БИОФИЗИКА, 2021, том 66, № 2, с. 350-357
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
574.5: 51.7
ГИПОТЕЗЫ ОБ АДАПТАЦИИ ВОДОРОСЛЕЙ
К ПЕРИОДИЧЕСКИМ ФАКТОРАМ СРЕДЫ
© 2021 г. В.Г. Ильичев*, О.А. Ильичева**
*Южный научный центр РАН, 344006, Ростов-на-Дону, ул. Чехова, 41
E-mail: vitaly369@yandex.ru
**Донской государственный технический университет,
344022, Ростов-на-Дону, ул. Социалистическая, 162
E-mail: oaili@yandex.ru
Поступила в редакцию 25.06.2020 г.
После доработки 12.10.2020 г.
Принята к публикации 20.10.2020 г.
Предложена новая эколого-эволюционная модель водорослей малой размерности. В ней реализо-
вана одновременная динамика переменных (биомассы популяций) и параметров-границ интервала
температурной толерантности [a - d, a + d]. При заданном периодическом режиме температуры
данные параметры стремятся к некоторым финальным, так называемым эволюционно-устойчи-
вым значениям. В результате расчетов установлено, что на плоскости (a, d) все эволюционно-
устойчивые параметры располагаются в форме «радуги», наиболее удаленные точки которой соот-
ветствуют холоднолюбивым (диатомовым) и теплолюбивым (синезеленым) водорослям. Проведен
анализ адаптации водорослей к двум периодическим факторам среды (температуры и солености) с
заданными интервалами толерантности по температуре [a - d, a + d] и по солености [b - c, b + c].
Установлено, что область эволюционно-устойчивых параметров в плоскости (a, b) зависит от сте-
пени синхронности факторов среды. В одних случаях эти параметры лежат на линейном отрезке, а
в других вариантах возможно их неожиданное расположение на нелинейном овале.
Ключевые слова: модель, адаптация, эволюционно-устойчивые параметры, температура, соленость .
DOI: 10.31857/S0006302921020162
Существуют разные подходы к построению
Возможное потепление климата вызовет де-
моделей адаптации и микроэволюции. Создание
формацию не только температурного, но и других
достаточно эффективных эколого-эволюцион-
факторов среды. Так, в разных районах планеты
ных моделей связано с одновременным развити-
изменится уровень осадков, что, в свою очередь,
ем математической экологии ([4, 5] и др.) и мате-
трансформирует там объем и сезонный режим
матической генетики ([6, 7] и др.). Однако прямо-
речного стока. В частности, вариация донского
линейная попытка соединить эти формализации
стока всегда порождает преобразования солено-
приводит к построению эколого-эволюционных
сти Азовского моря - ключевого регулятора его
моделей очень большой размерности. Это замед-
ляет скорость расчетов и затрудняет проведение
продуктивности [1]. Известные модели ([2, 3]
«асимптотических» прогнозов динамики экоси-
и др.) ориентированы преимущественно на опи-
стем на эволюционно-значимые времена.
сание поведения продукционных процессов. Од-
нако для адекватной оценки долговременных
В данной работе на основе идей эволюционно-
экологических последствий при новых условиях
го моделирования предложены новые компью-
среды требуются иные методы прогнозирования,
терные схемы [8], реализующие эти процессы. По
сути, эволюция - это «естественный интеллект
а именно учет одновременной динамики и мик-
экосистем». Важное достоинство предлагаемого
роэволюции морских биоценозов. Игнорирова-
подхода - небольшая размерность моделей, что
ние механизмов биологической адаптации при-
позволяет проводить долгосрочные прогнозы (на
водит к занижению показателей биологической
эволюционно-значимые времена) за приемлемое
продуктивности.
машинное время.
Сокращениe: ЭУ-параметры - эволюционно-устойчивые
Ниже на основе эколого-эволюционной моде-
параметры.
ли обнаружена взаимосвязь оптимальных темпе-
350
ГИПОТЕЗЫ ОБ АДАПТАЦИИ ВОДОРОСЛЕЙ
351
ратур и оптимальных соленостей развития для
каждая из которых является носителем своих эко-
эволюционно-зрелых видов водорослей. Это поз-
логических параметров. В результате их антаго-
воляет существенно снизить биологу выбор воз-
низма большая часть конкурентов погибает, а
можных исходов при адаптации водорослей.
оставшиеся (одна или несколько) - являются но-
сителями «оптимальных» параметров.
АДАПТАЦИЯ ВОДОРОСЛЕЙ
Несколько другой подход был предложен в ра-
К ПЕРИОДИЧЕСКОЙ ТЕМПЕРАТУРЕ.
боте [14], автор которой считал, что в процессе
ГИПОТЕЗА КРИТИЧЕСКИХ ЗНАЧЕНИЙ
адаптации происходит поиск так называемых
«эволюционно-устойчивых» параметров (ЭУ-па-
Исследование характера адаптации популя-
раметры) с использованием теоретико-игровых
ций к температурному режиму среды является до-
соображений о выборе оптимальной стохастиче-
вольно популярной темой [9]. Здесь можно выде-
ской стратегии поведения. Этот удачный термин
лить два подхода, которые можно условно назвать
может быть применен и в детерминированной
внутренним и внешним.
ситуации. А именно, носителем ЭУ-параметра
Во внутреннем подходе делается упор на про-
является популяция, которая не вытесняется сво-
странственных преобразованиях структур неко-
ими мутантами с параметрами, близкими к ис-
торых макромолекул клетки при деформации
ходным.
температуры среды. Так, полагают, что при на-
греве она «раскручивается», а при охлаждении -
Напомним, основной недостаток традицион-
«скручивается» [10].
ного подхода - необходимость задания в модели
Во внешнем подходе ключевую роль играет
большого числа субпопуляций (1000 и более) на
концепция Ф. Добжанского [11] о жестком поли-
каждом трофическом уровне. Возникает проти-
морфизме природных популяций. Суть ее в том,
воречие между требованиями биологической
что популяция представляет собой набор узко-
адекватности и высокой скорости вычислений.
специализированных субпопуляций (форм) с
Здесь компромиссное решение основано на
возможно разными скоростями роста. В этом слу-
изобретении новой модельной конструкции
чае изменение температуры среды вызывает пе-
«Адаптация». Она имитирует эволюционный
рестройку доминирующих форм, отчего меняют-
процесс в малом, имеет небольшую размерность
ся продукционные характеристики популяции в
и поэтому обладает высоким быстродействием. В
целом.
качестве иллюстрации ее практического приме-
Ниже будет использован второй подход. Он
нения будет проведен анализ динамики и микро-
составляет основу эволюционной экологии, а его
эволюции фитоценоза при ежегодном периоди-
функции проще поддаются математическому
ческом изменении температуры водной среды.
описанию. Этот путь достаточно универсален и
был применен при поиске закономерностей ко-
Так, хорошо известно, что каждая группа во-
адаптации маршрутов миграции взаимодейству-
дорослей имеет свой температурный интервал то-
ющих рыбных популяций [12].
лерантности I(a,d) = [a - d, a + d], внутри него
Обычно в эколого-эволюционных моделях
происходит рост водорослей, скорость которого
«прямолинейно» соединены экологические и ге-
(R) в зависимости от температуры воды θ задается
нетические процессы [13]. Предполагается, что
некоторой функцией, например, следующим
любая популяция состоит из ряда субпопуляций,
симметричным образом:
2
2
[d
– (θ -
a)
]
R a,d)
=
r
при θ
∈
I a,d), иначе
R
=
0.
(1)
3
d
Здесь r - некоторая суточная константа роста
иногда используются и «несимметричные» функ-
водорослей (положим r = 3).
ции [16] для описания роста водорослей, но это
Принято говорить: a является оптимальной
не вызывает существенных изменений в резуль-
температурой развития, d - ширина интервала I
татах.
(для конкретной группы водорослей). График
Следует отметить, что естественная гипотеза
функции R в зависимости от q представляет собой
«колокольчик» (см. рис. 1).
Э. Пианки обманчиво проста и требует опреде-
ленных уточнений. Так, исход борьбы двух кон-
В формуле (1) воплощена гипотетическая идея
Э. Пианки [15]: для эволюционно-зрелых попу-
курентов с разными (но равновеликими по пло-
ляций площадь под графиком R не изменяется
щади) «колокольчиками» может зависеть от хода
при вариации параметров a и d. Отметим, что
температурной кривой. Представляет интерес
БИОФИЗИКА том 66
№ 2
2021
352
ИЛЬИЧЕВ, ИЛЬИЧЕВА
видно, что в процессе эволюции могут реализо-
ваться только ЭУ-параметры.
Изложим теперь основную идею модельного
поиска пары ЭУ-параметров, основанную на ме-
тодах эволюционного моделирования [8]. Здесь
важно отметить, что базовое понятие вытеснения
популяции (достижения ею нулевой численности
на асимптотике) требует больших затрат машин-
ного времени. Поэтому ниже воспользуемся «бо-
лее слабым» аналогом явления «вымирания».
Приведем два возможных варианта.
Первый вариант основан на принципе Олли
[19]. Так, будем говорить:
Рис. 1. График функции роста водорослей R в зависи-
мости от температуры при фиксированном a и раз-
X0 вымирает, если ее численность становится
ных d.
меньше ε,
где ε - малое положительное число. В качестве
проведение соответствующих лабораторных ис-
такого «абсолютного» порога можно взять тысяч-
следований.
ную долю от среднемноголетней биомассы водо-
В рамках известной модельной схемы Контуа
рослей.
[17] динамика численности одного вида водорос-
Второй вариант опирается на относительное
лей задается базовым разностным уравнением:
соотношение численностей конкурентов в се-
мействе {X0,X1,…,Xn}.. Ниже под Xk будем подра-
xt+1 = xt[1 - α + R(θt,a,d)⋅K/(K + xt)],
(2)
зумевать не только название, но и среднюю за пе-
где xt - текущая численность водорослей; α - су-
риод (один год) численность k-й популяции в ре-
точный коэффициент смертности (α = 0.1); K -
жиме, близком к асимптотическому. Будем
величина корма (азота, фосфора и т. д.), положим
говорить, что
ниже K = 1. Шаг в модели составляет одни сутки.
X0 вытесняется, если X0/Xk < ε для некоторого k,
Отметим, что правая часть модели (2) пред-
где критическое значение ε - малая константа,
ставляет однородную функцию первой степени
например, можно положить ε = 0.1. Такого мутан-
относительно переменных x и K. Это сильно
та (с номером k) будем называть сильным. Если
сближает такие модели с линейными и упрощает
таких сильных мутантов несколько, то выбираем
исследование. В последнее время такого сорта
самого сильного, который ниже будет определять
модели приобретают все большую популярность
направление эволюционного процесса. Здесь
[18].
подразумевается достаточно монотонное асимп-
В дальнейшем ключевую роль играет понятие
тотическое поведение средних численностей по-
эволюционной устойчивости. Так, пару парамет-
пуляций, обусловленное простыми нелинейно-
стями в моделях. Но в более сложных ситуациях
ров (a*,d*) назовем эволюционно-устойчивой
реализуется колебательная динамика, и тогда
(ЭУ-параметрами), если соответствующая (ис-
сильные мутанты попеременно могут сменять
ходная) популяция не вытесняется в сообществе
друг друга. Так, в работе [8] на с. 59 показано, что
конкурентов (форм или мутантов) с близкими к
в семействе из трех групп микроорганизмов {A, B,
(a*,d*) значениями параметров. По сути, эволю-
C} имеет место неожиданная последовательность
ционная устойчивость - это способность не про-
конкурентных исходов: A вытесняет B, B вытес-
игрывать в конкурентной борьбе.
няет C, C вытесняет A. В этом редком случае нель-
Отметим, что поиск популяцией ЭУ-парамет-
зя корректно выделить сильного мутанта.
ров имеет прежде всего «оборонительное» значе-
Обычно эти различные варианты «слабого» от-
ние, но зачастую здесь реализуется и побочный
бора, заложенные в эколого-эволюционную мо-
эффект - увеличение биомассы популяции водо-
дель, приводят к близким результатам. Но в ряде
рослей. Разумеется, свойство эволюционной
случаев второй вариант оказывается предпочти-
устойчивости может зависеть от заданного в мо-
тельнее. Ниже будет использован именно второй
дели количества конкурентов. Однако, если их
вариант. Пусть (a0,d0) - исходный набор пара-
число достаточно велико, то результат эволюци-
метров температурного интервала толерантности
онного поиска, как правило, однозначен.
в исходной популяции X0. Зафиксируем некото-
Разумеется, можно модифицировать данное
рое число m - число мутантов {X1,…,X4}, каждый
определение эволюционной устойчивости отно-
из которых имеет свои параметры. Данные коэф-
сительно любого набора мутантов. При этом оче-
фициенты выбираются близкими к исходным па-
БИОФИЗИКА том 66
№ 2
2021
ГИПОТЕЗЫ ОБ АДАПТАЦИИ ВОДОРОСЛЕЙ
353
раметрам [20]. Это производится следующим об-
разом. Сначала определим величины:
a- = a0 - ξ1, a+ = a0 + ξ2, d- = d0 - ζ1, d+ = d0 + ζ2,
где все ξi и ζi - малые положительные фиксиро-
ванные числа. Их можно выбирать случайным
образом в некотором малом промежутке (0,Δ).
Считаем, что параметры мутантов следующие:
X1 → (a-,d-), X2 → (a+,d-), X3 → (a-,d+), X4 →
→ (a+,d+).
Для сокращения формул ниже будем говорить,
что (ai,di) - параметры Xi.
Рис. 2. Выделены ЭУ-параметры (a,d), полученные
Формализация «чисто» конкурентной борьбы
по модели «Адаптация».
в семействе близких популяций {X0,X1,…,X4} на
базе модели (2) задается системой:
= 14 - 10cos(2πt/T).
θt
i
i
X
=
X
[1−α+
R(θ
,a ,d
) / (1+Σ
)],
(3)
t+1
t
t
i
i
t
Здесь и всюду ниже T = 365, а t принимает дис-
0
1
4
где
Σ
= X
+X
+
+X
и i = 0.1,…,4. При фик-
кретные значения от 1 до 365.
t
t
t
t
сированных параметрах производим прогноз на
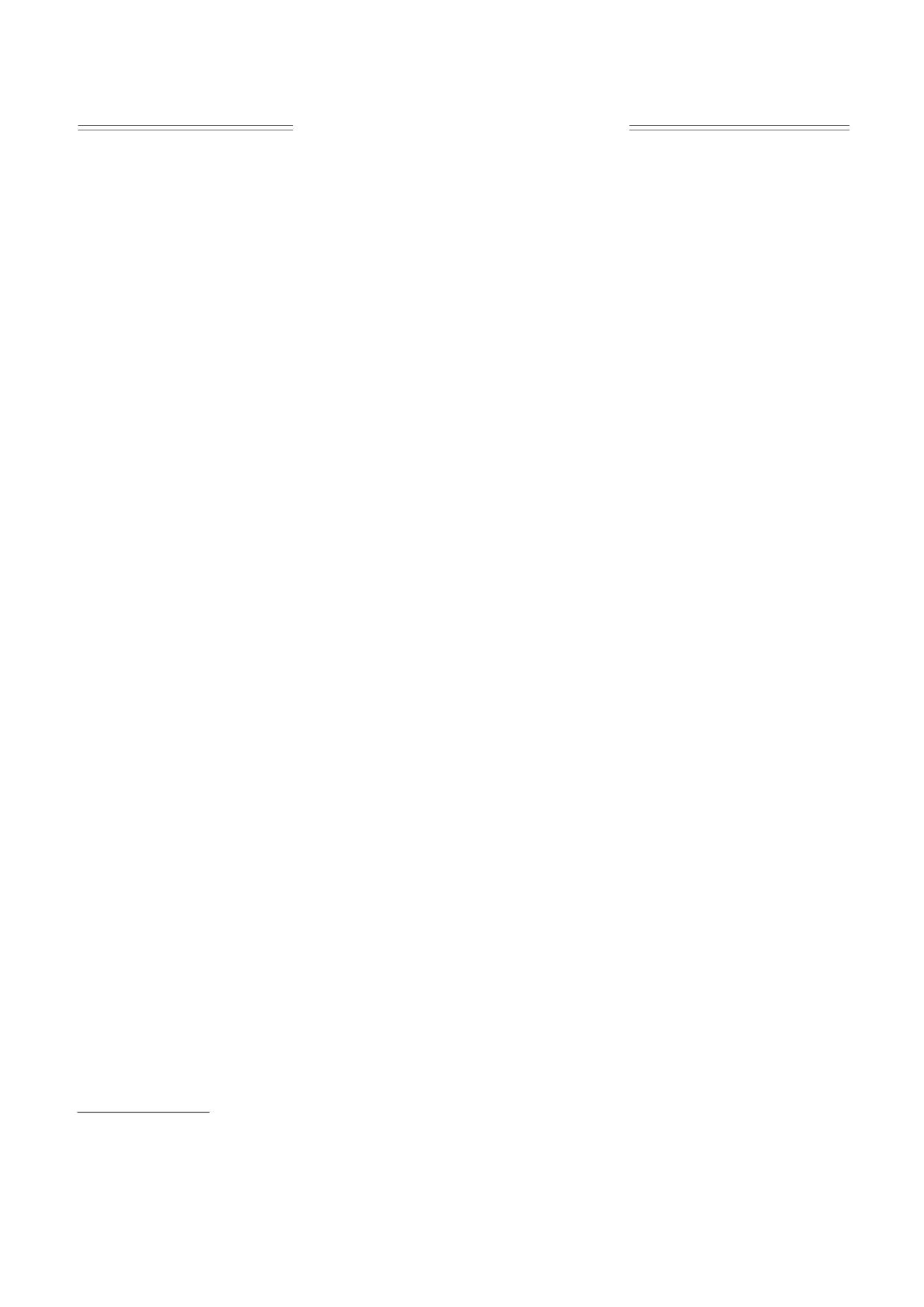
Актуальна задача: какие пары (a*,d*) оказыва-
длительный период времени (20 лет). Здесь могут
ются ЭУ-параметрами? При решении данной за-
возникать две ситуации:
дачи задаем стартовые значения (a,d), а затем под
1. «Плохая», когда исходная популяция выми-
действием модели «Адаптация» происходит изме-
рает (в слабом смысле). Тогда (a0,d0) заведомо не
нение переменных и медленное изменение пара-
являются ЭУ-параметрами, и их следует модифи-
метров. Полученные в результате расчетов на
цировать. Так, пусть, например, у первого мутан-
многолетний период финальные значения явля-
та наибольшая средняя за год численность среди
ются ЭУ-параметрами. При компьютерных экс-
остальных мутантов, тогда построим новые пара-
периментах были испробованы различные точки
метры исходной популяции:
старта из области параметров (AD): 4 ≤ a ≤ 24 и
(1 - ε)a0 + εa1 и (1 - ε)d0 + εd1,
(4)
1 ≤ d ≤ 11. Оказывается, что финальных пар (ЭУ-
параметров) совсем немного, и они образуют (см.
где малый положительный параметр ε можно
рис. 2) «радугу» (с малой сердцевиной).
считать скоростью микроэволюции. Фактически
Конструкция «Адаптация» позволяет выявить
формулы (4) означают малый сдвиг исходных па-
еще одно любопытное обстоятельство. Так, каж-
раметров в сторону параметров самого сильного
дая начальная точка - узел решетки AD - притя-
мутанта. Разумеется, это лишь одно из возмож-
гивается к своей паре ЭУ-параметров, лежащей
ных направлений эволюционного процесса.
на «радуге». Какие же точки «радуги» являются
2. «Хорошая», когда исходная популяция не
наиболее «притягательными»?
вымирает. Значит, она конкурентоспособна в
рамках заданного набора мутантов.
Ими оказались три пары параметров: Mini =
= (5, 1), Maxi = (23, 1) и Midl = (14, 10). Точнее, не
Теперь следует произвести новую генерацию
сами точки, а их малые окрестности. Строго по-
группы мутантов с новыми параметрами, близки-
казано [21, 22], что в периодической среде множе-
ми к коэффициентам исходной, например, путем
ство ЭУ-параметров плотное, т.е. может содер-
случайного выбора каждого коэффициента в ма-
жать целые окрестности
лом интервале
«вариабельности»
(0,Δ). Затем
проводим очередное конкурентное испытание на
Естественно, точка Mini соответствует холод-
20 лет. Если таких (одновременно хороших) ис-
нолюбивым (например, диатомовым), а точка
пытаний накопилось достаточно много (напри-
Maxi - теплолюбивым (например, синезеленым)
мер, 100 испытаний), то можно с большой долей
водорослям. В лабораторных экспериментах,
уверенности считать данные коэффициенты ЭУ-
действительно, обнаружен бимодальный харак-
параметрами.
тер температурной адаптации: «любимые темпе-
В качестве важного примера обсудим ситуа-
ратуры» бывают двух типов - достаточно высокие
цию, когда внутригодовая температура изменяет-
и довольно низкие [9]. Вероятно, это эволюцион-
ся по фиксированному закону в пределах от 4
ные реликты прошлых долговременных измене-
до 24:
ний климата - потепления и похолодания.
БИОФИЗИКА том 66
№ 2
2021
354
ИЛЬИЧЕВ, ИЛЬИЧЕВА
В отличие от этих стеногалинных (узкоспеци-
Заметим, что в работе [23] приведена непре-
ализированных) водорослей, третья точка Midl
рывная модельная схема адаптации параметра а,
характерна для эвригалинных (широкоспециали-
включающая в себя основную популяцию и двух
зированных) видов. Хотя они редко наблюдаются
мутантов. При малом d тестовые расчеты показа-
в Азовском море, но возможно этот (до поры до
ли, что в зависимости от выбора начального зна-
времени) скрытый вид является эволюционным
чения ЭУ-параметр а оказывается, как правило,
резервом азовского фитоценоза. А именно, в слу-
вблизи одного из критических значений (локаль-
чае многолетнего апериодического (внутригодо-
ных экстремумов) θt. Обнаружены и гораздо «ме-
вого) температурного режима он будет иметь кон-
нее притягательные» ЭУ-параметры а - это неко-
курентное преимущество. Напротив, если перио-
торые точки перегиба θt. В частности, значение 14
дический температурный режим в точности
соответствует точке перегиба данной синусои-
повторяется из года в год, то конкурентное пре-
дальной температурной кривой. В целом резуль-
имущество принадлежит стеногалинным видам.
таты по непрерывной и дискретной схемам прак-
Отметим, что нелинейность Контуа в модели
тически совпадают.
(3) допускает те или иные модификации. В этом
случае трансформируется и соответствующее
множество ЭУ-параметров. Однако если эти ва-
ОДНОВРЕМЕННАЯ АДАПТАЦИЯ
риации не слишком сильные, то указанные выше
ВОДОРОСЛЕЙ К СОЛЕВОМУ
точки («три богатыря») Mini, Maxi и Midl всегда в
И ТЕМПЕРАТУРНОМУ РЕЖИМАМ СРЕДЫ
нем присутствуют.
Несколько более реалистичным представляет-
Соленость и рост. Кратко обсудим адаптацию
ся следующее задание температуры среды:
водорослей к солености среды. Функционально
влияние солености на динамику фитопланктона
θt = 14 - 10cos(2πt/T) + h(t),
подобно действию температуры. Так, каждый вид
где h(t) - случайная величина (белый шум) с рав-
водорослей обладает своим интервалом солевой
номерным распределением (нулевым средним и
толерантности J(b,c) = [b - c, b + c], в пределах ко-
заданной дисперсией σ). В этом случае ширина
торого они способны развиваться. При описании
«эволюционного оптимального» интервала толе-
скорости роста используем прежнюю гипотезу
рантности будет увеличиваться с ростом σ.
Пианки:
2
2
[c
−
(S -b)
]
ρ(S,b,c)
=
r
при
S
∈
J(b,c), иначе
r
=
0.
3
c
ческое изменение температуры и солености на-
Здесь r - некоторая константа суточного роста
кладывается «белый шум» с небольшой диспер-
водорослей.
сией:
Разумеется, по-прежнему актуальна проблема
θt = 14 - 10cos(2πt/T) + hθ(t)
поиска ЭУ-параметров (b,c). С использованием
и St = 7 - 3cos(2πt/T - ϕ) + hS(t),
(5)
схемы адаптации из предыдущего параграфа
установлено, что и в данном случае наиболее
где hθ(t) и hS(t) - случайные величины с равно-
«притягательными» оказываются коэффициенты
мерным распределением; у обоих нулевое сред-
b, близкие к экстремальным значениям (миниму-
нее и дисперсии 1 и 1/2 соответственно. Здесь ϕ -
мы или максимумы) температурной кривой с ма-
возможный временной сдвиг в годовом поведе-
лым коэффициентом с. Согласно работе [8] это
нии указанных факторов. Например, при ϕ = 0
так называемая гипотеза критических значений.
имеет место синхронная динамика, а при ϕ = π -
Например, для периодического солевого ре-
противоположное изменение (увеличение одного
жима водоема, «похожего» на Таганрогский за-
фактора сопровождается уменьшением другого).
лив, St = 7 - 3cos(2πt/T) найдены два «наиболее
Совместный учет данных факторов на рост водо-
притягательных» параметра: b = 4.5 и b = 9.5 при
рослей может быть реализован многими способа-
c = 0.5. Они характерны для разных стеногалин-
ми. Ниже выбрана мультипликативная версия
ных видов - пресноводных синезеленых и мор-
функции роста (Ω):
ских диатомовых водорослей. Реже возникает па-
Ω(θ,S,a,d,b,c) = R(θ,a,d) ρ(S,b,c).
ра ЭУ-параметров b = 7.0 при c = 3.5, соответству-
ющая эвригалинной водоросли.
Иногда среднегодовое значение Ω может ока-
Модификация модели «Адаптация». Обсудим
заться слишком малым, тогда популяция будет
поиск ЭУ-параметров (a,d,b,c), когда на периоди- вымирать. Для сохранения ее живучести в модели
БИОФИЗИКА том 66
№ 2
2021
ГИПОТЕЗЫ ОБ АДАПТАЦИИ ВОДОРОСЛЕЙ
355
следует выбирать достаточно большое значение r.
Далее полагаем r = 20.
Теперь базовая модель динамики одной попу-
ляции водорослей представляется в форме:
xt+1 = xt[1 - α + Ω(θt,S t,at,dt,bt,ct)/(1 + xt)].
(6)
На ее основе построим модифицированную
адаптивную конструкцию, в которой наряду с ис-
ходной популяцией задано и 16 мутантов со свои-
ми параметрами. Кратко ее суть заключается в
следующем. Пусть (a0,d0,b0,c0) - текущие (базо-
вые) параметры исходной популяции. Обозначим
параметры мутантов через:
Рис. 3. ЭУ-параметры (a,b) при синхронном измене-
a- = a0 - δ1, a+ = a0 + δ2,
нии температуры и солености. Структура типа «воз-
растающий отрезок».
d- = d0 - η1, d+ = d0 + η2,
b- = b0 - δ3, b+ = b0 + δ4,
(7)
представительным спектром параметров. Тесто-
вые прогнозы продемонстрировали слабую зави-
c- = c0 - η3, c+ = c0 + η4,
симость области ЭУ-параметров от m.
где все δi и ηi - положительные числа из малого
Определенную роль играет и величина «вариа-
интервала «вариабельности» (0,Δ). Их можно вы-
бельности» Δ. Чем она больше, тем уже множе-
бирать случайным образом.
ство ЭУ-параметров.
Параметры мутантов представляют собой
Приведем результаты отдельных тестовых рас-
четов по данной модели. Во-первых, параметры d
комбинации данных вариаций от (a-,d-,b-,
и c (ширина интервала толерантности) оказались
c-) до (a+,d+,b+,c+). Удобно их обозначить
малыми величинами. Во-вторых, подозреваем,
через (a1,d1,b1,c1) и т.д. вплоть до
что все возможные пары ЭУ-параметров a и b об-
(a16,d16,b16,c16). Теперь модель конкурентно-
разуются как комбинации (прямое произведе-
ние) критических значений температуры и соле-
го сообщества «исходная популяция + мутан-
ности. Следовательно, кажется, что таких пар
ты» имеет следующий вид:
ЭУ-параметров будет не так уж много. Однако не-
i
i
ожиданно возникла иная ситуация.
X
=
X
[1−α+Ω(θ
,S
,a ,
d b,c
) / (1+Σ
)],
(8)
t+1
t
t
t
i
i
i
i
t
Напомним здесь возможную стратегию расче-
0
1
16
где
Σ
= X
+X
+X
и i = 0,1,…,16. Здесь схема
тов - это задание решетки в пространстве пара-
t
t
t
t
адаптации будет аналогична предыдущей, а
метров (a,d,b,c). Далее, узлы решетки выбирают-
именно, если при фиксированных коэффициен-
ся в качестве начальных параметров, затем про-
тах в модели (8) выявляется сильный мутант (на-
слеживается их судьба вплоть до достижения ими
пример, первый), происходит пересчет базовых
финальных значений. В результате в четырехмер-
параметров по формулам:
ном пространстве возникает область ЭУ-пара-
метров. Разумеется, такая полная программа ис-
(1 - ε)a0 + εa1, (1 - ε)d0 + εd1
следований требует больших затрат машинного
и (1 - ε)b0 + εb1, (1 - ε)c0 + εc1,
времени.
Поэтому воспользуемся ниже укороченной
где скорость микроэволюции ε - малый положи-
схема расчетов. Считаем, что старт «второстепен-
тельный параметр.
ных» параметров (d,c) производится с фиксиро-
После этого согласно формулам (7) строятся
ванных низких значений, например, с 2, а старт
коэффициенты мутантов.
начальных «первостепенных» параметров (a,b)
Если выявить сильного мутанта не удалось, то
происходит из узлов решетки [4,24]´[4,10] c ша-
производим новую случайную генерацию пара-
гом 2. Далее прослеживается микроэволюция
метров всех мутантов и т.д.
всех четырех параметров. Приведем основные ре-
зультаты расчетов.
Отметим, что данная схема адаптации предъ-
являет более жесткие требования к эволюцион-
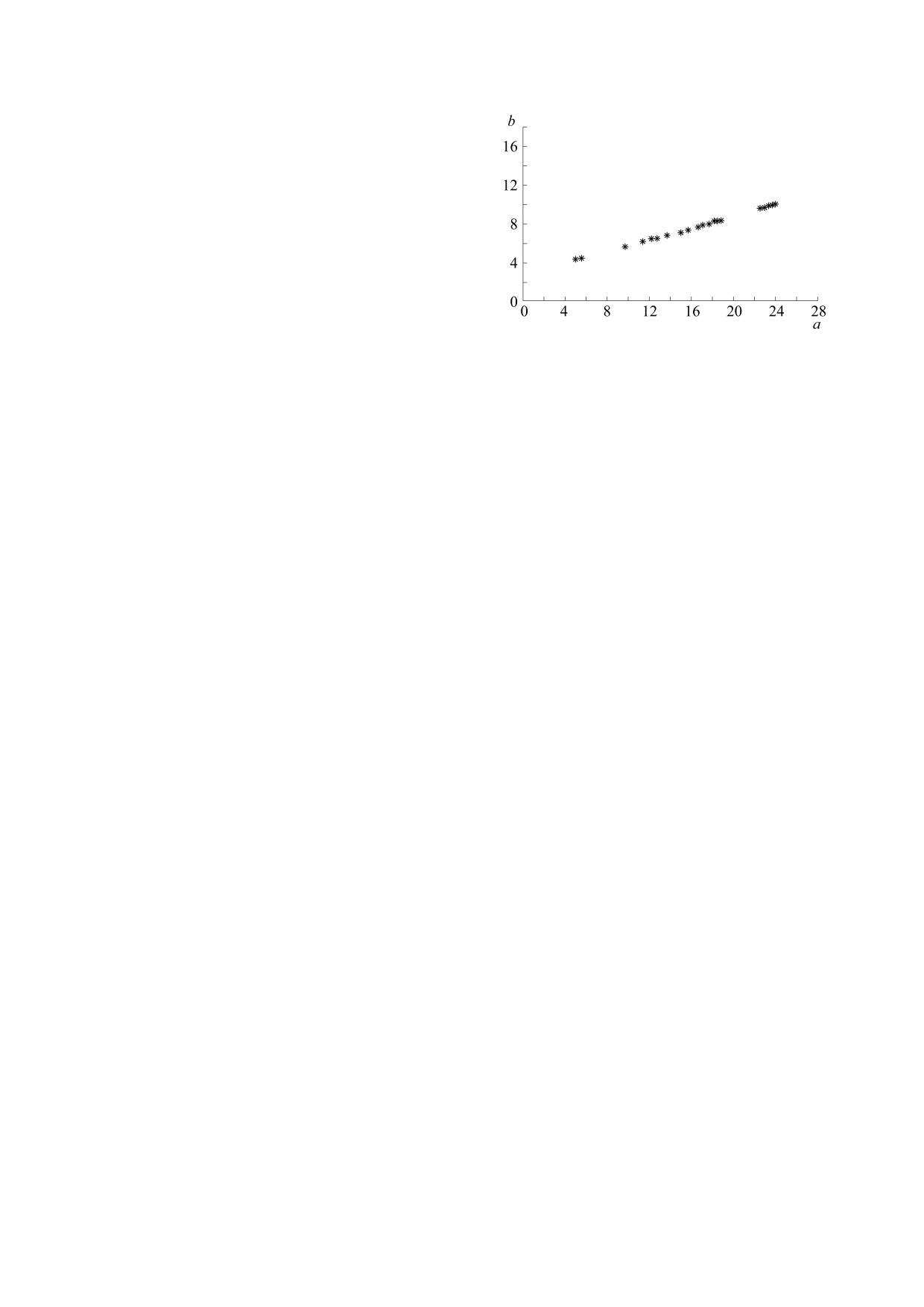
Синхронное поведение факторов. Формально
ной устойчивости: чем больше мутантов, тем
это равносильно тому, чтобы положить в динами-
труднее исходной популяции выдержать их кон-
ке факторов (5) сдвиг ϕ = 0. Оказалось, что все
курентное давление. Разумеется, можно задать и
ЭУ-параметры (a,b) располагаются на возрастаю-
другое количество (m) мутантов с достаточно
щем отрезке (см. рис. 3), соединяющем точки
БИОФИЗИКА том 66
№ 2
2021
356
ИЛЬИЧЕВ, ИЛЬИЧЕВА
Рис. 4. ЭУ-параметры (a,b) при строго асинхронном
Рис. 5. ЭУ-параметры (a,b) при «ортогональном»
изменении температуры и солености. Структура типа
изменении температуры и солености. Структура типа
«убывающий отрезок».
«овал».
(4,4) и (24,10). При этом финальные параметры d
том (против часовой стрелки) областей ЭУ-пара-
и c оказались близки к единице.
метров.
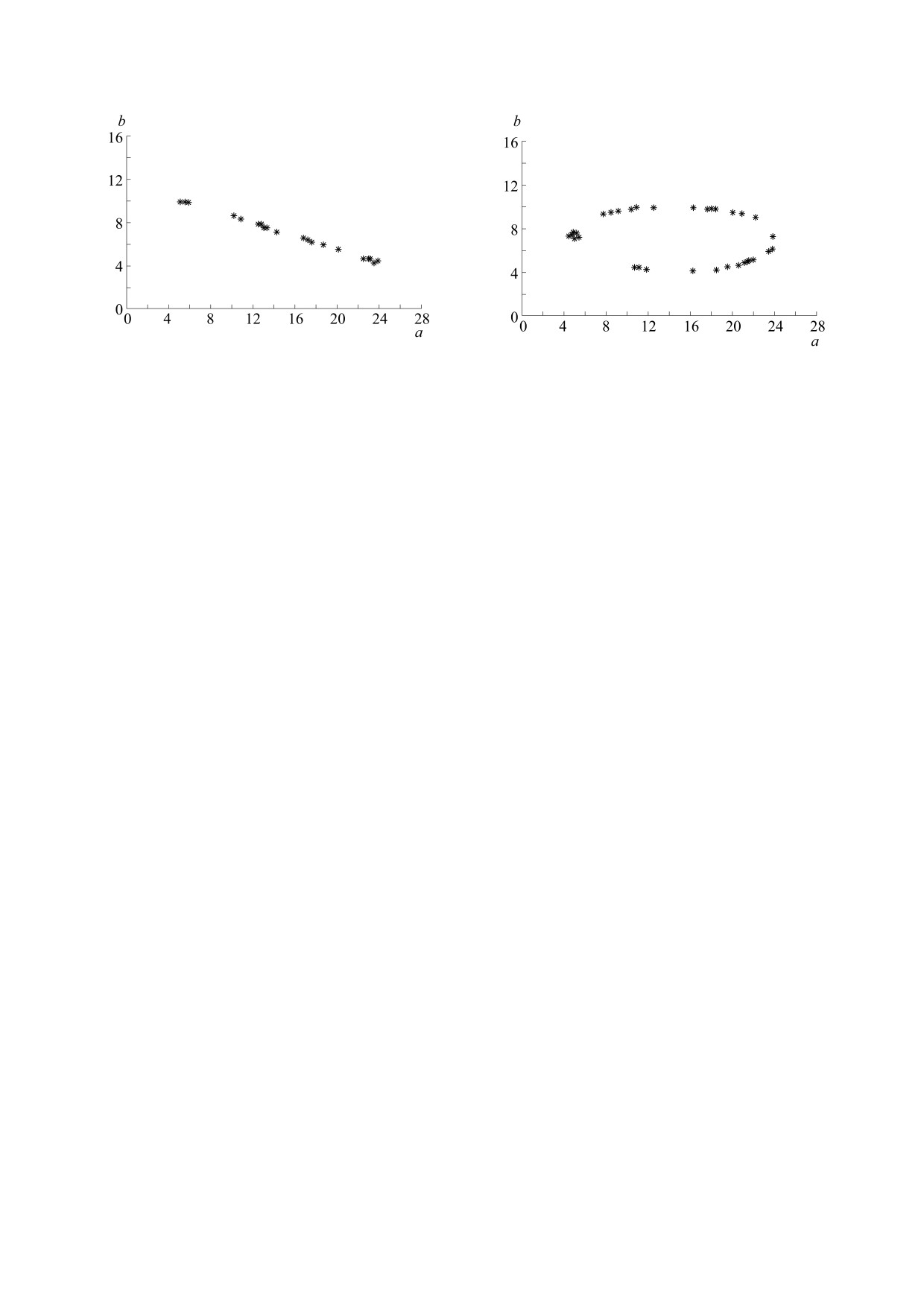
Строго асинхронное поведение факторов. Здесь
Представляет интерес аналогичный поиск ЭУ-
в динамике температуры и солености (5) полага-
параметров, когда скорость роста водорослей за-
ем ϕ = π. В этом случае ЭУ-параметры образуют
дается иным образом. Так, весьма популярно сле-
убывающий отрезок (см. рис. 4) с вершинами
дующее задание этой функции (принцип узкого
(4,10) и (24,4).
места):
«Ортогональное» поведение факторов. Полага-
Ω(θ,S,a,d,b,c) = min{R(θ,a,d) ρ(S,b,c)}.
ем ϕ = π/2 - это промежуточный вариант в свете
Расчеты показали, что теперь семейство ЭУ-
предыдущих экспериментов. Так, на одних вре-
параметров (a,b) похоже на предыдущее, но явля-
менных отрезках данные факторы ведут себя син-
ется куда более «тощим». Так, при синхронном
хронно, а на других - асинхронно. Поэтому неяс-
изменении температуры и солености остаются
но, как будет здесь располагаться множество ЭУ-
лишь две окрестности точек - (4,4) и (24,10). При
параметров. Оказалось, что здесь ЭУ-параметры
асинхронном поведении указанных факторов
лежат на некотором овале, вписанном в дискрет-
остаются три окрестности: (4,10), (14,7) и (24,4).
ную решетку начальных значений (см. рис. 5). Ве-
роятно, ЭУ-параметры не заполняют сплошь ука-
занные одномерные структуры на рис. 3-5. Грубо
ЗАКЛЮЧЕНИЕ
говоря, появление «дыр» неизбежно!
На основе идей эволюционного моделирова-
Любопытно, что при ϕ = 0 и ϕ = π множества
ния предложены новые быстродействующие схе-
ЭУ-параметров являются линейными структура-
мы адаптации параметров популяций. Показано,
ми - отрезками. Кажется «вполне естественным»:
что эволюционно-устойчивые параметры роста
при ϕ = π/2 множество ЭУ-параметров является
водорослей составляют «тощее» множество, а его
«полусуммой» антисимметричных отрезков ВО и
геометрическое расположение тесно связано с
УО. Возникает парадоксальная ситуация: итог
критическими точками (значениями минимумов
может зависеть от способа усреднения - по вер-
и максимумов) динамики факторов среды.
тикали или по горизонтали. Поэтому «естествен-
Разумеется, возможна и целесообразна экспе-
ный» подход приводит к разным построениям и
риментальная проверка полученных результатов.
не годится. Теперь не удивительно, что компро-
Так, в проточном культиваторе можно искус-
миссом оказывается нелинейный объект (овал).
ственно поддерживать постоянное содержание
Обсудим данные результаты с точки зрения
биогенных веществ и задавать определенный пе-
непрерывной деформации угла ϕ на участке
риодический температурный режим. Далее, в
этот аквариум можно засеять тот или иной вид
I = [0,π]. Путем дополнительных расчетов уста-
новлено, что множество ЭУ-параметров подвер-
водорослей и после длительных наблюдений оце-
гается двум «испытаниям»: при росте ϕ от 0 до π/2
нить итоговые интервалы температурной толе-
происходит постепенное «раздувание» отрезка
рантности.
ВО в овал, а при увеличении ϕ до π овал медленно
Обнаружено, что в процессе адаптации водо-
сплющивается в отрезок УО. В целом данные
рослей существенную роль играет взаимное пове-
процессы сопровождаются медленным поворо-
дение факторов среды. Здесь могут возникать как
БИОФИЗИКА том 66
№ 2
2021
ГИПОТЕЗЫ ОБ АДАПТАЦИИ ВОДОРОСЛЕЙ
357
линейные (отрезки), так и нелинейные (овалы)
6. J. B. S. Holdane, The Causes of Evolution (Longmans,
геометрические области ЭУ-параметров.
Lond., 1932).
7. R. A. Fisher, The genetical theory of natural selection
(Clarendon Press, Oxford, 1930).
ФИНАНСИРОВАНИЕ РАБОТЫ
8. В. Г. Ильичев, Устойчивость, адаптация и управле-
ние в экологических системах (Физматлит, М.,
Публикация подготовлена в рамках реализа-
2009).
ции Государственного задания ЮНЦ РАН (про-
9. В. Б. Вербицкий, Дис. … д-ра биол. наук (Институт
ект АААА-А18-118122790121-5).
биологии внутренних вод, Борок, 2012).
10. Н. Н. Яковлев, Живое и среда (Наука, Л., 1986).
11. Тh. Dobzhansky, Genetics of the Evolutionary Process
КОНФЛИКТ ИНТЕРЕСОВ
(Columbia Univ. Press, N.Y., 1970).
Авторы заявляют об отсутствии конфликта
12. В. Г. Ильичев и О. А. Ильичева, Биофизика 63 (2),
интересов.
373 (2018).
13. Ю. М. Свирежев и В. П. Пасеков, Основы матема-
тической генетики (Наука, М., 1982).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
14. J. M. Smith, Evolution and the theory of games (Cam-
bridge Univ. Press, N.Y., 1982).
Настоящая работа не содержит описания ис-
следований с использованием людей и животных
15. Э. Пианка, Эволюционная экология (Мир, М., 1981).
в качестве объектов.
16. Ю. Н. Сергеев, А. А. Колодочка, Х. Д. Круммель
и др., Моделирование процессов переноса и транс-
формации вещества в море (ЛГУ, Л., 1979).
СПИСОК ЛИТЕРАТУРЫ
17. В. Е. Contois, J. Gen. Microbiol. 21, 40 (1959).
18. Ю. В. Тютюнов и Л. И. Титова, Журн. общ. биоло-
1. А. М. Бронфман и Е. П. Хлебников, Азовское море.
гии 79 (6), 448 (2018).
Основы реконструкции (Гидрометеоиздат, Л.,
19. W. C. Alle, A. E. Emerson, D. Park, and K. Smith,
1985).
Principies of Animalls Ecology (W. B. Saunders Co,
2. Г. Ю. Ризниченко и А. Б. Рубин, Биофизическая ди-
Philadelphia, 1949).
намика продукционных процессов (ИКИ, Ижевск,
20. В. Г. Ильичев, Л. В. Дашкевич и В. В. Кулыгин,
2004).
Природа, № 8, 54 (2019).
3. А. Б. Медвинский, С. Е. Петровский, И. А. Тихо-
21. В. Г. Ильичев, Автоматика и телемеханика, № 11,
нова и др., Успехи физ. наук 172 (1), 31 (2002).
115 (1996).
4. V. A. Kostitzin, Symbios, parazitisme of evolution (Her-
22. В. Г. Ильичев, Биофизика 50 (3), 567 (2005).
mann, Paris, 1934).
23. В. Г. Ильичев, В. В. Кулыгин и Л. В. Дашкевич,
5. В. Вольтерра, Математическая теория борьбы за
Компьютерные исследования и моделирование 9
существование (Наука, М., 1976).
(6), 981 (2017).
Hypotheses Concerning Algal Adaptation to Periodic Environmental Factors
V.G. Ilyichev* and O.A. Ilyicheva**
*Southern Scientific Center, Russian Academy of Sciences, ul. Chekhova 41, Rostov-on-Don, 344006 Russia
**Don State Technical University, ul. Socialisticheskaya 162, Rostov-on-Don, 344022 Russia
A new ecological-evolutionary model of small-sized algae has been proposed to describe the simultaneous
dynamics of variables (population biomass) and parameters representing temperature tolerance limits [a - d,
a + d]. In the given periodic temperature regime, the parameters tend to some final, so-called evolutionarily
stable values. Calculation results showed that on the plane (a,d) all evolutionarily stable parameters are locat-
ed in the form of a “rainbow”, the farthest points of which correspond to cold-loving (diatomic) and heat-
loving (blue-green) algae. Analysis of data relating to algal adaptation to two periodic environmental factors
(temperature and salinity) within given ranges of temperature [a - d, a + d] and salinity [b - c, b + c] toler-
ance is performed. It has been established that the domain of evolutionarily stable parameters in the plane
(a,b) depends on the degree of synchronism of environmental factors. In some cases, these parameters lie on
a linear segment, and in other cases, their unexpected location on a nonlinear oval is possible.
Keywords: model, adaptation, evolutionarily stable parameters, temperature, salinity
БИОФИЗИКА том 66
№ 2
2021