БИОФИЗИКА, 2021, том 66, № 2, с. 271-276
БИОФИЗИКА КЛЕТКИ
УДК 577.23 + 576.311
ТРАНСПОРТ ГЛИЦЕРИНА В МИТОХОНДРИИ
© 2021 г. А.И. Даль*,
**
Н.Л. Векшин
*Пущинский государственный естественно-научный институт,
142290, Пущино Московской области, просп. Науки, 3
**Институт биофизики клетки РАН - обособленное подразделение ФИЦ «Пущинский научный центр биологических
исследований РАН», 142290, Пущино Московской области, Институтская ул., 3
E-mail: aleksandrina.dal@mail.ru
Поступила в редакцию 18.05.2020 г.
После доработки 04.09.2020 г.
Принята к публикации 06.12.2020 г.
Рассмотрен вопрос о поступлении молекул глицерина (в нефосфорилированной форме) внутрь ми-
тохондрий. С помощью спектрофлуориметрии и ИК-Фурье-спектроскопии продемонстрировано
эффективное связывание глицерина с митохондриальными мембранами из печени крысы. Метода-
ми полярографии и турбидиметрии показано, что в малых концентрациях (0.1-0.4 мМ) глицерин
повышает проницаемость внутренней мембраны для субстратов - НАДН и сукцината, а в больших
концентрациях сильно повреждает мембраны. Глицерин в малых концентрациях усиливает потреб-
ление кислорода интактными митохондриями (в изотонической среде) на НАДН или на сукцинате,
а в больших - угнетает. В гипотонической среде глицерин сразу начинает подавлять НАДН-дегид-
рогеназу, но не сукцинатдегидрогеназу. Полученные данные позволяют заключить, что глицерин
может сам по себе напрямую транспортироваться (пассивно по градиенту) внутрь митохондрий, а
не только в форме глицерофосфата. Предполагается, что глицерин, взаимодействуя с липидами и
белками, формирует во внутренней мембране каналы для проникновения себя и субстратов.
Ключевые слова: митохондрии, глицерин, потребление кислорода, НАДН-дегидрогеназа, сукцинатде-
гидрогеназа, дыхательная цепь
DOI: 10.31857/S0006302921020071
Общепринято, что глицерин транспортирует-
внутрь митохондрий. Выяснение этих вопросов и
ся внутрь митохондрий в форме альфа-глицеро-
было целью данной работы.
фосфата, образующегося в цитоплазме [1]. В
печени имеется фермент глицеролкиназа, ката-
МАТЕРИАЛЫ И МЕТОДЫ
лизирующий в цитозоле фосфорилирование гли-
церина в глицерофосфат, который проникает в
Фракцию митохондрий из печени крысы вы-
митохондрии и превращается в 3-фосфоглицери-
деляли по стандартной методике с небольшими
новый альдегид. Внутри митохондрий имеется
модификациями [2], все растворы охлаждали на
своя фосфоглицериндегидрогеназа, отдающая
льду. Печень крысы помещали в 40 мл ледяной
электроны и протоны в дыхательную цепь, что
среды выделения, содержащей 250 мМ сахарозы,
сопровождается синтезом двух молекул АТФ.
0.5 мМ ЭГТА и 10 мМ HEPES (pH 7.5), после чего
Значительная часть глицерина, образующегося
ее продавливали через пресс, добавляли 40 мл той
при гидролизе липидов, используется для их ре-
же среды и затем проводили гомогенизацию в
синтеза [1].
стеклянном гомогенизаторе с тефлоновым пести-
Ранее в нашей лаборатории было обнаружено,
ком. Полученный гомогенат центрифугировали
что некоторое количество добавленного глицери-
10 мин при 1000 g для осаждения неразрушенных
клеток. Осадок отбрасывали, а супернатант цен-
на ингибирует сукцинат-зависимое дыхание ми-
тохондрий (хотя мало влияет на шунтирующие
трифугировали 20 мин при 3500 g (тяжелая фрак-
реакции переноса электронов к искусственным
ция митохондрий). Полученный осадок осторож-
акцепторам) [2]. Представляет интерес подробнее
но гомогенизировали в 5-8 мл среды выделения и
изучить способность глицерина влиять на актив-
затем брали для опытов (хранили на льду и ис-
пользовали в течение нескольких часов).
ность дегидрогеназ дыхательной цепи. Главное -
важно понять, может ли глицерин сам по себе (в
В полярографических опытах изотоническая
нефосфорилированном виде) проникать извне среда инкубации митохондрий содержала 150 мМ
271
272
ДАЛЬ, ВЕКШИН
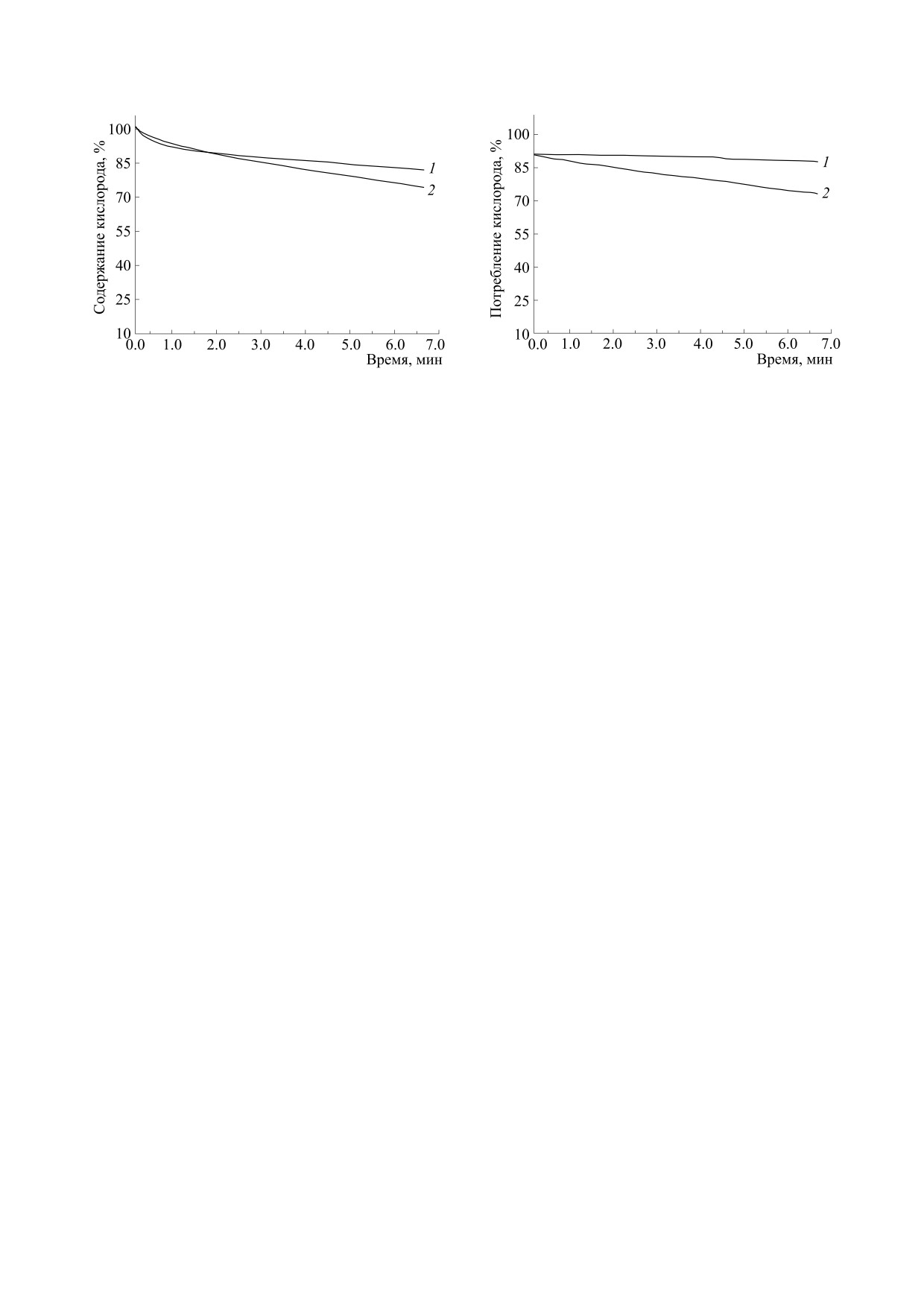
Рис. 1. Потребление кислорода в суспензии печеноч-
Рис. 2. Потребление кислорода в суспензии митохон-
ных митохондрий (2 мг /мл) в изотонической среде
дрий в изотонической среде при добавке 1 мМ сукци-
при добавке 1 мМ НАДН (1) и то же в присутствии
ната (1), то же самое в присутствии 400 мкМ глицери-
400 мкМ глицерина (2). По оси ординат - содержа-
на (2).
ние кислорода в процентах (100% соответствуют
250 мкМ в воде при нормальном атмосферном давле-
нии).
гих реагентов, мешающих измерениям в средней
ИК-области.
сахарозы и
10 мМ трис-фосфатного буфера
Спектры триптофановой флуоресцении бел-
(pH 7.0). Гипотоническая среда инкубации содер-
ков разбавленных суспензий митохондрий (0.1-
жала только 40 мМ хлорида натрия (pH 7.0).
0.3 мг/мл) регистрировали на спектрофлуоримет-
ре SLM-4800 (SLM Inc., США) при возбуждении
Измерение потребления кислорода суспензи-
на 286 нм в кварцевых микрокюветах с длиной
ей митохондрий в герметичной ячейке объемом
оптического пути 0.4 см.
2.6 мл при перемешивании (в изотонической или
гипотонической среде при добавлении сукцината
или НАДН) проводили полярографическим ме-
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
тодом с использованием термооксиметра «Экс-
Скорость дыхания изолированных митохон-
перт-001» (ООО «Эконикс», Россия) с электро-
дрий лимитируется в первую очередь проницае-
дом Кларка.
мостью внутренней мембраны для добавляемых
Светорассеяние митохондрий измеряли тур-
субстратов дыхательной цепи [4]. Внутренняя
бидиметрически при 700 нм в кювете с длиной
мембранная НАДН-дегидрогеназа, активный
оптического пути 1 см на спектрофотометре «ПЭ-
центр которой обращен в матрикс, слишком мед-
5400УФ» (ООО «ПромЭкоЛаб», Санкт-Петер-
ленно окисляет НАДН, добавленный к свежевы-
бург) по оптической плотности мутной суспен-
деленным нативным митохондриям в изотониче-
зии. Концентрацию митохондриального белка
ской среде, именно из-за своей низкой проница-
определяли на том же приборе по оптической
емости (нет никакой
«внешней» НАДН-
плотности триптофановых остатков при 286 нм
дегидрогеназы в наружной мембране [2, 4]). Мо-
(после разрушения митохондрий детергентом до-
лекула сукцината, имеющая гораздо меньший
децилсульфатом натрия, устраняющим светорас-
размер, чем молекула НАДН, лучше проникает
сеяние) [3].
внутрь митохондрий и поэтому окисляется с бо-
лее заметной скоростью [4].
Спектры ИК-поглощения митохондриальных
суспензий, помещенных и высушенных на про-
При добавлении небольших (0.1-0.4 мМ) ко-
зрачных пластинах фторида кальция, регистри-
личеств глицерина к суспензии свежевыделенных
ровали на ИК-Фурье-спектрометре FT-801 («Си-
печеночных митохондрий, помещенных в изото-
мекс», Новосибирск). Образцы получали путем
ническую среду, наблюдается активация НАДН-
центрифугирования суспензий без глицерина
зависимого (рис. 1, табл. 1) и сукцинат-зависимо-
(контроль) и в присутствии 100-400 мкМ глице-
го дыхания (рис. 2, табл. 1). Активация вызвана
рина (опыт). Супернатанты отбрасывали и высу-
тем, что молекулы глицерина повышают прони-
шивали 50 мкл осадка на пластинах с помощью
цаемость внутренней мембраны. Глицерин,
конвекции теплого воздуха. Для ИК-спектроско-
встраиваясь в отдельные участки липидной фазы,
пии митохондрии специально выделяли в 300 мМ
вероятно, нарушает их (рис. 3). В результате
хлорида натрия - без добавления сахарозы и дру-
НАДН и сукцинат легче входят внутрь, где и
БИОФИЗИКА том 66
№ 2
2021
ТРАНСПОРТ ГЛИЦЕРИНА В МИТОХОНДРИИ
273
Таблица 1. Скорость потребления кислорода (в мкМ/мин/мг) в суспензии печеночных митохондрий
Глицерин, мкМ
0
100
200
400
900
Скорость на НАДН в изотонии
28 ± 3
43 ± 3
39 ± 3
50 ± 4
41 ± 3
Скорость на сукцинате в изотонии
11 ± 3
17 ± 3
26 ± 3
81 ± 5
34 ± 3
Скорость на НАДН в гипотонии
120 ± 5
90 ± 5
50 ± 4
40 ± 4
-
Скорость на сукцинате в гипотонии
145 ± 5
110 ± 5
120 ± 5
130 ± 5
-
Примечание. Концентрация митохондрий в суспензии - 2 мг/мл. Изотоническая среда: митохондрии выделяли в среде,
содержащей сахарозу - 250 мМ, ЭГТА - 0.5 мМ, HEPES - 10 мМ, pH 7.4; для полярографии использовали изотоническую
среду, содержащую сахарозу - 150 мМ, трис-фосфатный буфер - 10 мМ, pH 7.0. Гипотоническая среда: митохондрии
выделяли в 300 мМ NaCl, для полярографии использовали дистиллированную воду (2 мл), добавляя туда митохондрии
(0.5 мл), концентрация NaCl в итоге составляла 40 мМ..
окисляются в активных центрах своих дегидро-
ляет дыхание уже в малой концентрации (рис. 5).
геназ.
Нужно отметить, что в гипотонических условиях
кислород при окислении НАДН потребляется не
Зависимость дыхания от концентрации глице-
только на цитохромоксидазе, но и непосред-
рина имеет сложный характер как в случае
ственно на НАДН-оксидазе
[4]. Добавление
НАДН, так и в случае сукцината (рис. 4, табл. 1).
глицерина в небольших концентрациях в гипото-
При малых количествах глицерина в обоих случа-
нических условиях приводит к существенному
ях имеет место ускорение, обусловленное повы-
угнетению НАДН-зависимой дыхательной ак-
шением мембранной проницаемости, а в более
тивности (рис. 6, табл. 1). Местом приложения
высоких - угнетение, что может быть связано как
глицерина тут является сам НАДН-дегидрогеназ-
с прямым действием глицерина на дегидрогена-
ный комплекс I, а не другие комплексы дыхатель-
зы, так и со слишком сильным повреждением
ной цепи, включая цитохромоксидазу. Это следу-
мембран или (и) торможением ферментов за счет
ет из того, что сукцинатоксидазная активность (в
высокой микровязкости при связывании глице-
условиях гипотонии) глицерином почти не угне-
рина в активном центре. О повреждении свиде-
тается (рис. 6).
тельствует уменьшение светорассеяния суспен-
зии митохондрий. Так, при добавлении глицери-
Подавление НАДН-зависимого дыхания гли-
на свыше 0.9 мМ наблюдалось падение величины
церином трудно объяснить тривиальным повы-
светорассеяния (измеряемого по величине опти-
шением вязкости раствора, так как вязкость воз-
ческой плотности при 700 нм) с 1.7 до 0.8, что обу-
растает не слишком сильно: с ~1 сП (водный рас-
словлено выходом оптически плотного матрикс-
твор) до 1.8 сП (0.4 мМ глицерина) [5]. Против
ного содержимого наружу из-за резкого повыше-
тривиального эффекта вязкости говорит также
ния проницаемости внутренней мембраны (этот
вопрос был подробно рассмотрен в работе [4]).
В гипотонической же среде, когда проницае-
мость внутренней мембраны велика и НАДН
окисляется достаточно быстро, глицерин подав-
Рис. 4. Зависимости скорости сукцинат-оксидазной
(1) и НАДН-оксидазной (2) реакции в суспензии ми-
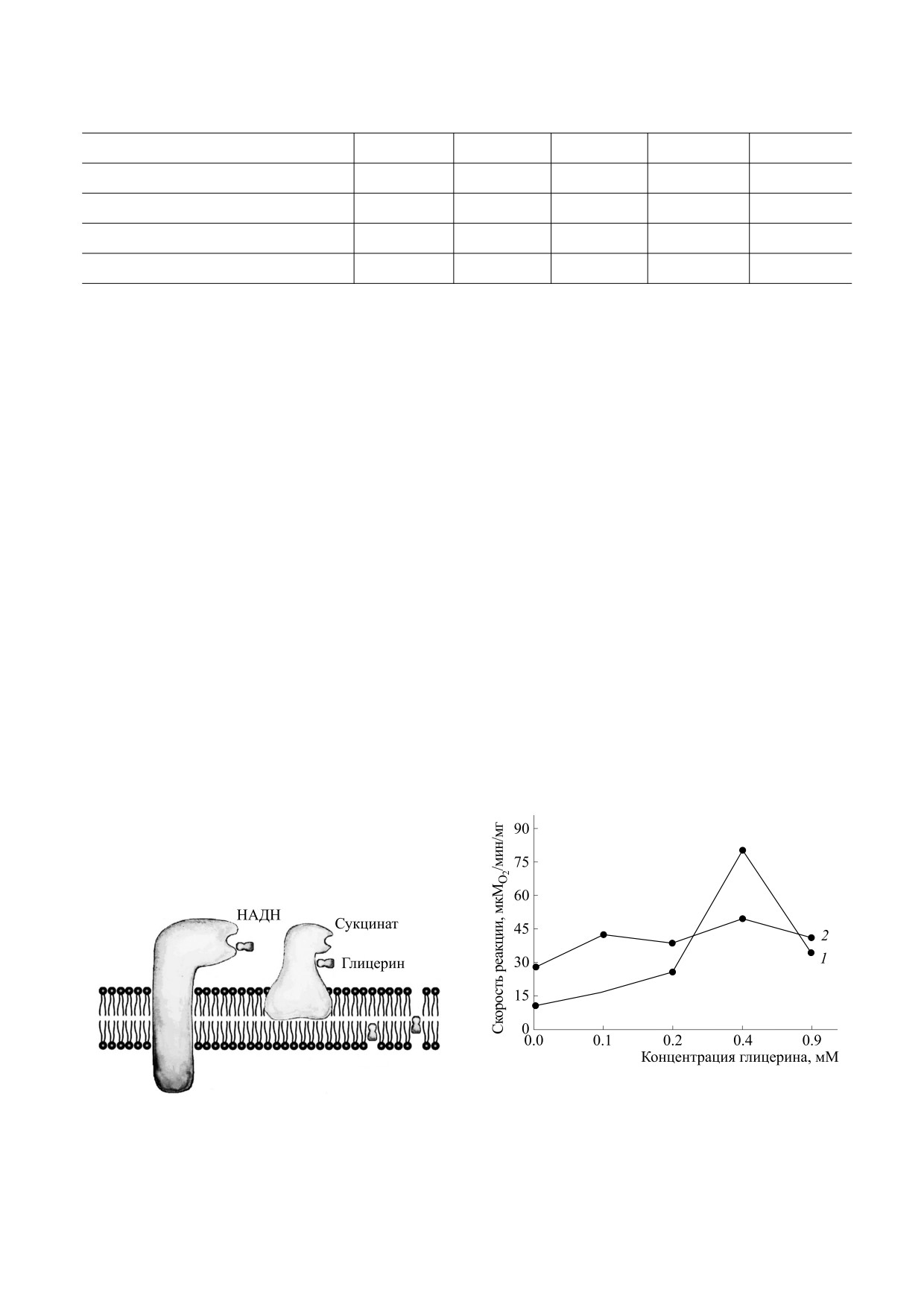
Рис. 3. Схема точек приложения глицерина во внут-
тохондрий в изотонической среде от концентрации
ренней митохондриальной.
глицерина.
БИОФИЗИКА том 66
№ 2
2021
274
ДАЛЬ, ВЕКШИН
Рис. 5. Потребление кислорода в суспензии митохон-
Рис. 6. Зависимости скорости сукцинат-оксидазной
дрий в гипотонической среде (40 мМ NaCl) при добавке
(1) и НАДН-оксидазной (2) реакции в суспензии ми-
1 мМ НАДН в присутствии 400 мкМ глицерина (1) и без
тохондрий в гипотонической среде от концентрации
глицерина (2).
глицерина.
то, что глицерин почти не влияет на скорость сук-
вертом случае глицерин мешает диффундировать
цинатзависимого дыхания (табл. 1). Из получен-
убихинону, который должен снимать электроны с
ных данных следует, что глицерин действует
НАДН-дегидрогеназы.
непосредственно на НАДН-дегидрогеназный
Судя по действующим концентрациям глице-
комплекс I (рис. 3), но не на дыхательные ком-
рина, его константа связывания с мембранами
плексы II, III и IV.
митохондрий достаточно велика и составляет не
Возможны четыре причины угнетающего дей-
менее 104 М-1. Глицерин может хорошо связы-
ствия глицерина на окисление НАДН комплек-
ваться с мембранными липидами и белками за
сом I (рис. 3): а) повышение микровязкости в
счет формирования сразу трех водородных связей
районе активного центра фермента; б) аллостери-
своими тремя гидроксильными группами.
ческое ингибирование; в) конкурентное ингиби-
рование; г) повышение микровязкости липидной
Существенное связывание глицерина с мито-
фазы. В первых двух случаях молекула глицерина
хондриальными мембранами подтверждается
воздействует на фермент путем образования с
прямыми данными ИК-Фурье-спектроскопии
ним трех водородных связей. В третьем случае
(рис. 7). Главный вклад в ИК-поглощение теней
молекула глицерина, входя в карман фермента,
митохондрий, потерявших матрикс, дают мем-
мешает вхождению туда молекулы НАДН. В чет-
бранные белки [4]. Широкая интенсивная полоса
Рис. 7. Спектры ИК-поглощения высушенных теней митохондрий (на поверхности окошка из фтористого кальция)
без добавок (1) и с мембранно-связанным глицерином (2). Митохондрии выделяли из печени в растворе 0.3 М NaCl
(без сахарозы, триса и ЭГТА).
БИОФИЗИКА том 66
№ 2
2021
ТРАНСПОРТ ГЛИЦЕРИНА В МИТОХОНДРИИ
275
Таблица 2. Зависимость интенсивности триптофановой флуоресценции белков митохондрий от добавления
глицерина и дейтерированного глицерина
Интенсивность триптофановой флуоресценции
То же + 0.1 мМ
То же + 0.1 мМ
Длина волны, нм
митохондрий, отн. ед.
глицерина
дейтероглицерина
320
0.94
0.57
1.28
355
1.05
0.71
1.48
380
0.53
0.36
0.72
в районе 3300 см-1 обусловлена валентными ко-
ляет прежде всего работу НАДН-дегидрогеназно-
го комплекса I.
лебаниями ОН-групп и NH-групп (белковый
Амид А); дуплет в районе 2900 см-1 обусловлен
Факт связывания глицерина с мембранами на-
валентными колебаниями ОН- и СН-групп бел-
тивных митохондрий подтверждается данными
по изменению их белковой триптофановой флуо-
ков и липидов; небольшая полоса в районе
ресценции (табл. 2). Глицерин тушит триптофа-
1660 см-1 обусловлена валентными колебаниями
новую флуоресценцию, акцептируя вибронную
СО-групп (белковый Амид I), а также деформа-
энергию на валентные колебания групп -ОН и -
ционными колебаниями ОН-групп молекул свя-
СН, а дейтероглицерин (D8), наоборот, усилива-
занной воды, оставшейся после высушивания об-
ет - благодаря увеличению квантового выхода
разца; небольшой пик при 1540 см-1 (Амид II)
флуоресценции триптофана в окружении тяже-
вызван
деформационными
колебаниями
лых групп -ОD и -CD.
NH-групп. При связывании глицерина с мито-
хондриями резко усиливается широкая полоса
БЛАГОДАРНОСТИ
при 3300-3400 см-1, что связано с валентными
колебаниями трех ОН-групп глицерина. Дефор-
Авторы выражают признательность А.В. Ча-
мационный дуплет глицерина при 1050 см-1 в
плыгиной, М.С. Фроловой и А.Н. Дойниковой за
мембранах заметно усилен в сравнении со спек-
помощь в работе.
тром самого глицерина (на рисунке сам глицерин
не показан). Поскольку при связывании глицери-
КОНФЛИКТ ИНТЕРЕСОВ
на наблюдается снижение белкового пика при
1540 см-1, то это говорит о том, что молекулы гли-
Авторы заявляют об отсутствии конфликта
церина связываются не только в липидной фазе,
интересов.
но также образуют комплексы с поверхностями
мембранных белков. Кроме того, при связывании
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
глицерина с мембранами усиливается пик при
2300 см-1, принадлежащий углекислому газу,
Все применимые международные, националь-
имеющемуся в мембранах и в глицерине (фоно-
ные и институциональные принципы ухода и ис-
вый сигнал углекислого газа воздуха был вычтен).
пользования животных при выполнении работы
Общая приподнятость ИК-спектра теней митохон-
были соблюдены.
дрий в присутствии глицерина по сравнению с те-
нями митохондрий без глицерина обусловлена
большим светорассеянием этого образца (поэтому
СПИСОК ЛИТЕРАТУРЫ
при оценке интенсивностей белковых полос вклад
1. М. Кокс и Д. Нельсон, Основы биохимии Лениндже-
этой бесструктурной полосы вычитали).
ра (Москва, 2017).
Таким образом, на основании полученных ре-
2. И. В. Шарова и Н. Л. Векшин, Биофизика 49 (5),
зультатов можно заключить, что глицерин спосо-
814 (2004).
бен сам по себе напрямую транспортироваться
3. N. L. Vekshin, Photonics of Biopolymers (Springer, Ber-
(пассивно по градиенту) внутрь митохондрий, а
lin, 2002).
не только в форме глицерофосфата. Он эффек-
4. Н. Л. Векшин, Биофизика митохондрий (Фотон-
тивно связывается с митохондриальными мем-
век, Пущино, 2019).
бранами, формируя каналы и повышая проница-
емость внутренней мембраны для субстратов -
5. Краткий справочник физико-химических величин,
НАДН и сукцината. Проникая внутрь, он подав-
сост. Н. М. Барон и др. (Химия, Л., 1974)..
БИОФИЗИКА том 66
№ 2
2021
276
ДАЛЬ, ВЕКШИН
Glycerol Transport in Mitochondria
A.I. Dal* and
**
N.L. Vekshin
*Pushchino State Institute of Natural Sciences, prosp. Nauki 3, Pushchino, Moscow Region, 142290 Russia
**Institute of Cell Biophysics, Russian Academy of Sciences, Institutskaya ul. 3, Pushchino, Moscow Region, 142290 Russia
Particular emphasis is placed on the entry of glycerol molecules (in a non-phosphorylated form) into the mi-
tochondria. Spectrofluorimetry and FTIR spectroscopy have revealed that glycerol is effectively bound to
mitochondrial membranes from rat liver. By using polarography and turbidimetric methods it was shown that
at low concentrations (0.1-0.4 mM) glycerol increases the permeability of the inner membrane for substrates,
such as NADH and succinate, and at high concentrations it strongly damages membranes. Low glycerol con-
centrations increase but high concentrations of glycerol suppress oxygen consumption of intact mitochondria
suspended in an isotonic medium with NADH or succinate added. In a hypotonic medium, glycerol imme-
diately begins to suppress NADH dehydrogenase, but not succinate dehydrogenase. The data obtained pro-
vide evidence for a conclusion that glycerol itself can directly transported (passively along a gradient) into the
mitochondria, and not only in the form of glycerophosphate. It is assumed that glycerol, while interacting
with lipids and proteins, forms channels in the inner membrane for the penetration of itself and substrates.
Keywords: mitochondria, glycerol, oxygen consumption, NADH dehydrogenase, succinate dehydrogenase, respi-
ratory chain
БИОФИЗИКА том 66
№ 2
2021