БИОФИЗИКА, 2021, том 66, № 2, с. 384-407
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 595.76, 51.76, 532.22
НЕПРЕРЫВНАЯ МОДЕЛЬ ТРЕХ СЦЕНАРИЕВ ИНФЕКЦИОННОГО
ПРОЦЕССА ПРИ ФАКТОРАХ ЗАПАЗДЫВАНИЯ ИММУННОГО ОТВЕТА
© 2021 г. А.Ю. Пеpеваpюxа
Санкт-Петербургский институт информатики и автоматизации PАН,
199178, Санкт-Петербург, 14-я линия, 39
E-mail: temp_elf@mail.ru
Поступила в редакцию 08.06.2020 г.
После доработки 17.11.2020 г.
Принята к публикации 20.11.2020 г.
Рассмотрены модели развития инфекции как регулируемые нелинейные процессы. Актуальным
вопросом для анализа оказались существенные различия в динамике протекания вызванного коро-
навирусом нового заболевания. Множество факторов рассматриваются как причины явления, ко-
гда инфекция протекает столь вариативно у разных людей и распространяется в локальных популя-
циях с различной скоростью. Репликация копий вируса в зараженных им клетках и выработка отве-
та от иммунных клеток организма это два взаимообусловленных процесса с последействием и
зависящие от предшествующих состояний. Моделирование их свойств требует получения разных
сценариев без возмущений параметров. Важнейшим фактором является эффективность иммунного
ответа, включающая время реакции трех компонентов сложнейшей системы организма. Нами
предложено вычислительное описание сценариев развития инфекции на основе уравнения с двумя
параметрами запаздывания. Переходы между фазами течения инфекционного заболевания в моде-
ли обусловлены аспектами уровня первичного заражения и запаздывающей реакции при выработки
иммунного ответа. Вариации дозы заражения и времени ответа способны приводить к переходу
острой фазы заболевания с выраженными симптомами в хроническое течение либо к летальному
результату. В разработанной модели рассмотрен бессимптомный вариант развития - быстрого эф-
фективного подавления вируса после краткой фазы его активной репликации, но сохранении ма-
лого количества в организме. Анализ режимов поведения модели соответствует теории, что изна-
чальное количество вирионов способно повлиять на качество иммунного ответа. Индивидуальные
различия в протекании заболевания COVID-19 и формирование типов иммунного ответа к вирусу
явно велики, но сейчас недостаточно понятны. Модельные сценарии ориентируются на изученные
варианты протекания вирусного гепатита С.
Ключевые слова: моделирование инфекционных процессов, запаздывающая регуляция, иммунный ответ,
модель хронизации гепатита С, начальная доза заражения, эпидемии, скорость распространения виру-
са, бессимптомный сценарий, вариабельность COVID-19.
DOI: 10.31857/S0006302921020204
В статье развивается тематика исследований
ством темпы распространения инфекционных
предыдущей работы о моделировании экстре-
заболеваний в субпопуляциях. Заражение пато-
мальных и особых переходных режимов суще-
генным вирусом можно отнести к частной форме
ствования агрессивного вида в новой среде [1], но
инвазионных процессов, но протекающих с ак-
с использованием иного математического аппа-
тивным сопротивлением, где решается исход
рата для этой актуальной задачи. Для специфиче-
борьбы систем защиты организма и эволюциони-
ской биологической коллизии необходимо кон-
рующего в нем паразита.
струировать и выбирать наиболее подходящий
Цель работы заключается в обосновании фе-
метод моделирования. Общим должен оставаться
номенологической модели, способной описать
принцип рассмотрения логики развития ситуа-
видимые различия в форме протекания заболева-
ции - как набора вычислительных сценариев.
ния в зависимости от начальной дозы вируса при
Процессы взаимодействия видов редко протека-
инфицировании и времени, которое затрачено
ют по единственно возможному пути [2]. Значи-
для выработки эффективного ответа. Поэтому ес-
тельна вариативность развития динамики инва-
ли в модели вспышки численности вредителей
зий при вселении чужеродных видов в разные
леса мы использовали итерационную хаотиче-
ареалы [3]. Аналогично отличаются непостоян-
скую динамику траектории с набором управляе-
384
НЕПРЕРЫВНАЯ МОДЕЛЬ ТРЕХ СЦЕНАРИЕВ
385
мых бифуркаций, то для объяснения вариантов
По сравнению с эпидемической волной в
исхода в ситуации противоборства с вирусом
США в Мексике количество заболевших увели-
больше подходит непрерывная модель. Формали-
чивается на порядок медленнее. В США по об-
зуем запаздывающую регуляцию при формирова-
щим данным видим пример худшей формы стре-
нии ответной реакции в уравнении с учетом пред-
мительной эпидемии, реализации затяжного Π-
пороговой величины вирусной нагрузки, когда
образного пика. Разнообразие в формах каче-
влияние инфекции на организм становится зна-
ственной динамики эпидемий по отдельным
чимым фактором. Введенная предкритическая
штатам США столь сильно выражено, что специ-
концентрация вместе с двумя показателями за-
алистам трудно обосновать прогноз заболеваемо-
паздывания ответа должна различаться для раз-
сти в соседних штатах.
ных организмов, что объяснит вариативные сце-
Разница в темпах распространения вируса так
нарии протекания заболевания при различной
очевидна, что со стороны Мексики шутят о готов-
начальной дозе заражения. Модель разрабатыва-
ности самим достроить стену на границе. Вирус
лась для описания сценариев хронизации вирус-
оказался бессилен во Вьетнаме - на момент на-
ного гепатита С после острой фазы. В модели рас-
писания статьи ни одного летального исхода в их
смотрен «бессимптомный» вариант развития с
статистике. После 180000 проведенных тестов
сохранением возбудителя - актуальное обстоя-
20 апреля 2020 г. Вьетнам заявил об отражении
тельство необычного протекания новой болезни
угрозы и снятии ограничительных мер. Нацио-
COVID-19.
нальные стереотипы поведения не стоит игнори-
В год пандемии интересно обсудить, почему
ровать в предсказаниях. Итальянцы могли силь-
скорость распространения инфекции так нерав-
нее пострадать из-за своей общительности и раз-
номерна и при этом наблюдается несколько вари-
говорчивости
[6], и этому есть физическое
антов фазы затухания локальных эпидемий, в том
обоснование. В исследовании о путях передачи
числе с точки зрения эффективности ограничи-
вируса [7] объясняется, что при интенсивном раз-
тельных мер. В первой части статьи анализируем
говоре создаются вихри микроскопических ка-
текущую информацию о качественной динамике
пель слюны, разносящих вирус, и чем громче раз-
эпидемии вируса SARS-CoV-2, предложим неко-
говор, тем больше разбрасывается вирионов и
торые объяснения различий с точки зрения не-
выше вероятность переноса возбудителя.
определенности регуляции процессов. Отметим
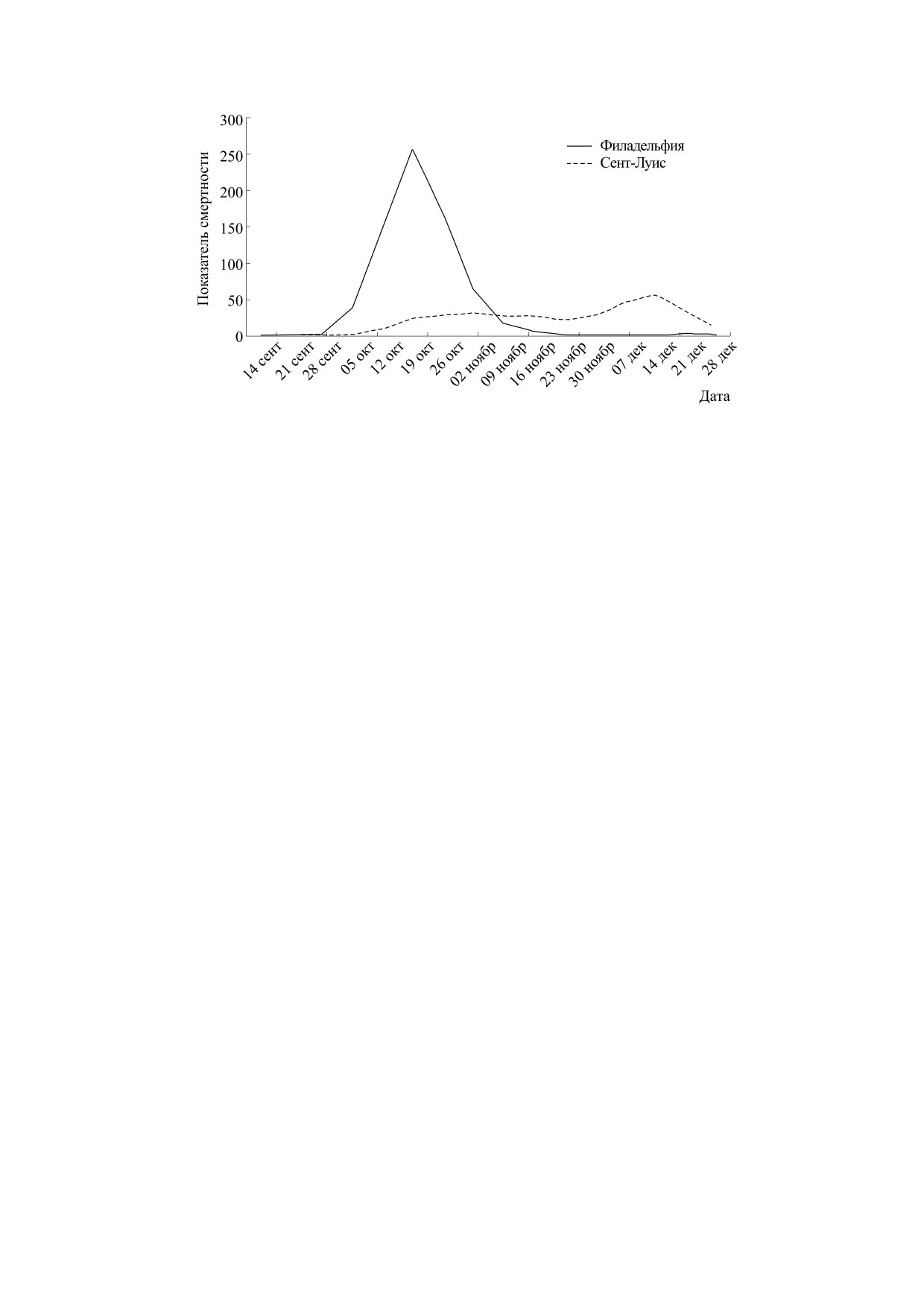
Пример разных последствий «испанки» в двух
необычные аспекты - переходные типы и мате-
городах США сто лет назад стал хрестоматийным.
матическую классификацию сценариев в отдель-
Графики эпидемии гриппа (считали смертность
ных регионах, на которые специалистам нужно
на 100000 человек) в Филадельфии и Сент-Луисе
обратить пристальное внимание при моделирова-
показывают два основных сценария. На дихото-
нии сроков завершения эпидемического процес-
мию пути развития эпидемического процесса
са и прогнозировании рецидивов распростране-
1919 г. опирались в марте директивы при вводе
ния нового РНК-вируса, возможно мутировав-
ограничений мобильности. Так обосновывают
шего.
изоляционные меры, которые успели ввести в
Сент-Луисе, и отменить все мероприятия и удач-
но избежали взрывообразной фазы процесса - с
РАЗЛИЧИЯ И ОБЩНОСТЬ
Λ-образным пиком (смертности). Объединять в
ЭПИДЕМИЧЕСКИХ ПРОЦЕССОВ
статистике столь несхожие локальные сценарии
ПРИ ГЛОБАЛИЗАЦИИ
совершенно нецелесообразно, агрегированные
данные по заболеваемости введут в заблуждение.
Давно известен факт, что эпидемии в странах
Спустя два месяца эпидемии единичный Λ-пик
развиваются по различным сценариям. Напри-
уже не кажется таким ужасным на фоне экономи-
мер, существенные отличия установлены для ви-
ческого коллапса. Во многих городах США «вы-
руса Эбола в регионах Африки [4]. Есть несколь-
ход на плато» сменяется вдруг повторным ростом.
ко отличительных черт и странных явлений у
Замедлившаяся динамика заражений коронави-
современной пандемии, что расширит простран-
русом неожиданно вновь ускоряется и таким об-
ство для построения гипотез. После окончания
разом дифференциация эпидемических сценари-
пандемии SARS-CoV-2 многие эпидемиологи и
ев в 2020 г. увеличивается.
математики продолжат несколько лет анализиро-
вать эффективность изоляционных мер в разных
Вопрос о причинах дихотомии эпидемических
странах, сравнивать графики и строить модель-
сценариев - краеугольный [8]. Либо в первые две
ные прогнозы для будущих эпидемий, проводя
недели произойдет стремительный рост, когда
обработку текущих данных о новом бетакорона-
ресурсы системы здравоохранения не справ-
вирусе; разумеется, они придут к противополож-
ляются с потоком тяжелых больных, либо про-
ным выводам, что мы уже видели по многим дру-
цесс перейдет в плавную (или спорадическую)
гим примерам [5].
форму. Отметим, что Филадельфия с классиче-
БИОФИЗИКА том 66
№ 2
2021
386
ПЕPЕВАPЮXА
Рис. 1. Сравнение развития в 1918 г. эпидемии испанки (по смертности) в двух городах США.
ским Λ-пиком является портовым городом и цен-
Обоснована теория [10], что эпидемия (как и
тром коммуникаций. Следовательно, изначально
вспышка численности инвазионного вида) стре-
в исходных условиях для запуска эпидемии в двух
мительно ускоряется при достижении некоторо-
городах предполагается существенное отличие.
го порога - концентрации заболевших в по-
Каждый исходный распространитель вызывает
пуляции - и аналогично резко затухает, когда
цепочку заражений и иногда непредсказуемо
образовывается достаточный коллективный им-
массовых.
мунитет у переболевших и невосприимчивых.
Математически идея базируется [11], в том чис-
Логично, что в портовый город проникает
ле, на модификациях пороговой теоремы Уитла,
большее количество инфицированных и из оча-
когда эпидемия представляется как ветвящийся
гов заражения, возможно, скрытых носителей,
случайный процесс [12], в котором для всех есть
так называемых «нулевых пациентов», что может
априорная вероятность перехода узла из времен-
сократить интервал перехода к ускоряющемуся
ного состояния «больной» в положение «имму-
росту числа заражений и не оставить времени для
низированный» [13].
принятия необходимых мер. На графики, приве-
После достижения 500 случаев в день эпиде-
денные на рис. 1, часто ссылаются как на нагляд-
мия в США, судя по обновляемому графику, дей-
ный назидательный пример, но нужно признать,
ствительно резко ускорилась. Число заболеваю-
что два города в точке отсчета не были в одинако-
щих в сутки моментально увеличилось в четыре
вых исходных условиях. Точно не доказано, отку-
раза. Это выглядит на графике как «скачок» через
да появился вирус в 1918 г. и был ли повар военно-
порог, где за созданием критической концентра-
го лагеря в Канзасе первой жертвой. После по-
ции стоит цепь событий. На самом деле эти люди
вторного пика в крупных городах с большей
болели уже давно, так как график выявленных
смертностью «испанка» загадочно навсегда ис-
случаев - это фиксация состояния эпидемиче-
чезла из Европы в июне 1919 г. [9], хотя «синюю
ского процесса (с неполными данными) с не-
смерть» ждали и в 1920 г. Прогнозировать время
определенным запаздыванием. Реальное порого-
завершения эпидемии - сложная и достаточно
вое состояние - концентрация инфицирован-
многофакторная задача для детерминированных
ных, после которого запущена эруптивная фаза
моделей.
вспышки в очаге, пройдено немного левее по оси
времени и при меньшем числе (фиксированных)
Критическая ситуация в США с коронавиру-
случаев заражения.
сом наблюдалась Нью-Йорке в апреле 2020 г. Это
крупнейший центр китайской диаспоры и исто-
На обновляемом графике текущей статистики
рически эмигрантские ворота Северной Амери-
по странам мира (например, на сайте Financial
ки. Рано или поздно новый вирус привезут
Times) видно, что карантинные ограничительные
повсюду - это следствие глобализации. В неко-
меры эффективно сработали, если были введены
торые города прибудет сразу много распростра-
после первых единичных случаев, как было четко
нителей вируса. Цепочки распространения умно-
организовано в Южной Корее. Когда «stayhome»
жатся, и тогда местные больницы окажутся сразу
вводится позже и вяло, то это совсем не быстро
перед фактом серии эпидемических волн.
меняет качество процесса. Эпидемический про-
БИОФИЗИКА том 66
№ 2
2021
НЕПРЕРЫВНАЯ МОДЕЛЬ ТРЕХ СЦЕНАРИЕВ
387
Рис. 2. Моделирование порогового сценария популяционной вспышки затуханием (из работы [1]).
цесс оказывается инерционным. Ирландия ввела
Как оказалось, китайские антиэпидемические
«stayhome» 20.03.2020 г. и ужесточила карантин
методы работают только в Китае. Гонконг и Син-
24.03.2020 г. К 08.04.2020 г. Ирландия вышла на
гапур уже заявляли о победе над эпидемией, но
«плато» - по гладкой логистической кривой было
власти Сингапура ждут повторные вспышки из-
380 новых случая, потом эпидемия вдруг резко
за потребности в иностранных рабочих. После
ускорилась и 09.04.2020 г. наблюдалось 500 случа-
пика статистика ежедневных случаев заражения
часто становится перемежающейся, действует
ев, а 11.04.2020 г. уже оказалась превышена «пси-
хологически важная отметка» в 1500 случаев за
фактор неоднородной задержки в процедуре вы-
явления. Чаще больной выявляется, когда возни-
сутки. Ирландия - интересный пример, когда до-
кают симптомы и уже есть иммунный ответ. Ва-
стижение модельного равновесия, где рост дол-
жен фактор скрытого развития инфекционного
жен остановиться («плато»), оказывается вдруг
процесса внутри организма, что создает эффект
пороговым значением и неожиданно для всех
комплексного запаздывания при распростране-
резко ускоряет эпидемию. К 05.05.2020 г. суточ-
нии, потому временный спад числа заражений не
ные показатели Ирландии так и не снизились до
нужно трактовать как успех. В Иране с июня со-
уровня, наблюдавшегося 8 апреля. Вероятно,
общают об уже очевидной «второй волне» эпиде-
итоговые показатели по смертности на 100000 че-
мии после затишья.
ловек не будут сильно различаться с «бескаран-
тинной» Швецией. Гипотеза, которая возникает
Эпидемия в Германии развивалась по плавно-
му сценарию (тут акцентируем внимание на вто-
после анализа, состоит в том, что если принцип
рой производной) по сравнению с Италией и Ис-
«сидим дома» запоздал, то пика никак не избе-
панией, где от старта до первого максимума про-
жать, работает принцип «если опоздали, то опоз-
шло соответственно 16 и 20 дней. Сценарий с
дали».
быстрым пиком похож на пороговый запуск
Швеция не вводила принцип «сидим дома»,
вспышки численности после длительной стаби-
там не было классического эпидемического Λ-пи-
лизации в окрестности неустойчивого равнове-
ка, но будет тянуться долго перемежающаяся во-
сия из нашей предыдущей работы [1] (см. рис. 2).
После «плато» здесь развивается турбулентный
латильная динамика роста/сокращения новых
режим.
случаев. 21.04.2020 г. в Швеции было выявлено
700 заболевших, а в Италии, где давно введено
В случае эпидемии сходство наблюдается в не-
положение «stai a casa», - 3500 случаев. При каче-
которых случаях у фазы порогового перехода от
ственном анализе эпидемического процесса дан-
флуктуаций к быстрому росту. Завершение эпи-
ные заболеваемости по мегаполисам нужно ана-
демии редко бывает аналогично резким, как при
лизировать отдельно от агрегированной стати-
падении численности размножившихся насеко-
стики по всей стране, что особенно актуально для
мых. Так, в Италии и в Испании долгое медлен-
Москвы, Нью-Йорка, Мехико, где плотность на-
ное снижение ежедневных случаев перемежается
селения много больше, чем в среднем по стране.
локальными пиками.
Некоторые страны (Франция) вносят задним
COVID-19 распространяется быстрее «свиного
числом сразу ранее пропущенные случаи, что ис-
гриппа» cо средним инкубационным периодом
кажает сценарий процесса и создает ложные мак-
5.5 суток [14], но с диапазоном инкубационного
симумы в статистике заражений.
периода от 0 до 15 суток (по данным из Уханя
БИОФИЗИКА том 66
№ 2
2021
388
ПЕPЕВАPЮXА
[15]). Уже в Ухане отмечали возможностью пере-
лей вариаций вирусного гликопротеина, у других
дачи вируса на бессимптомной стадии - при
авидитет получается с малой долей вирионов.
развитии инфекции при попустительстве адап-
Иммунитет реагирует на антигены, отдельные ча-
тивного иммунитета. Интервал до основной не-
сти белков возбудителя, но эти участки бывают
специфической реакции организма на респира-
изменчивы. Одни участки изменятся быстро, а
торную инфекцию - повышения температуры (и
другие консервативны. Количественные показа-
до выброса пирогенных интерлейкинов) у
тели иммунного ответа по антителам не говорят
COVID-19 в среднем больше [16], чем был в
об эффективности подавления вируса, когда не
2003 г. у родственного SARS-вируса. В работе [17]
находится стабильно аффинных вариантов имму-
отмечено, что нейтрализующие антитела были
ноглобулинов. Это вероятностный фактор эф-
обнаружены (с использованием псевдовирусных
фективности ответа. Известны ситуации, когда
частиц) в 79, 92 и 98% проб, взятых соответствен-
вирус уходит, но спровоцированное системное
но через 13-20, 21-27 и 28-41 сутки после появ-
воспаление останется.
ления симптомов у зараженных сотрудников
В 2020 г. сложилась уникальная возможность
больниц, которым не требовалась госпитализа-
для специалистов по моделированию следить за
ция. Это однозначно медленная реакция по срав-
эпидемическими траекториями стран в реальном
нению с ответом на большинство вирусов острых
режиме и делать быстро проверяемые прогнозы,
респираторных инфекций. О T-киллерах данных
но предсказания рушатся одно за другим из-за
нет. По важной, но неясной еще причине медиа-
особых свойств COVID-19. В сложившейся ситу-
торы иммунитета значительной доли инфициро-
ации информативными данными являются изме-
ванных не сильно торопятся активироваться в от-
нения доли положительных тестов от числа про-
вет на SARS-CoV-2. Вирусу это выгодно для
веденных за сутки, но такие сведения не публику-
скрытого распространения, но может оказаться,
ются. Невозможно строить долгосрочные
что видимая неспешность ответа предохраняет от
модельные прогнозы по будущему COVID-19, по-
большего риска, когда начальная доза не расце-
ка не установлено главное - как формируется им-
нивается как критическая.
мунитет, с какой скоростью реагируют разные
Биологические причины такой большой для
компоненты системы и как долго сохраняется за-
щита у переболевших. Интересно для моделиро-
острых респираторных вирусных инфекций дис-
вания получить данные о самой ранней стадии
персии латентной фазы болезни еще долго оста-
нутся предметом исследований и дискуссий ме-
развития COVID-19, так как в нелинейных систе-
диков. Уделяют внимание необычным сопутству-
мах переходные режимы и пороговые эффекты
ющим признакам - другим невыраженным и
распространения инфекции могут зависеть от на-
чальных условий. Другой важный вопрос - влия-
необычным для острых респираторных вирусных
инфекций симптомам, таким как потеря обоня-
ет ли начальная доза заражения и на скорость раз-
ния и вкуса [18]. Разумно предположить, что есть
вития пневмонии, и на иммунный ответ, который
пороговое значение концентрации вирионов, по-
бывает агрессивен сродни цепной ядерной реак-
сле которого резко проявляются симптомы. В ра-
ции и притом безрезультатен. Актуальная модель
боте [19] итальянскими медиками опубликованы
вариантов развития нового заболевания должна
обладать чувствительностью к N(0).
неожиданные результаты. Они дважды протести-
ровали большую часть (90 и 80%) жителей муни-
ципалитета. 40% выявленных случаев названы
ОСОБЕННОСТИ ВОСПРИЯТИЯ
бессимптомными. Итальянцы утверждают, что
ЭПИДЕМИЧЕСКОЙ СТАТИСТИКИ
определенная ими вирусная нагрузка у носителей
вируса с явными симптомами и «бессимптомны-
Наличие статистических данных о распростра-
ми» людьми не отличалась. Наблюдение сейчас
ненности вируса не гарантирует правильных вы-
не имеет достоверного объяснения. Осторожнее
водов. Систематически тестировать все населе-
говорить не о «бессимптомном», а о вялотекущем
ние начали сразу только Исландия и отчасти Ав-
сценарии болезни. Математически это значит со-
стралия, когда поняли, что инфицированный
хранение репликации вируса в стабильном рав-
через три дня уже является распространителем.
новесии (на неопределенный срок). Если симп-
Для будущих работ по анализу COVID их данные
томы не появились через неделю, не факт, что
послужат эталоном статистики. На острове поня-
они не проявятся через год. Есть вирусы, которые
ли, что изолировать нужно инфицированных, а
годами пребывают в субклинической стадии (на-
не здоровых. Деньги же эффективнее тратить на
пример, ВИЧ). Другие средства сдерживания ин-
средства для массового тестирования. На рис. 3
фекции могут начать истощаться или вирус смо-
приведено сравнение с данными процентного
жет их обманывать путем антигенного дрейфа в
распределения коронавирусной статистики Ни-
организме. Объясняется различие скорости про-
дерландов, где тестировали только попадавших в
грессирования болезни ВИЧ тем, что у одних лю-
лечебные учреждения с симптомами острых ре-
дей антитела способны связываться большей до-
спираторных вирусных инфекций. Не зная этого
БИОФИЗИКА том 66
№ 2
2021
НЕПРЕРЫВНАЯ МОДЕЛЬ ТРЕХ СЦЕНАРИЕВ
389
Рис. 3. Распределение относительного числа заражений коронавирусом по возрастным группам по данным из
факта, впоследствии биолог или статистик может
вое репродуктивное число явно увеличилось [19].
решить, что популяция исландцев имела какие-
Пусть среднестатистический больной COVID-19
то генетические отличия в восприимчивости к
заражает 3.5 других, но есть способные заразить и
вирусам, накопленные в долгой изоляции на ост-
намного больше. Существует гипотеза суперрас-
рове при низком генетическом разнообразии.
пространителей и указываются примеры такой
активности [21]. Так, например, объясняется воз-
Данные из Исландии разрушают миф о том,
никшая эпидемия ВИЧ в 1980 г. из-за действий
что коронавирусом болеют чаще люди старшей
одного канадского бортпроводника, заразившего
возрастной группы. Чаще болеют люди среднего
сотни человек по всей Северной Америке, хотя
возраста. У пожилых в среднем интенсивнее про-
вирус попал в США десятилетиями ранее. Пер-
являются осложнения. Тогда если тестировать
вый подтвержденный случай ВИЧ был датирован
только тех, кто обратился к врачам, то максимум
1966 г. в Сент-Луисе, но быстрая фаза эпидемии
придется на 60+. COVD - это не первая инфек-
не разразилась во время «лета любви» 1969 г., а
ция, которую более юные переносят легче. Мно-
медленно тлела целых 11 лет. В XIX в. описаны
гие знают, как неприятно заболеть краснухой в
единичные случаи распространителей брюшного
зрелом возрасте. Наш иммунитет - обучаемая са-
тифа, которые от симптомов тифа не страдали, но
монастраиваемая система защиты из нескольких
вызывали массовые заражения.
уровней, которой легче воспринимать и побеж-
дать новое в молодости, чем в дальнейшем срочно
В классических моделях с использованием си-
переучиваться для внезапных угроз. Когда суще-
стем обыкновенных дифференциальных уравне-
ствует простое объяснение, то для математиче-
ний триггерные эффекты и импульсную актив-
ских моделей лучше использовать наиболее ло-
ность учесть затруднительно. Актуально рассмат-
гичные связи между фактами.
ривать именно импульсные сценарии с учетом
массовых случаев одномоментного заражения и
их пролонгированных последствий, эффект
ПРОБЛЕМЫ ПРОГНОЗИРОВАНИЯ
инерции в динамике эпидемии. Триггерами уско-
И СОПОСТАВЛЕНИЯ ПОСЛЕДСТВИЙ
рения эпидемии стали футбольные матчи: напри-
ЭПИДЕМИЙ
мер, Аталанта против Валенсии. Одно событие
привело к появлению группы инфицированных
Для вызывающих эпидемии вирусов устанав-
активных распространителей болельщиков «ти-
ливается оценка базового репродуктивного числа
фози» в Бергамо, что спровоцировало Λ-образ-
R0. Это число предполагает, сколько один носи-
ный пик эпидемии с оцененной на данный мо-
тель в среднем заражает других. Для вируса SARS-
мент смертностью 12%. Именно смертность явля-
CoV-2 определяли R0 = 2.0 по результатам мате-
ется ключевой характеристикой при сравнении
матического моделирования [20] и R0 = 2.5 [15] по
разных эпидемий, а не только скорость роста за-
статистике из Уханя (есть основания считать, что
болевших.
данные занижались), но это значение R0 скорее
Сравнивать можно то, что сопоставимо -
является статистическим показателем для спра-
страны, близкие по уровню доступности медици-
вочников. Эта характеристика не объясняет стре-
ны и ведущие непредвзятую статистику. Столь
мительных локальных вспышек. В Италии базо-
драматичную разницу в показателях смертности
БИОФИЗИКА том 66
№ 2
2021
390
ПЕPЕВАPЮXА
стран (например, соседние Испания - 10%, Пор-
вания - и на две части Spike-белка, и на альтерна-
тугалия - 4%) объяснить чрезвычайно сложно
тивные N-протеины с анализом их частотного
уровнем качества медицины. Объяснение может
распределения. Косвенным подтверждением ре-
базироваться в рамках гипотезы «иммунологиче-
альности гипотезы будет расхождение - значи-
ского импринтинга». Если вы первым переболели
мая доля отрицательных (стандартизованных)
в детстве вирусом гриппа типа HxNx, то стали на-
тестов на антитела для подтвержденных ПЦР-те-
всегда устойчивы именно ко всем схожим с дан-
стами случаев COVID-19 c сильными
ным типом штаммам, но перед HxNу окажитесь
симптомами и положительных тестов на антитела
беззащитны и будете болеть каждый год, так как
при нескольких отрицательных ПЦР-тестах. Воз-
иммунная система не заметит различий - активи-
ражение против объяснения на базе идеи «перво-
зируются В-клетки памяти, хранящие старый ан-
родного антигенного греха» - в условиях глоба-
тиген. Из истории вакцинации помним, что ко-
лизации в Европе циркулирующие типы вирусов
ровья оспа обеспечивала стойкость к натураль-
должны были равномерно перемешаться. Генети-
ной, тогда их неразличимость играла на нашей
ки уже ищут объяснения различий смертности от
стороне. В работе [22] в серии опытов на мышах
COVID-19 в частоте вариантов участков генов ги-
был подтвержден эффект «слепого пятна» (blind
стосовместимости, отвечающих за иммунитет.
spot) иммунной системы - предел различимости
Методов моделирования распространения бо-
вариантов вирусных белков. В результате зара-
лезней известно несколько, как мы видим, не все
женные новым штаммом гриппа продуцируют
они оказываются пригодны для прогнозирования
антитела против старых вариантов белков вирус-
пандемии и, главное, методов оценки будущих
ных штаммов. Плохо связывающиеся с мишенью
показателей смертности. Популярны модифика-
антитела только мешают. В гипотезе предполага-
ции SIRS-модели распространения, исследован-
лись типы гриппа нейраминидаза/гемагглюти-
ные как в форме систем дифференциальных
нин, но коронавирусы человеческой популяции
уравнений, так и в дискретной алгоритмической
просто менее изучены, вакцины от них не тести-
форме. Алгоритмы применялись как полностью
ровали, тут может вмешиваться фактор неопреде-
детерминированные [22], так и с учетом вероят-
ленности, увеличивая дифференцировку сцена-
ностных характеристик исхода - заражения, вы-
риев болезни. Нет гарантии, что максимум часто-
здоровления, смерти [23]. Существуют камераль-
ты из многообразия возможных антител всегда
ные SIRS-модели, где выделяют субпопуляции и
придется на нужный нам белок. Несколько бета-
различные группы риска при распространении
коронавирусов давно присутствуют среди наших
особого рода инфекций. В работе [24] рассмотре-
сезонных острых респираторных вирусных ин-
ны аспекты применения терапии и выделены от-
фекций, но раньше распространенность типов
дельно инфицирование «чувствительным вари-
коронавирусов в регионах пристально не анали-
антом» и заражение «устойчивым вариантом». В
зировали. Из данной гипотезы следует, что люди,
автоматных SIRS-моделях «заболевший» с неко-
которые не сталкивались с коронавирусами,
торой вероятностью становился «мертвым» или
справляются с COVID-19 легче, чем уже заражав-
«иммунным», но пока непонятно, как включить
шиеся другими представителями из семейства
формат переходов в состояние «бессимптомных
Orthocoronavirinae. Люди старшего возраста могли
носителей». В Италии сообщают, что молодые
переболеть близким по антигенным признакам
люди со слабыми симптомами остаются заразны-
штаммом в 80-е гг. прошлого века, и сейчас одно
ми носителями (у них подтверждают наличие
из звеньев их иммунитета генерирует малоэффек-
РНК-вируса) в течение двух месяцев, но для мо-
тивный ответ. Формирование иммунного ответа
дели нужны данные динамики иммунного ответа.
при презентации антигенов получает некоторую
В настоящее время нет единых критериев, по
вероятностную составляющую, неравномерное
которым правительства стран должны вводить
распределение по частоте. Трансмембранные
ограничительные меры для остановки зараже-
Spike-белки у видов РНК-вирусов значительно
ний. Вводить ограничительные меры эффектив-
отличаются. Участки торчащих шипов априори
но всем странам вместе и сразу. Всемирная орга-
считаются иммунодоминантными, но консерва-
низация здравоохранения 11 марта 2020 г. объ-
тивные нуклеопротеины, массивные белки, упа-
явила о пандемии, но уже постфактум. Прогнозы
ковывающие РНК в вирион, тоже иммуногенны
и математические модели сценариев распростра-
(допустимые мишени для Т-клеток). Клетки им-
нения новой коронавирусной инфекции у руко-
мунной памяти, хранящие антигены нуклеопро-
водства организации имелись, но они не сыграли
теинов, могут стать помогающим фактором при
особой роли в принимаемых решениях. Наличие
столкновении с вирусом, использующим моди-
многих математических моделей [25] не помогает
фицированные участки «шипов короны».
при отсутствии воли у чиновников Всемирной
Для проверки гипотезы «слепого пятна» в им-
организации здравоохранения в принятии реше-
мунной памяти интересно сравнивать варианты
ний и обязательных для всех алгоритмах дей-
антител при легком и летальном развитии заболе-
ствий. О возможной эпидемии при передаче че-
БИОФИЗИКА том 66
№ 2
2021
НЕПРЕРЫВНАЯ МОДЕЛЬ ТРЕХ СЦЕНАРИЕВ
391
ловеку одного из SARS-вирусов, которые цирку-
ФОРМАЛИЗАЦИЯ ЗАПАЗДЫВАЮЩЕЙ
лирует в популяции рукокрылых, говорилось еще
РЕГУЛЯЦИИ В ПОПУЛЯЦИОННЫХ
в 2007 г. (см. работы [26, 27]).
ПРОЦЕССАХ
Важный вопрос для прогнозов - когда опти-
Феноменологические модели могут быть акту-
мально выходить из ограничительных мер, не
альны, когда истинные механизмы нам пока не-
провоцируя повторный всплеск, или понять, что
известны и непосредственно их описать нет воз-
повторного увеличения заболеваемости в данном
можности, либо их сложность не подлежит фор-
сценарии никак не избежать. Опыт былых эпиде-
мализации. Математическому моделированию
мий показывает, что часто бывают повторные пи-
функционирования подсистем иммунитета по-
ки через два-четыре месяца после затухания пер-
священо значительное число работ [28]. В по-
вой волны инфекции, как это было и с «испан-
строении моделей долго придерживались разде-
кой», и с чумой в Марселе в 1720 г. Прогноз
ления на гуморальный и клеточный виды иммун-
возвратной волны эпидемии - актуальная задача
ного ответа. В работах группы Г.И. Марчука даже
моделирования именно сейчас. Важно отличать
был рассмотрен аспект старения иммунной си-
фазу осцилляции при колебательном развитии
стемы из-за повышенной «антигенной нагрузки»
эпидемического процесса и повторную вспышку
[29]. Однако до результатов последних лет струк-
как события. Критически важно для моделирова-
тура врожденного и сложность запуска адаптив-
ния и построения прогнозов знать ключевые ас-
ного иммунитета не была ясна. Исследования
пекты: как формируется постоянный иммунитет,
подтипов клеток Т-хелперов и алгоритма выбора
сколько он действует, возможна ли «реактива-
рецепторов далеко не закончены. Открытия
ция» вируса при сохранении скрытого очага в ор-
Р. Меджитова и Ч. Джейнуэя [30] в трактовке
ганизме. Появляются сообщения, что иногда ви-
биологической кибернетики означают, что систе-
рус исчезает, но его вновь находят в организме,
ма регуляции иммунных ответов работает триг-
это тревожно и неясно, насколько такие случаи
герно, т.е. каскадом сигналов, и относится к па-
редки.
радигме дискретно-непрерывных и гибридных
типов динамических процессов [31].
Недостатком многих SIRS-моделей, по наше-
Существует проблема моделирования непо-
му мнению, является отсутствие зависимости ка-
средственного взаимодействия вредителя и леса
чественных режимов поведения от начального
или вируса и организма как несопоставимость
состояния в момент запуска локального эпиде-
временных шкал этих борющихся видов (в част-
мического процесса. Различие в исходном коли-
ности, вирион ВИЧ живет до полусуток). Если
честве зараженных (по одной из гипотез) может
классифицировать феноменологические модели
являться объяснением столь разных эпидемиче-
биологии, то в случае иммунитета и вируса мы
ских сценариев в разных регионах, помимо дру-
описываем не взаимодействия типа «хищник-
гих факторов, основным из которых является
жертва» или «паразит-хозяин», эта коллизия
слабоформализуемая в уравнениях характеристи-
ближе к «модели войны» - противостояния двух
ка - плотность социальных контактов.
сражающихся армий. В моделях мы можем рас-
Для современного моделирования эпидемии
считывать эффективность применения средств
востребовано, чтобы показатель смертности был
поражения противников, не описывая внутрен-
не априорным параметром в системе уравнений,
нюю механику их оружия. Также актуально рас-
пусть заданным случайной величиной с извест-
сматривать в моделях именно сценарии заверше-
ным законом распределения по возрастным груп-
ния экстремального взаимодействия.
пам, но функционалом. Модель станет прогно-
Популярным способом построения биологи-
стической, если вероятную смертность оценить в
ческих моделей со сложной динамикой является
вычислительных экспериментах из демографиче-
использование уравнений с запаздыванием [32]
ских и иммунологических показателей субпопу-
(синоним «эредитарности» у Вольтерра). В таком
ляций.
подходе регуляция текущего процесса отбрасыва-
ется на некоторое время в прошлое состояние си-
Сегодня вычислительные мощности позволя-
стемы [33]. В биологии запаздывание отражает
ют строить детализированные сетевые модели пе-
целый комплекс действующих временных факто-
редачи вируса в гетерогенном социуме, но свой-
ров [34] и не ограничивается временем достиже-
ства вируса по скрытому распространению не
ния зрелости. Внутриклеточные процессы, на-
позволяют в данный момент ориентироваться на
пример, при экспрессии белков с использовани-
медицинскую статистику. Традиционные методы
ем матричной РНК включают несколько
моделирования, безусловно, полезны для рас-
временных характеристик [35].
смотрения локальных вариантов смены фаз про-
цесса с изменением доли выздоравливающих в
Факторы запаздывания реакций важны для
зависимости от динамики числа заражений и сце-
ситуации выработки иммунного ответа. Они ока-
нариев течения инфекции в организме.
зываются зависимыми на узком промежутке вре-
БИОФИЗИКА том 66
№ 2
2021
392
ПЕPЕВАPЮXА
мени от частоты случившихся встреч активных
часть единого затухающего колебательного про-
клеток с вирусом. В организме есть процедуры
цесса. В работе [42] мы указывали отличия для
немедленного реагирования на вторжение и це-
двух ситуаций - повторной вспышки вредителей
ленаправленного, ориентированного на эту ми-
эвкалиптового леса как отдельного стохастиче-
шень, но опаздывающего, так как дендритные
ского события и реализации продолжающихся
клетки сперва должны выставить антигены виру-
колебаний листовертки в Канаде как целостного
са (по аналогии, как представляет фоторобот по-
длительного популяционного явления.
лиция).
Для уравнений
«логистических» моделей
В. Вольтерра первым в математической биоло-
Ферхюльста, Гомпертца, Ричардса и др. выпол-
гии рассмотрел уравнения с N(t - τ). Самые
няется условие ∀t, N(0) < K, N(t) ≤ K, что не соот-
известные биологические модели с запаздывани-
ветствует быстрым инвазивным сценариям. Сце-
ем - это модели Хатчинсона, Николсона и Го-
нарий при ∃tσ < tm, N(tσ) > K и N(tm) < K соответ-
полсами, имеющие ряд модификаций [36]. Урав-
ствует хрестоматийной σ-образной кривой,
нение Хатчинсона не было сформулировано са-
показанной в книге «Экология» Ю. Одума для ти-
мим Хатчинсоном в 1948 г., как часто можно
пичного сценария динамики роста инвазионной
прочитать (старые работы оцифрованы и доступ-
популяции с «овершутом» N(tσ) > K, но без фазы
ны, теперь можно увидеть труды классиков, а не
просто ссылаться на них по привычке). В работе
экстремальной вспышки. Популяция вселенца не
[37] он кратко изложил гипотезу влияния про-
может моментально остановить прирост числен-
шлых состояний на эффективность воспроизвод-
ности из-за достижения K-ниши (она о нем не
ства, что совсем не являлось основной идеей той
подозревает), потому происходит превышение
статьи. В работе [38] было указано, что модель
N(tσ) > K до момента остановки прироста N(tσ) =
предложена Райтом, но в работе [39] видим уже
0 и невынужденного снижения до приемлемого
несколько иную запись:
равновесия с биотическим окружением. Это пре-
вышение не относится к экстремальным переход-
y′(t) = -αy(t - 1)[1 + y(t)], α > 0.
ным режимам. Рост численности останавливает-
По-видимому, Р. Мэй [40] первым выписал
ся сам по себе (N(t∞) = K), как регулируемый без
уравнение в таком привычном виде:
внешнего противодействия. Динамика, представ-
ленная уравнением (1), может описывать сцена-
dN
N(t
−τ)
=
rN t)
1−
(1)
рий только «мирного», не разрушающего среду
(
)
dt
K
заполнения экологической ниши с конкурент-
Уравнение (1) фактически является усложне-
ным вытеснением и без резкого снижения чис-
нием «логистической» модели Ферхюльста с вве-
ленности после «овершута» - превышения апри-
дением запаздывания τ для регуляции, которая
орной емкости ниши. Известны случаи полной
определена значением K, важнейшим понятием
гибели чрезмерно агрессивной чужеродной попу-
емкости «экологической ниши», теоретически
ляции. Для многих актуальных ситуаций дости-
обоснованной именно экологом Г. Хатчинсоном.
жение точки нулевого прироста численности со-
K - больше чем константа в модели, это принцип
всем не означает баланса со средой. Например,
внутривидовой регуляции теоретической эколо-
K - предельное число доступных для заражения
гии [41], исходящий из существования априори
вирусом клеток, что определяется в организме
предельно допустимой равновесной величины
эффективностью действия интерферонов.
популяции, существующей ∀t. Из математиче-
ской модели возникли популярные теории об «r-
Решение уравнения (1) диссипативно: N(t) ≤
и K-стратегиях» эволюции видов, хотя у Ферх-
≤ (-1) + exp(rτ) независимо от N(0) [43]. Свой-
юльста это было умозрительным математическим
ства уравнения (1) с образованием орбитально
описанием для социума Европы середины XIX
устойчивого цикла и негармоническими колеба-
века и никогда не предполагалось для экстре-
ниями хорошо известны [45] как образование ав-
мальных ситуаций при инвазиях новых видов.
товолн в системах с запаздыванием [44]. Траекто-
рия уравнения (1) переходит в циклический ре-
Уравнение при небольших запаздываниях τ
жим для популяции с репродуктивным
феноменологически описывает реально наблю-
потенциалом r после бифуркации Андронова-
дающийся режим затухающих популяционных
Хопфа при значении произведения бифуркаци-
колебаний N(t) → K. Траектория (1) при rτ ≈ π/2 ± ε,
онных параметров rτ > π/2 [46]. В биологических
N(0) < K опишет характер волнообразной эпиде-
задачах уравнение (1) анализируют с константной
мии - например, повтор фазы роста числа случа-
предысторией ∀t ∈ [-τ,0], N(t) = N(0). В матема-
ев COVID в Техасе (рис. 4а), но не в Нью-Йорке и
тических работах рассматривается однопарамет-
не в Италии. В Иране без усреднения на графике
= λ[1
наблюдается (рис. 4б) явный осцилляционный
рическая форма записи:
-
N(t
−
1)],
пото-
сценарий эпидемии, который многие назовут
му эти результаты трудно перевести обратно в
«вторая волна», но математически эта «волна»
терминологию экосистемных процессов. Обоб-
БИОФИЗИКА том 66
№ 2
2021
НЕПРЕРЫВНАЯ МОДЕЛЬ ТРЕХ СЦЕНАРИЕВ
393
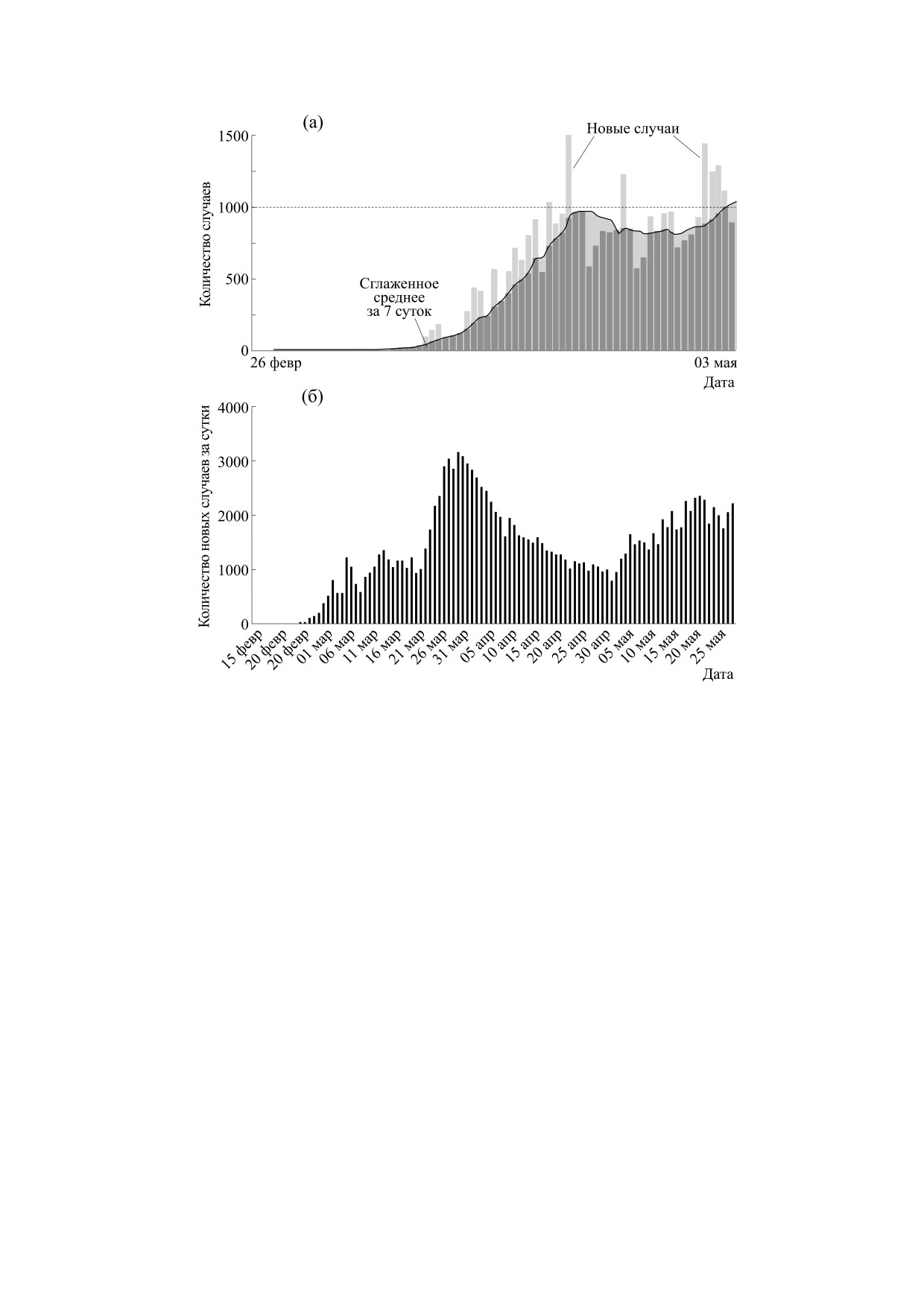
Рис. 4. (a) - Динамика новых случаев COVID в Техасе со сглаживанием - усреднением по данным за семь суток
(данные New York Times); (б) - осциллирующая эпидемии COVID в Иране.
щение уравнения Хатчинсона для учета возраст-
equation) не использует в явном виде значение
ной структуры популяции можно записать так:
предельной емкости ниши:
-τ2
∂
1
dN = rN
(t
−τ)exp(
−bN
(t
−τ))
−δN t).
(3)
N =r
1−
dv s)N(t
+
s)
N
(2)
(
)
dt
∂t
K -τ
1
В работе [48] модель в форме дифференциаль-
Недостатки уравнения Хатчинсона и его обоб-
ного уравнения предложена для описания коле-
щений хорошо известны специалистам в области
баний с большой амплитудой у лабораторной по-
практической экологии [47]. При увеличении ам-
пуляции мух в экспериментах Николсона. Урав-
плитуды минимумы релаксационного цикла (1)
нение
(3)
использует для формализации
становятся длительными и глубокими, фактиче-
внутривидовой конкуренции при повышении
ски попадают в ε-окрестность нуля. Для интер-
плотности известную функцию Рикера, только с
претации свойств изолированной популяции ре-
запаздыванием при репродуктивном сомножите-
жим нереалистичен. Обоснованно сказать, что
−bx
n
−1
ле, аналог уравнения
x
=
ax
e
−δx
. Итера-
n+1
n
n
модель теряет возможности сущностной биоло-
ция функции Рикера, давно применяемая в про-
гической интерпретации поведения при больших
мысловой ихтиологии модель пополнения попу-
значениях τ и не может описать резкие стреми-
ляции рыб с изъятием, имеет хаотический режим
тельные изменения.
по сценарию удвоения периода Фейгенбаума.
Альтернативное известное уравнение осцил-
Циклы итераций Ψn(x) = Ψ(n + p)(x), p = 2φ не под-
лирующей динамики «мясных мух» (blowflies ходят описания популяционных флуктуаций.
БИОФИЗИКА том 66
№ 2
2021
394
ПЕPЕВАPЮXА
Удвоение периода p рождает пары точек вокруг
гораздо сильнее первого. Подобный сценарий
давно известен в динамике активности насеко-
неустойчивой точки x*, но популяционные цик-
мых-вредителей как «sawtooth-like oscillation» -
лы - это последовательные перестановки с мак-
пилообразная вспышка численности с разнове-
*
*
симумом в конце периода
x
<
x
,i
=
0..p
−1
p- -1
p−i
ликими затухающими пиками. В совершенно
Анализ уравнения (3) относится к области хаоти-
разных биологических процессах может наблю-
ческой динамики [49] и выходит за рамки статьи.
даться идентичная динамика смены фаз, что объ-
Мальтузианский коэффициент r уже отражает
ясняется нами общностью математической фор-
разность между естественной рождаемостью и
мы их функций регуляции [53].
смертностью, тогда δ в уравнении (3) соответ-
Модельный сценарий актуален в предположе-
ствует дополнительной убыли как внешнему фак-
нии, что эпидемия полностью не затухнет, но по-
тору.
теряет «пандемический» статус и станет обычным
Максимумы колебательного режима после би-
среди других спорадически активных острых ре-
фуркации рождения цикла в модели Гополсами
спираторных вирусных инфекций. Вредители по-
[50] более соответствуют представлениям о регу-
сле окончания вспышки не исчезают, но их ста-
лярных колебаниях:
новится мало, и их активность не фиксируется в
лесных отчетах. В модели (5) вызвать новый пере-
dN
K -N(t
−τ)
=
rN t)
(4)
ходный режим с колебаниями способно резкое
dt
K
+
jN(t
-τ)
изменение значения N(t) - без бифуркаций.
Экологическим моделям с отклоняющимся
Сравнение подтверждает, что затухание эпи-
аргументом посвящено множество работ, обзор
демии после пика - процесс немонотонный и до-
многообразия модификаций моделей c запазды-
статочно волатильный, со стохастической компо-
ванием приведен в работах [51, 52].
нентой и локальными всплесками. Гладкое сни-
жение - признак умышленной фальсификации
Далее предложим модификации модели с точ-
статистики для отчета. При сознательном иска-
ки зрения описания редких и специфичных био-
жении статистики уменьшается дисперсия ряда.
логических процессов, которые подразумевают
В реальной эпидемии, как видим, обычное явле-
вариативность сценариев. Эпидемии, к нашему
ние - это разброс числа ежедневных случаев зара-
счастью, не формируют устойчивые циклы. Ко-
жения при колебаниях >25%.
лебания в популяционных процессах чаще спора-
дические, без постоянного периода - перемежа-
Для дальнейших модификаций целесообразно
ющиеся флуктуации. Описать сценарий спонтан-
принять гипотезу, что экологическая ниша пред-
ного завершения быстрой фазы инвазии важно в
ставляется относительным регулирующим фак-
том случае, когда достижение равновесия
тором, а не фиксированным - незыблемым апри-
limt→∞ N(t) = K является нереальным сценарием
орным значением. Переход к вспышке в реаль-
ных популяциях насекомых не может начинаться
для среды. Циклическая динамика - не главное
от единичных особей. Нужно учитывать, что на
ключевое свойство для эпидемических моделей.
графиках динамики вспышек численности при-
водятся данные о поражениях леса, а не о числен-
МОДИФИКАЦИЯ МОДЕЛИ
ности самих вредителей. В малых группах эффек-
ДЛЯ ЦИКЛИЧЕСКОЙ АКТИВНОСТИ
тивность размножения низка и зависит от слу-
ПОПУЛЯЦИЙ
чайных событий. Для запуска вспышки нужна
достаточно плотная группа особей, которая при
Для получения реалистичных с точки зрения
эффективном размножении способна вызвать
экодинамики форм осцилляций нами предложе-
вспышку. Мы предложили простую модифика-
но следующее уравнение с запаздыванием на ос-
цию уравнения (4) с неравномерным увеличени-
нове уравнения (3), но с формализацией нели-
ем нелинейности в числителе и знаменателе при
нейного порогового противодействия, которое
регуляции воспроизводства и относительностью
имеет режим спонтанного затухания осцилля-
ниши:
ций, возникших после бифуркации Андронова-
Хопфа:
2
dN
С -N
(t
−τ)
=
rN t)
,С <K
(6)
3
dN
N(t
− τ
dt
K +γN
(t
−τ)
=
rN(t
−τ)exp(−bN(t
−τ))
−
q
(5)
dt
A
-
N
(t
-τ)
Таким образом, с помощью уравнения (6)
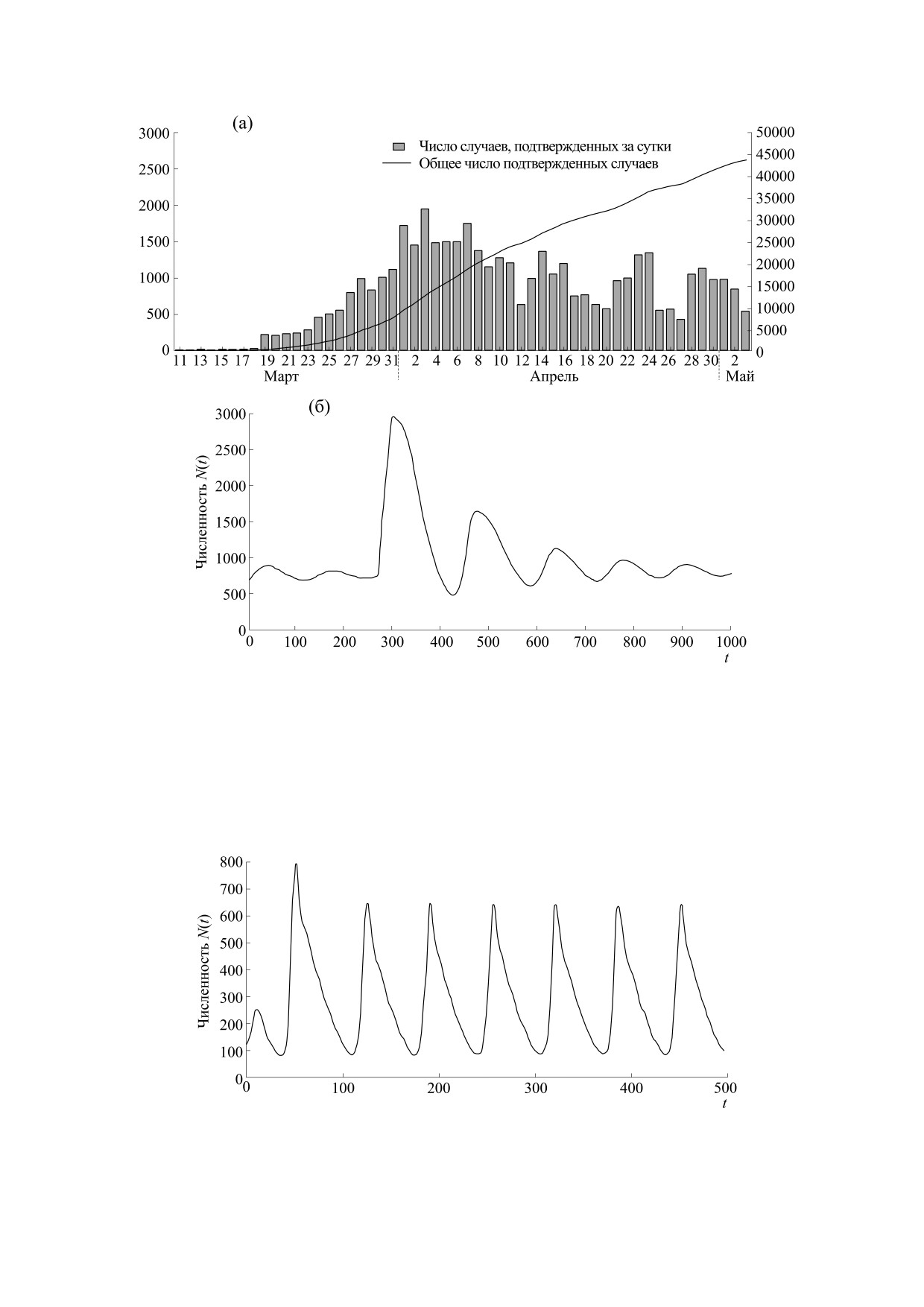
Модель (5) может описывать специфическую
можно получить режим пилообразных колебаний
динамику эпидемии COVID в штате Мичиган в
(рис. 6), где серии пиков численности начинают-
форме серии затухающих пиков после основной и
ся после бифуркации Андронова-Хопфа от
резкой вспышки (рис. 5). В штате Вайоминг эпи-
окрестности существовавшего до бифуркации
демия коронавируса аналогично развивалась
стационарного состояния, когда первый тре-
пульсирующими пиками, но второй пик там был
угольный пик переходного режима имеет боль-
БИОФИЗИКА том 66
№ 2
2021
НЕПРЕРЫВНАЯ МОДЕЛЬ ТРЕХ СЦЕНАРИЕВ
395
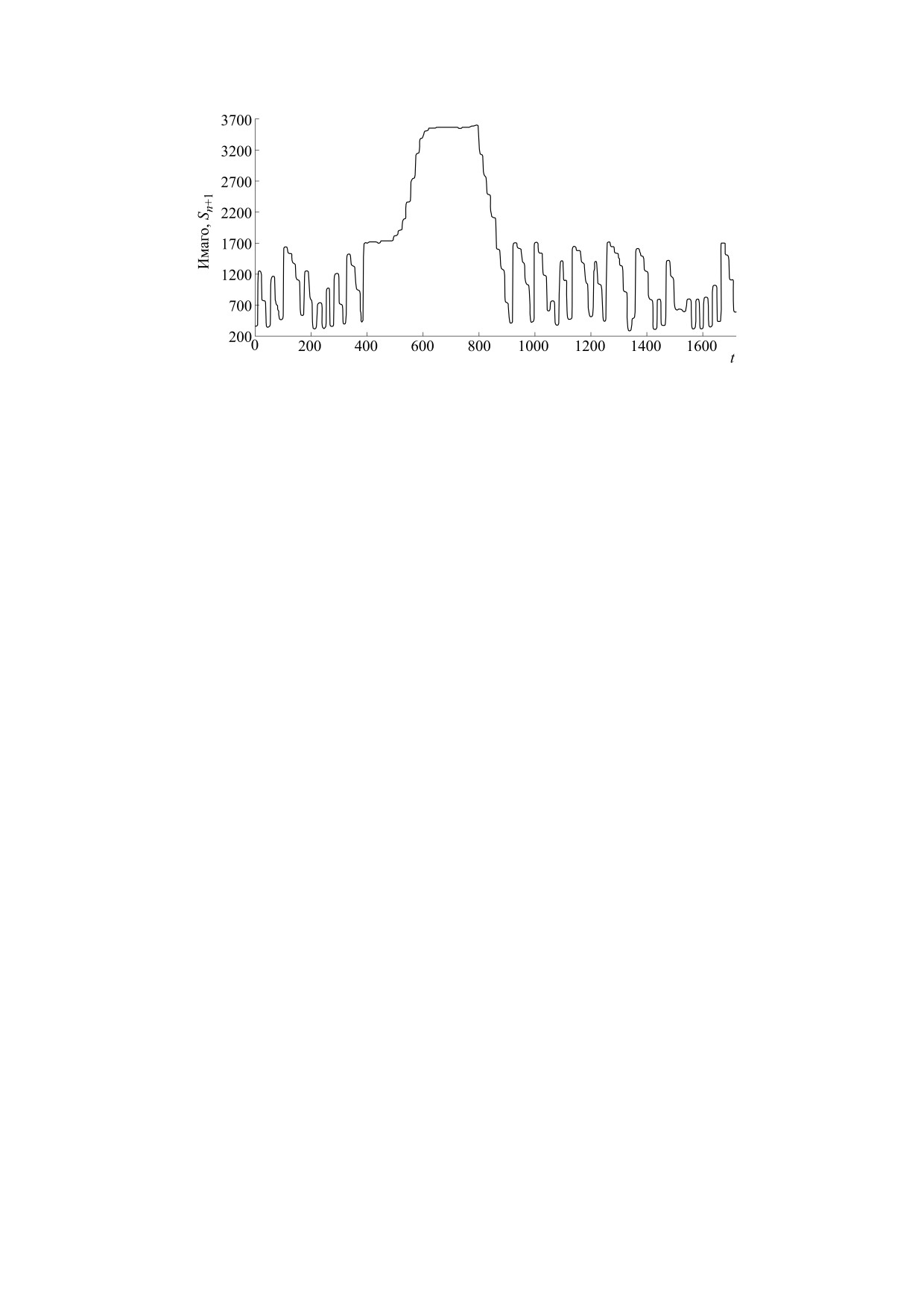
Рис. 5. (a) - Серия уменьшающихся пиков эпидемии в штате Мичиган; (б) - спонтанно затухающие осцилляции в
нашей модели (5) после бифуркации из-за увеличения параметра r.
шую амплитуду, чем последующие. В случае урав-
низмы вызывает потерю устойчивости у суще-
нения (6) для запуска пульсирующей вспышки
ствовавшего ранее в среде равновесия.
требуется первичный индуцирующий фактор.
Сопротивление среды вторжению агрессивно-
Некоторое воздействие на регулирующие меха- го вида можно учесть с помощью дополнительно-
Рис. 6. Λ-образные пики пульсирующей вспышки в модели (6) без изменения параметров.
БИОФИЗИКА том 66
№ 2
2021
396
ПЕPЕВАPЮXА
Рис. 7. Динамика поражения двух регионов лесов Аляски после начала осциллируюшей вспышки елового лубоеда
Dendroctonus rufipennis (из отчета службы лесного хозяйства [54]).
го нелинейного члена Ψ с запаздыванием в пра-
насекомыми, потому для подтверждения адекват-
вой части уравнения (6):
ности при моделировании экстремальной актив-
ности в биологических процессах лучше ориен-
2
dN
С -N
(t
−τ)
k
тироваться на данные из Канады. Динамика, пред-
=
rN t)
−Ψ[N
(t
−
h)].
(7)
3
dt
K +γN
(t
−τ)
ставленная на рис. 7, подтверждает гипотезу -
популяционные колебания насекомых вредите-
С помощью уравнения (7) можно получить се-
лей лесов в природе, в отличие от устойчивых
рию разновеликих пиков и колебания с нестаци-
циклов «вольтерровских» систем уравнений, -
онарным периодом. Специфический режим со-
это нестабильные переходные режимы и спора-
ответствует сценарию запуску пульсирующей
дические явления в экосистемах.
вспышки насекомых с Λ-образными перемежаю-
На рис. 8 показана иная регулярно осциллиру-
щимися пиками, который внезапно начинается
ющая активность комаров вида Aedes aegypti, яв-
после достижения значительной средней числен-
ляющихся переносчиками вируса лихорадки
ности. На рис. 7 показана динамика поражения
Денге в Южной Азии. В работе [55] отмечено, что
площади лесов в двух регионах Аляски из-за ак-
порог, от которого начинается новый пик, повы-
тивности жука-вредителя Dendroctonus rufipennis,
шается из-за потепления.
которая наблюдается на протяжении десятилетия
в форме вспышки с серией Λ-пиков. В Канаде и
В модели (6) значима близость численности
на Аляске открыты многолетние достоверные
инвазивной популяции к некоторому предпоро-
сведения о состоянии лесного фонда за вековой
говому уровню, при котором давление популяции
период и размерах поражения лесов различными
вселенца (или вируса) на среду уже велико. Для
Рис. 8. Пики активности переносчиков лихорадки Денге на юге Китая (из работы [55]).
БИОФИЗИКА том 66
№ 2
2021
НЕПРЕРЫВНАЯ МОДЕЛЬ ТРЕХ СЦЕНАРИЕВ
397
каждой биосистемы (или организма) этот уро-
акций активации, ее среднее значение может ва-
вень различен. Наблюдаемые у некоторых насе-
рьировать в локальных субпопуляциях (не обяза-
комых вспышки «пилообразной» формы - появ-
тельно из-за генетических факторов).
ление колебаний с быстрой серией пиков (saw-
Уравнения Хатчинсона и Николсона не могут
tooth-like oscillations)
- относятся к типам
описать завершение вспышки или эпидемии, ко-
экстремальной динамики. Подобные сценарии
торая в реальности не способна быть устойчивым
выходят за рамки теории плотностной регуляции
циклом, продолжаться при t → ∞. Вспышки чис-
и стремления биосистем к равновесию. Относи-
ленности насекомых и эпидемии вирусов - это
тельное балансовое равновесие «емкость ниши»
ограниченные во времени переходные режимы,
становится нижним значением для вспышечной
имеющие фазы и развития и окончания. Наша
активности. Регулярные серии вспышек бабочек
модель (5) показывает возможный путь заверше-
Malacosoma disstria и Lymantria dispar обычно име-
ния, но не обладает вариабельностью. Все экстре-
ют тенденцию затухания. Нам важна общность
мальные процессы в биологии обязательно завер-
регулирующих функций у таких явлений.
шаются, пусть даже состоянием полной деграда-
ции среды и гибелью бурно размножавшегося
ИНТЕРПРЕТАЦИЯ АГРЕГИРОВАННОГО
вида. Возможны несколько сценариев заверше-
ЗАПАЗДЫВАНИЯ
ния, в частности, достижения минимального
уровня равновесия, скрытого очага инфекции, не
Параметр «запаздывания» не обусловлен ха-
оказывающего видимого воздействия на среду. В
рактеристиками биологического вида, длиной
предыдущей работе [1] о триггерной итерацион-
этапов его жизненного цикла. Бабочка листо-
ной модели вспышки численности псиллид мы
вертка Choristoneura fumiferana в Нью-Брансуке
отметили, что описание сценария запуска эруп-
генерирует длительные циклы активности [56] с
тивной фазы и моделирование завершения
десятилетними промежутками между катастро-
вспышки - это две отдельные задачи, и сложнее
фическими вспышками. Интервалы между пика-
именно самостоятельное снижение численности.
ми не соотносятся с длиной развития поколений
У вируса меньше требований к среде, потому в
насекомых. Запаздывание, которое включено в
случае инфекции триггерным действием будет
популяционные модели (1) и (2), - это именно
обладать включение механизмов его подавления.
феноменологическое описание изменений в ди-
Рассмотрим «безбифуркационную» модель для
намике численности [57]. Таким способом рас-
сценариев развития инфекции.
ширяются возможности для описания изменений
режимов поведения траектории, но при измене-
нии параметров. В известных уравнениях исклю-
МОДЕЛИРОВАНИЕ РАЗЛИЧИЙ
чен фактор изменения качественной динамики
СЦЕНАРИЕВ ИНФЕКЦИОННОГО
сценариев в зависимости от начальных условий
ПРОЦЕССА
[58]. Динамика инфекционных заболеваний не
обязана следовать единственному сценарию, так
Комплексом ответных действий организма,
как иммунитет у всех индивидуален, но есть ма-
направленных на подавление проникшей инфек-
ловероятные варианты и частные случаи.
ции, управляет сложнейшая система взаимодей-
Бифуркационным параметром в уравнении (6)
ствующих разнотипных клеток. Иммунный ответ -
мы считаем не репродуктивный потенциал r, а ве-
это кооперативная реакция. Каждому звену мо-
личину τ. По нашему представлению, запаздыва-
жет быть определена узкая роль или несколько,
ющая регуляция в модели - это адаптационная и
как у B-лимфоцитов [59]. Каждое звено в иммун-
агрегированная характеристика. Величину τ мы
ных реакциях требует определенных шагов в ал-
полагаем результатом взаимодействия вида и
горитме активации, требуется время для их вклю-
конкретной биотической среды, формирующая-
чения. Иммунитет включает два больших связ-
ся в данном ареале. В действующую величину за-
ных структурных комплекса - врожденный и
паздывания (5) входят онтогенетическая задерж-
адаптивный. Для задачи моделирования инфек-
ка этапов развития, скорость восстановления ре-
ционных сценариев главное их отличие - разное
сурсов, время выработки ответа со стороны
время активации. Необходимо захватить антиге-
паразитов и патогенов, миграция. Запаздывание
ны, провести презентацию антигенов для Т-кле-
при анализе моделей имеет смысл разделить на
ток «хелперов» (показать «особые приметы») и
репродуктивное «регуляционное» и «адаптацион-
начать размножать именно требуемые антиген-
ное», которое необходимо популяции (или орга-
специфичные клетки-киллеры, что не может
низму) на выработку приспособительных реак-
происходить мгновенно. Презентация антигена -
ций, данная величина может изменяться в про-
процесс, в результате которого зрелый Т-лимфо-
цессе инвазии. В организме, если речь идет о
цит мигрирует в ткани с целью распознать и убить
борьбе иммунитета и вируса, запаздывание - ин-
там зараженную клетку, экспрессирующую на
дивидуально зависимая характеристика всех ре-
своей мембране этот антиген. Сократить время
БИОФИЗИКА том 66
№ 2
2021
398
ПЕPЕВАPЮXА
получения ответа от адаптивного комплекса кле-
обобщенный график позволил бы свести задачу
ток - проблема.
на язык математического анализа с построением
функциональной зависимости начальной дозы
Репликация вируса, в отличие от смены поко-
заражения и реакции иммунитета. Потом ее нуж-
лений насекомых, процесс непрерывный, поэто-
но аналогично тормозить, так, антитела подклас-
му для задачи анализа сценариев оправдано ис-
са IgG4 снижают воспаление, подавляя актив-
пользовать дифференциальное уравнение с за-
ность фагоцитирующих клеток [64]. В гипотети-
паздыванием. Интересно моделировать смены
ческой зависимости дозы и скорости реакции
фаз течения заболевания исходя из внутренней
наиболее интересны пороговые эффекты. Дан-
логики противодействия инвазии в сценариях без
ные экспериментов показывают, что при разви-
индукции «извне» и бифуркаций из-за колебаний
тии вирусных заболеваний начальная доза спо-
внутренних параметров. Изменения параметров
собна определять качественные характеристики и
возможны в долгосрочной перспективе. Перехо-
темп иммунного ответа, поэтому должна рас-
ды между фазами при первичной инфекции нело-
сматриваться в модельных вычислительных экс-
гично описывать колебаниями характеристик и
периментах как влияющий фактор на реализа-
явными бифуркационными переменами.
цию сценария болезни.
Основная задача в модифицированной модели -
Изменения поведения уравнений (5) и (6) мы
рассмотреть сценарии спонтанного завершения
наблюдаем при бифуркациях, вызываемых мо-
экстремального процесса, перехода его в иное ка-
ментальными изменениями значений управляю-
чество после фазы быстрого роста численности
щих параметров, которые далеко не всегда можно
вирионов. В некоторых ситуациях считается важ-
объяснить сущностными причинами. В каждом
ным начальное число попавших в организм вири-
случае нужно теоретически обосновать механиз-
онов [60]. Зависимость течения заболевания от
мы увеличения значения r (или τ), которое прово-
начального N(0) может быть нелинейной [61]. Во
цирует пульсирующую вспышку. Все характери-
многих случаях необходимо принимать в расчет
стики видов в природе изменчивы, но обычно
дозозависимый иммунный ответ «dose-dependent
плавно.
response», что важно для эффективной вакцина-
ции [62]. После контакта с антигеном дендрит-
Предложим новую модель исходя из значимо-
ные клетки должны взаимодействовать с лимфо-
сти субкритического уровня присутствия патоге-
цитами. При малом N(0) вирус может ускользнуть
на H - предпороговой величины вирусной на-
от иммунных «сторожевых».
грузки N(0) < H < K. Вместо квадратичной или
В работе [63] проводились эксперименты, в
экспоненциальной выберем логарифмическую
которых две группы мышей заражали разными
функцию регуляции:
дозами гриппа, низкой и очень высокой. Разница
в 1000 раз интересна в опытах, в реальной жизни
dN
K
=
rN t)ln
(H -N(t
−
h)).
(8)
различие дозы в 10 раз уже велико. Было показа-
dt
N(t
-τ)
но, что начальная инфекционная доза регулирует
Величина H отражает в уравнении (8) конкрет-
раннюю реакцию и выброс иммуномедиаторов
ное состояние организма при взаимодействии
цитокинов. Оказалось, что чем больше начальная
иммунитета с данной инфекцией, тогда как K иг-
доза вируса, тем сильнее симптомы воспаления,
рает роль априорно заданной пороговой величи-
быстрее начинается индукция интерферонов и
ны, но не тождественной емкости ниши из логи-
неожиданно наблюдалась более ранняя активная
стической модели. Уровень предельного насыще-
инфильтрация Т-клеток с рецептором CD8 в лег-
ния в нашем понимании
- это состояние,
кие. «Поразительно, хотя титры вируса в легких
которое достижимо и может быть незначительное
были значительно выше у мышей, зараженных
превышено в кратком времени, но не сохраняемо
высокой дозой в первые дни заражения, относи-
в среде. Роль этой характеристики оказывается
тельная вирусная амплификация была значи-
двоякой: чем показатель меньше, тем слабее чув-
тельно выше (в 125 раз) на пике вирусной нагруз-
ствуем симптомы, но тем ближе критический по-
ки у мышей, зараженных низкой дозой вируса».
рог, переход через который необратим.
Начальная доза, согласно этим экспериментам,
влияла только на реакцию клеточного иммуните-
Рассмотрим два основных сценария новой мо-
та, генерация нейтрализующих антител в дина-
дели (8). Будем считать, что h может быть только
мике оставалась независимой. Потом экспери-
незначительно больше регуляционного запазды-
ментаторы снова заражали выживших животных
вания τ, так что h = τ + ξ, но τ полагаем значитель-
и установили, что перенесшие большую дозу
ной величиной, а H = K/3 фиксируем. В первом
мыши приобрели устойчивость за счет оставших-
сценарии фаза замедляющегося «логистическо-
ся CD8.
го» роста численности будет идентична модели
Хатчинсона при rτ ≈ π/2 c «овершутом» N(tm) > K,
К сожалению, авторы не использовали инте-
гральные показатели иммунного ответа, тогда
N(t) → K, но достигнутое равновесие после зату-
БИОФИЗИКА том 66
№ 2
2021
НЕПРЕРЫВНАЯ МОДЕЛЬ ТРЕХ СЦЕНАРИЕВ
399
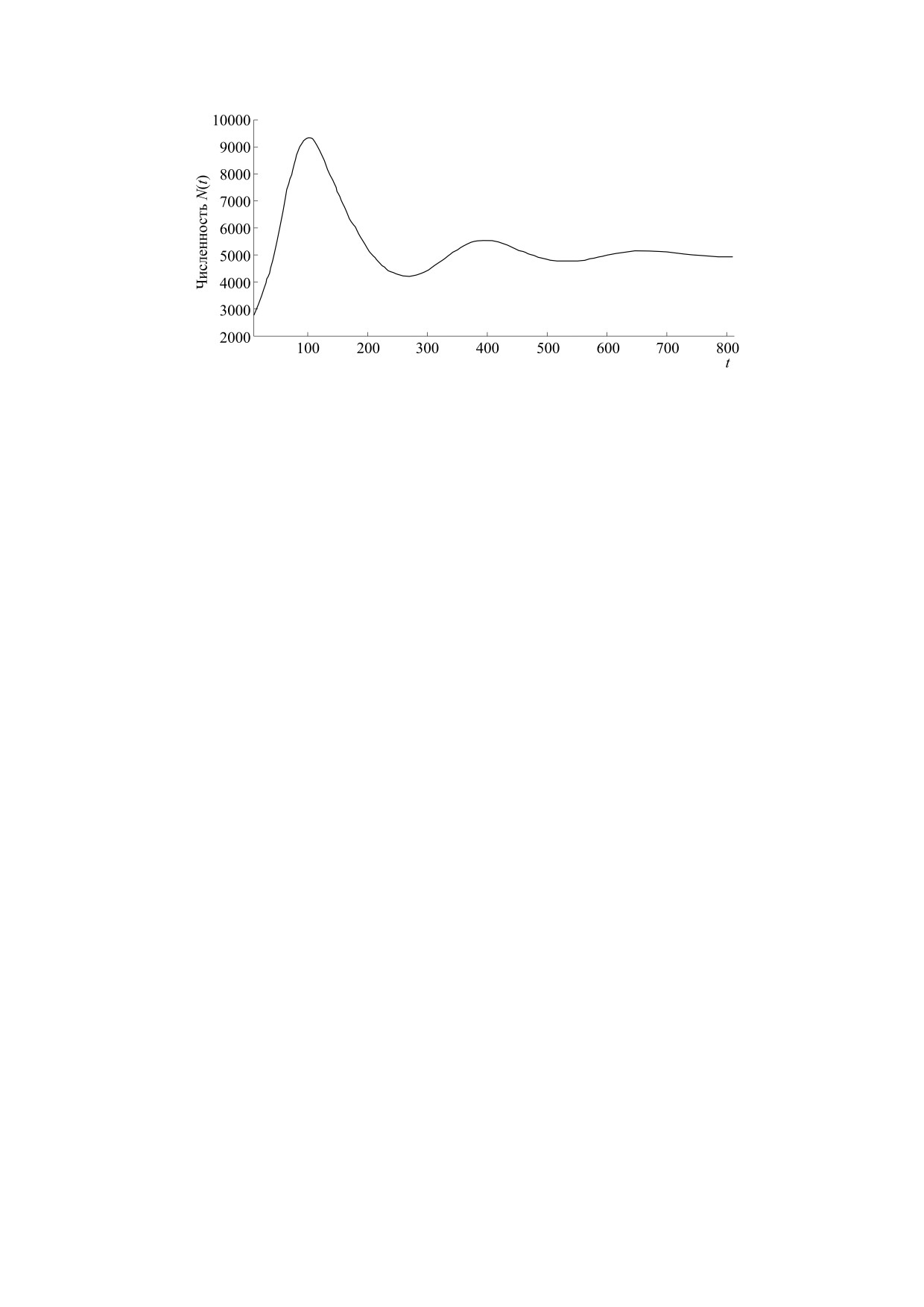
Рис. 9. Недиссипативная траектория решения модели (8) после состояния N(t) ≈ K в режим N(t) → ∞ при τ = 58,
K = 15000, H = 5000, r = 0.00000335, N(0) = 190, ξ = 2.
хающих осцилляций оказывается неустойчиво. В
роста N(t) → K происходит переход от кратковре-
этом сценарии нет конечного равновесия (рис. 9).
менных колебаний в окрестности K к флуктуаци-
ям при значительно меньшей численности в
Траектория t > tm резко покидает окрестность
окрестности H, которые медленно затухают
K, так что limt→∞ N(t) = ∞ и решение модели (8)
(limt→∞ N(t) = Н). Мы видим более стремитель-
при выбранном значении N(0),τ становится более
ную фазу роста, но меньшее время инфекции в
не диссипативным. Сценарий означает разруше-
окрестности K. В вычислительном эксперименте,
ние среды в том случае, когда имеющийся исход-
представленном на рис. 10, траектория приходит
ный иммунитет недостаточен для контроля инва-
в устойчивое равновесие после переходного ре-
зии. После фазы острой виремии и следующего
жима N(t) ≈ K при аналогичных значениях пара-
после max N(t) этапа tK > 2τ длительной псевдо-
метров модели (8). Мы не рассматриваем условие
стабилизации тяжелого заболевания в этом вари-
N(0) > H как недопустимое в реальности даже во
анте происходит безвозвратная деструкция сре-
время пандемии, заведомо летальные дозы вируса
ды. В вычислительном эксперименте расчет пе-
вкалывают только в экспериментах мышам.
реходного режима после выброса траектории
заканчивается сообщением численного метода с
Решение уравнения (8) имеет альтернативные
контролем точности об ошибке переполнения
сценарии асимптотического поведения без изме-
при остановке вычислений.
нения ключевых характеристик взаимодействия.
Если начальная численность при заражении
В контексте именно заражения организма виру-
станет ближе к H, то в модели (8) реализуется со-
сом значения r, τ, H - постоянные параметры.
вершенно иной сценарий. После сходной фазы
Важно значение начальной численности N(0) от-
Рис. 10. Равновесие траектории модели (8) после N(t) ≈ K в состояние N(t) → H, N(0) = 1099.
БИОФИЗИКА том 66
№ 2
2021
400
ПЕPЕВАPЮXА
Рис. 11. Сценарий бессимптомной хронизации инфекции c max N(t) < K при уменьшенном регуляционном
запаздывании в модели (8), N(0) = 1200.
носительно субкритичекой емкости среды H.
Полученный в модели (8) качественный ха-
Предполагается N(0) ≥ μ0 - инфекция не начина-
рактер динамики заболевания наиболее типичен
ется от околонулевого количества. Величины H и
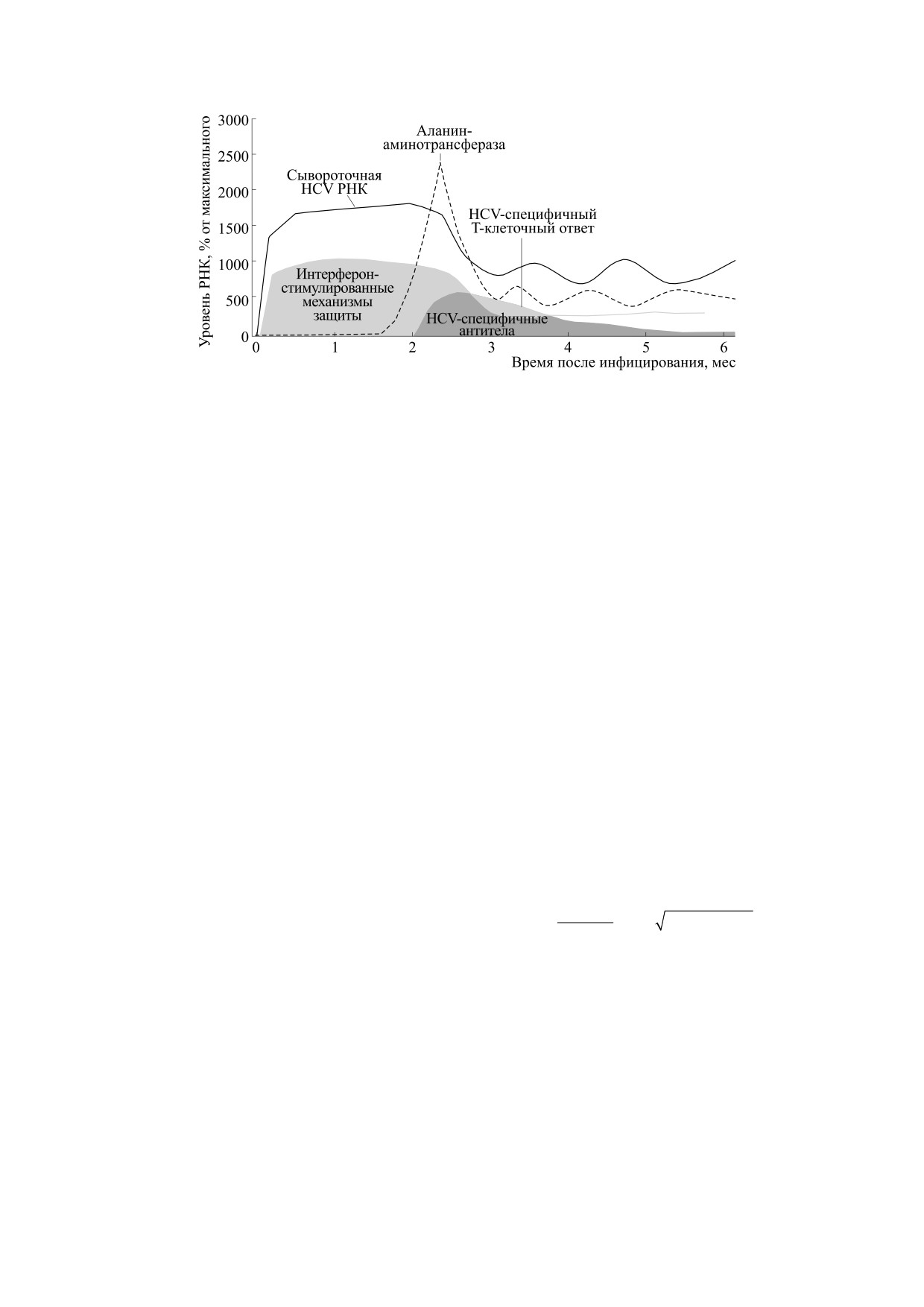
для заражения гепатитом С. На рис. 12 представ-
K мы трактуем как характеристики организма,
лено реально встречающееся перетекание острой
свойства его врожденного иммунитета. K - вари-
фазы инфекции вирусным гепатитом С в хрони-
ативная величина в рамках популяции, что объяс-
ческое течение [65].
няет различную тяжесть симптомов. Значение H
На графиках от начала инфекции показано как
консервативнее, но не сохраняется с возрастом и
резко после достижения вирусом предельной
является естественным изменчивым параметром
окрестности K появляется аланинаминотрансфе-
среды, влияющим на поведение модели. Сокра-
раза. Внутриклеточный фермент аланинамино-
щение величины запаздывания до 3/4 h в модели
трансфераза обнаруживается в крови в значи-
(8) приводит к сценарию с менее выраженной
тельной концентрации при разрушении клеток
острой фазой инфекции - мягкому течению за-
печени - тогда проявляются характерные острые
болевания (рис. 11). Непосредственное измене-
симптомы. Пик выброса аланинаминотрансфе-
ние параметров влечет более значительное изме-
разы происходит с запаздыванием по сравнению
нение положения max N(t) в переходном режиме.
с ростом концентрации РНК вируса гепатита С в
В переходной динамике симптомы восприни-
крови. Интерферон-стимулированные механиз-
маются краткими и незначительными из-за от-
мы защиты активируются быстрее [66], но они не
сутствия стадии N(t) ≈ K, но асимптотическое со-
могут подавить вирус и в сценарии выполняют
стояние процесса аналогично сценарию с силь-
роль демпфирующего регулятора. Появление эф-
ными симптомами. Биологическое объяснение
фективных лимфоцитов, способных снизить
сценария в том, что врожденный иммунитет и ан-
концентрацию вирионов вирусного гепатита С и
титела быстро остановили распространение ви-
следом уменьшать присутствие фермента алани-
руса, но не убили все зараженные клетки - созда-
наминотрансферазы, регистрируется со значи-
лось длительное равновесие. Можно сказать, что
тельным запаздыванием. Нейтрализующие спе-
тут иммунитет стал «близорук» и недоработал (без
цифические антитела в этом сценарии появляют-
действия Т-киллеров), но логичнее считать, что
ся еще позднее. Экспрессия белков от большой
его реакцию просто затормозили. Существует
группы чувствительных к интерферонам генов
петля отрицательной обратной связи у иммунных
интерферон-стимулированных механизмов за-
реакций, где начинают действовать клетки Т-су-
щиты (далеко не все гены изучены) тормозит рас-
прессоры, подавляющие активность других лим-
пространение вируса в клетках [67], что создает
фоцитов. Третий сценарий коварен, так как чело-
предельный K-уровень - уровень временного не-
век может долго не понимать, что он серьезно за-
устойчивого баланса, при котором размножение
болел. Одностороннее увеличение в модели (8)
инфекции остается гарантированно контролиру-
репродуктивной активности rm >> r логично вы-
емым. Превышение неустойчивого уровня K для
зовет быстрый режим N(t) → ∞ - как необрати-
количества вирусных частиц временно допусти-
мый летальный вариант развития процесса с со-
мо, как мы видим в сценарии хронизации с зату-
всем недолгим пребыванием около неустойчиво-
хающими осцилляциями, но время превышения
го равновесия.
не должно превосходить τ.
БИОФИЗИКА том 66
№ 2
2021
НЕПРЕРЫВНАЯ МОДЕЛЬ ТРЕХ СЦЕНАРИЕВ
401
Рис. 12. Динамика концентрации РНК вируса гепатита С и выработка разных типов иммунного ответа при переходе
острой инфекции в хронический сценарий (согласно работе [65]).
Противовирусные внутриклеточные белки ин-
Равновесное состояние не исключает возмож-
терферон-стимулированных механизмов защиты
ности повторных пиков инфекции. Многие ви-
и ZAP (англ. Zinc-finger Antiviral Protein) матема-
русные инфекции рецидивируют при долгой им-
тически переводят экспоненциальный рост в ло-
муносупрессии. Иммунный ответ на рис. 12 при
гистическую кривую. Эти регуляторы создают
уменьшении присутствия вируса гепатита С сни-
время на выполнение алгоритма ответа от клеточ-
жается, в плазме остаются обычно белки IgG. Для
ного иммунитета. Передовые средства защиты
возмущающего воздействия в модели (8) даже не
всегда работают с рождения и спасали бы, только
нужно менять ключевой параметр r, а резко ва-
рьировать H или τ, что вызовет новый переход-
при эволюции некоторые вирусы научились про-
ный режим с фазой роста инфекции.
тиводействовать синтезу интерферонов и избе-
гать антивирусного белка ZAP. Клетки врожден-
ного иммунного комплекса, не проходящие «ан-
ВАРИАНТЫ МОДИФИКАЦИИ
тигенную настройку», справляются со многими
ИНФЕКЦИОННЫХ СЦЕНАРИЕВ
вирусами плохо и с побочными последствиями.
С ТЕРАПИЕЙ
Своевременная генерация ответа в форме T-кле-
ток с рецептором CD8 имеет решающее значение
«Хронизация» в модели (8) - невынужденный
для остановки инфекции, но потому мы и болеем
переход траектории к стабильному равновесию
гриппом неделю, что максимум их концентрации
через окрестность неустойчивого. Актуален сце-
наблюдается в опытах через семь суток [68].
нарий окончательной гибели вторгшегося агрес-
сивного вида в разрушаемой им среде. Модель
Реакция противодействия в рассмотренной
эрадикации с N(t∞) = 0 можно предложить при
группе сценариев не способна полностью пода-
использовании идеи уравнения Базыкина
-
вить вирус, но может перевести инфекцию в со-
включить явно критическое значение численно-
стояние, когда оставшийся очаг вируса не угро-
сти для существования вида L, но в степени 1/3:
жает жизнеспособности организма, однако ста-
новится скрытой угрозой. Вирус Эбола таится в
dN
K
3
клетках Сертоли. Интервал времени от зараже-
=r
ln
N t)
N(t
−κ)
−
L, 0
< κ<τ
(9)
dt
N(t
-τ)
ния до начала действия специфических клеток
иммунитета - важный фактор сопротивляемости
В моделях с критическим нижним значением
организма, но в нашей модели дополнительно
численности L требуется N(0) > L, и в таком сце-
значим первичный уровень проникшей в орга-
нарии (9) получим полное уничтожение очага бо-
низм инфекции. За двумя параметрами запазды-
лезни, но не рассмотрим варианты стремитель-
вания в модели скрываются, во-первых, свойства
ной инвазии от исходно малой группы проник-
алгоритма репликации и сборки вирионов, во-
ших патогенных организмов [69]. Вирусам не
вторых, эффективность сложной группы медиа-
нужно искать партнеров для размножения, им до-
торов иммунного ответа - интерлейкиновая си-
статочно где-то скрыться от средств иммунитета.
стема взаимной стимуляции клеток. Запаздыва-
По оценкам, сделанным в работе [70], заражение
ние - атрибут системы «репликация-подавле-
натуральной оспой способен вызвать единичный
ние».
вирус. К счастью, обычные сезонные острые ре-
БИОФИЗИКА том 66
№ 2
2021
402
ПЕPЕВАPЮXА
спираторные вирусные инфекции и даже COVID
тивной составляющей иммунного ответа бывают
не так беспощадны.
значительными, но они могут реализоваться с
С точки зрения вычислительного моделиро-
критическим для организма опозданием, что
вания подсистемы иммунитета удобно класси-
приводит к реализации летального сценария. В
фицировать по времени начала их реакции на
отдельных случаях они становятся недостаточно
три группы. Третьим звеном будем считать ней-
специфичными - плохо взаимодействуют с под-
трализующие аффинные к постоянным антиге-
типом вируса. Обосновано включить в инфекци-
нам вируса иммуноглобулины, клетки-киллеры.
онную модель нелинейную составляющую
Для математической формализации не обяза-
тельно знать их физиологические механизмы, но
Ψ[Nk(t - h] активного противоборства специаль-
важно время триггерного включения и попра-
ными средствами терапии или например, цито-
вочные факторы, корректирующие их эффект
токсическими лимфоцитами, клетками с рецеп-
воздействия. Количественные показатели адап-
тором СD8 следующим образом:
dN
K
N(t
−
2h)
=
rN t)ln
(H -N(t
−
h))
−
q
(10)
dt
N(t
-τ)
H
1
-
N(t
-2τ)
Специальные клетки способны уничтожать
низации - это асимптотически устойчивое состо-
внутриклеточных паразитов, оставшихся там, ку-
яние, а тривиальное равновесие неустойчиво. Ес-
да антитела проникнуть не могут. Дополнением в
ли терапия начинает применяться в устойчивом
модели (10) можно получить сценарий очень мед-
положении системы, то она оказывается малоэф-
ленного асимптотического приближения N(t) → 0.
фективной и достичь окончательного успеха
В реальности активность лимфоцитов со време-
трудно. Терапию нужно начинать применять при
нем начинает аналогично ингибироваться сред-
переходном режиме и неустойчивом балансе, то-
ствами самого организма [71]. Интересно рас-
гда возможно быстрое достижение излечения,
смотреть модельный сценарий, когда длитель-
минуя хронизацию. Математическое решение
ность активности иммунных клеток-киллеров,
может выразиться в переводе процесса снова в
проходящих сложную процедуру отбора и актив-
острую фазу, это вызовет включение иммунного
ных к конкретной мишени антигену, зависит не-
ответа одновременно с началом терапии, но кли-
посредственно от начального состояния при за-
ническая практика данный способ не одобрит.
ражении. Для таких целей нужно формализовать
Возражение - положение K критического не-
составляющую Ψ[N(t - h, N(0)], что мы рассмот-
устойчивого баланса у истощенного борьбой ор-
рим в следующей работе, когда будут получены
ганизма при повторной острой фазе может ока-
данные о фазах протекания коронавирусной ин-
заться эфемерным; в нашем случае чем большего
фекции и динамике вырабатываемого организ-
промежуточного значения достигает N(t), тем
мом разных форм иммунного ответа. Есть все ос-
сильнее симптомы и риски.
нования полагать, что будет установлено большее
разнообразие вариантов развития заболевания и
соответственно описан ряд динамических сцена-
ЗАКЛЮЧЕНИЕ
риев противоборства с компонентами нашего им-
В статье мы сравнили три оригинальные моде-
мунитета. Подтверждаются наблюдения, что не-
ли (5), (6) и (8) для специфического развития про-
сбалансированный иммунный ответ, названный
цессов в биосистемах. В приведенных уравнениях
в работе [72] «дисрегуляция профиля экспрессии
экстремальная или пульсирующая динамика ин-
воспалительных цитокинов» (для группы, не
вазии способна начинаться (и завершаться) раз-
только интерлейкина-6), осложняет тяжесть по-
личным образом: 1) вследствие увеличения пока-
следствий у заболевания COVID.
зателя репродуктивной активности и бифурка-
Антивирусную терапию целесообразно моде-
ций; 2) из-за резких сдвигов численности самой
лировать другим способом, функционалом
популяции без изменений параметров; 3) в урав-
уменьшения репродуктивного потенциала r(t),
нении (8), актуальном для анализа фаз инфекции
так как известные препараты против вирусного
при запаздывании иммунного ответа, эти транс-
гепатита С и вируса иммунодефицита человека
формации свойств переходного режима происхо-
направлены на подавление репликации и инги-
дят в зависимости от начального количества вы-
бирование вирусной полимеразы, но без лизиса.
зывающего заболевание вируса и его новой сре-
Такой метод актуален, чтобы прогнозировать ди-
ды, которое в наших вычислительных сценариях
намику после отмены терапии.
различалось в пять раз. В модели (8) возможны и
Отметим важный практический вывод из мо-
бифуркационные сценарии изменений поведе-
дельных сценариев: равновесие в сценарии хро-
ния. Метаморфозы предельных множеств траек-
БИОФИЗИКА том 66
№ 2
2021
НЕПРЕРЫВНАЯ МОДЕЛЬ ТРЕХ СЦЕНАРИЕВ
403
тории для анализа быстрых фаз инвазий не столь
леть фазу острой инфекции. Важнейшая рассчи-
значимы. Отдельно оценены модели с явным не-
тываемая в модели (8) характеристика переходно-
линейным противодействием в форме дополни-
го процесса - тот интервал времени tK, который
тельного функционала воздействия - по вариан-
траектория проводит в окрестности предельного
ту (7).
уровня заражения. Включенные в модель (8) t - τ
и t - h мы полагаем агрегированными характери-
В новой модели (8) последовательно рассмот-
стиками.
рены альтернативные имитационные сценарии:
летального развития инфекционного процесса,
В предыдущей нашей итерационной модели
критически опасного течения острой инфекции
[1] псевдостабилизация происходила перед
с сохранением хронического очага и бессимп-
вспышкой насекомых у порогового состояния. В
томного сценария c max N(t) ≈ 2/3K. Два вариан-
новой модели пороговое состояние будет дли-
та имеют существенно различающийся переход-
тельно устойчивым, но равновесие при высокой
ный режим, но аналогичную асимптотику. В до-
численности K - репеллерное. Для опасного па-
статочно простом и наглядном уравнении нет
тогенного вируса нет продолжительной стабиль-
необходимости для каждого из трех сценариев
ной высокой численности, но возможно сохране-
настраивать репродуктивный r-коэффициент
ние хронического очага. Существуют различные
вируса или изменять другие параметры и вызы-
сценарии смены фаз течения многих инфекцион-
вать бифуркации. В вычислительных экспери-
ных заболеваний даже внутри субпопуляции, что
ментах на основе формализации в уравнении за-
ярко демонстрирует COVID-19. Свойства мо-
паздывающего взаимодействия мы варьировали
дельных сценариев объясняют наблюдения ита-
начальные условия при попадании инфекции по
льянских медиков об одинаковости средней ви-
отношению к субкритической для данного виру-
русной нагрузки. Математическое объяснение
са концентрации, специфичной для текущего со-
состоит в том, что имеется разный уровень пре-
стояния инфицированного организма. Рассмот-
дельного критического насыщения, но предпоро-
ренные три сценария не означают, что смены ре-
говый асимптотический уровень одинаков и со-
жимов поведения модели
(8) не способны
храняется долго. В работе [18] говорилось о срав-
вызываться непосредственно возмущением ко-
нении средних показателей, но бывает так, что
эфициентов r и τ, но способ описания переходов
усреднение данных не все говорит о сути явления,
между стадиями заболевания импульсами пара-
важна величина разброса значений внутри сопо-
метров не столь интересен. Структура предпола-
ставляемых групп.
гает спектр модификаций для анализа сценариев
О динамике иммунного ответа и последова-
элиминации вируса N(t) → 0 с дополнительным
тельности стадий коронавирусной инфекции в
активным воздействием нелинейного характера
настоящее время информации уже много, но она
в варианте модели (10).
противоречива. Логика подсказывает, что найдут
много разнообразных вариантов усиления ответа.
Включенный в модель (8) при рассмотрении
У пожилых людей с низким уровнем интерлейки-
инфекционного процесса параметр K по своему
на-7 и сниженной чувствительностью к γ-интер-
смыслу не отождествляется с емкостью «экологи-
ферону [73] наблюдается иммуносценция - оста-
ческой ниши» или величине доступного для зара-
навливается пролиферация T-лимфоцитов и им-
жения множества клеток просто потому, что ле-
мунный ответ становится недостаточным.
тальный исход тогда наступал значительно ранее
Однако летальные случаи отмечены у молодых и
достижения такого уровня. Положение K - уро-
здоровых, не имеющих сопутствующих заболева-
вень инфекции, который может существовать в
ний людей, чаще всего - у медиков. Смертность
системе только ограниченное время, но среда
врачей пока является главным аргументом влия-
данное значение не выдерживает, и этот уровень
ния дозы инфекции.
не будет постоянным для популяции. Соответ-
ственно процесс переходит в терминальную ста-
Публикуются сообщения, что самые тяжелые
дию, либо снижается до стабильно равновесного
последствия коронавируса для организма связа-
состояния. При эффективном и быстром сраба-
ны с гиперактивной реакций клеток собственно-
тывании иммунитета критическое состояние K не
го иммунитета [74]. Синдром высвобождения
достигается. Разумеется, в реальных процессах
цитокинов существует как долгое системное вос-
режим N(t) → 0 реализоваться не должен. Вариант
паление, но «цитокиновый шторм» после каскад-
потери диссипативных свойств решения модели
ного выброса сигнальных молекул - экстремаль-
считаем терминальным сценарием при N(t) = 5Н.
ная необратимая фаза [75]. После быстрой гипер-
Очевидно, в модели (8) есть простой и быстрый
цитокенемии апоптозу подвергаются и здоровые
летальный сценарий при очень большом и не-
клетки.
компенсируемом репродуктивном r-параметре,
Предположим, что алгоритм регуляции позво-
подобный N(t) = N(0)ert. Чем больше оказывают-
ляет корректировать ответ, быстрый массирован-
ся значения K и r, тем труднее организму преодо-
ный или отложенный по времени, но ориентиро-
БИОФИЗИКА том 66
№ 2
2021
404
ПЕPЕВАPЮXА
ванный на известные мишени. В таком ракурсе
симптомов сходных человеческим вирусом им-
выбираемая иммунитетом при «бессимптомном»
мунодефицита. У устойчивых обезьян оказалась
течении неспешная тактика управления ответом
ограничена иммунная активация, не запускалась
оказывается рациональным и предосторожным
пролиферация иммунных клеток CD4. Их денд-
подходом. Запускается механизм созревания аф-
ритные клетки слабо выбрасывают сигналы тре-
финности, который алгоритмически выглядит
воги цитокины при введении в опытах вируса
как задача поиска в пространстве из доступных
[78]. Математически имеем следующий сценарий
аминокислотных последовательностей антител с
в наших обозначениях: ∀N(0) < K: limt→∞ N(t) = H,
повышенными константами связывания с анти-
∀t > 0, N(t) ≤ H.
геном. С точки зрения биологической кибернети-
ки, если нет признаков экстремальности и ситуа-
Маленькие обезьяны получили оптимизаци-
ция позволяет, то оптимально выдержать время
онное решение проблемы инфекции вирусом им-
для создания средств с высоким сродством к ан-
мунодефицита обезьян - не борясь с вирусом.
тигенам: иммуноглобулинов IgG3 и зрелых CD8-
Когда мы научимся оптимально управлять им-
киллеров, чем немедленно выбрасывать в легкие
мунными реакциями, то получим замечательный
огромные порции нейтрофилов и макрофагов.
инструментарий борьбы даже с новыми угрозами
По всей видимости, SARS-CoV-2 способен про-
без острой фазы. Геном SARS-CoV-2 большой
никать в эти клетки и убивать лейкоциты, запус-
[79] по сравнению с его родственниками, он ко-
кая таким образом петлю разрушительной обрат-
дирует с неясной целью несколько новых корот-
ной связи воспаления в альвеолярные мешочки
ких белков [80]. Удалось экспрессировать 26 из
легких. Для сценария гибкого регулирования не-
29 его белков. Пять вирусных белков действи-
обходимо функционирование промежуточного
тельно направлены на противоборство с внутри-
звена, роль которого могут успешно выполнить
клеточным уровнем иммунитета [81]. Физически
NK-клетки, количество которых составляет <10%
РНК будет реплицироваться и упаковываться
от всех лимфоцитов. Важно исходное число
дольше и, в силу непреодолимой статистики, со
средств сдерживания ответа - IgG4, Т-супрессо-
случайными ошибками, делая полуфункцио-
ры, количество которых ≈ 5% в популяции всех
нальные варианты вирионов внутри организма,
CD4 клеток, но они не разделяются в обычных
но это увеличит антигенную дифференциацию
анализах.
инфекции.
Если видим слабую корреляцию гуморального
Актуально провести эксперименты, насколько
иммунного ответа, тиров антител с падением ви-
величина начальной дозы вируса может повы-
русной нагрузки, то вероятно влияние эффекта
шать вероятность запуска смертельного автоим-
антителозависимого усиления инфекции, когда
мунного сценария. В цитируемой ранее нами ра-
несовершенные антитела создают непрочные со-
боте [63] отмечалось, что индукция цитокинов
единения с вирусом, помогая таким образом ви-
была значительно выше у мышей, инфицирован-
русу проникать в лейкоциты, освобождаться и
ных высокой дозой, по сравнению с мышами, ко-
начинать репликацию в моноцитах и макрофагах.
торые первоначально получали низкую дозу ви-
Из-за данного эффекта провалилась компания
руса гриппа. Если предположить сильную зави-
вакцинации от лихорадки Денге на Филиппинах.
симость между начальной дозой вируса и
Оказалось [76], что непривитые и без антител бо-
(максимальной) концентрацией основных типов
лели вирусом легче, чем привитые, но с недоста-
цитокинов, то функциональная связь отразится
точно аффинными, плохо связывающимися IgG.
не гладкой кривой, а ступенчатой функцией. В
В Китае к началу лета 2020 г. не заявили о созда-
норме же ответ плавный [82].
нии вакцины, наиболее логично предположение,
что разработчики столкнулись с признаками ан-
Обратим внимание на работу [83], где в опытах
тителозависимого усиления инфекции (пусть да-
с туляремией было отмечено, что гиперцитокине-
же в 5% случаев, но это увеличивает риски). Со-
мия следует за начальной задержкой экспрессии
временное развитие молекулярного 3D-модели-
цитокинов. Значит, наиболее вероятен порого-
рования позволит решить проблему, точно
вый запуск процесса. Сценарий шторма матема-
выбрать оптимальный участок белка и собрать
тически возможен, когда внезапно достигается
ген для его экспрессии в вакцинном штамме.
порог и естественные средства иммуносупрессии
Известны примеры избегания серьезных по-
не в состоянии контролировать цепную быструю
следствий именно при своевременном сдержива-
активацию всех других типов клеток. Для стреми-
нии иммунной реакции [77]. Наиболее резонанс-
тельных переходов востребован метод триггерно-
ный случай успешного контроля ответа - вирус
го описания изменения в развитии сценариев. В
иммунодефицита обезьян у некоторых их видов
случае COVID начальная доза может быть не про-
(например, мангобеев Cercocebus atys) не вызыва-
сто значимым, но важнейшим фактором риска
ет иммунодефицит, хотя реплицируется в орга-
развития летального сценария посредством стре-
низме, но у других приматов ведет к развитию
мительной гиперактивации воспалительных ре-
БИОФИЗИКА том 66
№ 2
2021
НЕПРЕРЫВНАЯ МОДЕЛЬ ТРЕХ СЦЕНАРИЕВ
405
акций от собственной иммунной системы. Поро-
8. Э. М. Кольцова, Е. С. Куркина и А. М. Васецкий,
говое значение часто ближе, чем мы ожидаем.
Comput. Nanotechnol. 3 (1), 99 (2020).
Модель с неустойчивым достижимым равно-
9. А. Б. Белов и П. В. Куликов, Эпидемиология и вак-
весием K актуальна для прикладных задач экоди-
цинопрофилактика 18 (5), 109 (2019).
намики: биологической борьбы с опасными ин-
10. L. Allen, An introduction to stochastic processes with ap-
вазивными видами [84], управления рыбными ре-
plications to biology (Pearson Prentice Hall, 2003).
сурсами [85] и биоремедиации водоемов после
11. Г. Ш. Цициашвили и М. А. Осипова, Дальнево-
нефтяного загрязнения [86]. Уравнение предпо-
сточ. мат. журн. 2 (1), 58 (2001).
лагает широкие возможности дальнейших моди-
12. А. В. Нагаев и А. В. Старцев, Мат. заметки 3 (2), 179
фикаций, как для оценки стратегии антивирус-
(1968).
ной терапии, так и сценариев заражения с учетом
вакцинации, сокращающей время выработки эф-
13. Ш. К. Форманов, А. Н. Старцев и С. С. Седов,
фективного ответа от клеточного иммунитета.
Дискретная математика 25, 103 (2013).
Большинство современных вакцин оцениваются
14. X. He, E. H. Y. Lau, and P. Wu, Nature Med. 26, 672
по величине титров антиген-специфических ан-
(2020).
тител в сыворотке или слизистой после вакцина-
15. National Health Commission of the People’s Republic
ции, но авторы работы [87] обосновывают как бо-
of China. Decision of awarding outstanding groups and
лее значимый фактор оценки по скорости реак-
individuals in the National Health System for excellent
ции от клеток иммунной памяти. В работе [88]
work in the prevention and control of Covid-19
отмечается «узкое место» работы самого эффек-
(March 5, 2020).
тивного звена иммунитета, скорость продвиже-
16. H. Nishiura, N. M. Linton, and A. R. Akhmetzhanov,
ния Т-лифоцитов в лимфатические узлы может
Int. J. Infect. Dis. 93 (2), 284 (2020).
быть просто ограничена пропускной способно-
17. S. Fafi-Kremer, T. Bruel, Y. Madec, et al., medRxiv
стью венул с высоким эндотелием.
preprint (2020). DOI: 10.1101/2020.05.19.20101832
18. G. De Natale, V. Ricciardi, and G. De Luca, J. Clin.
ФИНАНСИРОВАНИЕ РАБОТЫ
Med. 9, 1564 (2020).
Работа выполнена при финансовой поддержке
19. E. Lavezzo, E. Franchin, and C. Ciavarella, medRxiv
Российского фонда фундаментальных исследова-
preprint (2020). DOI: 10.1101/2020.04.17.20053157
ний (грант № 17-07-00125) и Проекта научных ра-
20. L. Ferretti, C. Wymant, M. Kendall, et al., Science 368
бот СПИИРАН по бюджетной теме № 0073-2019-
(6491), eabb6936 (2020).
0003.
21. J. A. Al-Tawfiq and A. J. Rodriguez-Morales, J. Hospi-
tal Infect. 105 (1), 111 (2020).
КОНФЛИКТ ИНТЕРЕСОВ
22. J. H. Kim, I. Skountzou, and R. Compans, J. Immu-
nol. 183 (1), 3294 (2009).
Автор заявляет об отсутствии конфликта инте-
ресов.
23. А. В. Гаврилина и С. В. Соколов, Процессы управ-
ления и устойчивость 11 (1), 229 (2018).
24. А. В. Шабунин, Прикладная нелинейная динами-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
ка 27 (2), 5 (2019).
Настоящая работа не содержит описания ка-
25. А. Н. Герасимов, Мат. моделирование 31 (3), 109
ких-либо исследований с использованием людей
(2019).
и животных в качестве объектов.
26. E. Waltersa, M. I. MeslébcIan, and M. Hall, Epidemics
25 (1), 8 (2018).
СПИСОК ЛИТЕРАТУРЫ
27. V. C. C. Cheng, Clin. Microbiol. Rev. 20 (4), 60 (2007).
1. A. Y. Perevaryukha, Biophysics 61 (2), 334 (2016).
28. M. A. Gouilh, S. J. Puechmaille, and J. P. Gonzalez,
Infection, Genetics and Evolution 11 (7), 1690 (2011).
2. M. Jeschke, Diversity and Distributions 20 (11), 1229
(2014).
29. А. М. Молчанов, В. Г. Назаренко и И. Г. Шатур-
3. А. В. Pуcаков, А. Б. Медвинcкий, Б. Д. Ли и
ный, Биофизика 16 (4), 667 (1971).
М. М. Гоник, Биофизика 54 (4), 733 (2009).
30. Т. Е. Санникова, Г. И. Марчук, А. А. Романюха и
4. В. В. Нечаев, К. В. Жданов и Г. И. Гришанова,
А. И. Яшин, Успехи геронтологии 12, 091 (2003).
Журнал инфектологии 7 (2), 21 (2015).
31. R. Medzhitov, P. Preston-Hurlburt, and C. A. Janeway,
5. А. Б. Медвинский, Биофизика 51 (6), 1033 (2006).
Nature 388 (6640), 394 (1997).
6. S. Asadi, S. Wexler, and D. Cappa, Sci. Rep. 9 (6), 2348
32. A. Y. Perevaryukha, Cybernetics and Systems Analysis
(2019).
52 (4), 623 (2016).
7. L. Anderson, P. Turnham, R. Griffin, and C. Clarke,
33. Yu. S. Kolesov and D. I. Shvitra, Lithuanian Math. J.
Risk Analysis 40 (5), 902 (2020).
19 (1), 81 (1979).
БИОФИЗИКА том 66
№ 2
2021
406
ПЕPЕВАPЮXА
34. G. Bocharov and F. Rihan, J. Comput. Appl. Math.
62. A. Paulo and M. Correia-Neves, PLoS One 5 (7),
125 (2), 183 (2000).
e11655 (2010).
35. A. Y. Morozov, M. Banerjee, and S. V. Petrovskii, J.
63. V. Davids, W. Hanekom, and J. Gelderbloem, Clin.
Theor. Biol. 396 (1), 116 (2016).
Vaccine Immunol. 14 (2), 198 (2007).
36. О. Ф. Воропаева и С. Д. Сенотрусова, Мат. моде-
64. I. Marois, A. Cloutier, E. Garneau, and V. Richter, J.
Leukocyte Biol. 92 (1), 107 (2012).
лирование 29 (9), 135 (2017).
65. A. M. Collins and K. J. Jackson, Frontiers Immunol. 4
37. R. E. Baker and G. Röst, J. Nonlinear Sci. 30 (2), 397
(2), 235 (2013).
(2020).
66. E. Shin and S. Sung, Nature Rev. Immunol. 16, 509
38. G. E. Hutchinson, Ann. N. Y. Acad. Sci. 50 221 (1948).
(2016).
39. S. A. Kashchenko, Automatic Control and Comput.
67. Н. А. Арсентьева и А. В. Семенов, Инфекция и им-
Sci. 47 (3), 470 (2013).
мунитет 2 (4), 687 (2012).
40. E. M. Wright, J. für die Reine und Angewandte Math-
68. J. Schoggins and C. Rice, Curr. Opin. Virology 1 (6),
ematik 194 (1) 66 (1955).
519 (2011).
41. R. M. May, Stability and Complexity of Ecosystems
69. K. L. Yap, I. F. Ada, and G. L. McKenzie, Nature 273
(Princeton University Press, 1974).
(5659), 238 (1978).
42. J. Van Dyken, J. Theor. Biol. 460 (1), 115 (2019).
70. А. Ю. Переварюха, Журн. общ. биологии 81 (3), 174
43. А. Ю. Переварюха, Биофизика 65 (1), 138 (2020).
(2020).
44. А. Ю. Колесов и Н. Х. Розов, Мат. сборник 202 (6),
71. N. Mark, A. E. Hubbard, and M. Jones, Appl. Biosafe-
51 (2011).
ty 9 (3), 118 (2004).
45. T. L. Kaplan and T. A. Yorke, J. Math. Anal. Appl. 48
72. N. Hanna, R. Kalderon, and D. Nelken, Immunology
(2), 317 (1974).
29 (3), 433 (1975).
73. G. Siracusano, C. Pastori, and L. Lopalco, Frontiers
46. А. Ю. Колесов и Н. Х. Розов, Труды Мат. института
Immunol. 11, 1049 (2020).
им. Стеклова 277, 101 (2012).
74. И. М. Кветной и М. А. Пальцев, Нейроиммуноэндо-
47. С. В. Лазаренко, Мат. машины и системы 1, 28
кринные механизмы старения и возрастной патоло-
(2019).
гии (Наука, СПб, 2012).
48. М. В. Глаголев, А. Ф. Сабреков и В. М. Гончаров,
75. P. Mehta, D. F. McAuley, and M. Brown, Lancet 395
Динамика окружающей среды и глобальные изме-
(10229), 1033 (2020).
нения климата 9 (2), 40 (2018).
76. J. Teijaro, K. Walsh, S. Rice, et al., Proc. Natl. Acad.
49. W. S. Gurney, S. P. Blythe, and R. M. Nisbet, Nature
Sci. USA 111 (10), 3799 (2014).
287, 17 (1980).
50. С. Д. Глызин, Модел. и анализ информ. систем 3,
77. S. Halstead and P. Russell, Vaccine 34 (14), 1643
365 (2017).
(2016).
51. K. Gopalsamy, Stability and Oscillations in Delay Dif-
78. N. R. Klatt, G. Silvestri, and V. Hirsch, Cold Spring
ferential Equations of Population Dynamics (Springer,
Harb. Perspect. Med. 2 (1), a007153 (2012).
Netherlands, 1992).
79. J. Mandl, A. Barry, and T. Vanderford, Nature Med.
52. N. A. Bacaër, Short History of Mathematical Population
14, 1077-1087 (2008).
Dynamics (Springer, Berlin, 2011).
80. F. A. Isihak, M. A. Hamad, and N. G. Mustafa, Ин-
53. S. Ruan, in Delay Differential Equations and Applica-
фекция и иммунитет 10 (2), 247 (2020).
tions (Springer, Berlin, 2006), pp. 477-517.
81. E. S. Brielle, D. Schneidman-Duhovny, and M. Linial,
54. A. Y. Perevaryukha, Cybernetics and Systems Analysis
Viruses 12 (5), 497 (2020).
55 (1), 141 (2019).
82. L. Malle, Nature Rev. Immunol. 20, 351 (2020).
55. U.S. Forest Service. 2018. Forest Health Conditions in
Alaska - 2017. U.S. Forest Service, Alaska Region, An-
83. M. W. Deem and P. Hejazi, Annu. Rev. Chem. Biomol.
chorage, AK. Publication R10-PR-43, 74 pp.
Engineer. 1 (1), 247 (2010).
56. R. Lia, L. Xua, and N. Bjørnstadc, Proc. Natl. Acad.
84. C. A. Mares, S. S. Ojeda, and E. G. Morris, Infection
Sci. USA 116, 3624 (2019).
and Immunity 76 (7), 3001 (2008).
57. T. Royama, W. E. Mackinnon, and E. G. Kettela, Ecol-
85. A. Y. Perevaryukha, Cybernetics and Systems Analysis
ogy 86 (5), 1212 (2005).
56 (1), 139 (2020).
58. L. Geritz and S. A. Kisdi, J. Math. Biol. 65, 1411
86. Е. А. Криксунов, А. Е. Бобырев и В. А. Бурмен-
(2012).
ский, Журн. общ. биологии 71 (5), 436 (2010).
59. Y. Kuang, Delay Differential Equations with Applications
to Population Biology (Academic Press, New York,
87. V. A. Gushchin, A. I. Sukhinov, and A. V. Nikitina,
1993).
Comput. Math. and Math. Physics 58 (8), 1316 (2018).
60. P. Klein, J. Sterzl, and J. Dolezal, J. Math. Biol. 13 (1),
88. J. Sarkander, S. Hojyo, and K. Tokoyoda, Clin. Trans-
67 (1981).
lat. Immunol. 5 (12), e120 (2016).
61. T. J. Lunn, O. Restif, and J. Peel, Phil. Trans. R. Soc. B
89. А. В. Бондарь, Ю. П. Гумовская и А. В. Полевщи-
374, 20190016 (2019).
ков, Биофизика 61 (5), 927 (2016).
БИОФИЗИКА том 66
№ 2
2021
НЕПРЕРЫВНАЯ МОДЕЛЬ ТРЕХ СЦЕНАРИЕВ
407
Continuous Model for the Three Scenarios of the Infection Routes Given Delayed
Immune Response Factors
A.Yu. Perevaryukha
St. Petersburg Institute for Informatics and Automation, Russian Academy of Sciences,
14-ya Linia 39, St. Petersburg, 199178 Russia
The article describes models of the course of an infection as controlled nonlinear processes. Understanding
significant differences observed in the trajectory of the disease caused by a new coronavirus SARS-CoV-2 is
of critical importance at the moment. Myriad of factors are regarded as causes of disease when symptoms are
highly variable in different people and the transmission rate of infection with the new coronavirus through
local population varies. Viral replication by which the virus makes copies of itself within a host cell and is able
to infect other cells and the development of an immune response to its antigens within an organism are two
interdependent processes which have aftereffects and depend on the previous conditions of the cell and virus
populations. Different scenarios without altering input parameters are necessary for modeling the properties
of these states. The most important factor is the effectiveness of the immune response, including the time of
defense responses from the three levels of the immune system, the complex system in the body. We have pro-
posed a computational description of infection development scenarios based on a delayed differential equa-
tion with two values of the delayed time. In a new model, transitions between phases during infectious disease
result from the level of initial infection and a delayed immune response to infection. The variation in infec-
tious dose and response time can lead to the transition from the acute phase of the disease with pronounced
symptoms to a chronic stage, or fatal outcome. In the created model, we calculated and described asymptom-
atic transmission of viral infection by considering the possibility of rapid and effective suppression of the virus
after active virus replication that occurs a short time but maintaining a small amount of the virus in the body.
An analysis of the model’s behavior is consistent with the theory that the initial number of HCV virions can
affect the quality of the adaptive immune response. There are big differences in the trajectory of COVID-19
disease and the formation of the types of the immune system’s response to coronavirus and these processes
are not yet well understood. Model scenarios are developed based on the trajectories of the studied hepatitis
C virus variants.
Keywords: modeling infection routes, delayed regulation, immune response, model for hepatitis C virus chronicity,
initial infection dose, epidemics, virus spread rate, asymptomatic scenario, COVID-19 variability
БИОФИЗИКА том 66
№ 2
2021