. 2), а скорость элонгации
Ранее движение транскрипционного пузыря в
зависит от нуклеотидной последовательности
плазмиде pTTQ18 моделировалось нами как дви-
транскрибируемого участка [4]. Кинетика фор-
жение кинка в потенциальном поле этой плазми-
мирования пузыря и скорость его движения вдоль
ды [12]. Поведение кинка в такой модели полно-
молекулы ДНК в процессе транскрипции явля-
стью определялось энергетическим профилем
ются ключевыми факторами, обусловливающи-
потенциального поля плазмиды. Аналогичный
ми вариацию в уровне экспрессии различных ге-
математический аппарат применялся для расчета
нов в диапазоне 10000 раз [5]. Механо-динамиче-
потенциального поля плазмиды pBR322 в работе
ские и энергетические свойства нуклеотидов
[13]. В настоящей работе мы используем этот ап-
играют важную роль при формировании тран-
парат для расчета энергетического профиля плаз-
скрипционного пузыря. Известно, что это много-
миды pPF1 и различных ее модификаций, полу-
стадийный процесс, сопровождающийся образо-
ченных путем интегрирования в область, распо-
ванием промежуточных состояний и вращатель-
ложенную между двумя генами белков Egfp и
ной динамикой отдельных оснований [6-9].
mCherry, однородных последовательностей и не-
больших фрагментов Escherichia coli, взятых из так
Внутренняя (конформационная) лабильность
называемых промоторных островов - областей,
ДНК и ее способность реагировать на точечные
высоко насыщенных промотороподобными
возмущения в упорядоченной структуре могут
участками [14]. Цель данной работы - рассчитать
рассматриваться в качестве физической основы
энергетические профили плазмиды pPF1 и ее мо-
для реализации ее основной функции - хранения
дификаций и использовать полученные результа-
и экспрессии генетической информации. С этой
ты для предсказания поведения кинка, что в свою
точки зрения одним из наиболее перспективных
очередь позволит сделать выводы о том, в какой
направлений является использование математи-
области плазмиды с наибольшей вероятностью
ческого моделирования конформационной по-
произойдет нуклеация транскрипционного пузы-
движности кольцевых ДНК для предсказания ди-
ря и в каком направлении (в сторону гена Egfp
намического поведения локальных возмущений
или mCherry) пойдет процесс транскрипции.
БИОФИЗИКА том 66
№ 2
2021
250
ГРИНЕВИЧ и др.
Рис. 3. Последовательность кольцевой плазмиды pPF1. Ген зеленого белка Egfp отмечен светло-серым маркером, ген
красного белка mCherry - черным маркером, ген устойчивости к канамицину Kan - темно-серым маркером, сайт
узнавания эндонуклеазы рестрикции BglII - жирным белым шрифтом на темно-сером фоне.
БИОФИЗИКА том 66
№ 2
2021
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ
251
СТРУКТУРА ПОСЛЕДОВАТЕЛЬНОСТИ
ПЛАЗМИДЫ PPF1
Полная последовательность нуклеотидов
плазмиды pPF1 приведена на рис. 3. Общая длина
последовательности равна 5557 п.о. Она содержит
три гена: ген зеленого белка Egfp с координатами
(333-1049), помеченный на рис. 3 светло-серым
маркером, ген красного белка mCherry с коорди-
Рис. 4. Схематическое изображение последователь-
натами (1134-1841), помеченный черным марке-
ности второго участка. Q - точка, указывающая ме-
ром, и ген устойчивости к канамицину Kan с ко-
сто, куда будут встроены небольшие модельные по-
ординатами (2569-3381), помеченный темно-се-
следовательности в компьютерных экспериментах.
рым маркером. Жирным шрифтом белого цвета
на темно-сером фоне показан сайт узнавания эн-
донуклеазы рестрикции BglII (AGATCT) с коор-
Как и в экспериментах по искусственным ге-
нетическим манипуляциям in vitro, в наших ком-
динатами (1089-1094).
пьютерных экспериментах мы будем встраивать
Кроме генов белков, последовательность плаз-
во второй участок небольшие модельные после-
миды pPF1 содержит три промежуточные участ-
довательности. Сначала для отработки метода в
ка. Назовем их вторым (1050-1133), четвертым
качестве модельных будут использованы одно-
(1842-2568) и шестым участками (3382-5557) +
родные последовательности: polyA, polyT, polyG,
+ (1-332).
polyC, а затем - небольшие фрагменты последо-
вательности Escherichia coli.
Основное внимание при генетических мани-
пуляциях уделяется второму участку, так как он
расположен между генами белков Egfp и mCherry
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ
и имеет удобный для манипуляций нуклеотид-
КОНФРМАЦИОННОЙ ПОДВИЖНОСТИ
ный состав. На рис. 4 показана структура этого
МОЛЕКУЛЫ ДНК
участка. В центре последовательности находится
Для математического описания конформаци-
сайт узнавания эндонуклеазы рестрикции BglII
онной подвижности некоторого фрагмента моле-
AGATCT длиной 6 п.о. Слева и справа от него на-
кулы ДНК, которое включало бы возможность
ходятся короткие последовательности длиной
образования открытых состояний, воспользуемся
39 п.о. каждая, причем короткая последователь-
системой из 2N связанных нелинейных диффе-
ность справа от рестриктазного сайта является
ренциальных уравнений, имитирующих угловые
обратно комплементарной к короткой последо-
отклонения азотистых оснований в обеих поли-
вательности слева от рестриктазного сайта.
нуклеотидных цепочках [15]:
2
d
φ t)
n,1
'
I
n,1
–
K
n,1
[
φ
n+1,1
t) –
2φ
n,1
t) +φ
n-1,1
(t)]+
2
dt
+
k
n,1-2
R
n,1
(R
n,1
+
R
n,2
) sinφ
n,1
–
k
n,1-2
R
n,1
R
n,2
sin(φ
n,1
–
φ
n,2
)=
(1)
d
φ
t)
n,1
=-β
n,1
+
M
0
,
dt
2
d
φ
n
,2
t)
'
I
n
,2
-
K
n,2
[
φ
n
+1,2
t)
- 2φ
n,2
t)
+ φ
n
-1,2
(t)] +
2
dt
+
k
R
(
R
+
R
)
sinφ
–
k
R
R
sin(φ
– φ
) =
(2)
n,1-2
n
,2
n
,1
n
,2
n,2
n
,1-2
n,1
n
,2
n,2
n
,1
d
φ
n,2
t)
= -β
n
,2
+
M
0
dt
Здесь φn,i(t)
- угловое отклонение n-го
характеризующая жесткость (на растяжение)
азотистого основания i-й цепочки; In,i - момент
2
сахаро-фосфатного остова;
β
=αR
;
α
-
n,i
n,i
инерции n-го азотистого основания i-й цепочки;
коэффициент диссипации; kn,i
- константа,
Rn,i - расстояние от центра масс n-го азотистого
характеризующая
взаимодействие
между
основания i-й цепочки до сахаро-фосфатного
основаниями внутри пар; M0
- постоянный
2
остова;
K'
n,i
=KR
n,i
;
K
- константа,
внешний момент; i = 1, 2; n =1, 2, … N, N - число
БИОФИЗИКА том 66
№ 2
2021
252
ГРИНЕВИЧ и др.
пар оснований в рассматриваемом фрагменте.
нить коэффициенты In,i, Rn,i, K'n,i, kn,1-2 и βn,i ме-
Уравнения (1) и (2) упрощаются, если усред-
тодом концентраций [16]:
I
n,i
→
I
i
=
I
CA A,i
+
I
T T
C
,i
+
I
CG G,i
+
I
CC C,i
,
R
n,i
→
R
i
=
R
C
A A,i
+
R
T T
C
,i
+
R
CG G,i
+
R
CC C
,
i
,
'
'
'
'
'
'
K
n,i
→
K
i
=
K
CA A,i
+
K
CT T,i
+
K
CG G,i
+
K
CC C,i
(3)
k
n,1-2
→
k
1-2
=
k
A-
(
T A
C
,1
+
C
T,2
)
+
k
G-
(
C G
C
,1
+C
C,2
),
,
,
,
,
,
,
n
i
i
A A
i
T T
i
G G
i
C C
i
β
→β
=β
C
+β
C
+β
C
+β
C
где Cj,i = Nj,i/N - концентрация оснований j-го количество азотистых оснований j-го типа в i-й
последовательности.
типа (j = А, Т, G, С) в i-й цепи (i =1, 2); Nj,i -
После проведения процедуры усреднения (3)
уравнения (1) и (2) преобразуются к виду:
2
d
ϕ
n,1
t)
'
I
1
−
K
1
[
ϕ
n+1,1
t)
−
2ϕ
n,1
t)
+ϕ
n−1,1
(t)]+
2
dt
+
k
R
(
R
+
R
) sin
ϕ
−
k
R
R
sin(
ϕ
−ϕ
)
=
(4)
1-2
1
1
2
n,1
1−2
1
2
n,1
n,2
d
ϕ
n
,1
t)
= -β
1
+
M
0
,
dt
2
d
ϕ
t)
,2
'
n
I
−
K
[
ϕ
t)
−
2
ϕ
t)
+ϕ
(t)]+
2
2
1,2
,2
1,2
2
n
n
n
+
−
dt
+
k
1-2
R
2
(
R
1
+
R
2
) sin
ϕ
n,2
−
k
1−2
R
1
R
2
sin(
ϕ
n,2
−ϕ
n
,1
)
=
(5)
d
ϕ
t)
n
,2
= -β
2
+
M
0
dt
В континуальном приближении эти уравнения приобретают следующий вид:
'
2
I
1
ϕ
1tt
−
K
1
a
ϕ
1zz
+
k
1-2
R
1
(
R
1
+
R
2
) sin
ϕ
1
−
k
1−2
R
1
R
2
sin(
ϕ
1
−ϕ
2
)
=
(6)
= -β
ϕ
+
M
,
1
1
t
0
'
2
I
ϕ
−
K
a
ϕ
+
k
R
(
R
+
R
) sin
ϕ
−
k
R
R
sin(
ϕ
−ϕ
)
=
2
2tt
2
2
zz
1-2
1
1
2
2
1−2
1
2
2
1
(7)
= -β
2
ϕ
2
t
+
M
0
С помощью приближения среднего поля [17] два связанных уравнения (6) и (7) трансформируются
в два независимых уравнения:
'
2
2
I
1
ϕ
1
tt
−
K
1
a
ϕ
1
zz
+
k
1-2
R
1
sin
ϕ
1
= -β
1
ϕ
1
t
+
M
0
,
(8)
'
2
2
I
ϕ
−
K
a
ϕ
+
k
R
sin
ϕ
= -β
ϕ
+
M
(9)
2
2
tt
2
2
zz
1-2
2
1
2
2t
0
Односолитонные решения этих уравнений - кинки:
ϕ
k
,1
(
z
,
t
)
=
4arctg
{
exp
[(
γ
k
,1
/
d
k
,1
)(
z
−υ
k
,1
t
−
z
0,1
)]}
,
(10)
ϕ
k
,2
(
z
,
t
)
=
4arctg
{
exp
[(
γ
k,2
/
d
k
,2
)(
z
−υ
k
,2
t
−
z
0,2
)]}
Кинки используют для имитации открытых
'
2
1/2
C
i
=
(
K
i
a
/
I
i
)
- скорость звука в i-й цепочке,
состояний
[18,
19]. Здесь υk,i
- скорость
'
2
1/2
движения кинка в i-й цепочке (i
=1,
2),
d
=
(
K
a
/
V
)
- размер кинка в i-й цепочке,
k
,
i
i
i
2
2
−1/2
2
γ
=
(1−υ
/
C
)
- фактор Лоренца,
V
=
k
R
k,i
k,i
i
i
1-2
i
БИОФИЗИКА том 66
№ 2
2021
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ
253
Минимальные значения энергии кинка E0,i, рассчитанные для шести участков основной и комплементарной
последовательностей плазмиды pPF1
E0,1
E0,2
Номер участка
Координаты участка, п.о.
(10-17 Дж)
(10-17 Дж)
1 (Egfp)
333-1049
0.1639
0.1658
2
1050-1133
0.1566
0.1566
3 (mCherry)
1134-1841
0.1673
0.1673
4
1842-2568
0.1585
0.1585
5 (Kan)
2569-3381
0.1596
0.1596
6
(3382-5557) + (1-332)
0.1631
0.1631
Полная энергия кинка в i-й цепочке Ei опреде-
формула (12) для расчета минимальных значений
ляется следующей формулой [20]:
энергии E0,i, необходимых для образования кинка
в каждом из шести участков i-й цепочки ДНК
E
0,i
(i = 1,2): в трех участках, отвечающих генам (Egfp,
E
i
=
,
(11)
2
mCherry, Kan) и трех участках, отвечающих про-
υ
k,i
1-
межуточным областям (второй, четвертый и ше-
2
Ci
стой участки). Результаты расчета приведены в
где E0,i - минимальное значение энергии, необ-
таблице. Построенные энергетические профили
для основной и комплементарной последователь-
ходимое для образования кинка:
ностей плазмиды pPF1 приведены на рис. 5.
'
E
0,i
=
8
K
Vi i
, (i =
1, 2).
(12)
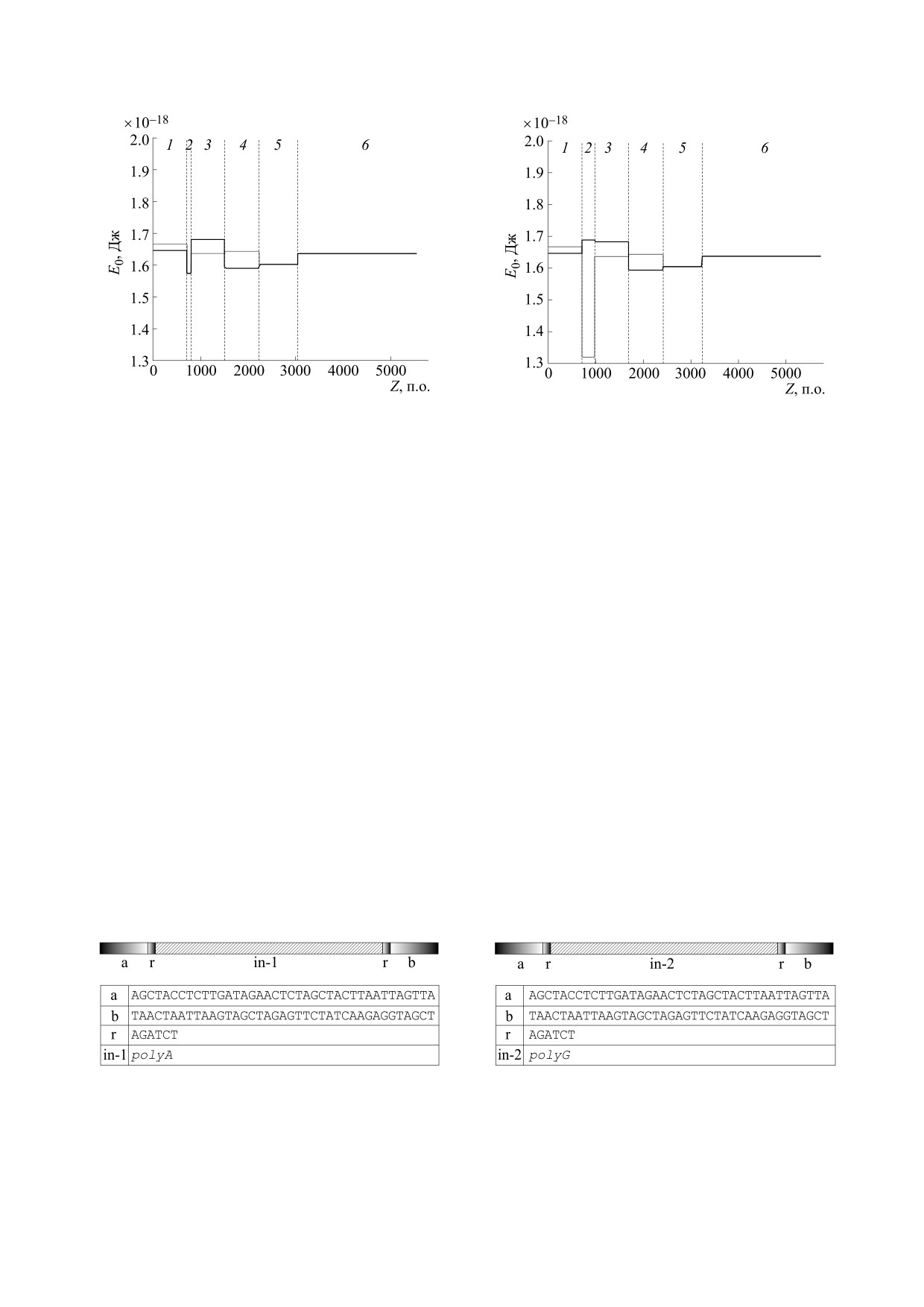
Из рис. 5 видно, что самая глубокая потенци-
альная яма наблюдается во втором участке и ос-
В случае малых скоростей (υk,i <<Ci) форму-
новной (черная кривая), и комплементарной (се-
ла (11) приобретает вид:
рая кривая) последовательностей. Это говорит о
том, что вероятность образования кинка во вто-
2
E
0,i
m
k,i
υ
k,i
ром участке выше, чем в других. Видно также, что
E
=
=
E
+
(13)
i
0,
i
2
во втором участке черная и серая кривые совпада-
2
υ
k,i
ют. Это означает, что энергии активации кинка в
1-
2
C
основной и в комплементарной последователь-
i
ностях одинаковые. Таким образом, с энергети-
E
0,i
ческой точки зрения кинку все равно, в какой из
где
m
k
,i
=
- масса кинка в i-й цепочке.
2
2C
двух этих последовательностей активироваться.
i
Формула (13) свидетельствует о том, что кинк
Какого-либо предпочтения нет.
можно моделировать как квазичастицу,
обладающую массой mk,i, скоростью υk,i и
ЭНЕРГЕТИЧЕСКИЕ ПРОФИЛИ
обладающую потенциальной энергией E0,i. В
МОДИФИЦИРОВАННЫХ ПЛАЗМИД
следующем разделе мы воспользуемся
квазиоднородным приближением и формулой
Компьютерные эксперименты с однородными
(12) для расчета потенциальной энергии кинка
последовательностями, интегрированными в плаз-
миду pPF1. В первом компьютерном эксперимен-
внутри каждого из участков. .
те плазмида pPF1 модифицировалась вставкой во
второй участок однородной модельной последо-
ЭНЕРГЕТИЧЕСКИЙ ПРОФИЛЬ
вательности polyA длиной 183 п.о. Схематическое
ПЛАЗМИДЫ PPF1
изображение последовательности второго участ-
ка со вставкой представлено на рис. 6.
При построении энергетического профиля
плазмиды pPF1 использовались значения коэф-
Результаты расчета энергетических профилей
фициентов уравнений (1) и (2) из работы [12] и
основной и комплементарной последовательно-
БИОФИЗИКА том 66
№ 2
2021
254
ГРИНЕВИЧ и др.
Рис. 7. Энергетический профиль основной (черная
Рис. 5. Энергетический профиль основной (черная
кривая) и комплементарной (серая кривая) последо-
кривая) и комплементарной (серая кривая) последо-
вательностей модифицированной плазмиды pPF1 с
вательностей плазмиды pPF1. Цифрами 1-6 обозна-
однородной последовательностью polyA, интегриро-
чены участки последовательностей. Вертикальные
ванной во второй участок. Цифрами 1-6 обозначены
пунктирные линии обозначают границы между
участки. Вертикальные пунктирные линии обознача-
смежными участками.
ют границы между смежными участками.
стей модифицированной плазмиды представле-
грированной в нее последовательностью polyG
ны на рис. 7.
представлены на рис. 9.
Из рис. 7 видно, что во втором участке компле-
Из рис. 9 видно, что самая глубокая потенци-
ментарной последовательности polyT (серая кри-
альная яма наблюдается во втором участке ком-
вая) наблюдается самая глубокая потенциальная
плементарной последовательности polyC (серая
яма, а в том же участке основной последователь-
кривая). В этом же участке основной последова-
тельности polyG (черная кривая), напротив, на-
ности polyA (черная кривая), напротив, наблюда-
ется барьер. Это говорит о том, что вероятность
блюдается барьер. Таким образом, наибольшая
активации кинка во втором участке комплемен-
вероятность локального расплетания ДНК будет
наблюдаться в комплементарной последователь-
тарной последовательности выше, чем в том же
ности второго участка.
участке основной последовательности.
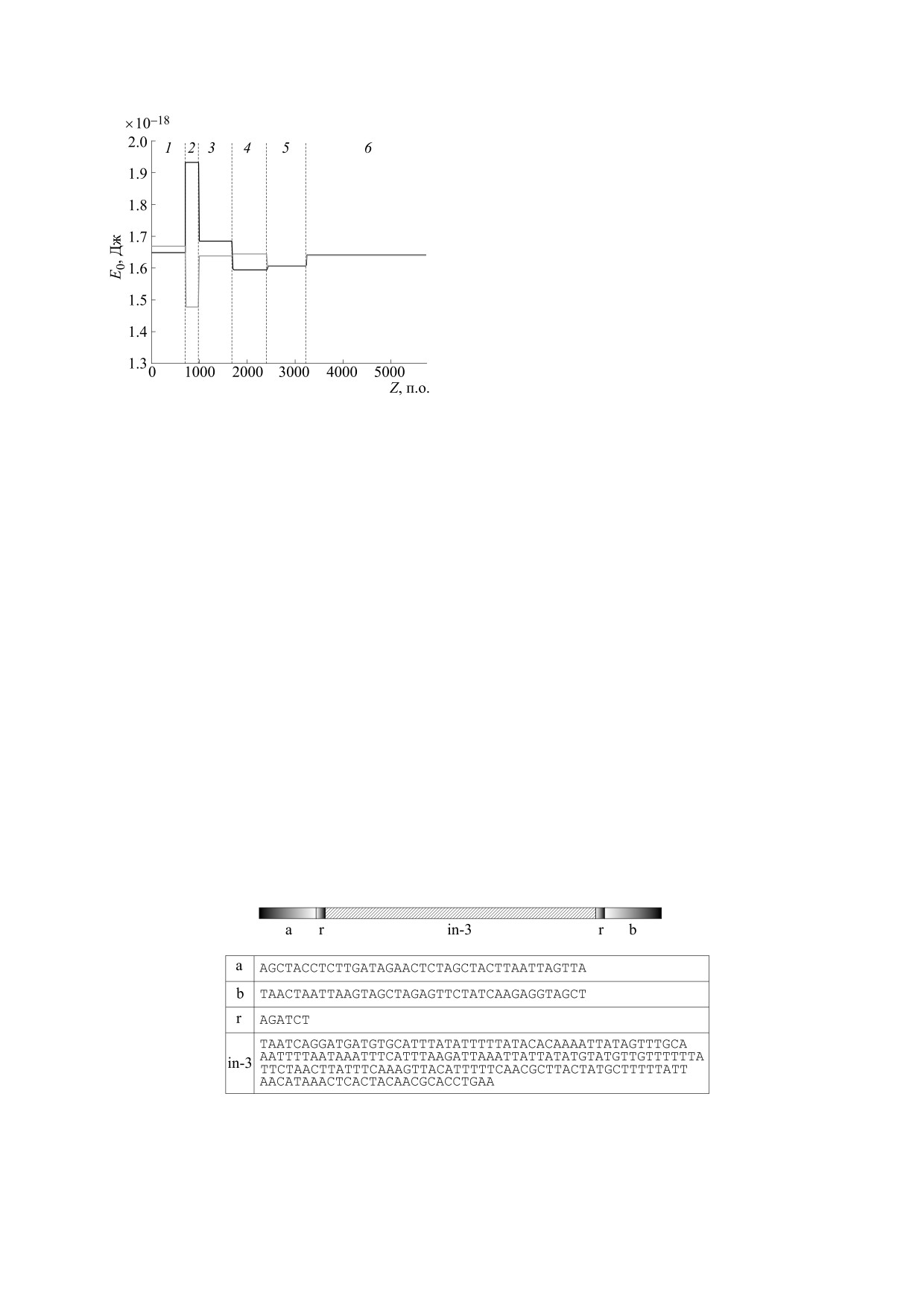
Во втором компьютерном эксперименте плаз-
Компьютерные эксперименты с неоднородными
последовательностями, интегрированными в плаз-
мида pPF1 модифицировалась вставкой во вто-
миду pPF1. В следующем компьютерном экспери-
рой участок другой однородной последователь-
менте плазмида pPF1 модифицировалась встав-
ности (polyG) той же длины (рис. 8).
кой во второй участок небольшого фрагмента ре-
Результаты расчета энергетических профилей
альной последовательности Escherichia coli,
основной и комплементарной последовательно-
взятого из так называемых промоторных остров-
стей модифицированной плазмиды pPF1 с инте-
ков - областей высоко насыщенных промотор-
Рис. 8. Схематическое изображение последователь-
Рис. 6. Схематическое изображение последователь-
ности второго участка плазмиды pPF1 с интегриро-
ности второго участка с интегрированной в него по-
ванной в него модельной однородной последователь-
следовательностью polyA.
ностью polyG.
БИОФИЗИКА том 66
№ 2
2021
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ
255
ния пузыря: от 5'- к 3'-концу. Такое движение со-
ответствует движению в направлении гена крас-
ного белка. Можно ожидать, что в данном случае
преимущественно будет синтезироваться белок
mCherry.
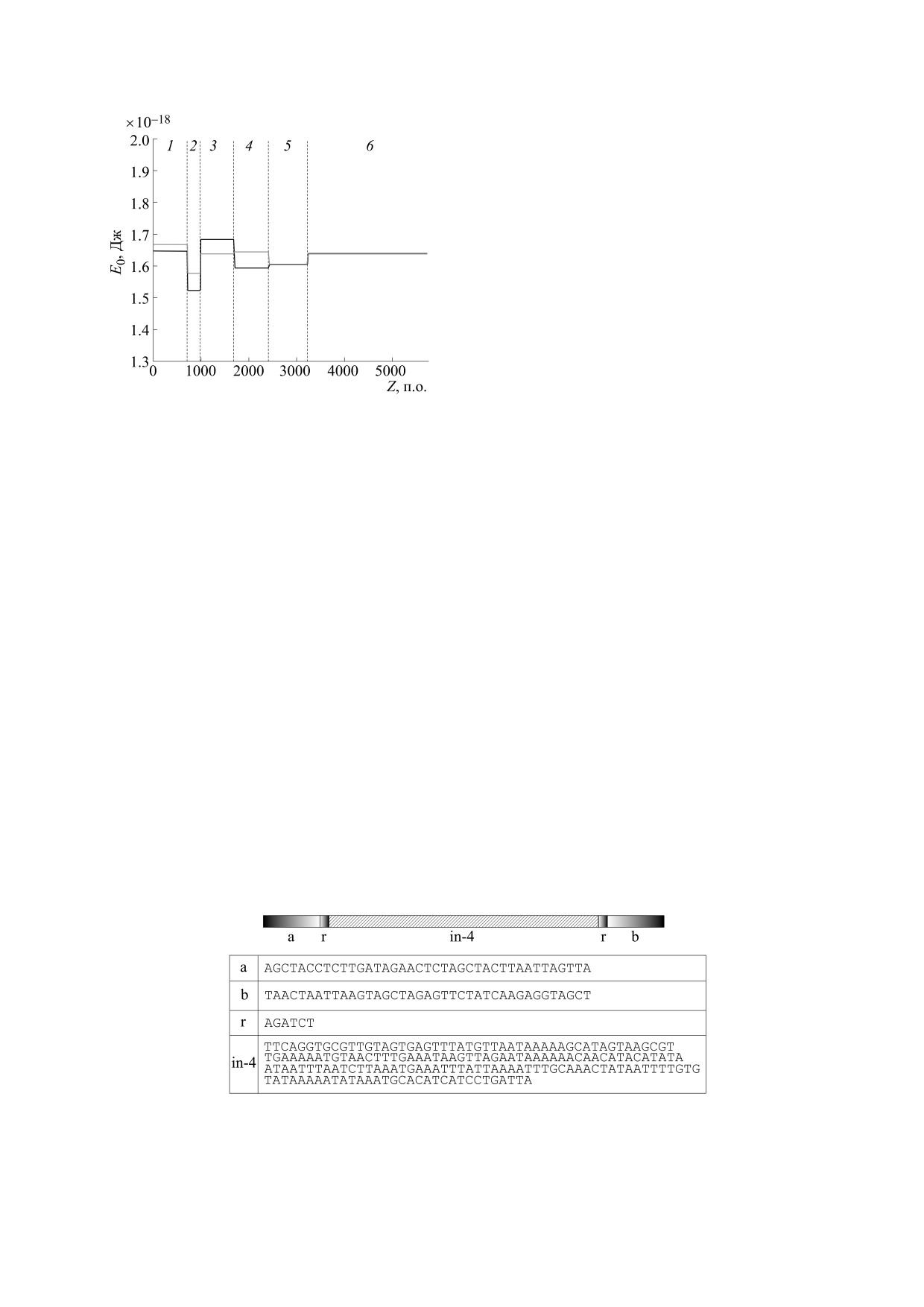
В следующем компьютерном эксперименте во
второй участок плазмиды pPF1 был встроен фраг-
мент последовательности обратно комплемен-
тарной к последовательности фрагмента, кото-
рый встраивался во второй участок в предыдущем
эксперименте. Последовательность второго
участка, полученная в результате такой модифи-
кации, показана на рис. 12, а на рис. 13 представ-
лены результаты построения энергетического
профиля модифицированной плазмиды.
Из рис. 13 видно, что в отличие от предыдуще-
го случая самая глубокая потенциальная яма при-
Рис. 9. Энергетический профиль основной (черная
ходится на второй участок комплементарной по-
кривая) и комплементарной (серая кривая) последо-
вательностей модифицированной плазмиды pPF1, с
следовательности (серая кривая). Таким образом,
однородной последовательностью polyG, интегриро-
с более высокой вероятностью кинк будет акти-
ванной во второй участок. Цифрами 1-6 обозначены
вироваться не в основной, а в комплементарной
участки. Вертикальные пунктирные линии обознача-
последовательности второго участка. С учетом
ют границы между смежными участками.
того, что движение пузыря всегда происходит от
5'- к 3'-концу, можно ожидать, что с большей ве-
подобными участками [14]. На рис. 10 представ-
роятностью пузырь будет двигаться в направле-
нии гена зеленого белка, для которого в данной
лена схема второго участка после модификации.
конструкции нематричной нитью является ком-
Результаты расчета энергетического профиля
плементарная последовательность. Таким обра-
основной и комплементарной последовательно-
зом, вероятность синтеза белка Egfp будет выше
стей модифицированной вышеуказанным спосо-
по отношению к mCherry.
бом плазмиды pPF1 представлены на рис. 11.
Из рис. 11 видно, что самая глубокая потенци-
альная яма приходится на второй участок основ-
ОБСУЖДЕНИЕ И ВЫВОДЫ
ной последовательности (черная кривая). Это го-
ворит о том, что наибольшая вероятность актива-
В данной работе с помощью методов нелиней-
ции кинка будет наблюдаться во втором участке
ной математики были рассчитаны энергетиче-
основной последовательности, соответствующей
ские профили потенциальных полей плазмиды
нематричной нити ДНК. В данном случае это
pPF1 и ее модификаций. Модификации плазми-
можно трактовать следующим образом: именно
ды получали вставкой во второй участок, распо-
второй участок ответственен за разрушение водо-
ложенный между генами зеленого и красного
родных связей и нуклеацию транскрипционного
белков, небольших фрагментов однородных
пузыря. Развитие нуклеации на основной после-
(синтетических) и неоднородных (природных)
довательности указывает на направление движе-
последовательностей.
Рис. 10. Схематическое изображение последовательности второго участка плазмиды pPF1 с интегрированным в него
первым фрагментом реальной последовательности Escherichia coli (in-3).
БИОФИЗИКА том 66
№ 2
2021
256
ГРИНЕВИЧ и др.
основаниями внутри пар играет ключевую роль в
процессе образования локально расплетенного
участка. Чем легче основание, тем легче происхо-
дит нуклеация.
Были рассчитаны энергетические профили
модифицированных плазмид с интегрированны-
ми во второй участок двумя небольшими фраг-
ментами реальной последовательности Escherich-
ia coli, содержащими промоторные участки. Ис-
ходя из вида полученных профилей, удалось
предсказать, в какой области плазмиды с наи-
большей вероятностью произойдет нуклеация
транскрипционного пузыря и в каком направле-
нии (в сторону гена Egfp или mCherry) пойдет
процесс транскрипции. Это говорит о том, что в
модельном эксперименте приоритетное направ-
ление транскрипции, а следовательно, и роль ни-
Рис. 11. Энергетический профиль основной (черная
тей ДНК в качестве матричной или нематричной
кривая) и комплементарной (серая кривая) последо-
обусловлены энергетическими характеристика-
вательностей модифицированной плазмиды pPF1,
ми вставляемой последовательности, независимо
где во второй участок интегрирован небольшой фраг-
от ее ориентации по отношению к генам репор-
мент последовательности Escherichia coli (in-3). Циф-
рами 1-6 обозначены участки последовательностей.
терных белков. Эта определяющая роль нуклео-
Вертикальные пунктирные линии обозначают грани-
тидов нематричной нити имеет эксперименталь-
цы между смежными участками.
ное [7, 21-23] и теоретическое [24] подтвержде-
ние.
При моделировании энергетического профи-
Показано, что во всех случаях самая глубокая
ля однородных и природной последовательно-
потенциальная яма наблюдается на втором участ-
ке. Это говорит о том, что этот участок наиболее
стей, использованных в данной работе, компле-
ментарные нити ДНК различались по своей по-
предпочтителен для образования кинка (или нук-
тенциальной способности к активации кинка.
леации транскрипционного пузыря). В случае не-
Асинхронный характер дестабилизации двойной
модифицированной плазмиды кинк с равной ве-
спирали и преимущественная лабильность пири-
роятностью активируется в обоих (в основной и в
мидинов при взаимодействии с бактериальными
комплементарной) последовательностях. В слу-
хеликазами были подтверждены эксперимен-
чае с интегрированной в плазмиду последова-
тальными методами [25].
тельностью polyA кинк с большей вероятностью
активируется в комплементарной последователь-
Полученные результаты показали, что исполь-
ности polyT. В случае интегрированной в плазми-
зованная математическая модель - удобный и
ду последовательности polyG кинк с большей ве-
эффективный инструмент для исследования ди-
роятностью активируется в комплементарной по-
намических характеристик открытых состояний в
следовательности polyC. Это свидетельствует о
молекуле ДНК, частным случаем которых являет-
важной роли такого физического параметра, как
ся транскрипционный пузырь [3]. Однако модель
масса основания, которая вместе с водородными
обладает рядом ограничений и приближений. В
взаимодействиями между комплементарными
ней рассматриваются только вращательные дви-
Рис. 12. Второй участок плазмиды с интегрированной в него вторым фрагментом реальной последовательности
Escherichia coli (in-4).
БИОФИЗИКА том 66
№ 2
2021
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ
257
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
Настоящая работа не содержит описания ис-
следований с использованием людей и животных
в качестве объектов.
СПИСОК ЛИТЕРАТУРЫ
resources/plasmid-files/?set=pet_and_duet_vectors_
(novagen)&plasmid=pET-28b(%2B) (дата обраще-
ния: 01.11.2020).
2. I. S. Masulis, Z. Sh. Babaeva, S. V. Chernyshov, et al.,
Sci. Rep. 5, 11449 (2015).
3. S. Borukhov and E. Nudler, Trends Microbiol. 16 (3),
126 (2008).
4. J. Y. Kang, T. V. Mishanina, R. Landick, et al., J. Mol.
Biol. 431 (20), 4007 (2019).
5. M. T. Jr. Record, W. S. Reznikoff, M. L. Craig, et al., in
Рис. 13. Энергетический профиль основной (черная
Escherichia coli and Salmonella Cellular and Molecu-
кривая) и комплементарной (серая кривая) последо-
lar Biology, Ed. by F.C. Neidhardt (ASM Press, Wash-
вательностей модифицированной плазмиды pPF1,
ington, DC, USA, 1996), pp. 792-821.
где во второй участок интегрирован небольшой фраг-
мент последовательности Escherichia coli (in-4). Циф-
6. H. Boyaci, J. Chen, R. Jansen, et al., Nature 565
рами 1-6 обозначены участки последовательностей.
(7739), 382 (2019).
Вертикальные пунктирные линии обозначают грани-
7. J. Chen, C. Chiu, S. Gopalkrishnan, et al., Mol Cell. 78
цы между смежными участками.
(2), 275 (2020).
8. E. F. Ruff, M. T. Jr. Record, and I. Artsimovitch. Bio-
molecules 5 (2), 1035 (2015).
жения и не учитываются продольные, попереч-
9. J. T. Winkelman, I. O. Vvedenskaya, Y. Zhang, et al.,
ные и радиальные динамические степени свобо-
Science 351 (6277), 1090 (2016).
ды нуклеотидов. Используется приближение
10. P. J. Caudrey, J. C. Eilbeck, and J. D. Gibbon, Nuovo
среднего поля, при котором явная динамика ос-
Cimento B 25 (2), 497 (1975).
нований одной нити описывается в среднем поле,
11. V. G. Ivancevic and T. T. Ivancevic, J. Geometry and
формируемом комплементарной нитью. Не учи-
Symmetry in Physics 31, 1 (2013).
тывается спиральность молекулы ДНК, и не рас-
12. A. A. Grinevich, A. A. Ryasik, and L. V. Yakushevich,
сматривается сам процесс инициации и образо-
Chaos, Solitons & Fractals 75, 62 (2015).
вания транскрипционного пузыря, который фор-
13. L. V. Yakushevich and L.A. Krasnobaeva, Math. Biol.
мируется при взаимодействии РНК-полимеразы
Bioinform. 14 (1), 327 (2019).
с ДНК. Не смотря на эти ограничения модели,
14. K. S. Shavkunov, I. S. Masulis, M. N. Tutukina, et al.,
она позволяет на основе расчета энергетических
Nucl. Acids Res. 37 (15), 4919 (2009).
профилей рассматриваемых молекул ДНК сде-
15. L. A. Krasnobaeva and L. V. Yakushevich, J. Bioin-
лать качественную оценку вероятности нуклеа-
form. Comput. Biol. 13 (1), 1540002 (2015).
ции пузыря и направления его движения, резуль-
16. L. V. Yakushevich and L. A. Krasnobaeva, Int. J. Non-
таты которой согласуются с экспериментальны-
linear Mech. 43, 1074 (2008).
ми данными.
17. L. V. Yakushevich, J. Biol. Phys. 43, 113 (2017).
18. W. Englander, N. R. Kallenbach, A. J. Heeger, et al.,
Результаты моделирования показали, что эф-
Proc. Natl. Acad. Sci. USA 77, 7222 (1980).
фективность транскрипции в заданном направ-
19. Л. В. Якушевич, Л. А. Краснобаева, А. В. Шапова-
лении синтеза РНК зависит от энергии актива-
лов и др., Биофизика 50, 450 (2005).
ции кинка в нематричной нити. Полученные
20. Л. В. Якушевич и А. А. Рясик, Компьютерные ис-
данные свидетельствуют о целесообразности
следования и моделирование 4 (1), 209 (2012).
применения расчетного метода на основе аппара-
21. Y. Zhang, Y. Feng, S. Chatterjee, et al., Science 338
та нелинейной динамики для прогнозирования
(6110), 1076 (2012).
биологических характеристик природных после-
22. E. Heyduk, K. Kuznedelov, K. Severinov, et al. J. Biol.
довательностей ДНК.
Chem. 281 (18), 12362 (2006).
23. M. E. Karpen and P. L. deHaseth, Biomolecules 5 (2),
668 (2015).
КОНФЛИКТ ИНТЕРЕСОВ
24. N. S. Roy, S. Debnath, A. Chakraborty, et al., Phys.
Chem. Chem. Phys. 20 (14), 9449 (2018).
Авторы заявляют об отсутствии конфликта
25. F. Colizzi, C. Perez-Gonzalez, R. Fritzen, et al., Proc.
интересов.
Natl. Acad. Sci. USA 116 (45), 22471 (2019).
БИОФИЗИКА том 66
№ 2
2021
258
ГРИНЕВИЧ и др.
Mathematical Modeling of the Behavior of the Transcription Bubble
in the pPF1 Plasmid and Its Modified Versions.
A link between the Plasmid Energy Profile and the Direction of Transcription
A.A. Grinevich, I.S. Masulis, and L.V. Yakushevich
Institute of Cell Biophysics, Russian Academy of Sciences,
Institutskaya ul. 3, Pushchino, Moscow Region, 142290 Russia
In this study, we use the kink solutions of the nonlinear sine-Gordon equation to apply the methods of math-
ematical modeling to investigate the dynamics of the transcription bubble in the pPF1 plasmid. Based on the
calculated energy profile for the pPF1 plasmid and its modified versions, it was shown that the minimum po-
tential energy of the kink formation or transcription bubble nucleation corresponds to the region between the
genes of the Egfp and mCherry proteins. The insertion of homogeneous sequences into the region between
Egfp and mCherry showed that the kink is more likely to be activated in polyT or polyC as compared to polyA
or polyG, which indicates the dependence of nucleation of the transcription bubble on the molecular weight
of base pairs. In the case of insertions into the region between Egfp and mCherry of small fragments of the
native sequence of Escherichia coli, the model identifies DNA strands with the highest probability of nucle-
ation of the transcription bubble, and, accordingly, determines the direction (towards the Egfp or mCherry
gene) of transcription, indicating a link between the direction of transcription and the energy profile of the
plasmid.
Keywords: mathematical modeling, nonlinear sine-Gordon equation, kink, solitons, pPF1 plasmid, DNA, tran-
scription bubble
БИОФИЗИКА том 66
№ 2
2021