БИОФИЗИКА, 2021, том 66, № 2, с. 372-383
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 534.75
ПОДСТРОЙКА ЭХОИМПУЛЬСОВ, ОТРАЖЕННЫХ
ОТ РАСПОЛОЖЕННЫХ НА РАЗНЫХ ДИСТАНЦИЯХ ОБЪЕКТОВ,
ПОД СЛУХОВУЮ ЧУВСТВИТЕЛЬНОСТЬ У ДЕЛЬФИНОВ:
МОДЕЛЬНЫЕ ИССЛЕДОВАНИЯ
© 2021 г. Л.К. Римская-Корсакова*, А.А. Родионов**, М.П. Иванов***
*АО «Акустический институт имени академика Н.Н. Андреева», 117036, Москва, ул. Шверника, 4
E-mail: lkrk@mail.ru
**Институт океанологии РАН (Санкт-Петербургский филиал),
199034, Санкт-Петербург, Университетская наб., 5
E-mail: sppp@spbrc.nw.ru
***Санкт-Петербургский государственный университет,
199034, Санкт-Петербург, Университетская наб., 7/9
E-mail: 20mivanov@mail.ru
Поступила в редакцию 03.05.2020 г.
После доработки 14.07.2020 г.
Принята к публикации 23.12.2020 г.
Дельфины при эхолокации во время охоты излучают зондирующий импульс и принимают
эхоимпульс, на основании которого обнаруживают и идентифицируют жертву в фазе поиска. В фазе
захвата дельфины нуждаются в стабильной слуховой реакции на эхоимпульс. Для объяснения
особенностей восприятия эхоимпульсов в фазах поиска и захвата моделировали вызванную
импульсами реакцию ансамбля высокочастотных периферических волокон. Пара импульсов с
центральными частотами в
70 кГц, разными межимпульсными интервалами и пиковыми
амплитудами была моделью зондирующего импульса и эхоимпульса. Импульсы поступали на вход
модели, имеющей разные этапы обработки, включая преобразование рецепторного потенциала
волосковых клеток в последовательность потенциалов действия ансамбля волокон слухового нерва.
Показано, что для воспроизведения временной структуры импульсов в реакции ансамбля
необходимо соответствие возникшего рецепторного потенциала порогам реакций большинства
волокон ансамбля. При коротких интервалах между импульсами в паре воспроизведению
временной структуры второго импульса (эхоимпульса) способствовало повышение порогов
реакций волокон ансамбля за счет рефрактерных свойств, вызванных действием первого импульса
(модели зондирующего импульса). При больших интервалах между импульсами в паре за
воспроизведение структуры второго импульса (эхоимпульса) могли отвечать вариации пиковых
значений амплитуд первого импульса (модели зондирующего импульса). Показано, что метод
моделирования - удобный способ для выявления различий способов согласования эхоимпульсов
со слуховой чувствительностью дельфинов в фазах поиска и захвата жертвы.
Ключевые слова: эхолокация, периферическое кодирование, волокна слухового нерва, воспроизведение
временной структуры эхоимпульсов.
DOI: 10.31857/S0006302921020198
дБ отн. 1 мкПа). Анализируя эхоимпульсы, дель-
Дельфины используют эхолокацию для нави-
гации и во время охоты [1, 2]. Они излучают зон-
фины определяют направление на источник с
дирующие импульсы и принимают отраженные
точностью ±1°, обнаруживают и распознают объ-
от объектов эхоимпульсы. Зондирующие импуль-
екты, незначительно отличающиеся по размеру,
сы имеют малую длительность (<100 мкс), высо-
форме и материалу. Эхолокатор дельфина имеет
кую частоту (50-130 кГц) и интенсивность (~ 230
преимущества перед средствами гидроакустики
при поиске малоразмерных объектов, располо-
Сокращения: ЗИ - зондирующий импульс, ЭИ - эхоим- женных в толще воды, на дне, под слоем ила или
пульс, СВП - слуховые вызванные потенциалы, ВВК -
подо льдом, т.е. при наличии шумов, донной и
внутренние волосковые клетки, ВСН - волокна слухового
нерва.
объемной реверберации, рассеяния звуков на
372
ПОДСТРОЙКА ЭХОИМПУЛЬСОВ
373
границах сред. Поэтому изучение эхолокацион-
ния высокочастотных звуков в интересах объяс-
ных механизмов у дельфинов, в том числе метода-
нения особенностей слухового приема дельфина-
ми моделирования, имеет фундаментальное и
ми эхоимпульсов, отраженных от объектов, рас-
практическое значение.
положенных на коротких и дальних дистанциях.
Также ставились задачи определения целесооб-
Дельфины кормятся во время охоты, в которой
разности варьирования амплитуд ЗИ в пачках
выделяют фазы поиска и захвата жертвы [3]. Слу-
при поиске жертвы на дальних дистанциях и
ховой анализ эхоимпульсов в фазе поиска позво-
определения причин, по которым вызванная эхо-
ляет дельфину обнаружить и идентифицировать
импульсом реакция ансамбля могла быть ста-
жертву, а в фазе захвата - иметь стабильную от
бильной в фазе захвата жертвы.
нее реакцию.
Стабилизацию слуховых реакций на эхоим-
Считается, что на периферии слуховой систе-
пульсы наблюдали при лоцировании дельфинами
мы млекопитающих тонкая временная структура
алюминиевого цилиндра на дистанциях в 1-8 м,
сигналов с частотами выше 4 кГц не воспроизво-
т. е. при интервалах между зондирующими им-
дится, поскольку импульсные отклики базиляр-
пульсами (ЗИ) и эхоимпульсами (ЭИ) в 1-10 мс
ной мембраны сглаживают реакции рецепторных
[4-6]. Зарегистрированные на этих дистанциях
внутренних волосковых клеток (ВВК), обладаю-
слуховые вызванные потенциалы (СВП) ствола
щих интегрирующими свойствами [12]. Постоян-
мозга дельфинов в ответ на ЭИ были постоянны-
ную времени интегрирования оценивают при-
ми. Постоянству амплитуд СВП при уменьшении
мерно в 0.2 мс. Сглаживание компенсируют реак-
дистанции, как полагали авторы, способствовало
ции наборов волокон слухового нерва (ВСН),
снижение слуховой чувствительности к ЭИ за
связанных с одной ВВК, а также генерация по-
счет повышения эффекта маскировки, вызван-
тенциалов действия (спайков) ВСН, приурочен-
ной ЗИ, но рост пиковых амплитуд ЭИ за счет
ных к возрастающей части рецепторного потен-
уменьшения затухания звука в воде. Временная
циала ВВК [13, 14]. Ограничения числа ВСН,
маскировка способствовала подстройке чувстви-
имеющих свойства восстановления возбудимо-
тельности слуха дельфина под интенсивность от-
сти (рефрактерностью и адаптацией), могут при-
раженного от объекта ЭИ, и поэтому получению
вести к появлению особенностей перифериче-
стабильной информации о жертве в фазе захвата
ского кодирования временной структуры высо-
на дистанциях в 1-8 м.
кочастотных импульсов, рассмотрению которых
посвящена данная работа. Под периферическим
На средних дистанциях до объектов, составля-
кодированием импульсов мы понимали преобра-
ющих примерно 20-60 м, дельфины излучают
зование аналогового рецепторного потенциала
каждый следующий ЗИ после приема предыду-
ВВК в последовательность спайков ансамбля
щего эхоимпульса. При этом дельфины варьиру-
ВСН.
ют в широком диапазоне интенсивность зондиру-
ющих импульсов при обнаружении или распо-
Следует отметить, что возникшая в ответ на
знавании объектов как в экспериментальном
высокочастотный импульс реакция ансамбля
бассейне [1, 7], так и при свободном плавании [8,
ВСН вызывает последовательные реакции ней-
9]. На целесообразность вариаций интенсивно-
ронных структур прямого слухового пути, кото-
сти ЗИ как способа подстройки эхоимпульсов
рые ответственны за формирование СВП и слухо-
под слуховую чувствительность дельфина в точке
вые реакции дельфинов в целом. Учитывая по-
приема указывали данные моделирования
требность дельфинов в быстром реагировании на
свойств периферического кодирования импульс-
принятые эхоимпульсы, учитывая короткий путь
ных звуков [10].
формирования слуховых реакций, можно ожи-
На больших дистанциях до объектов, превы-
дать проявлений свойств периферического коди-
шающих 100 м, дельфины излучали пачки ЗИ с
рования в характеристиках СВП и слуховых реак-
периодами следования в 140-160 мс [11]. Внутри
циях дельфинов.
пачки имели место вариации интенсивностей ЗИ
и задержек между ЗИ от 15 до 20 мс. При таких за-
держках временная маскировка не должна влиять
МОДЕЛИРОВАНИЕ РЕАКЦИЙ АНСАМБЛЯ
на слуховое восприятие отдельных эхоимпульсов
ВОЛОКОН СЛУХОВОГО НЕРВА
в пачке [6]. Поэтому можно предположить, что
В работе была использована модель волокна
вариации интенсивности ЗИ в пачке могли под-
слухового нерва [10, 13-15], включающая не-
страивать интенсивность эхоимпульсов под слу-
сколько этапов преобразований входного сигнала
ховую чувствительность дельфина в точке приема
X(t). К ним относятся:
и тем самым способствовать идентификации
жертвы (объекта) на этапе его обнаружения.
1. Полосовая фильтрация сигнала X(t), воспро-
Целью данной работы было моделирование
изводящая фильтрующие свойства базилярной
свойств временного периферического кодирова-
мембраны (выходная функция Y(t)).
БИОФИЗИКА том 66
№ 2
2021
374
РИМСКАЯ-КОРСАКОВА и др.
2. Детектирование и интегрирование, участву-
течение 50 мкс и были близки профилям и дли-
ющие в формировании рецепторных потенциа-
тельностям зондирующих импульсов дельфинов-
лов моделей ВВК (выходная функция R(t)).
афалин [1]. Центральная частота импульсов была
равна 70 кГц. Эхоимпульсы, отраженные от ре-
3. Формирование синаптического потенциала
альных жертв дельфинов, а также от мишеней,
модели волокна слухового нерва (выходная функ-
ция G(t)) в виде пуассоновского случайного про-
используемых в экспериментах, чаще всего со-
стоят из наборов отражений зондирующих им-
цесса со средней и дисперсией, пропорциональ-
пульсов, интервалы между которыми могут быть
ные функции R(t); для воспроизведения спонтан-
менее ста микросекунд. Поэтому моделями эхо-
ной активности дисперсию функции G(t)
импульсов были пары и тройки коротких импуль-
увеличивали на некоторую величину.
сов с межимпульсными интервалами меньшими
4. Генерация потенциалов действия (спайков
100 мкс.
Pi) при условии, что функция G(t) превышает по-
Характеристическая частота моделей ВСН, со-
роговую функцию H(t);
ответствующая частоте настройки фильтра бази-
5. Изменение пороговой функции H(t) модели
лярной мембраны, была равна 70 кГц. Шаг вы-
волокна после генерации спайка, включающее
числения составлял 1 мкс. В ответ на однократное
мгновенное повышение порога и его постепен-
предъявление одиночных, пар или троек импуль-
ное восстановление за 20-30 мс до начального
сов получали реакцию ансамбля из 300 моделей
значения, а также повышение текущего значения
волокон в виде гистограммы Pi(t). Пиковые ам-
порога на остаточное значение порога, бывшее в
плитуды импульсов выражали в дБ относительно
момент генерации нового спайка после генера-
амплитуды одиночного импульса на пороге реак-
ции предыдущего спайка. Такой механизм повы-
ции ансамбля.
шения порога позволял достоверно воспроизво-
дить в моделях ВСН свойства рефрактерности и
адаптации реальных ВСН [16].
РЕЗУЛЬТАТЫ
Параметры моделей ВСН были подобраны
Восстановление временной структуры импуль-
так, чтобы воспроизвести известные свойства ре-
сов в реакции ансамбля моделей волокон. На
акций реальных ВСН наземных млекопитающих.
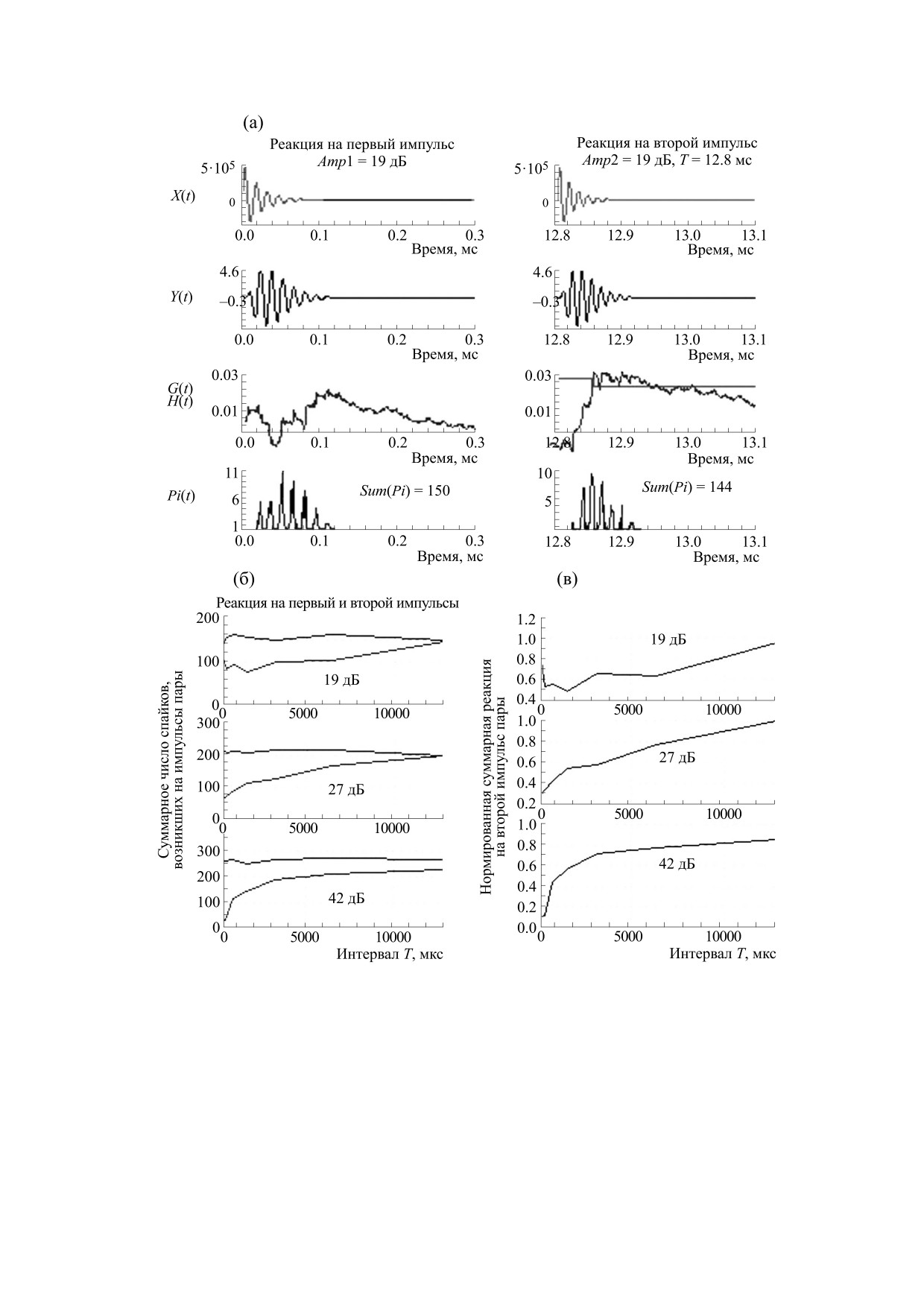
рис. 1а представлены реакции разных блоков мо-
Дельфины не являются обычными лабораторны-
дели на пару надпороговых импульсов с амплиту-
ми животными, и прямые исследования реакций
дами в 19 дБ и межимпульсными интервалами Т в
ВСН у дельфинов невозможны. Поэтому адекват-
12.8 мс. Показаны профили входных сигналов
ность использованной модели подтверждали пу-
X(t) и импульсных откликов базилярной мембра-
тем воспроизведения свойств реальных СВП
ны Y(t), сглаженные рецепторные потенциалы
дельфинов, особенностей слухового восприятия
ВВК G(t), а также суммарные реакции ансамбля
импульсных звуков у дельфинов [17-19]. Модель-
волокон Pi(t). Данные рис. 1а показали, что про-
ные СВП получали путем свертки реакции ан-
филь функции G(t), вызванный первым импуль-
самбля ВСН с «элементарным» СВП, в основе ко-
сом, не соответствует профилю Y(t). За счет инте-
торого был реальный СВП дельфина, получен-
грирующих свойств ВВК потенциал G(t) сначала
ный на одиночный щелчок.
возрастал, а потом плавно спадал до нуля. Макси-
Следует отметить, что на этапе моделирования
мум потенциала G(t) приходился на окончание
импульсных откликов базилярной мембраны мы
функции Y(t). Интегрирующие свойства ВВК
не учитывали нелинейные свойства смещений
способствовали повышению отношения сиг-
базилярной мембраны [20]. Динамический диа-
нал/шум, но нарушали воспроизведение времен-
пазон используемой модели периферии был су-
ной структуры импульса. Структуру восстанавли-
щественно уже реального динамического диапа-
вала суммарная реакция ансамбля ВСН.
зона слуховой системы дельфина, достигающего
При низких пиковых амплитудах импульсов
почти 200 дБ отн 1 мкПа. Поэтому моделирова-
(19 дБ) каждое волокно ансамбля (из-за рефрак-
ние было направлено на выявление качественных
терности) могло генерировать не более одного
закономерностей кодирования тонкой времен-
спайка в момент, когда положительный градиент
ной структуры эхоимпульсов, у которых пиковые
потенциала G(t) превышал пороговую функцию
значения амплитуд менялись в динамическом
H(t). За счет внутреннего шума (спонтанной ак-
диапазоне модели периферии. Нас интересовали
тивности) профили функций G(t) в разных волок-
свойства преобразования профиля входного сиг-
нах ансамбля отличались случайным образом,
нала (X(t)) в последовательность потенциалов
поэтому возникшие в разных волокнах спайки
действия (спайков), которую генерировал ан-
были не синхронизированы. Появление спайков
самбль ВСН (Pi(t)).
было приурочено к разным частям возрастающей
Входными сигналами модели X(t) были оди-
части потенциала G(t), и поэтому суммарная ре-
ночные импульсы, а также пары или тройки им-
акция ансамбля Pi(t) восстанавливала профиль
пульсов, временные профили которых спадали в
функции Y(t) (рис. 1а). Данный способ кодирова-
БИОФИЗИКА том 66
№ 2
2021
ПОДСТРОЙКА ЭХОИМПУЛЬСОВ
375
Рис. 1. Реакции разных блоков модели на пару равновеликих импульсов с пиковыми амплитудами в 19 дБ и задержкой
Т в 12.8 мс (а); зависимости от задержки Т суммарного числа спайков, вызванных импульсами пары (б) и
нормированной суммарной реакции на второй импульс пары (в). Параметры на (б) и (в) - значения пиковых
амплитуд импульсов в дБ относительно амплитуды импульса на пороге. По осям абсцисс: на (а) - время в мс, на (б) и
(в) - величина интервала Т в мс; по осям ординат: на (а) - амплитуды в усл. ед.; на (б) - число спайков на каждом шаге;
на (в) - отношение суммарной реакции на второй импульс. Sum(Pi) - суммарное число спайков, возникшее в
ансамбле в ответ на эхосигнал. X(t), Y(t), G(t), H(t), Pi(t) и Sum(Pi) - см. объяснения в тексте.
БИОФИЗИКА том 66
№ 2
2021
376
РИМСКАЯ-КОРСАКОВА и др.
ния пар низко амплитудных импульсов в ансам-
блем волокон слухового нерва, позволяет воспро-
бле ВСН был назван стохастическим [21].
извести временную структуру низкоамплитудных
импульсов, входящих в пару, а также структуру
На рис. 1б показаны зависимости суммарного
пар импульсов. Воспроизведение структуры тем
числа спайков, возникших на импульсы пары, от
точнее, чем точнее соответствие потенциала G(t)
интервала между ними, а на рис. 1в - кривые вос-
порогу реакции волокон в ансамбле. Способ ко-
становления реакции на второй импульс как за-
дирования пар высокоамплитудных импульсов -
висимости нормированной амплитуды реакции
детерминистический. Восстановление реакции
на второй импульс от интервала между импульса-
на второй импульс определяется свойствами ре-
ми пары. Зависимости были получены при трех
фрактерности и адаптации волокон ансамбля.
амплитудах импульсов (19, 27 и 47 дБ).
Воспроизведение структуры пары импульсов за-
При амплитудах импульсов в 19 и 27 дБ и ме-
висело от интервала между ними.
жимпульсном интервале в 0.8 мс суммарное чис-
Моделирование эффекта стабилизации слухо-
ло спайков, возникших на первый импульс в па-
вых реакций. Учитывая рассмотренные выше
ре, соответствовало числу среагировавших воло-
свойства, можно объяснить эффект стабилиза-
кон и было больше числа спайков (волокон),
ции реакции на эхоимпульс, наблюдаемый у
возникших на второй импульс (рис. 1б). Умень-
дельфинов при эхолокации объектов на коротких
шение суммарной реакции Sum(Pi) на второй им-
дистанциях [6]. В модельном эксперименте вход-
пульс обусловлено конечным числом волокон в
ным сигналом X(t) была пара разновеликих им-
ансамбле, рефрактерными свойствами и низкими
пульсов (рис. 2). Первый импульс пары был моде-
значениями потенциалов G(t). При низких ам-
лью зондирующего импульса (ЗИ), а второй - мо-
плитудах на второй импульс могли ответить толь-
делью эхоимпульса (ЭИ). Чтобы учесть
ко те волокна, которые еще не ответили на пер-
уменьшение амплитуды ЭИ при увеличении за-
вый импульс. При интервале в 13 мс реакция на
держки между ЗИ и ЭИ за счет свойств распро-
второй импульс восстановилась полностью
странения звука в воде, амплитуду модели ЭИ
(рис. 1в): суммарные реакции Sum(Pi), а также
уменьшали согласно выражению Amp2
=
профили реакций на первый и второй импульс
= Amp1/(2TV), где Amp1 и Amp2 - амплитуды мо-
были одинаковыми (рис. 1а). Реакция на второй
делей ЗИ и ЭИ; T - задержка ЭИ относительно
импульс при увеличении интервала повышалась,
ЗИ; V - скорость звука в воде. Поэтому при пико-
так как с течением времени увеличивалось число
вой амплитуде модели ЗИ в 42 дБ при задержке
волокон, готовых ответить в случайный момент
между ЭИ и ЗИ в 39 мс амплитуда ЭИ составляла
времени. Чем меньше были амплитуды импуль-
6 дБ (рис. 2а), а вызванные ЗИ и ЭИ реакции
сов, тем меньше волокон отвечало на первый им-
Sum(Pi)
- 286 и 20 спайков соответственно
пульс, тем больше волокон отвечало на второй,
(рис. 2а).
тем меньше реакция на второй импульс зависела
от межимпульсного интервала.
При приближении к жертве дельфин генери-
ровал стабильные по амплитуде ЗИ и принимал
При наибольшей амплитуде импульсов в 42 дБ
ЭИ, амплитуды которых возрастали при умень-
практически все волокна в ансамбле ответили на
шении задержки. При высокой амплитуде ЗИ ан-
первый импульс (т.е. Sum(Pi) ~ 280) (рис. 1б). По-
самбль волокон кодировал пару, включающую
этому при интервале в 0.8 мс число спайков, вы-
ЗИ и ЭИ, детерминистическим способом, а реак-
званных вторым импульсом, было почти равно
ция на ЭИ была под влиянием свойств рефрак-
нулю. Восстановление реакции на второй им-
терности и адаптации волокон ансамбля. За счет
пульс отличалось от такового при амплитудах им-
двух взаимоисключающих процессов - повыше-
пульсов в 19 и 27 дБ (рис. 1в). При амплитуде в
ния амплитуды ЭИ вследствие распространения
42 дБ восстановление было более быстрым при
звука в воде и уменьшения реакции ансамбля,
задержках в 1-7 мс. Однако при задержке в 13 мс
вызванной ЭИ, вследствие рефрактерных
суммарная реакция на второй импульс была
свойств волокон - реакция на ЭИ была постоян-
меньше таковой на первый импульс. При высо-
ной при Т < ~10-13 мс (рис. 2г).
ких амплитудах на второй импульс отвечали те
волокна, которые уже ответили на первый им-
Таким образом, моделирование объяснило на-
пульс. Поэтому восстановление реакции на вто-
блюдаемую при коротких задержках Т стабилиза-
рой импульс соответствовало изменению порогов
цию слуховых реакции на эхоимпульсы у дельфи-
волокон вследствие рефрактерности и адаптации.
нов при их приближении к объекту [4-6] или
Такой способ кодирования пары импульсов был
жертве в фазе захвата.
назван детерминистическим [19].
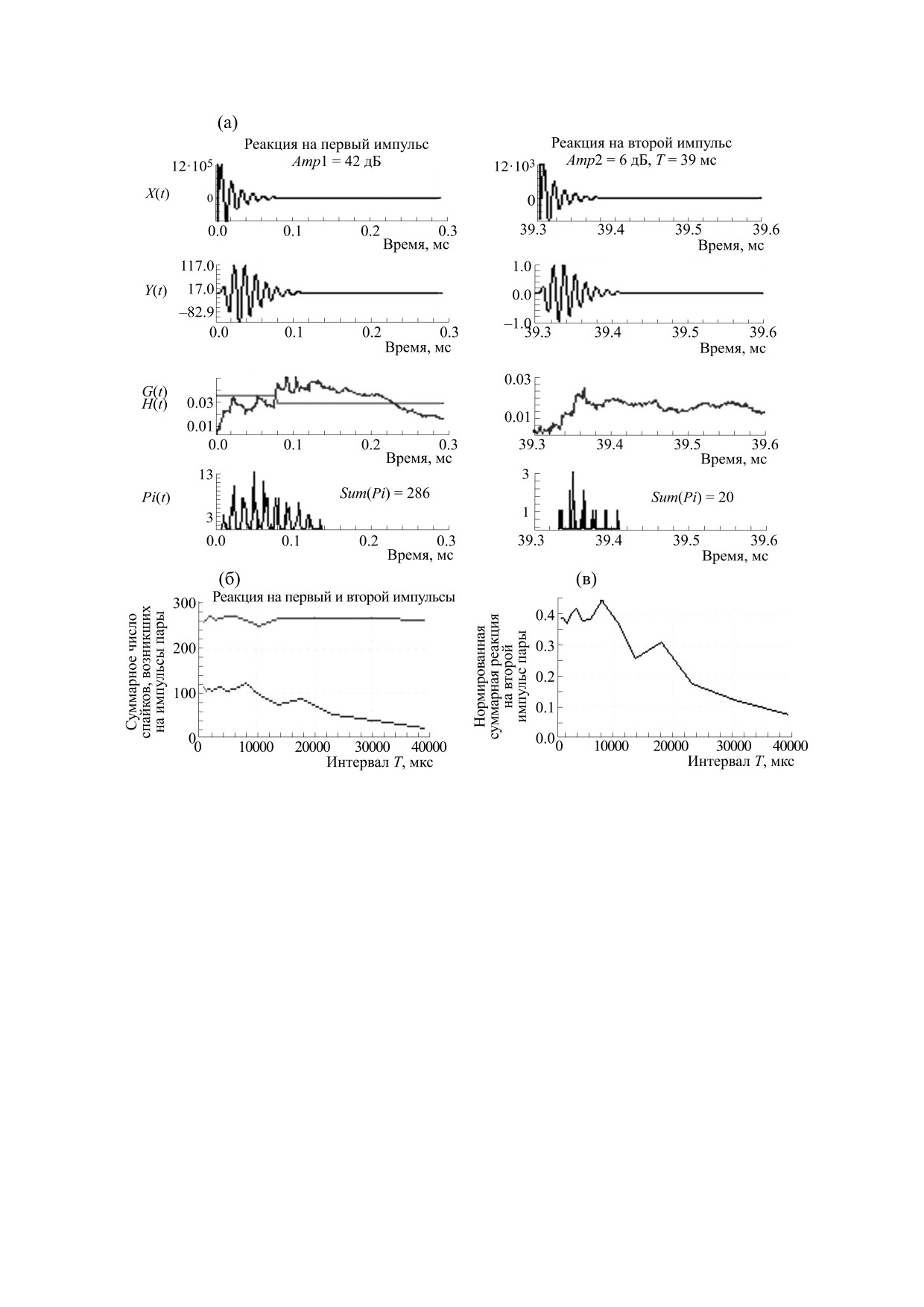
Роль варьирования амплитуд зондирующих им-
Таким образом, воспроизведение временной
пульсов в пачках при поиске жертвы на дальних ди-
структуры пары импульсов зависело от амплитуд
станциях. Оценим значение варьирования ам-
импульсов и интервалом между ними. Стохасти-
плитуд зондирующих импульсов в пачке, излуча-
ческий способ кодирования, реализуемый ансам-
емой дельфинами в фазе поиска жертвы на
БИОФИЗИКА том 66
№ 2
2021
ПОДСТРОЙКА ЭХОИМПУЛЬСОВ
377
Рис. 2. Реакции блоков модели на пару разновеликих импульсов, имеющих пиковую амплитуду первого и второго
импульсов в 42 и 6 дБ соответственно и с межимпульсным интервалом Т в 39 мс (а). Зависимости суммарного числа
спайков, возникших на импульсы пары (б) и нормированной суммарной реакции на второй импульс (в) от величины
интервала Т. Остальные обозначения, как на рис. 1.
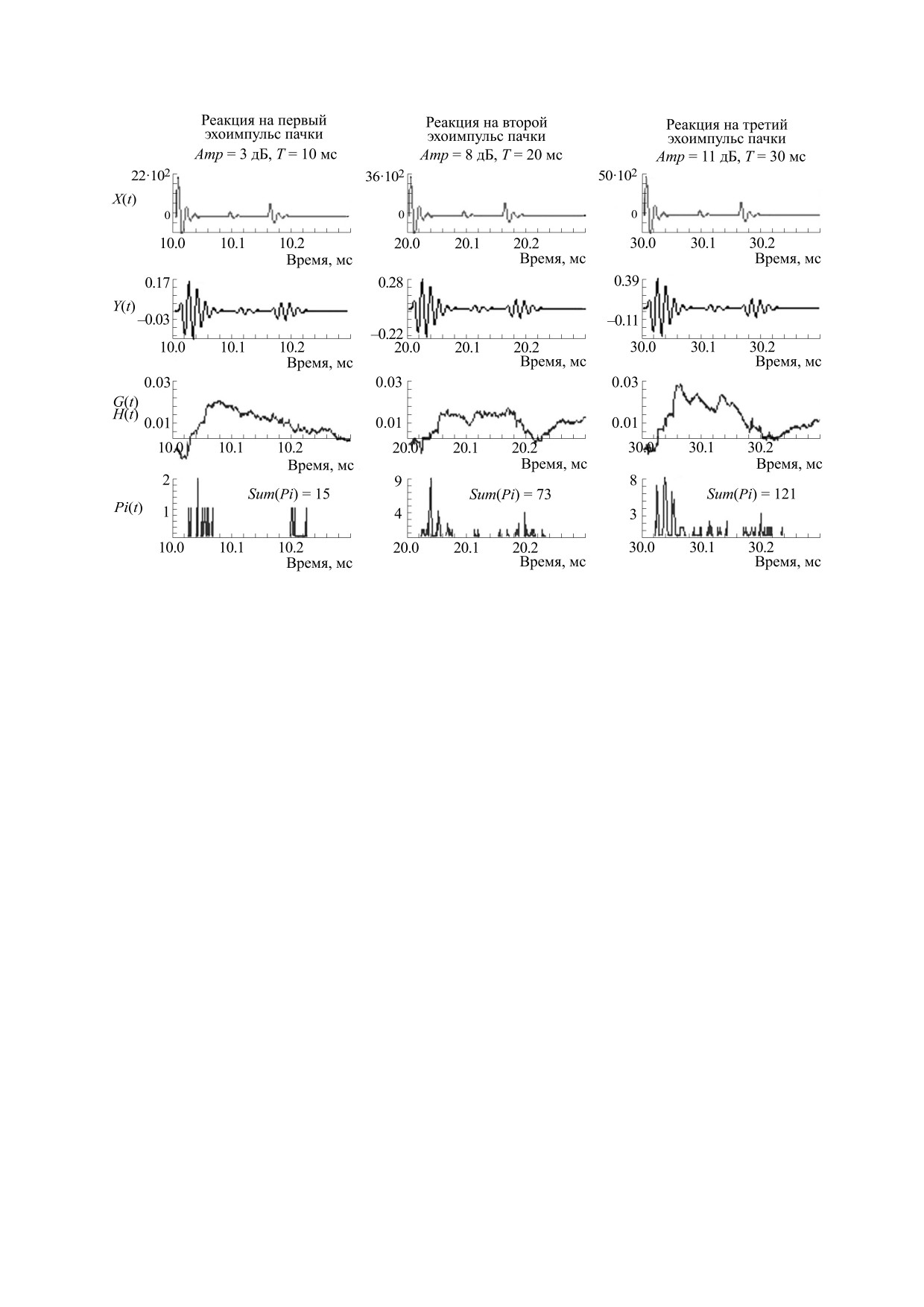
дальних дистанциях [11]. После излучения пачки
ситуация имеет место при приеме ЭИ, пришед-
ЗИ дельфин принимает пачку эхоимпульсов.
ших с дальних дистанций.
Временная структура ЭИ формируется при отра-
Значения пиковых амплитуд компонентов
жении ЗИ от жертвы и может включать в себя не-
ЭИ превышали пороги реакции волокон ансам-
сколько импульсных компонентов. В нашем слу-
бля не более чем на 15 дБ. При задержках между
чае моделью пачки ЭИ (рис. 3) были три импульс-
ЭИ в 10 мс реакция, вызванная предыдущим ЭИ,
ных комплекса, каждый из которых включал в
уже не влияла на реакцию, вызванную последую-
себя три импульсных компонента с одинаковыми
щим ЭИ (рис. 1г). Увеличение амплитуды Amp
частотами в 70 кГц, амплитудами, равными Amp,
приводило к росту суммарного числа спайков,
Amp/10, Amp/3, и интервалами между компонен-
вызванных ЭИ (рис. 3): при Amp в 3 или 8 или
11 дБ значения Sum(Pi) составили 15 или 73 или
тами в 90 и 70 мкс (функция X(t)). Интервалы
121 спайков.
между ЭИ были равны 10 мс. Амплитуду Amp ва-
рьировали вблизи порога реакции большинства
Ансамбль волокон кодировал каждый из трех
моделей волокон в ансамбле, полагая, что такая низкоамплитудных комплексов (модель ЭИ) сто-
БИОФИЗИКА том 66
№ 2
2021
378
РИМСКАЯ-КОРСАКОВА и др.
Рис. 3. Реакции разных блоков модели на пачку из трех импульсных комплексов (моделей эхоимпульсов пачки),
каждый из которых имел три импульсных компонента. Интервалы между комплексами в пачке - 10 мс. Импульсные
компоненты имели амплитуды Amp, Amp/10 и Amp/3. Задержки второго и третьего комплексов были равны 90 и 70 мкс.
Остальные обозначения, как на рис. 1.
хастическим способом. При таком способе каж-
ЗИ и прием пачек ЭИ позволяет дельфину надеж-
дое волокно в ансамбле могло ответить только
но обнаружить жертву на дальних дистанциях, а
один раз, поэтому эффекты маскировки отдель-
варьирование амплитуд ЭИ вследствие варьиро-
ных компонентов ЭИ при интервалах в 90 и
вания амплитуд ЗИ позволяет дельфину иденти-
70 мкс не возникали (рис. 3). Профиль реакция на
фицировать жертву на этапе обнаружения.
первый комплекс с амплитудой Amp в 3 дБ вос-
Воспроизведение временной структуры подпоро-
производил только два наибольших компонента.
говых импульсов. Интегрирующие свойства ВВК
Реакции на двух других комплекса с амплитудами
направлены на повышение отношения сиг-
Amp в 8 и 11 дБ выявили три компонента. При Amp
нал/помех, но могут быть полезными при обнару-
в 11 дБ реакция на средний наименьший компо-
жении эхоимпульсов дельфинами, например, в
нент комплекса была подчеркнута по сравнению
условиях реверберации.
таковым в профиле X(t). Это важное свойство ко-
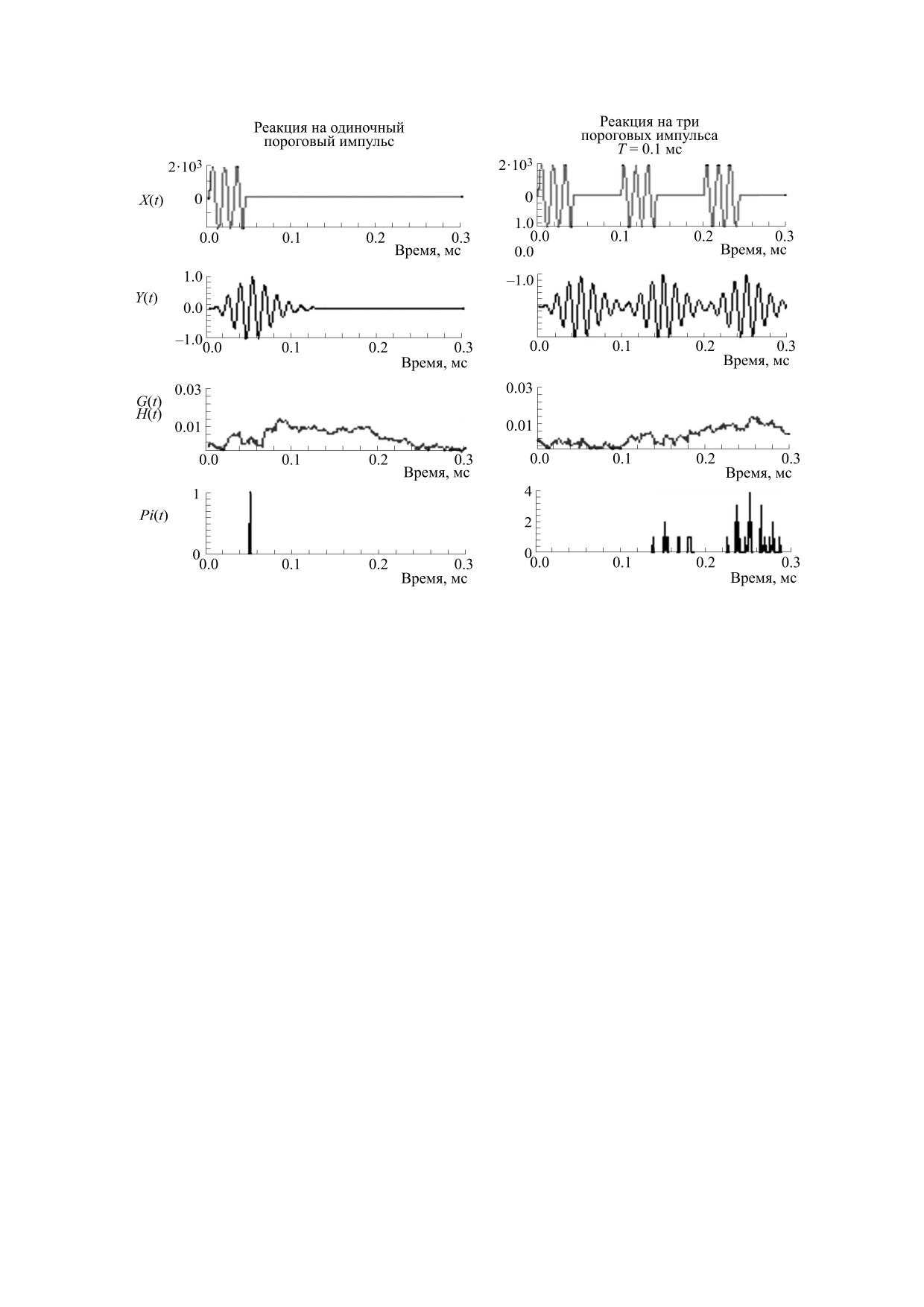
На рис. 4 сравниваются реакции, полученные
дирования импульсных звуков. За подчеркивание
на одиночный импульс, и три импульса с межим-
компонентов комплексов отвечали интегрирую-
пульсными интервалами в 100 мкс. Импульсы
щие свойства ВВК и стохастический способ ко-
имели подпороговые амплитуды. Одиночный
дирования в ансамбле ВСН.
импульс и три импульса могли быть закодирова-
Таким образом, при большой скважности пач-
ны только стохастическим способом. Реакция ан-
ки, состоящей из трех моделей ЭИ (рис. 3), пери-
самбля на одиночный импульс содержала
1
ферическое кодирование каждого из них осу-
спайк, возникший в случайный момент времени.
ществлялось стохастическим способом и не зави-
Однако реакция, вызванная тремя импульсами,
село от кодирования других импульсов.
точно воспроизвела реакцию на третий импульс,
Близкорасположенные импульсные компоненты
поскольку рецепторный потенциал G(t) имел на-
моделей ЭИ также не маскировали друг друга. Ва-
растающий профиль. Таким образом, воспроиз-
риации параметра Amp способствовали точности
ведению временной структуры импульсов спо-
воспроизведения временной структуры модели
собствовали интегрирующие свойства рецептор-
ЭИ. Можно предположить, что излучение пачек
ных потенциалов G(t) и стохастический способ
БИОФИЗИКА том 66
№ 2
2021
ПОДСТРОЙКА ЭХОИМПУЛЬСОВ
379
Рис. 4. Реакции разных блоков модели на одиночный импульс и три пороговых импульса с межимпульсными
интервалами в 100 мкс. Остальные обозначения, как на рис. 1.
кодирования импульсов в ансамбле, а также соот-
Таким образом, интегрирующие свойства ре-
ветствие потенциалов G(t) порогу реакции боль-
цепторных потенциалов G(t) и увеличение числа
шинства волокон в ансамбле (рис. 4). Группа из
волокон в ансамбле способствует повышению от-
трех импульсов могла воспроизводить случай об-
ношения пиковых амплитуд эхоимпульсов к
наружения эхоимпульсов в присутствии отраже-
уровню шума, при котором возможно воспроиз-
ний у дельфинов.
ведение временной структуры эхоимпульсов.
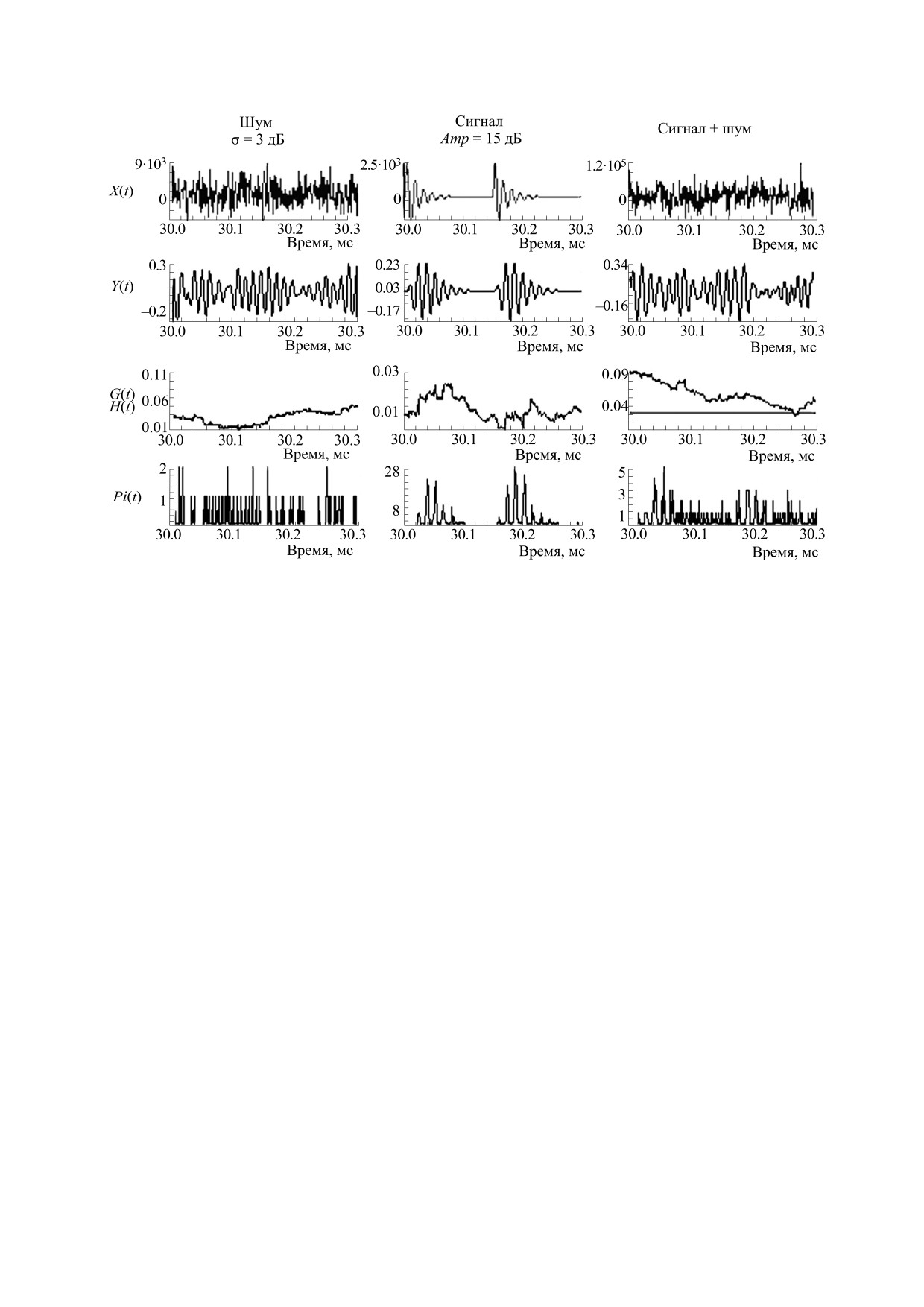
Воспроизведение временной структуры импуль-
сов в шуме. Пример выделение временной струк-
ОБСУЖДЕНИЕ
туры пары равновеликих импульсов в шуме пока-
Рассмотрим значение и место данной работы в
зан на рис. 5.
широком поле исследований эхолокационных
Дисперсия шума в этом примере соответство-
способностей дельфинов.
вала пиковой амплитуде импульсов. Поэтому
Целенаправленные изменения частоты следо-
профиль пары импульсов не проявился в профи-
вания и интенсивности зондирующих импульсов
ле смеси сигнала и шума (в функции X(t)). Однако
при лоцировании дельфинами объектов в экспе-
модели волокон обладали свойствами адаптации,
риментальных условиях выявлены давно [1, 2,
т.е. волокна могли менять свою чувствительность
22]. У диких дельфинов разных видов обнаружена
под действием продолжительных тонов или шу-
динамическая связь между частотой повторения,
мов (в моделях волокон повышение потенциалов
интенсивностью излучаемых зондирующих им-
G(t) вызывало повышение порогов H(t)). Пару
пульсов и дистанцией до объекта локации [8, 9].
импульсов предъявляли через 30 мс после начала
Специальная система регистрировала зондирую-
шума, т.е. после изменения порогов H(t) волокон.
щие импульсы (ЗИ), оценивала расстояние до
Поэтому ансамбль ВСН мог выявить присутствие
пары импульсов в шуме. Чем больше волокон в
дельфина, интенсивность (уровень звукового
ансамбле (в приведенном примере число волокон
давления) и частоту повторения ЗИ. По мере при-
было увеличено в три раза), тем точнее ансамбль
ближения к объекту дельфины корректировали
воспроизводил структуру входного сигнала.
частоту повторения. Интенсивности ЗИ, как пра-
БИОФИЗИКА том 66
№ 2
2021
380
РИМСКАЯ-КОРСАКОВА и др.
Рис. 5. Реакции разных блоков модели на полосовой шум, пару импульсов с межимпульсным интервалом в 150 мкс и
смесь пары импульсов и шума. Остальные обозначения, как на рис. 1.
вило, были высокими при низкой частоте повто-
частотой повторения. Авторы предположили, что
рения, но уменьшались при увеличении частоты
дельфины способны поддерживать излучаемую
повторения. Изменчивость интенсивности ЗИ
акустическую энергию относительно постоянной
при изменении дистанции до объекта наилучшим
для каждого цикла повышения давления, поэто-
образом аппроксимировала функция вида
му амплитуда ЗИ могла быть высокой при низкой
Amp(R) = 20logR, где R - дистанция. Выявленная
частоте повторения, но должна снижаться по ме-
зависимость указывала, что эхолокатору дельфи-
ре увеличения частоты повторения. Было также
на присущи свойства систем с автоматической
показано [8, 9], что пиковые амплитуды ЗИ, заре-
регулировкой усиления (АРУ). Однако регули-
гистрированные на одном и том же расстоянии до
ровка осуществлялась не при приеме эхоимпуль-
объекта, варьировали в диапазоне ±20 дБ. При-
сов, а при излучении зондирующих импульсов.
чины таких значительных вариаций точно неиз-
Это явление назвали динамическим управлением
вестны.
уровня эхолокационного источника. Есть мнение
В данной работе приводятся аргументы в поль-
[23], что динамическое управление является не
зу целесообразности вариаций интенсивности
результатом познавательного процесса, а есте-
зондирующих импульсов. Интенсивность эхоим-
ственным следствием способа генерации ЗИ у
пульсов зависит от интенсивности ЗИ, расстоя-
дельфинов. Дельфины генерируют каждый сле-
ния до объекта, силы цели объекта, поглощения
звука в среде. При отсутствии априорной инфор-
дующий ЗИ после приема отраженного от объек-
мации об объекте вариации интенсивности ЗИ
та ЭИ, поэтому частота повторения ЗИ повыша-
могут помочь дельфину выявить объект за счет
ется при приближении дельфина к объекту.
подстройки ЭИ под слуховую чувствительность,
Щелчки генерируются в назальной системе дель-
имеющую место в точке приема ЭИ. Однако надо
фина путем управления потоком воздуха через
признать, что значительные вариации интенсив-
акустические губы. Сначала дельфин создает по-
ности зондирующих импульсов у дельфинов име-
вышенное давление в своей назальной системе, а
ют место как на стадии поиска объекта, так и по-
затем испускает серию ЗИ с относительно низкой
сле его обнаружения.
БИОФИЗИКА том 66
№ 2
2021
ПОДСТРОЙКА ЭХОИМПУЛЬСОВ
381
Другим аргументом в пользу целесообразно-
при предъявлении тонального сигнала дельфин
сти варьирования интенсивности ЗИ мог быть
лоцировал пространство, в котором присутство-
способ кодирования высокочастотных импуль-
вала или отсутствовала мишень. При отсутствии
сов во внутреннем ухе дельфинов, направленный
мишени пороги были ниже. По мнению авторов,
на сохранение максимально полной частотной и
причиной наблюдаемых изменений порогов об-
временной информации об ЭИ. Под кодировани-
наружения могли быть стремянной рефлекс сред-
ем мы понимали преобразование аналогового ре-
него уха или эффект слуховой адаптации. Эти эф-
цепторного потенциала внутренних волосковых
фекты могли подавлять реакцию на ЭИ.
клеток в последовательность потенциалов дей-
Действительно, адаптацию, которая снижает
ствия (спайков) ансамбля волокон слухового нер-
слуховую чувствительность, вызывают длитель-
ва. Одиночное инерционное волокно не может
ные звуковые воздействия. На рис. 5 в данной ра-
воспроизвести временную структуру высокоча-
боте приведен пример, в котором адаптация, вы-
стотного звука, но, согласно принципу, это мо-
званная шумом, способствовала обнаружению
жет сделать реакция ансамбля ВСН. Принцип
импульсного сигнала. Помимо этого, проведен-
залпов был декларирован, но специально не ис-
ные исследования [27, 28] показали, что благода-
следован. Морфологические изменения в улитке
ря свойствам адаптации выделение и анализ им-
дельфинов подчеркивают роль реакции ансамбля
пульсных звуков, предъявляемых на фоне поме-
ВСН в слуховом анализе ЭИ. Количество рецеп-
ховых сигналов, может быть лучше, чем в
торных ВВК у дельфинов сопоставимо с таковым
тишине.
у других млекопитающих, в том числе человека,
Известно, что громкие звуки провоцируют
но число клеток спирального ганглия улитки,
связанных с одной рецепторной клеткой, у дель-
стремянной рефлекс, вызывающий защитное
фина заметно больше [24-26]. На ~3500 волоско-
снижение слуховой чувствительности уже на
вых клеток у человека приходится ~30000 спи-
уровне среднего уха. У летучих мышей чувстви-
ральных нейронов, а у афалины - ~95000 спи-
тельность слуха понижается за счет сокращения
ральных нейронов. В данной работе показано,
мышц в среднем ухе непосредственно перед излу-
что для точного воспроизведения тонкой времен-
чением ЗИ [29]. Однако роль среднего уха и меха-
ной структуры ЭИ в реакции ансамбля ВСН важ-
низмы его функционирования у дельфинов не яс-
на подстройка рецепторного потенциала G(t) под
ны [24, 25]. Цепочка слуховых косточек усилена и
чувствительность большинства волокон ансам-
жестко связана с кольцевыми связками, а сами
бля. При такой подстройке реализуется стохасти-
косточки плотнее и массивнее, чем у наземных
ческий способ кодирования ЭИ, эффективность
млекопитающих соответствующего размера. Та-
которого тем выше, чем больше число волокон в
кие изменения могут препятствовать рефлексу
ансамбле. Однако не только вариации интенсив-
среднего уха [23].
ности ЗИ могут способствовать подстройке.
Путем регистрации слуховых вызванных по-
ЗАКЛЮЧЕНИЕ
тенциалов у дельфинов [4, 5, 20] показано, что на
1. Слуховое периферическое кодирование вы-
коротких дистанциях до объектов за такую под-
сокочастотных эхолокационных импульсов
строку могло отвечать взаимодействие ЭИ и ЗИ.
включает в себя преобразование рецепторных по-
Высокоинтенсивный ЗИ мог маскировать ЭИ.
тенциалов волосковых клеток в последователь-
Согласно полученным нами данным, основой та-
ность потенциалов действия (спайков) ансамбля
кой маскировки, могли быть рефрактерные свой-
волокон. Моделирование показало, что ансамбль
ства волокон ансамбля, вызванные зондирую-
волокон может кодировать короткие звуки двумя
щим импульсом. За счет этих свойств, пороги ре-
способами. Низкоамплитудные импульсы могут
акции волокон в ансамбле повышались и
быть закодированы ансамблем волокон стохасти-
подстраивались под вызванный эхоимпульсом
ческим способом. При таком способе каждое во-
локно в ансамбле отвечает один раз, поэтому со-
рецепторный потенциал волосковых клеток, по-
ставляющие импульсных звуков не маскируют
этому реакция ансамбля воспроизвести времен-
друг друга. Этот способ кодирования позволяет
ной профиль ЭИ (рис.2).
восстановить тонкую временную структуру ко-
Еще один вариант слухового взаимодействия
роткого импульса. Восстановление тем точнее,
ЗИ и ЭИ на коротких дистанциях до объектов
чем ближе амплитуда одиночного импульса к по-
рассмотрен в работах [5, 6]. Авторы оценили слу-
рогу реакции большинства волокон слухового
ховую чувствительность касатки по порогам по-
нерва. Способ кодирования пар высокоампли-
явления СВП в ответ на тональные сигналы, ко-
тудных импульсов с интервалами, превышающи-
торые излучали во время лоцирования животным
ми единицы миллисекунд, детерминистический.
мишеней. Показано, что пороги возникновения
В этом случае в ответ на пару импульсов волокна
СВП, вызванные тональным сигналом одной и
в ансамбле отвечают более одного раза. Реакция
той же интенсивности, различались на 20 дБ, если
на второй импульс определяется свойствами вос-
БИОФИЗИКА том 66
№ 2
2021
382
РИМСКАЯ-КОРСАКОВА и др.
становления возбудимости волокон ансамбля
4. A. Ya. Supin, P. E. Nachtigall, and M. Breese, J.
(рефрактерностью и адаптацией), вызывающими
Acoust. Soc. Am. 115, 3218 (2004).
снижение возбудимости волокон после реакции
5. A. Ya. Supin, P. E. Nachtigall, and M. Breese, J.
на первый импульс. Поэтому восстановление
Acoust. Soc. Am. 123, 534 (2008).
временного профиля второго импульса пары мо-
6. В. В. Попов и А. Я. Супин, Слух китов и дельфинов
жет зависеть от амплитуд импульсов пары и за-
(ООО «Товарищество научных изданий КМК»,
держки между ними.
М., 2013).
2. Моделирование воспроизводит стабилиза-
7. Н. А. Дубровский, Л. К. Римская-Корсакова и
Р. П. Трошенкова, в сб. Докл. XI Всесоюз. акустич.
цию слуховых реакций на ЭИ, наблюдаемую при
конф-ии, секция Ц (М., 1991), сс. 9-12.
коротких задержках Т между ЗИ и ЭИ. Пара им-
8. W. W. L Au and K. J. Benoit-Bird, Nature 423, 861
пульсов (ЗИ и ЭИ) закодирована ансамблем во-
(2003).
локон детерминистическим способом. Реакция
на второй импульс (ЭИ) формируется с учетом
9. S. Li, D. Wang, K. Wang, and T. Akamatsu, J. Acoust.
Soc. Am. 120, 1803 (2006).
рефрактерных свойств волокон ансамбля, возни-
кающих под действием первого импульса, а также
10. Л. К. Римская-Корсакова и Н. А. Дубровский,
Акустич. журн. 52, 521 (2006).
свойствами распространения ЭИ в воде. Такое
взаимодействие может подстроить чувствитель-
11. М. П. Иванов, в сб. Труды IY межд. конф. «Морские
ности слуховой системы дельфина под амплитуду
млекопитающие Голарктики» (СПб, 2006), сс. 218-
222.
отраженного от объекта ЭИ и обеспечить ста-
бильную реакцию на ЭИ во время охоты в фазе
12. P. M. Sellick and I. J. Russell, Hear. Res. 2 (3-4), 439
(1980).
захвата.
13. Л. К. Римская-Корсакова и Н. А. Дубровский,
3. На средних дистанциях до объектов дельфи-
Сенсорные системы 4, 92 (1990).
ны излучают следующий ЗИ после приема ЭИ, но
14. N. A. Dubrovsky and L. K. Rimskaya-Korsakova, in
варьируют интенсивность ЗИ в широком диапа-
Marine Mammal Sensory Systems (Plenum Press, N.Y.,
зоне. Моделирование указывает на целесообраз-
1992), pp. 223-233.
ность варьирования амплитуд ЗИ. Варьирование
15. N.A. Dubrovsky and L.K. Rimskaya-Korsakova, Neu-
представляется механизмом подстройки интен-
ral Network World 6, 577 (1996).
сивности ЭИ под чувствительность слуха дельфи-
16. H. Г. Бибиков и Г. А. Иваницкий, Биофизика 30,
на в точке приема, сформированную окружаю-
141 (1985).
щими шумами.
17. Н. Г. Бибиков, Л. К. Римская-Корсакова, А. В. За-
Для обнаружения объектов на дальних дистан-
нин и Н. А. Дубровский, в сб. Электрофизиология
циях или объектов с малыми силами цели на
сенсорных систем морских млекопитающих, под
средних дистанциях дельфины могут излучать
ред. В. Е. Соколова (Наука, М., 1986), сс. 56-84.
пачки переменных по амплитуде ЗИ. Излучение
18. Л. К. Римская-Корсакова, Сенсорные системы 12,
пачек ЗИ и прием пачек ЭИ помогает дельфину
376 (1998).
надежно обнаружить жертву на дальних дистан-
19. Л. К. Римская-Корсакова и Н. А. Дубровский,
циях, а варьирование амплитуд ЗИ (и поэтому
Сенсорные системы 17, 1 (2003).
ЭИ) позволяет идентифицировать жертву на эта-
20. L. Robles and M. A. Ruggero, Physiol. Rev. 81, 1305
пе обнаружения.
(2001).
21. Л. К. Римская-Корсакова, Акустич. журн. 63, 436
КОНФЛИКТ ИНТЕРЕСОВ
(2017).
22. Л. Р. Гиро и Н. А. Дубровский, Морское приборо-
Авторы заявляют об отсутствии конфликта
строение, сер. Акустика. 2, 84 (1972).
интересов.
23. S. H. Ridgway, W. W. L. Au, Encyclopedia of Neurosci-
ence 4, 1031 (2009).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
24. D. R. Ketten, Bioacoustics 8, 103 (1997).
Настоящая работа не содержит описания ис-
25. D.R. Ketten, in Hearing bv Whales and Dolphins, Ed. by
следований с использованием людей и животных
W. W. L. Au, A. N. Popper, and R. R. Fay, (Springer-
Verlag, N.Y., 2000), pp. 43-108.
в качестве объектов.
26. E. G. Wever, J. G. McCormic, H. Palin, and
S. H. Ridgway, Proc. Math. Acad. Sci. USA 68, 2381
СПИСОК ЛИТЕРАТУРЫ
(1971).
1. В. М. Белькович и Н. А. Дубровский, Сенсорные
27. Л. К. Римская-Корсакова, Изв. РАН. Сер. физ. 70,
основы ориентации китообразных (Наука, Л., 1976).
138 (2006).
2. W.W. L. Au, Sonar of Dolphins (Springer, N. Y., 1993).
28. L. K. Rimskaya-Korsakova, J. Optic. Technol. 72, 383
(2005).
3. В. М. Белькович, Е. Е. Иванова и О. В. Ефремен-
кова, в сб. Поведение и биоакустика дельфинов
29. L. A. Simmons, A. J. M. Moffat, and W. M. Masters, J.
(ИОАН, М., 1978), сс. 67-78.
Acoust. Soc. Am. 91, 1150 (1992).
БИОФИЗИКА том 66
№ 2
2021
ПОДСТРОЙКА ЭХОИМПУЛЬСОВ
383
Distance-Dependent Gain Control of Echo-Pulses to Auditory Sensitivity in Dolphins:
Simulation Experiments
L.K. Rimskaya-Korsakova*, A.A. Rodionov**, and M.P. Ivanov***
*JSC Andreyev Acoustics Institute, ul. Shvernika 4, Moscow, 117036 Russia
**Saint Petersburg Branch of the Institute of Oceanology, Russian Academy of Sciences,
Universitetskaya nab. 5, St. Petersburg, 199034 Russia
Saint Petersburg State University, Universitetskaya nab. 7/9, Saint-Petersburg, 199034 Russia
Dolphins use echolocation during hunting. They emit acoustic pulses and analyze returning echoes to detect
and identify the prey during the search phase. However, in the hunting capture phase, dolphins must have a
stable sonar system in response to echo information they receive. In order to explain the features of echolo-
cation in dolphins during the search and capture phases, the responses of the ensemble of high-frequency pe-
ripheral fibres were simulated. Pairs of pulses with central frequencies of 70 kHz, different inter pulse intervals
and peak amplitudes were models of probing and echo pulses of dolphins. Pulses arrived at the input of a
model with different stages of peripheral processing, including the transformation of the receptor potential of
inner hair cells into a sequence of action potentials (spikes) generated by the ensemble of auditory nerve fi-
bers. It is shown that an important condition for the reproduction of the fine temporal structure of an echo
pulse in a synchronous ensemble reaction is the correspondence of the arising receptor potential to the
thresholds of most fibers in the ensemble. For short delays between the pulses of the pair (in the capture
stage), this correspondence causes an increase in the fiber thresholds due to the refractory properties of the
fibers caused by the first pulse (models of the probing impulse). For long delays between the pulses of the pair
(in the search stage), variations in the peak amplitudes of echo pulses could help to such a correspondence
(models of the probing impulse). Modeling indicates a difference in the methods of matching echo pulses
with the auditory sensitivity of dolphins in the phases of search and capture of the prey.
Keywords: echolocation, peripheral coding, auditory nerve fibers, reproduction of the temporal structure of echo
pulses
БИОФИЗИКА том 66
№ 2
2021