БИОФИЗИКА, 2021, том 66, № 2, с. 358-364
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 577.3
ТЕМПЕРАТУРНАЯ ЗАВИСИМОСТЬ МЕХАНИЧЕСКИХ ХАРАКТЕРИСТИК
ДЕМЕМБРАНИЗОВАННЫХ МЕДЛЕННЫХ МЫШЕЧНЫХ
ВОЛОКОН КРОЛИКА
© 2021 г. С.Р. Набиев*, С.Ю. Бершицкий*, А.К. Цатурян**, Н.А. Кубасова**
*Институт иммунологии и физиологии УрО РАН, 620049, Екатеринбург, ул. Первомайская, 106
**Институт механики Московского государственного университета имени М.В. Ломоносова,
119192, Москва, Мичуринский просп., 1
E-mail: natalia@imec.msu.ru
Поступила в редакцию 04.12.2020 г.
После доработки 05.12.2020 г.
Принята к публикации 22.12.2020 г.
Изучены температурные зависимости напряжения и жесткости активно сокращающихся одиноч-
ных волокон медленной (m. soleus) мышцы кролика с помощью джоулевого скачка температуры.
Показаны различия в характеристиках волокон быстрой (m. psoas) и медленной (m. soleus) мышцы
кролика, вызванные, по-видимому, различиями в кинетике цикла миозиновой головки.
Ключевые слова: мышечное сокращение, изометрическое напряжение, скачок температуры.
DOI: 10.31857/S0006302921020174
Основополагающие работы в области исследо-
жения происходит в два шага. Сначала миозино-
вания механизма мышечного сокращения были
вая головка слабо присоединяется к актину, не
сделаны на быстрых волокнах скелетных мышц.
участвуя в генерации силы, но внося вклад в
Было показано, что развитие активной силы про-
жесткость мышцы, затем она «застегивается», т.е.
исходит за счет движения миозиновой головки,
переходит в прочно связанное состояние и уже в
присоединенной к актину, либо поворота части
ходе этого перехода развивает некоторую силу, и
головки, так называемого рычага, относительно
лишь на следующем шаге цикла взаимодействия с
ее актинсвязывающего домена, и предложены со-
актином происходит поворот рычага. Таким об-
ответствующие модели [1, 2]. Казалось бы, вели-
разом, сила, развиваемая мышцей в изометриче-
чина развиваемой силы должна быть прямо про-
ском сокращении, пропорциональна не общему
порциональна числу присоединенных к актину
количеству присоединенных головок, а лишь
миозиновых головок, и это число можно оценить
числу тех из них, которые присоединены в по-
по динамической (т.е. измеренной при быстрых
следнем состоянии с повернутым рычагом, а
изменениях длины) жесткости мышечного во-
жесткость - числу всех присоединенных головок.
локна. Однако эксперименты на активно сокра-
щающихся волокнах быстрых скелетных мышц,
В то же самое время исследование механики
выполненные при различных значениях темпера-
актин-миозинового взаимодействия в медленных
туры, показали, что наблюдаемый значительный
скелетных мышцах, ничуть не менее важных с
рост силы с ростом температуры - четырех-пяти-
физиологической точки зрения, носило лишь
кратный в мышце кролика при нагревании от 5°С
фрагментарный характер. В экспериментах на во-
до 35°С [3, 4] и в 1.7 раза при увеличении темпера-
локнах медленных мышц, часто проводившихся
туры мышцы лягушки от 4°С до 17°С [5, 6] - не
параллельно с экспериментами на волокнах
сопровождается, либо сопровождается неболь-
быстрых мышц этих же животных, были получе-
шим, не более 15%, ростом жесткости [3-6]. Это
ны классические биофизические характеристи-
наблюдение, подкрепленное анализом измене-
ки: кривая «сила-скорость» для волокон мышц
ний на рентгенодифракционных диаграммах ак-
морской свинки [8] и температурная зависимость
тивно сокращающихся мышечных волокон кро-
кальциевой чувствительности силы для мышц
лика в ответ на скачок температуры, привело к
крысы [9, 10]. Похожие эксперименты были вы-
созданию новой структурно-кинетической моде-
полнены примерно в то же время и в других лабо-
ли актин-миозинового взаимодействия [7]. Со-
раториях [11-13]. Было высказано предположе-
гласно этой модели, развитие активного напря-
ние, что наблюдаемые различия механических
358
ТЕМПЕРАТУРНАЯ ЗАВИСИМОСТЬ МЕХАНИЧЕСКИХ ХАРАКТЕРИСТИК
359
свойств медленных и быстрых волокон скелетных
мышц могут быть вызваны различием изоформ-
ного состава тяжелых цепей миозина в быстрых и
медленных мышцах с различной кинетикой их
рабочего цикла [14].
В сравнительных экспериментах на волокнах
мышц человека, содержащих различный набор
тяжелых цепей миозина, с использованием усо-
вершенствованной аппаратуры (более быстрый
мотор для изменения длины мышцы, динамиче-
ский контроль изменения длины саркомеров)
были измерены не только характеристики стаци-
онарной связи сила-скорость при укорочении и
растяжении этих мышц, но и их кинетические ха-
рактеристики - скорость восстановления силы
после малых и больших ступенчатых изменений
длины и динамическая жесткость волокон при
изометрических сокращениях и при растяжении
с различной скоростью [15]. Были выявлены су-
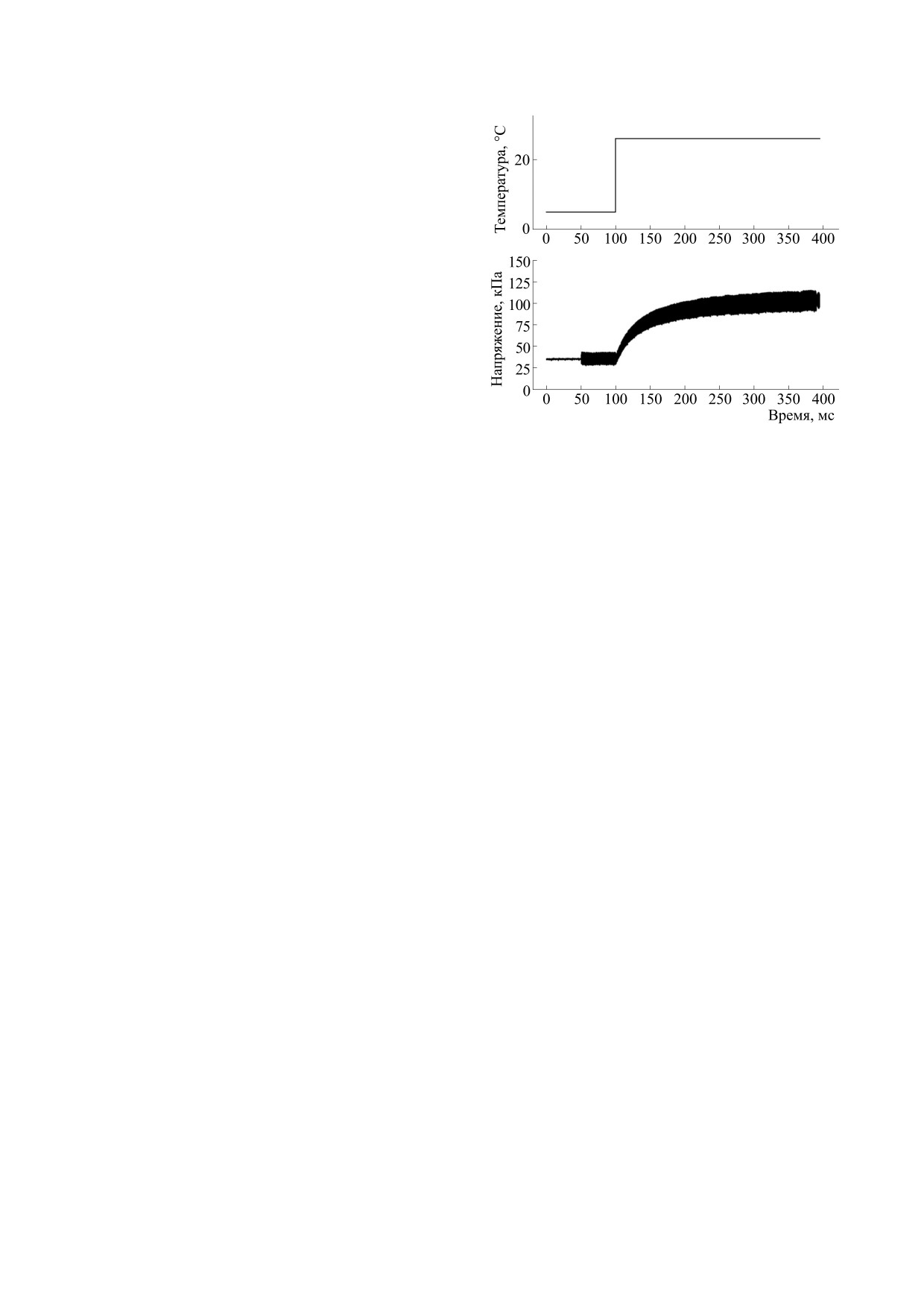
Рис. 1. Экспериментальный протокол. Синусоидаль-
щественные различия свойств медленных и быст-
ные изменения длины прикладывали к волокну за
рых мышц. Однако в этих экспериментах дина-
50 мс до скачка температуры. Продолжительность за-
мическая жесткость была измерена лишь при од-
писи варьировали так, чтобы было достигнуто уста-
ном значении температуры (12°С), далеком от
новившееся значение напряжения при высокой тем-
пературе.
физиологической для теплокровных. Темпера-
турная зависимость изометрического напряже-
ния и динамической жесткости волокон камбало-
ческой жесткости медленных волокон при более
видной (медленной) мышцы человека была ис-
высоких значениях температуры вплоть до фи-
следована в интервале температур 5-20°С [16].
зиологической.
Было показано, что динамическая жесткость во-
локон медленных мышц меняется в этом интер-
вале температур значительно сильнее, чем воло-
МАТЕРИАЛЫ И МЕТОДЫ
кон быстрых мышц. Следует заметить, что экспе-
Эксперименты были выполнены на лабора-
риментальные образцы волокон мышц человека
торной установке, основные узлы которой описа-
не относились к чистым быстрым и медленным
ны в работах [4, 18]. В качестве объекта исследо-
типам мышц, а содержали небольшую долю дру-
вания нами были выбраны демембранизованные
гих изоформ миозина [15].
медленные мышечные волокна из камбаловид-
Новые эксперименты на одиночных волокнах
ной (m. soleus) мышцы кролика. Состав раство-
камбаловидной мышцы кролика [17] подтверди-
ров, методика выделения и работы с одиночным
ли выводы, сделанные в работе [16]. Гель-элек-
мышечным волокном описаны в работе [19]. По-
трофорезный анализ показал, что содержание
сле завершения экспериментов волокна подвер-
быстрых изоформ миозина в волокнах m. soleus
гали гель-электрофорезному анализу, показыва-
кролика пренебрежимо мало. Было показано, что
ющему содержание в волокне различных изо-
при постоянной температуре 12°С увеличение
форм миозина. Во всех экспериментальных
уровня кальциевой активации волокна вело к
образцах была обнаружена только тяжелая цепь
увеличению как изометрического напряжения,
миозина 1 - медленная изоформа миозина.
так и жесткости полусаркомера. Авторами была
Волокно, закрепленное одним концом на ли-
предложена модель, объясняющая такое поведе-
нейном моторе, а другим - на датчике силы, пе-
ние напряжения и жесткости. Одно из важней-
ремещали из камеры с расслабляющим раство-
ших предположений модели состояло в том, что
ром в камеру с активирующим раствором при
жесткость одиночной миозиновой головки в во-
температуре 2°С, где его выдерживали около 5 с.
локнах медленной мышцы в три раза меньше, чем
Активированное волокно перемещали в воздуш-
в быстрой. Этот вывод согласуется с прямыми из-
ную камеру с температурой 5°C, где к волокну бы-
мерениями жесткости одиночных миозиновых
ли приложены синусоидальные изменения дли-
головок в оптической ловушке [16].
ны и джоулев скачок температуры (рис. 1). Ам-
Поскольку во всех вышеописанных экспери-
плитуда скачка температуры составляла от 15°С
ментах температура не превышала 20°С, нам по-
до 30°С. Скачок температуры получали пропуска-
казалось важным дополнить исследования тем-
нием высоковольтного импульса переменного
пературной зависимости напряжения и динами-
тока частотой 40 кГц и длительностью 0.2 мс. До-
БИОФИЗИКА том 66
№ 2
2021
360
НАБИЕВ и др.
Рис. 2. Изменение напряжения волокна в ответ на скачок температуры: (а) - абсолютные значения напряжения до
(5°С) и после скачка температуры; (б) - те же значения, нормированные на соответствующее напряжение до скачка
температуры. Разные символы соответствуют разным волокнам (n = 6). Светлые и темные символы соответствуют
двум сериям экспериментов.
полнительно для компенсации остывания волок-
Значения стационарного напряжения при раз-
на прикладывали малоамплитудный импульс пе-
личных значениях финальной температуры пока-
ременного тока продолжительностью до 400 мс.
заны на рис. 2. Видно, что основные изменения
Частота синусоидальных изменений длины во-
напряжения происходили в интервале темпера-
локна составляла 1.6 кГц, амплитуда - 0.1% дли-
тур от 5°С до 20°С, а в диапазоне 20-37°С измене-
ны волокна. Все измерения на волокне происхо-
ния были незначительны.
дили под контролем длины саркомеров, началь-
Абсолютные и нормированные на соответ-
ная длина саркомеров в центральной части
ствующее значение при 5°С величины напряже-
волокна была 2.4-2.5 мкм. К сожалению, не уда-
ния и жесткости, наблюдавшиеся в эксперимен-
лось получить достоверного сигнала изменений
тах, сведены в группы по значениям температу-
длины саркомеров в ответ на синусоидальные из-
ры, достигнутой после скачка, и показаны в
менения длины волокна до и после скачка темпе-
таблице. Видно, что разброс абсолютных значе-
ратуры. Количество циклов «активация-расслаб-
ний напряжения и жесткости был достаточно
ление» одного волокна составляло от 6 до 12.
большим, но после нормирования разброс суще-
ственно уменьшался. Изменения жесткости на-
Для обработки результатов экспериментов
блюдали при изменении температуры от 5°С до
была написана программа на языке Python. Жест-
20°С, в дальнейшем жесткость оставалась посто-
кость оценивали как отношение амплитуд си-
янной в пределах статистической погрешности.
нусоид, аппроксимирующих изменения на-
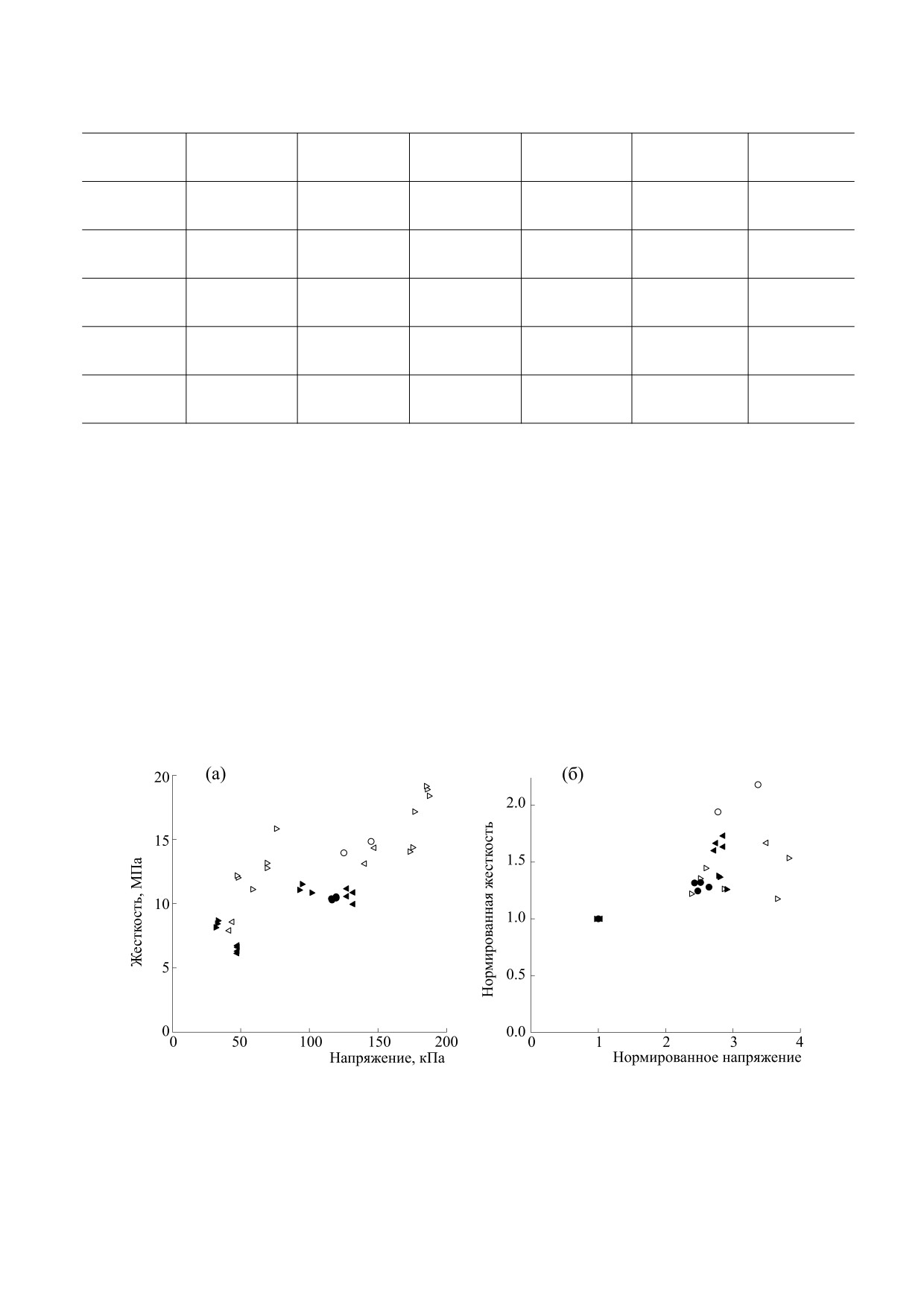
На рис. 3 показана зависимость абсолютных
пряжения и длины волокна. Интервал для
и нормированных значений жесткости от напря-
аппроксимации составлял два полных периода
жения.
колебаний длины с центром в выбранной точке
по времени.
Кинетику изменений напряжения и жесткости
характеризовали «полувременем», т.е. временем,
за которое изменение величины достигало поло-
РЕЗУЛЬТАТЫ
вины своего максимального значения. Темпера-
турные зависимости полувремени роста напря-
В наших экспериментах к волокнам были при-
жения и жесткости показаны в координатах Ар-
ложены скачки температуры от 15°С до 32°С, что
рениуса на рис. 4. Видно, что несмотря на
соответствовало финальным температурам во-
существенный разброс данных, напряжение име-
локна от 20°С до 37°С соответственно. При мень-
ет бόльшую температурную зависимость, чем
ших амплитудах скачка температуры не удавалось
жесткость. Угол наклона аппроксимирующей
обеспечивать подогрев волокна в течение такого
прямой для напряжения составлял -5.03 ± 0.95
времени, какое было необходимо для достижения
(n = 32, здесь и далее: взвешенное среднее ± взве-
медленными мышечными волокнами стационар-
шенное стандартное отклонение), соответствую-
ного напряжения.
щее значение для жесткости -2.86 ± 0.61 (n = 7).
БИОФИЗИКА том 66
№ 2
2021
ТЕМПЕРАТУРНАЯ ЗАВИСИМОСТЬ МЕХАНИЧЕСКИХ ХАРАКТЕРИСТИК
361
Экспериментальные данные
Средняя
Напряжение T,
t0.5T, мс
Жесткость S,
Snorm
t0.5S , мс
Tnorm
температура
кПа
MПа
5
44.1 ± 14.1
1
9.03 ± 2.65
1
1
(38)
(21)
21
120.0 ± 26.2
2.6 ± 0.5
47.0 ± 18.4
12.61 ± 2.61
1.51 ± 0.28
52.7 ± 15.2
(9)
(9)
(2)
(6)
(6)
(2)
26
132.6 ± 32.2
2.7 ± 0.5
24.5 ± 4.9
14.18 ± 3.66 (7)
1.54 ± 0.33 (7)
35.7 ± 9.5
(10)
(9)
(2)
(2)
31
137.9 ± 32.6
3.0 ± 0.5
14.5 ± 1.4
13.18 ± 2.97 (6)
1.47 ± 0.33
23.5 ± 1.4
(9)
(9)
(2)
(6)
(2)
37
121.0 ± 10.4
3.0 ± 0.3
10
10.10 ± 0.25
1.47 ± 0.22
18
(4)
(4)
(1)
(2)
(2)
(1)
Примечание. Значения, измеренные после скачка температуры, сгруппированы по финальной температуре. Отклонения тем-
пературы от среднего значения, указанного в таблице, не превышали 1°С. В каждой клетке показано среднее значение ± стан-
дартное отклонение. В скобках указано число измерений. Для сравнения относительных изменений напряжение и жесткость
нормировали на соответствующие значения до скачка температуры, при 5°С, нормированные значения обозначены Tnorm и
Snorm. t0.5T, t0.5S - полувремена ответов напряжения и жесткости.
Для оценки температурной зависимости жестко-
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
сти использовали два волокна (рис. 4б). Темпера-
Интересно сравнить полученные результаты с
турная зависимость напряжения для соответству-
соответствующими параметрами волокон быст-
ющей пары волокон (рис. 4а, точки) имела угол
рой (m. psoas) мышцы кролика [4, 20]. В экспери-
наклона -4.51 ± 0.41 (n = 14). Сгруппированные
ментах, описанных в работе [20], исследуемый
по величине финальной температуры значения
диапазон температур составлял 5-20°С и не до-
полувремени для напряжения и жесткости пока-
стигал значений, близких к физиологическим. В
заны в таблице. Можно заметить, что полувремя
работе [4] было показано, что рост изометриче-
для ответа жесткости всегда было больше, чем по-
ского напряжения происходит во всем диапазоне
лувремя для ответа напряжения.
температур 5-37°С. При температурах выше 30°С
Рис. 3. Изменение жесткости волокна в ответ на скачок температуры: (а) - абсолютные значения напряжения до
скачка температуры (5°С) и установившиеся после скачка температуры; (б) - те же значения, нормированные на
соответствующее напряжение до скачка температуры. Разные символы соответствуют разным волокнам (n = 6).
Светлые и темные символы соответствуют двум сериям экспериментов.
БИОФИЗИКА том 66
№ 2
2021
362
НАБИЕВ и др.
Рис. 4. Температурные зависимости изменений напряжения (а) и динамической жесткости (б) в ответ на скачок
температуры, выраженные графиками Аррениуса. По оси х отложены обратные температуры в градусах Кельвина,
умноженные на тысячу. По оси у - десятичный логарифм величины, обратной времени достижения напряжением
либо жесткостью половины разницы между стационарными значениями. Соответствующие
«полувремена»
обозначены t0.5T и t0.5S. Угловой коэффициент для напряжения составлял k = -5.0 (пунктирная линия, 6 волокон,
32 точки); для двух волокон, использовавшихся для определения температурной зависимости жесткости k = -4.5
(точечная линия, 2 волокна, 15 измерений). Для жесткости k = -2.9 (линия показана точками, 2 волокна, те же
символы, что и на рис. (а), 7 измерений.
рост замедлялся, однако напряжение по-прежне-
менения напряжения и жесткости в медленных
му увеличивалось с увеличением температуры [4].
m. soleus волокнах кролика в ответ на изменения
В наших экспериментах рост напряжения в диа-
температуры. Максимальные изменения были за-
пазоне температур 20-37°С был постоянным, но
регистрированы в интервале температур от 5°С до
незначительным. Основные изменения напряже-
20°С, дальнейший рост напряжения был незна-
ния в медленных волокнах происходили в интер-
чительным, жесткость в описываемых экспери-
вале 5-20°С. Рост напряжения при изменении
ментах также не зависела от температуры при
температуры от 5°С до 20°С составил 2.61 (см.
температурах, больших 20°С. Отметим, что в ра-
таблицу), а от 5°С до 30°С был трехкратным. Не-
боте [21] волокно активировали при высокой тем-
большое и статистически недостоверное падение
пературе, что ведет к неоднородной активации и
напряжения при возрастании температуры до
разбросу длин саркомеров в волокне, а частоты
37°С, по-видимому, связано с недостаточной
синусоидальных колебаний не превышали
мощностью подогревающего импульса. В экспе-
500 Гц, что не позволяет достоверно оценить ди-
риментах на волокнах из m. soleus человека рост
намическую жесткость, т.к. на этих временах
напряжения в диапазоне температур от 5°С до
успевает произойти изменение структурного и
20°С составлял 3.98 [16].
механического состояния миозиновых головок.
Наиболее яркие отличия между двумя типами
Значения изометрического напряжения и жест-
мышц проявляются в температурной зависимо-
кости в волокнах m. soleus кролика были измерены
сти жесткости. Если в волокнах быстрой мышцы
при температуре 12°С и различных уровнях каль-
кролика изменение жесткости с температурой не-
циевой активации [17]. Cреднее напряжение при
значительно [4, 20], то в волокнах медленной
максимальном pCa составляло 141 кПа, что выше
мышцы человека четырехкратный рост напряже-
измеренного нами при 20°С. Податливость полус-
ния при изменении температуры от 5°С до 20°С
аркомера составляла 39.5 ± 4.7 нм/МПа, что для
сопровождался двукратным ростом жесткости
средней длины саркомера в указанных экспери-
(2.18 по данным в работе [16]). В наших экспери-
ментах 2.4 мкм соответствует удельной жесткости
ментах жесткость при изменении температуры от
2400/2/39.5
= 30.4 МПа. Титрованием уровня
5°С до 20°С росла в среднем в полтора раза, при
кальциевой активации был получен широкий диа-
дальнейшем увеличении температуры до 30-37°С
пазон значений напряжения волокна и измерены
жесткость не изменялась (таблица).
соответствующие значения жесткости. Интересно,
Наши результаты согласуются и с данными ра-
что, как и в наших экспериментах, при напряже-
боты [21], авторы которой также наблюдали из- ниях ниже 120 кПа жесткость с ростом напряжения
БИОФИЗИКА том 66
№ 2
2021
ТЕМПЕРАТУРНАЯ ЗАВИСИМОСТЬ МЕХАНИЧЕСКИХ ХАРАКТЕРИСТИК
363
существенно увеличивалась, но ее рост замедлялся
ла в 33 раза меньше, чем в быстрых; при 10°С раз-
до едва заметного при превышении этого уровня
личие становилось 84-кратным [25]. Снижение
напряжения.
напряжения в этих экспериментах было связано с
ассоциацией фосфата с комплексом миозин-
Известно, что в волокнах быстрой мышцы не-
АДФ, образовавшимся в результате сброса фос-
большой рост жесткости после скачка температу-
фата из активного центра миозиновой головки
ры опережает рост напряжения (см, например,
после гидролиза АТФ, и обращением ее силоге-
рис. 8 в работе [4]). Мы характеризовали кинети-
нерирующего шага. Эксперименты, в которых
ку нарастания напряжения и жесткости в ответ на
механические и структурные изменения в мед-
скачок температуры величиной полувремени из-
ленных и быстрых волокнах мышцы крысы были
менения соответствующей величины (таблица,
инициированы импульсным фотоиндуцирован-
рис. 4). В отличие от волокон быстрых мышц в
ным выделением АТФ в присутствии и в отсут-
наших экспериментах рост напряжения не отста-
ствие ионов Са2+, показали [26], что головки мед-
вал от роста жесткости и даже опережал его.
ленного миозина связывают АДФ намного проч-
Различия в кинетике механических процессов
нее и развивают активное напряжение примерно
для медленного и быстрого миозина были описа-
в пять раз медленнее, чем головки быстрого мио-
ны ранее. Максимальная скорость укорочения
зина. Простая кинетическая модель рабочего
медленных волокон была примерно вдвое мень-
цикла миозиновой головки в медленной мышце
ше, чем быстрых для мышц морской свинки [8],
была предложена в работе [16]. Тем не менее су-
крысы [13], трески [12] и человека [22] в широком
ществующих экспериментальных данных еще не-
диапазоне температур вплоть до физиологиче-
достаточно для определения всех параметров по-
ской для животных данного вида. Еще большее,
добных моделей.
более чем пятикратное, различие скорости дви-
жения актиновых филаментов по поверхности,
В нашей работе мы показали, что отличия в
покрытой медленным и быстрым миозином из
механических характеристиках медленных и
мышц крыс и кроликов, было обнаружено в экс-
быстрых волокон проявляются в широком диапа-
периментах в искусственной подвижной системе
зоне температур вплоть до физиологической. В
(in vitro motility assay) [23]. Аналогичные результа-
ответ на скачок температуры в волокнах быстрых
ты были получены в экспериментах, в которых
мышц происходит быстрый переход слабосвязан-
изучали движение не актиновых филаментов, а
ных миозиновых головок в силогенерирующее
реконструированных тонких нитей, причем для
состояние без существенного увеличения числа
реконструкции использовали тропонин и тропо-
присоединенных к актину головок [7]. Поскольку
миозин из медленных и быстрых мышц соответ-
в медленных волокнах при увеличении темпера-
ственно [24]. В этой же работе в опытах с оптиче-
туры не только напряжение, но и жесткость изме-
ской ловушкой также было обнаружено трехкрат-
няется существенно, мы полагаем, что в этих во-
ное увеличение среднего времени пребывания
локнах скачок температуры вызывает два процес-
одиночных головок медленного миозина в при-
са: увеличение доли силогенерирующих головок
соединенном к актину состоянии по сравнению с
за счет слабосвязанных и увеличение общего чис-
быстрым миозином [24]. Эти данные хорошо со-
ла присоединенных головок.
гласуются с результатами измерения АТФазной
активности медленных и быстрых волокон чело-
века, сокращающихся в изометрических и изото-
БЛАГОДАPНОCТИ
нических условиях при 10°С и 20°С [22], которые
Авторы благодарны Г.В. Копыловой и
показали, что доля времени, которое головка
миозина медленных мышц проводит в прочно
А.М. Кочуровой за помощь в анализе изоформ-
ного состава миозина в экспериментальных об-
связанном с актином состоянии, в несколько раз
разцах.
больше, чем для миозиновых головок в быстрых
волокнах, а также с оценкой доли присоединен-
ных к актину головок в активном изометриче-
ском сокращении медленных и быстрых волок-
ФИНАНСИРОВАНИЕ РАБОТЫ
нах кролика при 12°С - 0.47 и 0.29 coответственно
Работа выполнена при поддержке РФФИ,
[17].
грант № 18-04-00599, и госпрограмм AAAA-A18-
Различия кинетики биохимических процес-
118020590135-3 и АААА-А19-119012990119-3.
сов, лежащих в основе механической активности
медленных и быстрых мышц, еще более значи-
тельны. Скорость снижения механического на-
КОНФЛИКТ ИНТЕРЕСОВ
пряжения в ответ на фотоиндуцированное увели-
чение концентрации неорганического фосфата в
Авторы заявляют об отсутствии конфликта
медленных волокнах мышц кролика при 20°С бы-
интересов.
БИОФИЗИКА том 66
№ 2
2021
364
НАБИЕВ и др.
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
12. J. D. Altringham and I. A. Johnston, J. Physiol. 333,
421 (1982).
Все применимые международные, националь-
13. K. W. Ranatunga, J. Physiol. 351, 517 (1984).
ные и институциональные принципы ухода и ис-
14. P. J. Reiser, R. L. Moss, G. G. Giulian, et al., J. Biol.
пользования животных при выполнении работы
Chem. 260, 9077 (1985).
были соблюдены.
15. M. Linari, R. Bottinelli, M. A. Pellegrino, et al., J.
Physiol. 554, 335 (2004).
СПИСОК ЛИТЕРАТУРЫ
16. B. Brenner, N. Hahn, E. Hanke, et al., J. Muscle. Res.
Cell Motil. 33 (6), 403 (2012).
1. A. F. Huxley and R. M. Simmons, Nature 233, 533
17. V. Percario, S. Boncompagni, F. Protasi, et al., J.
(1971).
Physiol. 596 (7), 1243 (2018).
2. K. C. Holmes, Curr Biol. 7 (2), R112 (1997).
18. A. K. Tsaturyan, S. Y. Bershitsky, N. A. Koubassova,
3. S. Y. Bershitsky and A. K. Tsaturyan, Biophys. J. 56 (5),
et al., Biophys. J. 101 (2), 404 (2011).
809 (1989).
19. П. В. Кочубей и С. Ю. Бершицкий, Биофизика 59
4. S.Y. Bershitsky, A.K. Tsaturyan, J. Physiol. 540 (1), 971
(5), 967 (2014).
(2002).
20. М. Linari, M. Caremani, C. Piperio, et al., Biophys. J.
5. A. K. Tsaturyan, S. Y. Bershitsky, R. Burns, et al., Bio-
92 (7), 2476 (2007).
phys. J. 77 (1), 354 (1999).
21. G. Wang and M. Kawai, J. Physiol. 531 (1), 219 (2001).
6. G. Piazzesi, M. Reconditi, N. Koubassova, et al., J.
22. Z. H. He, R. Bottinelli, M. A. Pellegrino, et al., Bio-
Physiol. 549 (1), 93 (2003).
phys J. 79 (2), 945 (2000)
7. M. A. Ferenczi, S. Y. Bershitsky, N. Koubassova, et al.,
23. R. Rossi, M. Maffei, R. Bottinelli, et al., J. Appl. Physi-
Structure 13 (1), 131 (2005).
ol. 99 (6), 2239 (2005).
8. J. Gulati, Proc. Natl. Acad. Sci. USA 73 (12), 4693
24. Д. В. Щепкин, С. Р. Набиев, Н. А. Кубасова и др.,
(1976).
Бюлл. эксперим. биологии и медицины 169 (3), 310
9. D. J. Stephenson and D. A. Williams, J. Physiol. 317,
(2020).
281 (1981).
25. N. C. Millar and E. Homsher, Am. J. Physiol. 262 (5),
10. D. J. Stephenson and D. A. Williams, J. Physiol. 360, 1
C1239 (1992).
(1985).
26. K. Horiuti, N. Yagi, and S. Takemori, J. Physiol. 502
11. K. W. Ranatunga, Exp. Neurol. 70 (2), 211 (1980).
(2), 433 (1997).
Temperature Dependence of Mechanical Characteristics of Demembranized Rabbit
Slow Muscle Fibres
S.R. Nabiev*, S.Y. Bershitsky*, A.K. Tsaturyan**, and N.A. Koubassova**
*Institute of Immunology and Physiology, Ural Branch of the Russian Academy of Sciences,
Pervomayskaya ul. 106, Yekaterinburg, 620049 Russia
**Institute of Mechanics, Lomonosov Moscow State University, Mitchurinsky prosp. 1, Moscow, 119192 Russia
We have studied the temperature dependences of the tension and stiffness of actively contracting single fibers
from rabbit slow (soleus) muscle using joule temperature jump. We have shown differences in the character-
istics between fibers from rabbit fast (psoas) and slow (soleus) muscles, assuming that these differences are
due to differences in the kinetics of the myosin head cycle.
Keywords: muscle contraction, isometric tension, temperature jump
БИОФИЗИКА том 66
№ 2
2021