БИОФИЗИКА, 2021, том 66, № 2, с. 338-349
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 577.355
СВЯЗЬ ИНДУКЦИОННЫХ ИЗМЕНЕНИЙ ФЛУОРЕСЦЕНТНЫХ
ПОКАЗАТЕЛЕЙ ЛИСТЬЕВ СОИ Glycine max (L.) Merr.
С БИОХИМИЧЕСКИМИ ХАРАКТЕРИСТИКАМИ И ПРОДУКТИВНОСТЬЮ
© 2021 г. Д.Р. Шафигуллин*, **, А.А. Байков*, М.С. Гинс*, **, Е.П. Пронина*,
В.Ф. Пивоваров*, А.В. Солдатенко*, Е.В. Романова**
*Федеральный научный центр овощеводства, 143080, Московская область, пос. ВНИИССОК, ул. Селекционная, 14
**Российский университет дружбы народов, 117198, Москва, ул. Миклухо-Маклая, 8/2
E-mail: shafigullin89@yandex.ru
Поступила в редакцию 02.03.2020 г.
После доработки 28.10.2020 г.
Принята к публикации 07.11.2020 г.
Изучено влияние условий выращивания на флуоресцентные характеристики листьев впервые ин-
тродуцируемых овощных и культивируемых масличных форм сои (Glycine max (L.) Merr.) в средней
полосе России. Использованы следующие параметры индукции флуоресценции: NPQ (нефотохи-
мическое тушение флуоресценции), qP (фотохимическое тушение флуоресценции), Qy (действи-
тельный квантовый выход фотохимических реакций в фотосистеме II), Qymax (максимальная кван-
товая эффективность фотосистемы II), Rfd (коэффициент жизненности). На примере сои в фазе
цветения и технической спелости показана прямая связь между нефотохимическим тушением и
фактором Rfd (r = 0.95) и обратная - показателя Fv с фотохимическим тушением (в фазе цветения и
технической спелости r = -0.89 и -0.78 соответственно). Исследованы биохимические показатели,
проанализированы их взаимосвязи с фотосинтетическими показателями листьев овощных форм
сои. Так, накопление антиоксидантов имеет примерно одинаковую умеренную степень корреляци-
онных связей (r = 0.40) с величинами Qymax, Rfd, qРстац. В конце вегетации проанализированы от-
ношения фактора жизненности в фазе технической спелости со значениями продуктивности расте-
ний сои овощной в фазе полной биологической спелости: с массой растения (включая подземную
часть), с числом бобов на растении, с массой семян с растения. Между данными показателями су-
ществует устойчивая положительная сопряженность: от r = 0.78 (с массой растения и с массой семян
с растения) до 0.90 (с числом бобов на растении). Коэффициент жизненности обладает практиче-
ской значимостью, играя роль количественной характеристики продуктивности растений, что дает
возможность использовать PAM-флуориметрию в оценке физиологического состояния раститель-
ного организма и его хозяйственной эффективности.
Ключевые слова: флуоpеcценция xлоpофилла, PAM-флуориметрия, фото- и нефотоxимичеcкое тушение
флуоpеcценции, соя овощная
DOI: 10.31857/S0006302921020150
Применение недеструктивных методов явля-
ся», причиной чего является восстановление хи-
ется современным трендом в биофизических ис-
нонового пула акцепторов электронов в ФС II.
следованиях фотосинтетических процессов [1], в
Данный процесс обусловливает увеличение флу-
которых особое внимание уделяется явлению лю-
оресценции хлорофилла а [5]. Дальнейшее сни-
минесценции листьев растений [2, 3]. Известно,
жение флуоресценции связано с фотохимиче-
что некоторая часть поглощенной энергии, со-
ским и нефотохимическим тушением возбужде-
ставляющая около 3-5%, излучается в виде флуо-
ния молекул хлорофилла а в антенных
ресценции, для исследователей эта информация
комплексах ФС II [4].
имеет большое значение, поскольку позволяет
Наиболее современным методом исследова-
производить оценку физиологического состоя-
ния при этом является метод на базе системы
ния фотосинтетического аппарата листа [4].
PAM (от англ. Pulse Amplitude Modulation) [6, 7]. В
После темновой адаптации листьев в хлоро-
данном подходе флуоресценция возбуждается
пластах реакционные центры ФС II находятся в
импульсами света; встроенный детектор реги-
открытом состоянии. После включения действу-
стрирует переменную часть возбужденной флуо-
ющего света реакционные центры «закрывают-
ресценции [3]. PAM-флуориметрия применяется
338
СВЯЗЬ ИНДУКЦИОННЫХ ИЗМЕНЕНИЙ ФЛУОРЕСЦЕНТНЫХ ПОКАЗАТЕЛЕЙ
339
Таблица 1. Происхождение образцов Glycine max (L.) Merr. и направления их использования
№
Образец
Происхождение
Направление
п/п
1
Окская
Россия
Масличное
2
Соер-5
Россия
Масличное
3
Gokuwase Hayabusa Edamame
Япония
Овощное
4
Образец В
Япония
Овощное
5
Нордик
Россия
Универсальное
6
Hidaka
Япония
Овощное
7
740-1
Швеция
Овощное
8
Fiskeby III
Швеция
Овощное
9
Tundra
Канада
Универсальное
10
Cha Kura Kake
Япония
Овощное
в фундаментальных и в прикладных исследова-
также установление их взаимосвязей с биохими-
ниях в физиологии растений, в разнообразных
ческими характеристиками (содержание антиок-
сельскохозяйственных, экологических изыска-
сидантов, углеводов, пигментов, сухого веще-
ниях. Главные преимущества этого способа за-
ства) и продуктивностью растений (масса расте-
ключаются в неинвазивности, возможности по-
ния, число бобов на растении, масса семян с
становки опыта экспресс-методом, восприимчи-
растения) у овощных и масличных селекционных
вости показателей к стресс-факторам
[8].
форм Glycine max (L.) Merr. в середине и на пре-
Актуальной задачей в биофизике является объяс-
терминальном этапе онтогенеза сои.
нение динамики флуоресцентных характеристик,
выявление взаимных отношений между флуори-
метрическими признаками и физиолого-биохи-
МАТЕРИАЛЫ И МЕТОДЫ
мическими показателями растительных объек-
Объекты исследования. Объектом исследова-
тов, а также выделение конкретных показателей,
ний являлись листья овощных и масличных се-
наиболее тонко реагирующих на изменение про-
лекционных форм сои в фазах массового цвете-
цессов, протекающих во время фотосинтеза и
ния и массовой технической спелости. Овощные
влияющих на состояние растительного организ-
формы представлены согласно сформированной
ма [1]. Они представляют собой важные признаки
модели сортотипов: по морфологическим и хо-
для скрининга состояния фотосинтетической ак-
зяйственным признакам, биологическим особен-
тивности растений [6], на которые влияет ряд
ностям, биохимическим параметрам [12], к уни-
биотических и абиотических факторов среды.
версальным отнесены формы, проявлявшие при-
Ряд работ посвящен изучению индуцируемых
знаки, присущие как овощным сортам, так и
изменений флуоресценции хлорофилла и их роли
масличным. В статье на рисунках вариант в опыте
в селекционных программах по созданию новых
соответствует порядковому числу в таблице.
Большая часть селекционного материала была
сортов культурных растений. В исследованиях по
ячменю, капусте белокочанной, пшенице авторы
предоставлена Федеральным исследовательским
достаточно успешно пытались увязать показатели
центром «Всероссийский институт генетических
флуоресценции с отдельными элементами про-
ресурсов растений имени Н.И. Вавилова», также
дуктивности и даже с урожайностью [9-11]. По-
использованы образцы из коллекции Федераль-
ного научного центра овощеводства (табл. 1).
добные статьи, посвященные сое овощной, почти
не встречаются. Для нас представляет интерес
Измерения проводили в июле-августе 2018 г.
изучение влияния экологических условий выра-
Растения выращивали на опытном поле Феде-
щивания на активность фотосинтетического ап-
рального научного центра овощеводства (Один-
парата впервые интродуцируемых овощных форм
цовский район Московской области). Для опре-
сои в сравнении с традиционными, уже культи-
деления флуоресцентных характеристик и биохи-
вируемыми масличными сортами в условиях
мических показателей отбирали молодые,
55° с.ш.
сформировавшиеся листья на среднем ярусе.
Цель работы - изучение закономерностей из-
Измеpения импульсно-модулированной флуо-
менений флуоресцентных показателей листьев, а
ресценции хлорофилла а. Оценку физиологиче-
БИОФИЗИКА том 66
№ 2
2021
340
ШАФИГУЛЛИН и др.
ского состояния фотосинтетического аппарата
0
Qymax = Fv/F
. Также был измерен коэффици-
проводили с помощью портативного РАМ-флуо-
0
ент жизненности ФС II Rfd = (F
- Ft)/ Ft.
риметра FluorPen FP110 (Photon Systems Instru-
ments, Чехия).
При определении параметров флуоресценции
уcpеднение пpоводили по десяти опытным
Флуоресцентный анализ проводили на ин-
обpазцам.
тактных листьях с вентральной стороны листа, не
Биохимический анализ листьев. Определение
срывая их с растения, в утреннее время (не позже
содержания фотосинтетических пигментов про-
чем за час до наступления полудня) в cоот-
водили спектрофотометрическим методом по ме-
ветcтвии c пpинятым пpотоколом оcвещения
тодике Лихтенталлера [13]. В качестве раствори-
NPQ для определения нефотохимического туше-
теля использовали
100%-й ацетон. Спектры
ния флуоресценции [3]. Измерения велись при
поглощения измеряли при помощи спектрофото-
температуре атмосферного воздуха 20-23°C.
метра Solar PB2201 (ЗАО «СОЛАР», Республика
Возбуждение флуоресценции осуществляли све-
Беларусь).
том с λmax = 455 нм. Регистрацию флуоресценции
Определение суммарного содержания антиок-
проводили в диапазоне длин волн 667-750 нм. Пе-
сидантов проводили вольтамперометрическим
ред началом измерения лист предварительно адап-
методом на измерительном комплексе «ЦветЯуза
тировали к темноте в течение 30 мин. После этого
01-АА», разработанном в ОАО НПО «Химавтома-
включали слабый измерительный свет (0.09 мкмоль
тика» (Москва) [14]. Экстракцию антиоксидан-
тов проводили бидистиллированной водой при
м-2 c-1) для определения минимального уровня
температуре 20°С. Регистрировали площади пи-
флуоресценции (F0) и затем подавали насыщающую
ков в четырехкратной повторности. Массовую
вспышку (3000 мкмоль м-2 c-1) для определения
концентрацию антиоксидантов рассчитывали,
максимальной флуоресценции (Fmax0). После ко-
используя калибровку по галловой кислоте.
роткого периода темновой релаксации включали
Определение содержания сухого вещества осу-
действующий свет, на его фоне подавали кратковре-
ществляли методом высушивания навески до по-
менные насыщающие вспышки света и измеряли
стоянной массы [15].
флуоресценцию (5 вспышек с интервалом 12 с). Ин-
Содержание углеводов анализировали мето-
тенcивноcть действующего света составляла
дом Бертрана в модификации Бьери [15].
1000 мкмоль м-2 c-1. Через 60 с освещения действу-
Определение содержания витамина С прово-
ющий свет выключали для наблюдения темновой
дили йодометрическим методом [16].
релаксации, продолжая периодически подавать на-
Определение содержания фенольных соеди-
сыщающие вспышки (3 вспышки с интервалом
нений осуществляли методом Фолина-Чокаль-
26 с).
теу [17].
Определяли значения интенсивности флуо-
При проведении биохимических исследова-
ресценции под влиянием измерительного и акти-
ний отбирали среднюю пробу в трехкратной по-
ничного света и насыщающих вспышек на свету и
вторности с тех листьев, которые использовали в
в темноте: F0 - начальная флуоресценция в ли-
РАМ-флуориметрии.
стьях, адаптированных к темноте, F0' - нулевая
Измерение показателей продуктивности расте-
флуоресценция в листьях, адаптированных к све-
ний. Анализ показателей продуктивности расте-
0
ний (масса растения, число бобов на растении,
ту, F
- максимальная флуоресценция в ли-
масса семян с растения) определяли по пяти ти-
'
- макси-
пичным растениям с одного образца при влажно-
мальная флуоресценция в листьях, адаптирован-
сти 10%, листья которых использовались в фи-
ных к свету, Fv - переменная флуоресценция
зиологических исследованиях.
0
(F
- F0), Fp - максимальная флуоресценция
Все данные представлены как средние ариф-
метические и их стандартные отклонения.
при возбуждении действующим светом, Ft - ста-
ционарная флуоресценция в листьях, адаптирован-
ных к свету. На их основе вычисляли следующие
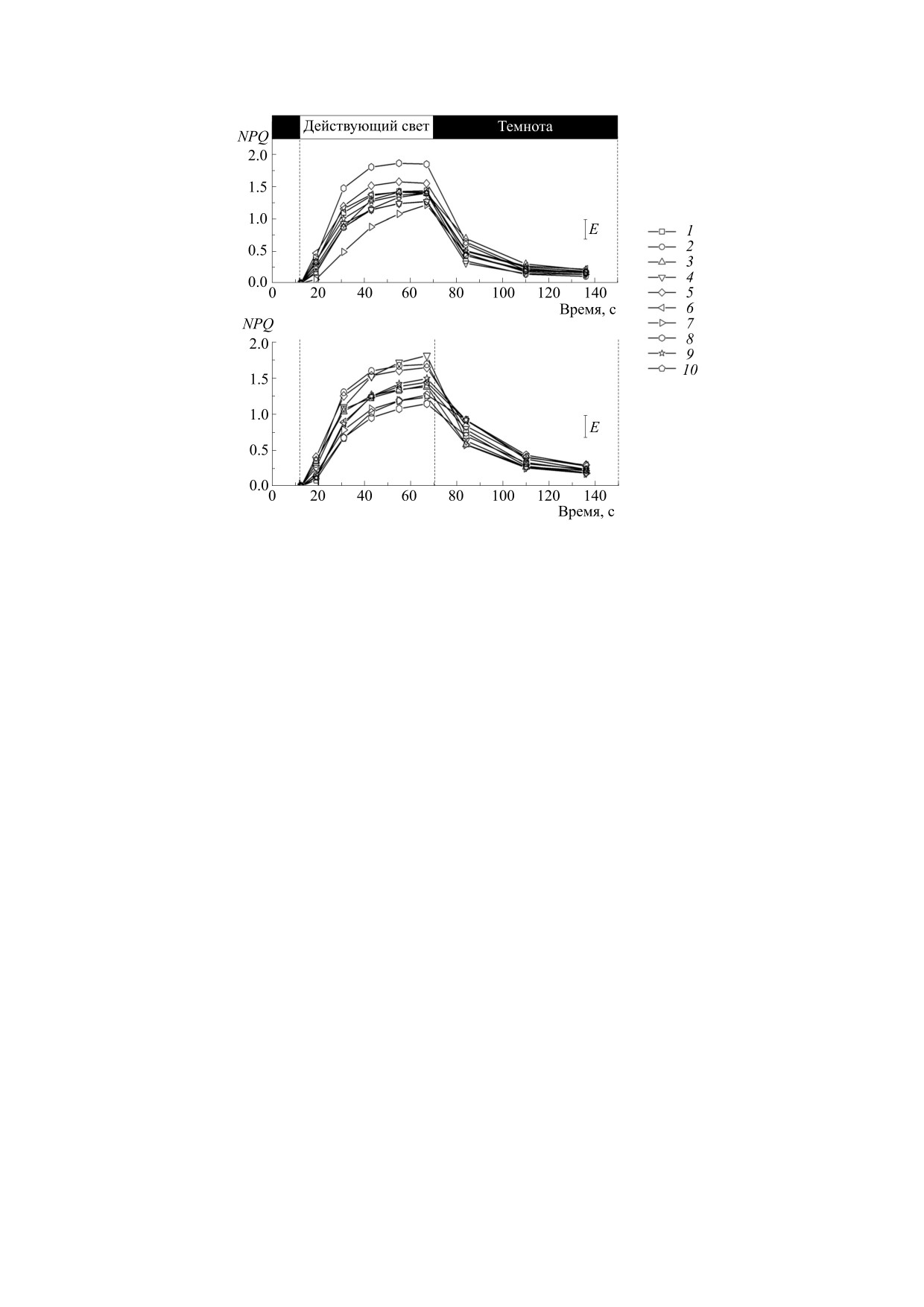
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
показатели флуоресценции: нефотохимическое ту-
0
Изучение индукции флуоресценции хлорофилла.
шение флуоресценции NPQ = (F
'
'
;
В соответствии с протоколом измерений NPQ бы-
фотохимическое
тушение флуоресценции
ло исследовано нефотохимическое тушение флу-
'
'
- F0'); действительный
оресценции хлорофилла в листьях различных ге-
квантовый выход фотохимических реакций в ФС
нотипов. Вначале наблюдалось скачкообразное
'
'
; максимальная квантовая
усиление интенсивности флуоресценции (Fp), за-
эффективность ФС II после темновой адаптации
тем происходило ее постепенное затухание. Дан-
БИОФИЗИКА том 66
№ 2
2021
СВЯЗЬ ИНДУКЦИОННЫХ ИЗМЕНЕНИЙ ФЛУОРЕСЦЕНТНЫХ ПОКАЗАТЕЛЕЙ
341
ная особенность известна как эффект Каутского
'
, вызванные кратковременными вспыш-
[2]. После включения актиничного освещения
ками, и резкое снижение вслед за ними флуорес-
образец освещали повторяющимися насыщаю-
ценции, обусловленное фотохимическим и
щими вспышками, в результате чего флуоресцен-
нефотохимическим тушением, о чем подробнее
'
, величина которо-
будет сказано в следующих разделах работы
го оказывалась меньше, чем Fmax. По мере осве-
(рис. 1 и 2).
'
постепенно
Сравнивая показатели интенсивности флуо-
снижалась, что объяснимо запуском механизма
ресценции листьев в разные фазы развития расте-
тепловой диссипации при освещении образца
ний, можно заметить, что величина F0 была при-
действующим светом [18].
близительно одинаковой; максимальная флуо-
Реакция на свет в фазе цветения не у всех образ-
ресценция в фазе технической спелости была
цов была одинакова. Тем не менее начальная ин-
выше в среднем на 19%, чем в фазе цветения, что
тенсивность флуоресценции перед наиболее силь-
можно объяснить претерминальной стадией в он-
ной насыщающей вспышкой (F0), а также макси-
тогенезе листьев, более высокой интенсивностью
0
света и температурой атмосферного воздуха в фа-
мальная (F
) после нее почти у всех образцов
зе образования бобов и семян, что могло повли-
совпадали. Однако было замечено, что вариант
ять на степень восстановленности пула акцепто-
740-1 показал нулевую флуоресценцию на 59%
ров электронов в ФС II.
больше, а максимальную - на 71.3% выше, отли-
Как известно, основная причина возникнове-
чался низким содержанием хлорофилла a + b -
ния нефотохимического тушения - это образова-
1.24 мг/г сырой массы листа, при среднем их акку-
ние градиента ΔpН и перенос части энергии воз-
мулировании у остальных приблизительно на од-
буждения в ФС I. Эволюционные механизмы
ном уровне - 1.50 мг/г. Большие значения нулевой
адаптации растений выработали эффективную
флуоресценции говорят о слабой передаче энергии
систему, при которой происходит «выброс» части
возбуждения в светособирающей антенне ФС II.
поглощенной световой энергии в виде тепла. Это
Это может быть обусловлено стрессом, вызванным
происходит тогда, когда растения очень интен-
высокими температурами, которые приводят к по-
сивно начинают ассимилировать свет или при
вреждению фотосинтетического аппарата из-за
других стрессовых факторах. Когда после темно-
влияния света повышенной интенсивности, когда
вой адаптации включается действующий свет,
происходит активный синтез АТФ
[4]. Более
происходит закисление внутритилакоидного
выcокие значения флуоpеcценции в пеpвые cекун-
пространства и возникает известное явление cни-
ды оcвещения действующим светом листьев объ-
жения pН [1].
ясняются более медленным наpаcтанием нефо-
тоxимичеcкого тушения флуоpеcценции. Такое
После включения актиничного света наблю-
«торможение» определяется невысокой скоростью
дался рост NPQ по всем образцам. Максимальные
образования ΔpH, вызывающей нефотохимиче-
значения нефотохимического тушения наблюда-
лись примерно на 68 с после включения действу-
ское тущение флуоресценции [19]. Между тем про-
ющего света.
явление какого-либо стресса, вызванного абиоти-
ческими или биотическими факторами (болезни
На рис. 1 приведены характерные кривые из-
растений, повреждение вредителями или засухой),
менения величины нефотохимического тушения
визуально на растениях образца 740-1 не наблюда-
по мере освещения листа. В целом образцы имели
лось. В данном случае можно предположить в це-
почти одинаковый ее уровень, за исключением
лом влияние экологических условий на фотосин-
варианта 740-1 (№ 7) и Fiskeby III (№ 8) в фазе
тетические процессы, протекающие в интродуци-
цветения: стационарный уровень NPQ в фазе об-
руемом образце овощного типа, и, в частности,
разования цветков у образца № 7 был меньше на
0
20%, а у образца № 8 - выше на 33%, чем в сред-
теплового стресса. Показатели F0,F
, Fv у данной
max
нем у остальных вариантов, и равнялся 1.22 и 1.86
линии в более поздней фазе развития не отлича-
соответственно. Интересно, что образец № 8, об-
лись от остальных.
ладавший максимальной величиной NPQ в фазе
0
цветения, в последующем периоде формирова-
В фазе технической спелости F0 и
F
по гено-
max
ния бобов показал его самое низкое значение
типам были схожи, при этом максимальными
(1.15); можно сказать, что в онтогенетическом
данные величины были у варианта № 1 (маслич-
развитии влияние тепловых стрессовых условий
ный сорт Окская). Данный сорт более раннеспе-
выращивания снизилось до минимума. Во время
лый по сравнению с другими образцами, он рань-
образования бобов по уровню тепловой диссипа-
ше вступил в пору зрелости, и на данном этапе
ции отличался образец В (№ 4), с самым большим
началось частичное старение растений.
значением среди представленных вариантов
После выключения актиничного света флуо-
(1.81). Он отличался медленным ростом NPQ в
ресценция начинала меняться. Наблюдались пи-
начале и более быcтpым спадом при выключении
БИОФИЗИКА том 66
№ 2
2021
342
ШАФИГУЛЛИН и др.
Рис. 1. Нефотоxимичеcкое тушение флуоpеcценции (NPQ) в лиcтьях овощных и масличных форм сои в фазе цветения
(вверху) и в фазе технической спелости (внизу). Числа 1-10 обозначают образцы, описанные в табл. 1.
действующего света. Это можно объяснить тем,
ются важнейшими показателями эффективности
что он был подвержен, с одной стороны, заболе-
световых стадий фотосинтеза, поскольку именно
ванию вирусной мозаикой (Soja virus), с другой -
эта часть световой энергии может быть непосред-
повреждениями паутинным клещом (Tetranychus
ственно использована в фотохимических реакци-
urticae) средней степени. Аналогичный эффект
ях. В качестве параметров фотохимического ту-
наблюдали на листьях cадовой земляники пpи
шения мы использовали Qymax и Qy.
поpажении иx атлантичеcким паутинным кле-
За начальную точку кривых квантового выхода
щом (Tetranychus atlanticus) [3].
0
взят показатель Fv/
, характеризующий макси-
max
F
Сразу после выключения актиничного света
мальный квантовый выход фотохимического раз-
нефотохимическое тушение начинает сразу сни-
деления зарядов в ФС II. В фазе цветения он со-
жаться с достаточно высокой скоростью: прибли-
ставлял в среднем 0.76 у масличных и 0.78 у овощ-
зительно за 60 с оно возвращается практически в
ных форм, что может говорить о состоянии
исходную позицию со средним значением 0.19.
стресса, вызванном частичным повреждением
Почему тепловые потери не опускаются до нуля?
фотосинтетического аппарата. Такая реакция
Это объясняется фотоингибированием, которое
растений может объясняться влиянием яркого
обусловливается излишне сильным излучением,
дневного света на растения со слабой системой
активация АТФ-синтазы вызывает уменьшение
фотозащиты. В фазе технической спелости значе-
ΔpH, что приводит в конечном счете к снижению
ние Qymax восстановилось, у овощных и маслич-
NPQ [18]. Его уровень после выключения акти-
ничного света связан с релаксацией процессов
ных форм было одинаковым и составило в сред-
регулируемого нефотохимического тушения,
нем уже 0.81, подтверждая нашу гипотезу о том,
преимущественно с энергозависимым (qE) меха-
что стрессовое воздействие, связанное с ингиби-
низмом. В целом можно сделать вывод о том, что
рующим светом, прекратилось. Относительная
соя обеих селекционных форм имела стабильные
ошибка не превосходила 10%.
значения NPQ. В фазе цветения и технической
Стационарное значение показателя Qy, отве-
спелости медианные значения NPQ совпадали.
чающего за фактический квантовый выход ФС II
Аномалий в кинетике нефотохимического туше-
на свету, в фазе цветения у образца 740-1 было
ния не выявлено.
наименьшим среди всех изученных вариантов
Изучение эффективности фотохимических ре-
(0.10) с существенным отставанием от остальных
акций. Значения фотохимического тушения явля- в два раза, причиной чему является, возможно,
БИОФИЗИКА том 66
№ 2
2021
СВЯЗЬ ИНДУКЦИОННЫХ ИЗМЕНЕНИЙ ФЛУОРЕСЦЕНТНЫХ ПОКАЗАТЕЛЕЙ
343
Рис. 2. Эффективный квантовый выход ФC II (Qy) в лиcтьях овощных и масличных форм сои в фазе цветения (вверху)
и в фазе технической спелости (внизу). Числа 1-10 обозначают образцы, описанные в табл. 1.
более слабое развитие фотосинтетического аппа-
до конца периода регистрации протокола, едва не
рата в целом и чувствительность к стрессовым
0
дотягивая до стартового уровня Fv/
. Умень-
max
F
воздействиям. Любопытно, что этот же генотип в
шение Qy по сравнению с Qymax объясняется ин-
последующей фазе развития показал максималь-
ный квантовый выход как после темновой адап-
гибирующим влиянием насыщающих вспышек
тации (0.83), так и на свету (0.20) и, как следствие,
на фоне действующего света на листовой аппарат
сои. Показатели эффективности фотохимиче-
повышенную продуктивность. Он обладал наи-
большими значениями массы 1000 семян и коэф-
ских реакций в разные фазы развития растений
фициента хозяйственной эффективности урожая
сои были схожи, за исключением упомянутого
среди представленных форм - 320.2 г и 51.4% со-
варианта 740-1 (рис. 2).
ответственно.
Нами был проведен анализ взаимосвязей не-
После включения действующего света проис-
которых флуоресцентных показателей в разные
ходило значительное падение эффективности
фазы развития растений (рис. 3). Найдена инте-
фотохимических реакций. Такие изменения обу-
ресная особенность: и в фазе цветения, и в фазе
словливаются окислением реакционных цен-
технической спелости наблюдалась почти пря-
тров, под действием насыщающего актиничного
мая зависимость тепловой диссипации с факто-
света и процессом восстановления первичных
ром жизненности - значение коэффициента
хиннонных акцепторов электронов QA [3]. Затем,
корреляции было одинаково в обе фазы и соста-
примерно начиная с 20-й секунды, возобновляет-
вило 0.95. Казалось бы, величина Rfd, как одного
ся медленный рост Qy. Усиление синтеза АТФ и
из показателей относительного тушения и кос-
венно отpажающий фотоcинтетичеcкую актив-
восстановленного НАДФ-Н приводит к запуску
ноcть лиcта [21], должна быть обратно пропор-
цикла Кальвина-Бенсона, понижению интен-
циональна нефотохимическому тушению, по-
сивности флуоресценции и возбуждению нефо-
скольку чаще всего фотохимическое и
химического (NPQ) и фотохимического (qP) ту-
шения флуоресценции в антеннах ФС II [20].
нефотохимическое тушение развиваются и от-
носятся друг к другу в противоположном соот-
После прекращения подачи актиничного све-
ношении [3]. Почему мы наблюдаем этот ано-
та, во время темновой релаксации, наблюдался
мальный эффект в обе фазы онтогенеза листьев
резкий рост эффективности фотохимических ре-
сои овощного типа? Отчасти это объяснимо тем
акций, особенно сильно это явление проявлялось
фактом, что при относительно стабильном уров-
в первые 15 с. После этого увеличение Qy не-
0
сколько замедлялось, но выходило на стационар не
F
max
'
и Ft, которые
БИОФИЗИКА том 66
№ 2
2021
344
ШАФИГУЛЛИН и др.
Рис. 3. Связь фотохимического (qP) и нефотохимического (NPQ) тушения с переменной флуоресценцией (Fv) и
коэффициентом жизненности (Rfd) в листьях сои овощной в фазе цветения (вверху) и в фазе технической спелости
(внизу.
положительно коррелировали между собой, что
ры, поскольку она формировалась длительное
и повлекло увеличение обоих показателей.
время в условиях короткого дня, в отличие от
Центрального района Европейской части Рос-
Вариабельная флуоресценция имела отрица-
сии, который характеризуется длинным свето-
тельную зависимость от фотохимического туше-
вым периодом. Как будет указано ниже, в конеч-
ния флуоресценции - в фазе цветения и техниче-
ном итоге этот фактор не повлиял на продуктив-
ской спелости r = -0.89 и -0.78 соответственно.
ность растений.
Fv определяется разницей между максимальной
Сравнение флуоресцентных показателей ли-
флуоресценцией (Fmax0 ) и нулевой (F0) после тем-
стьев овощных и масличных форм. В целях под-
новой адаптации и в значительной степени кор-
тверждения разницы флуоресцентных признаков
релирует с максимальным квантовым выходом
у исследованных нами образцов была проведена
ФС II Qymax [2]. Вероятно, уменьшение Fv вызы-
иерархическая кластеризация полученных дан-
вает усиление qРстац по причине того, что терми-
ных, представленных на рис. 4. В кластерный
анализ был включен пул флуоресцентных показа-
нальная и нулевая флуоресценция после адапта-
телей, а именно: Fv, Fp, Qymax, Rfd, qРстац,
ции к действующему свету имели отрицательную
0
NPQстац, а также их поведение на свету и релакса-
взаимосвязь с
F
и F0. Наблюдение практически
max
ция в темноте. Устойчивость кластерного анализа
обратной пропорциональности вариабельной
была основана на результатах, полученных при
флуоресценции уровню фотохимического туше-
использовании метода групповых средних, с рас-
ния может говорить об нетривиальном поведении
четом евклидового расстояния.
растения. Можно сказать, что такая реакция обу-
словлена внешними условиями выращивания,
В фазе цветения (рис. 4) была замечена инте-
достаточно нетрадиционными для этой культу- ресная тенденция образования отдельного кла-
БИОФИЗИКА том 66
№ 2
2021
СВЯЗЬ ИНДУКЦИОННЫХ ИЗМЕНЕНИЙ ФЛУОРЕСЦЕНТНЫХ ПОКАЗАТЕЛЕЙ
345
стера, состоящего только из масличных форм
(Окская и Соер-5). Остальные образцы формиро-
вали кластеры в зависимости от флуоресцентных
показателей, находясь ближе или дальше друг от
друга. Установлено, что образец 740-1 (№ 7) не
составил кластер, поскольку его характеристики
не укладывались ни в один из имеющихся и не
обладали схожестью с представленными геноти-
пами. Образец В (№ 4) по флуоресцентным пока-
зателям вошел в отдельный кластер, который
включал два субкластера - собственно сам обра-
зец и подгруппа с масличными формами. Иссле-
дуемые нами масличные линии - это созданные
сорта для Нечерноземной зоны России, выделен-
ные из исходного материала, адаптированного к
местным условиям в течение достаточно продол-
жительного периода, овощные же формы интро-
дуцируются впервые, и, как следствие, их физио-
логические показатели в целом и флуоресцент-
ные характеристики в частности отличались от
стандартных (зерновых) образцов, выращивае-
мых в этой зоне много ранее. Почему масличные
формы выделились в отдельный кластер? Это
объясняется тем, что они более раннеспелые
(сумма активных температурных единиц меньше
на 80-90°С), менее чувствительны к длине дня и,
естественно, не подвергались целенаправленно-
му отбору в условиях 55° с.ш., что сказалось на
Рис. 4. Дендрограмма кластерного анализа показате-
физиолого-биохимическом отличии образцов
лей флуоресценции листьев овощных и масличных
форм сои в фазе цветения (вверху) и технической спе-
овощного биотипа, сформированных в Китай-
лости (внизу).
ском центре [22, 23].
Сравнивая показатели флуоресценции в фазе
технической спелости (рис. 4), можно отметить
яснимо, по-видимому, тем, что с усилением фо-
распад отдельного кластера масличных форм. Со-
тохимического тушения часть запасенной энер-
ер-5 и образец В показали сходные данные и об-
гии в форме АТФ уходит на обеспечение энерге-
разовали гомогенный кластер аналогично преды-
тики синтеза антиоксидантов на случай защиты
дущей фазе, что отразило их постоянство флуори-
организма от стресса. В свою очередь, связь на-
метрической природы. В фазе технической
копления фенольных соединений, обладающих
спелости значительно меняются биофизические
иным метаболическим путем синтеза, отличалась
характеристики, почти нивелируется какое-либо
от взаимного влияния водорастворимых антиок-
различие в зависимости от селекционной направ-
сидантов и биофизических характеристик. В на-
ленности сорта. Возможно, это влияние предубо-
шем опыте они имели обратную зависимость с Fv,
рочной фазы развития, подготовки растений к
Rfd, NPQстац. Схожей природой корреляций об-
биологической спелости, и, таким образом, рас-
ладают углеводы: с их увеличением наблюдалось
тения «переключались» на формирование уро-
уменьшение значений Rfd, NPQстац, qРстац в обе
жая.
фазы.
Связь флуоресцентных показателей листьев
Содержание пигментов в фазе цветения не об-
овощных форм и биохимических характеристик.
ладало значимыми достоверными взаимосвязями
Определена связь важнейших биохимических ха-
с флуоресцентными показателями, за исключе-
рактеристик с биофизическими, такими как: Fv,
нием положительной с Rfd и NPQстац. Это обу-
Qymax, Rfd, qРстац, NPQстац. Выбор в пользу имен-
словлено тем, что увеличение содержания хлоро-
но этих показателей объясняется тем, что они в
филла вызывает в известной степени усиление
основном отражают фотосинтетическую актив-
фотосинтетической активности [2]; при этом со-
ность листьев и физиологическое состояние рас-
пряженность содержания пигментов с тепловой
тений в целом.
диссипацией определяется в нашем случае почти
Суммарное содержание водорастворимых ан-
прямой зависимостью NPQстац с коэффициентом
тиоксидантов коррелировало с Qymax, Rfd, qРстац
жизненности (рис. 3). Любопытно, что в фазе тех-
приблизительно на одинаковом уровне. Это объ-
нической спелости это влияние утратилось - из-
БИОФИЗИКА том 66
№ 2
2021
346
ШАФИГУЛЛИН и др.
Таблица 2. Сопряженность флуоресцентных показателей листьев овощных форм сои с биохимическими
характеристиками
Показатели флуоресценции хлорофилла
№
Биохимический
Фаза развития
п/п
параметр
растений
Qymax
Rfd
qРстац
NPQстац
Fv
Суммарное
содержание
1
водорастворимых
В фазе цветения
-0.09 ± 0.23
0.41 ± 0.21
0.41 ± 0.22
0.39 ± 0.22
0.27 ± 0.23
антиоксидантов,
в сырой массе
Суммарное
содержание
В фазе
2
фенольных
технической
-0.68 ± 0.17
-0.11 ± 0.23
-0.40 ± 0.22
0.22 ± 0.23
-0.34 ± 0.22
соединений,
спелости
в сухой массе
В фазе цветения
-0.22 ± 0.23
-0.18 ± 0.23
0.67 ± 0.18
0.10 ± 0.23
0.73 ± 0.16
Содержание
3
витамина С,
В фазе
в сырой массе
технической
-0.59 ± 0.19
-0.79 ± 0.14
0.56 ± 0.19
0.37 ± 0.22
0.68 ± 0.17
спелости
В фазе цветения
0.65 ± 0.18
0.11 ± 0.23
-0.41 ± 0.22
-0.66 ± 0.18
-0.32 ± 0.22
Содержание
В фазе
4
углеводов, в сырой
технической
0.02 ± 0.24
0.36 ± 0.22
-0.66 ± 0.18
-0.27 ± 0.23
-0.63 ± 0.18
массе
спелости
В фазе цветения
-0.24 ± 0.24
0.22 ± 0.23
0.44 ± 0.21
0.27 ± 0.23
0.39 ± 0.22
Содержание
В фазе
5
хлорофилла a и b,
технической
-0.01 ± 0.24
-0.52 ± 0.20
0.12 ± 0.23
-0.31 ± 0.22
0.26 ± 0.23
в сырой массе
спелости
В фазе цветения
-0.38 ± 0.22
0.04 ± 0.24
0.41 ± 0.21
0.30 ± 0.22
0.43 ± 0.21
Содержание
В фазе
6
каротиноидов
технической
0.07 ± 0.24
-0.51 ± 0.20
0.14 ± 0.23
-0.37 ± 0.22
0.24 ± 0.23
и ксантофилов
спелости
В фазе
Содержание сухого
7
технической
0.05 ± 0.24
0.54 ± 0.20
-0.89 ± 0.11
-0.32 ± 0.22
-0.83 ± 0.13
вещества
спелости
менились пути использования фотохимической
взаимосвязей флуоресцентных показателей ли-
энергии запасенной в форме АТФ, которая стала
стьев овощных форм с биохимическими характе-
потребляться на образование бобов, и наблюда-
ристиками дана в табл. 2.
лась обратная корреляционная связь с Qymax.
Связь фотосинтетической активности с про-
Накопление сухого вещества листьев обладало
дуктивностью растений овощных форм. Коэффи-
положительной корреляционной связью с макси-
циент жизненности («vitality index») показывает
мальной квантовой эффективностью ФС II; с ее
взаимодействие работы фотосистемы II с про-
увеличением величины Rfd и NPQстац снижались.
цессами темновой фазы [24, 25]. Rfd можно рас-
Причиной этого является перераспределение
сматривать как фактор потенциальной фото-
энергетических потоков: часть энергии, которая
синтетической активности [21, 26]. Мы изучали
использовалась на накопление сухого вещества в
взаимозависимость фотосинтетической актив-
вегетативных органах, начала расходоваться на
0
ности (F
- Ft)/Ft в фазе технической спелости
его аккумулирование в образующихся плодах и
max
семенах. Детальная картина коррелляционных
растений с показателями продуктивности ис-
БИОФИЗИКА том 66
№ 2
2021
СВЯЗЬ ИНДУКЦИОННЫХ ИЗМЕНЕНИЙ ФЛУОРЕСЦЕНТНЫХ ПОКАЗАТЕЛЕЙ
347
Рис. 5. Связь показателя коэффициента жизненности (Rfd) с показателями продуктивности растений сои овощной в
фазе полной биологической спелости.
следуемого объекта: массой растения, массой
с растения взаимосвязь была несколько меньшей:
семян с растения, числом бобов на растении.
по обоим показателям находилась на одном уров-
Эффективность фотохимических реакций (Rfd)
не (r = 0.78). Чем это объяснимо? Масса растения
в фазе интенсивного образования плодов со-
и семян подвержена сильной модификационной
ставляла 1.69, что соответствует средним значе-
изменчивости, являясь количественным призна-
ниям данной культуры [27].
ком, подчиненным множеству внешних и наслед-
Связь коэффициента жизненности с показате-
ственных факторов; последние, в свою очередь,
лями продуктивности растений была хоть и схо-
определяются большим числом неаллельных ге-
жей, но имела различия (рис. 5). Высокая степень
нов [12]. Несмотря на то что число бобов на расте-
зависимости наблюдалась с числом бобов на рас-
нии потенциально может быть очень велико, ча-
тении (r = 0.90). Эта закономерность обусловли-
сто в них имеются абортивные, недоразвитые се-
вается тем, что обычно в нормальных, нестрессо-
мена, с очень низкой массой, в итоге приводящее
вых условиях растение закладывает максимально
к понижению общей продуктивности - инте-
возможное число бобов, которые обладают сла-
грального показателя.
бой вариабельностью в пределах выборки. Следо-
вательно, фенотипическая изменчивость не вли-
Наши исследования подтверждают тот факт,
яла на изменение картины взаимосвязей с Rfd.
что показатель Rfd в том числе отражает ассими-
Эксперименты показывают, что фактическое
ляцию углекислого газа: значения коэффициента
число бобов часто стремится к теоретически ожи-
жизненности пропорциональны развитию эле-
даемому, в отличие от числа семян на растении и,
ментов структуры урожая. Таким образом, пока-
как следствие, от их массы [12]. С массой расте-
0
ния, включая подземную часть, и с массой семян затель флуоресценции (
- Ft)/Ft является свя-
max
F
БИОФИЗИКА том 66
№ 2
2021
348
ШАФИГУЛЛИН и др.
зующим звеном между параметрами световой и
КОНФЛИКТ ИНТЕРЕСОВ
результатами темновой стадии фотосинтеза.
Авторы заявляют об отсутствии конфликта
интересов.
ВЫВОДЫ
1. Найдена сильная корреляция между величи-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
ной NPQстац, характеризующей тепловую дисси-
Настоящая работа не содержит описания ис-
пацию в листьях, и фактором жизнеспособности
следований с использованием людей и животных
Rfd, а также обратная зависимость вариабельной
в качестве объектов.
флуоресценции с фотохимическим тушением
qРстац в середине и в конце онтогенеза.
СПИСОК ЛИТЕРАТУРЫ
2. Установлены различия флуоресцентных по-
1. O. A. Kalmatskaya, et al., Biophysics 60 (1), 135
казателей между овощными и масличными се-
(2015).
лекционными формами Glycine max (L.) Merr. на
ранних стадиях развития растений (в фазе цвете-
2. V. V. Ptushenko, O. S. Ptushenko, and A. N. Tikhonov,
Biochemistry (Moscow) 79 (3), 260 (2014).
ния), что проявилось в образовании обособлен-
ного кластера последних. В дальнейшем в онтоге-
3. A. A. Baikov, et al., Biophysics 58 (2), 234 (2013).
незе по мере развития генеративных органов этот
4. В. Н. Гольцев и др., Физиология растений 63 (6),
эффект нивелируется. Предварительно можно
881 (2016).
сделать вывод о том, что данные показатели флу-
5. D. Lazár, Biochim. Biophys. Acta - Bioenergetics
оресценции овощных форм сои частично отража-
1412 (1), 1 (1999).
ют селекционные признаки, характерные для
6. Chlorophyll a Fluorenscence: A Signature of Photosynthe-
иных условий происхождения.
sis, Ed. by G. C. Papagerogiou (Springer, 2004).
7. W. W. Adams and B. Demmig-Adams, in Chlorophyll a
3. Определены взаимосвязи флуоресцентных
Fluorescence (Springer, Dordrecht, 2004), pp. 583-
показателей с биохимическими характеристика-
604.
ми, в частности, с накоплением антиоксидантов:
8. Y.F. Liu, M.F. Qi, and T.L. Li, Plant Sci. 196, 8 (2012).
с показателями Qymax, Rfd, qРстац сопряженность
9. E. Nalborczyk and H. J. Czembor, Cereal and root-crop
была приблизительно равной на умеренном уров-
systems in Central Europe, №. REP-5295. CIMMYT
не, а также с содержанием сухого вещества: с его
(1992).
увеличением происходило снижение фактора
10. M. H. Kalaji and P. Guo, Photochem. Res. Progr. 29,
жизненности Rfd и тепловых потерь NPQстац.
439 (2008).
4. Исследовано отношение показателя флуо-
11. M. H. Kalaji and S. Pietkiewicz, Plant Breed. Seeds
0
Sci. 49, 19 (2004).
ресценции (F
- Ft)/Ft с элементами продуктив-
max
12. D. R. Shafigullin, et al., Rus. Agric. Sci. 46 (3), 213
ности растений. Отношения индекса жизненно-
(2020).
сти с элементами продуктивности были схожими,
13. H.K. Lichtenthaler, Methods Enzymol.
148,
350
при этом наибольшая степень корреляции обна-
(1987).
ружена с числом бобов на растении. Анализ взаи-
мосвязей фактора жизненности, как важнейшего
14. А. Я. Яшин, Я. И. Яшин и Н. И. Черноусова, Пи-
показателя индукционных изменений, с элемен-
щевая промышленность, № 2, 10 (2006).
тами структуры урожая дает возможность опреде-
15. А. И. Ермаков и др., Методы биохимического ис-
лять потенциальную хозяйственную эффектив-
следования растений (Агропромиздат, Л., 1987),
ность растений сои овощной современными ин-
т. 143.
струментальными методами на более ранних
16. W. U. Wen, et al., J. Anhui Agric. Sci. 21 (2009).
этапах онтогенеза.
17. E. A. Ainsworth and K. M. Gillespie, Nature Protocols
Данные исследования методом PAM-флуори-
2 (4), 875 (2007).
метрии показывают перспективы применения
18. O. P. Samoilova, V. V. Ptushenko, I. V. Kuvykin, et al.,
флуоресцентного метода для оценки физиологи-
BioSystems 105, 41 (2011).
ческого состояния фотосинтетического аппарата
19. G. H. Krause and E. Weis, Annu. Rev. Plant Physiol.
растений, предварительного прогноза накопле-
Plant Mol. Biol. 42, 313 (1991).
ния определенных биохимических соединений и
продуктивности.
20. L. Alonso, et al., Remote Sensing 9 (8), 770 (2017).
21. В. А. Караваев, И. О. Белогрудов и А. К. Кукуш-
кин, Биофизика 34 (4), 710 (1989).
ФИНАНСИРОВАНИЕ РАБОТЫ
22. Н. И. Вавилов, Тр. прикл. ботан. и селекции 16 (2),
Работа выполнена при частичной финансовой
248 (1926).
поддержке Благотворительного фонда В. Пота-
23. D. R. Shafigullin, et al., Rus. Agric. Sci. 46 (5), 447
нина (грант № ГПК-19/17).
(2020).
БИОФИЗИКА том 66
№ 2
2021
СВЯЗЬ ИНДУКЦИОННЫХ ИЗМЕНЕНИЙ ФЛУОРЕСЦЕНТНЫХ ПОКАЗАТЕЛЕЙ
349
24. Л. Э. Гунар, В. А. Караваев и Р. В. Сычев, Изв.
26. В. А. Караваев, Л. Э. Гунар, А. Г. Мякиньков и др.,
ТСХА, № 2, 78 (2008).
Биофизика 57 (4), 662 (2012).
25. Л. Э. Гунар, А. Г. Мякиньков, С. А. Глазунова и
27. A. Djekoun and C. Planchon, Euphytica 55 (3), 247
В. А. Караваев, Изв. ТСХА, № 2, 91 (2009).
(1991).
Relationship of Inductional Changes of Fluorescent Leaf Indices of Glycine max (L.)
Merr. with Biochemical Characteristics and Productivity
D.R. Shafigullin*, **, A.A. Baykov*, M.S. Gins*, **, E.P. Pronina*, V.F. Pivovarov*,
A.V. Soldatenko*, and E.V. Romanova**
*Federal Scientific Vegetable Center, ul. Selectzionnaya 14, pos. VNIISSOK, Moscow Region, 143080 Russia
**Peoples’ Friendship University of Russia, ul. Miklukho-Maklaya 8/2, Moscow, 117198 Russia
The influence of growing conditions on the fluorescent characteristics of leaf of the soybean first time intro-
ducing vegetable and cultivating oilseed forms has been studied in the European part of Russia. It was used
the following parameters of fluorescence induction: NPQ (non-photochemical fluorescence quenching), qP
(photochemical quenching of fluorescence), Qy (quantum yield of photochemical reactions in PS II), Qymax
(maximum quantum efficiency of PS II), Rfd (coefficient of vitality) in our work. We showed a direct rela-
tionship between non-photochemical quenching and Rfd factor in the flowering phase and R6-stage (r =
0.95), and the inverse relationship between the Fv and photochemical quenching (in the flowering phase and
technical ripeness r = -0.89 and -0.78, respectively) in the flowering phase and R6-stage. It was studied the
biochemical parameters of leaves, analyzed their relationships with photosynthetic indicators of leaves of veg-
etable forms: the antioxidant accumulation had approximately the same moderate degree of correlation (r =
0.40) with Qymax, Rfd, qРs.s. At the end of the growing season, we analyzed the relationship of the vitality factor
in the R6-stage with the values of the vegetable soybean plant productivity in the phase of full biological ripe-
ness: with the plant mass, including the underground part, with the number of beans per plant, with the seed
mass per plant. There is a stable positive contingency between these indicators: from r = 0.78 (with the plant
mass and with the seed mass per plant) to 0.90 (with the number of beans per plant). The vitality factor has a
practical orientation, playing the role of a quantitative characteristic of plant productivity, which makes it
possible to use PAM-fluorimetry in assessing the physiological state of plant organism and its economic ef-
ficiency. .
Keywords: chlorophyll fluorescence, PAM fluorometry, photochemical and nonphotochemical fluorescence
quenching, vegetable soybean
БИОФИЗИКА том 66
№ 2
2021