БИОФИЗИКА, 2021, том 66, № 3, с. 495-503
БИОФИЗИКА КЛЕТКИ
УДК 582.998.2:547.587
КИНЕТИКА ИНИЦИИРОВАННОГО ОКИСЛЕНИЯ ЛИПОСОМ
ИЗ ФОСФАТИДИЛХОЛИНА С ВВЕДЕННЫМИ В НИХ ЭКСТРАКТАМИ
АЛОЕ И ОПРЕДЕЛЕНИЕ ИХ АНТИОКИСЛИТЕЛЬНОЙ АКТИВНОСТИ
© 2021 г. Н.Н. Сажина*, П.В. Лапшин**, Н.В. Загоскина**
*Институт биохимической физики им. Н.М. Эмануэля РАН, 119334, Москва, ул. Косыгина, 4
**Институт физиологии растений им. К.A. Тимирязева РАН, 127276, Москва, Ботаническая ул., 35
E-mail: Natnik48s@yandex.ru
Поступила в редакцию 15.12.2020 г.
После доработки 10.01.2021 г.
Принята к публикации 20.01.2021 г.
Многочисленные виды растений рода Алое привлекают внимание исследователей различными про-
явлениями их биологической активности, в том числе антиоксидантной. В настоящей работе про-
ведено сравнительное исследование кинетики ингибирующего действия этанольных экстрактов
пяти видов Алое в модельной системе инициированного окисления фосфатидилхолиновых липо-
сом. Установлено, что экстракты A. marlothii и A. сongolensis обладают, соответственно, в 13 и 10 раз
большей антиоксидантной активностью, чем традиционные виды Алое: A. arborescens и А. vera. Зна-
чения суммарного содержания фенольных соединений экстрактов A. marlothii и A. сongolensis превы-
шают значения содержания фенольных соединений A. arborescens и А. vera в меньшей степени, чем
антиоксидантная активность (в пять-шесть раз). Это может свидетельствовать о наличии в экстрак-
тах A. marlothii и A. сongolensis очень активных фенольных антиоксидантов. Анализ влияния введен-
ных в липосомы экстрактов на их размер показал, что наиболее активные экстракты A. marlothii,
A. сongolensis и A. pillansii уменьшают средний размер липосом в сравнении с «чистыми» липосома-
ми, а экстракты с меньшей антиоксидантной активностью увеличивают, что связано, вероятно, с
различным изменением липидной структуры липосом компонентами экстрактов. Полученные ре-
зультаты позволяют рекомендовать A. marlothii, A. сongolensis и A. pillansii для изучения других видов
их биологической активности и создания на их основе новых лекарственных средств.
Ключевые слова: Алое, фенольные метаболиты, кинетика, окисление, липосомы, фосфатидилхолин.
DOI: 10.31857/S0006302921030108
тракты Алое помогают лечить гастриты и язвы
В мире насчитывается более 500 различных ви-
желудка [5]. Антидиабетическая и антигиперли-
дов рода Алое. Они являются важными источни-
ками биологически активных веществ [1]. К фи-
пидемическая активности компонент Алое позво-
ляют бороться с сахарным диабетом и жировой
тохимическим классам, присущим Алое, относят-
болезнью печени [6]. Отдельные компоненты
ся: антроны, хромоны, кумарины, пироны,
алкалоиды, флаваноиды, стиролы, энзимы, про-
Алое (алое-эмодин, алоин и др) обладают антиви-
теины, жирные кислоты и другие липидные ком-
русной, иммуномодулирующей активностями и
поненты, практически все витамины (В1, В2, В6,
используются для лечения заболеваний верхних
дыхательных путей [1, 7]. Некоторые соединения
В12, С, Е, бета-каротин и др.) [1]. Благодаря раз-
Алое обладают противоопухолевой активностью
ному биологическому действию соединений этих
[8, 9]. Было найдено, что алое-эмодин, один из
классов, Алое широко применяют в традицион-
основных компонентов Алое, ингибирует ак-
ной и современной медицине [2]. При лечении
тивность ацетилхолинэстеразы, а экстракт
гепатита и алкогольной интоксикации печени ис-
A. arborescens защищает IMR32, нейробластому
пользуются гепатопротекторные свойства Алое [3,
клеточной линии человека, от токсичности, вы-
4]. Благодаря пребиотической и антибактериаль-
званной бета-амилоидным пептидом, ответ-
ной активности в гастроэнтерологии сок и экс-
ственным за болезнь Альцгеймера [2, 10].
Сокращения: АОА - антиоксидантная активность, ФХ -
Одна из причин и стадий перечисленных бо-
фосфатидилхолин, ААРН - 2,2′-азо-бис(2-амидинопро-
пан)дигидрохлорид, СФС - суммарное содержание фе-
лезней - окислительный стресс, обусловленный
нольных соединений, ДК - диеновые конъюгаты.
перекисным окислением липидов в мембранах
495
496
САЖИНА и др.
клеток крови, печени, мозга, эпителия и других,
размером ≈ 5 мм и лиофильно высушивали до по-
поэтому для ингибирования окислительного про-
лучения сухой массы. Экстракты готовили из
цесса важно найти Алое с высоким содержанием
50 мг сухих листьев в 1.5 мл 70%-го этанола, пере-
активных антиоксидантов. Наиболее исследо-
мешивали в термостате в течение 45 мин при тем-
ванными и используемыми видами Алое являют-
пературе 45°С [20]. Далее центрифугировали с от-
ся Aloe arborescens Mill., Aloe ferox Mill. и Aloe vera
носительным ускорением 12000 g (13000 об/мин)
(L.) Burm. [1]. Однако встречаются мало изучен-
на центрифуге Hettich (Германия) и отбирали на-
ные виды с более активными антиоксидантными
досадочную жидкость (супернатант). Готовые
компонентами. В работе [11] амперометрическим
экстракты хранили при 4°С. Весовая концентра-
и хемилюминесцентным методами был проведен
ция всех полученных экстрактов составила
скрининг 15 видов Алое по оценке антиоксидант-
33.3 мг сухих листьев на 1 мл этанола. В качестве
ной активности (АОА) их экстрактов и найден
вещества сравнения использовали галловую кис-
вид Алое A. pillansii со значительно более высоким
лоту (Sigma-Aldrich, США). Суммарное содержа-
значением АОА, чем у традиционных видов Алое.
ние фенольных соединений (СФС), выраженное
Поиск продуцентов биологически активных ве-
в мг-экв галловой кислоты на 1 г сухих листьев,
ществ с высокими антиокислительными свой-
определяли спектрофотометрическим методом
ствами в других видах Алое востребован и являет-
(725 нм) с помощью реактива Фолина-Чокальтеу
ся актуальной задачей современной биологии и
[20]. Погрешность определения СФС составила
фармацевтики.
менее 3%.
В настоящее время для исследования биохи-
Приготовление суспензии липосом. Для приго-
мических процессов в мембранах клеток живых
товления липосом использовали суспензию сое-
организмов, в частности процесса перекисного
вого ФХ марки L-α-phosphatidylcholine P3644
окисления липидов, широко используют различ-
(Sigma-Aldrich, США) в фосфатном буфере
ные липосомные структуры как модели биомем-
(NaH2PO4 + Na2HPO4) с рН 7.4 и ионной силой
бран [12-15]. Инициируя окисление липосом
1 мM. В состав ФХ данной марки входили два ос-
разными способами, можно регулировать окис-
новных фосфолипида - фосфатидилхолин (55%)
лительный процесс различными субстанциями,
и фосфатидилэтаноламин
(25%). Содержание
встроенными в липосомы. Ингибирование окис-
ЖК в % к общему их количеству составило: паль-
ления липосом антиоксидантами дает более ре-
митиновая - 17%, стеариновая - 4%, олеиновая -
альную «физиологичную» модель для определе-
9%, линолевая - 60%, линоленовая - 7%. Сус-
ния их АОА. Поэтому в нашей работе мы исполь-
пензию готовили из навески ФХ с объемом буфе-
зовали модельную систему инициированного
ра, необходимым для получения концентрации
окисления липосом из соевого фосфатидилхоли-
1 мг/мл. Смесь перемешивали в шейкере, добав-
на (ФХ) при Т = 37°C [14-16]. Кроме использова-
ляя разные объемы (от 10 до 100 мкл) спиртовых
ния в качестве модели биологических мембран,
экстрактов Алое. Липосомы формировали c помо-
липосомы из ФХ можно использовать также как
щью ультразвукового гомогенизатора VCX-130
потенциальные наноконтейнеры для доставки в
(Sonics & Materials, США) при мощности 70 Вт в
ткани и клетки живых организмов различных ле-
течение 15 мин в режиме 15 с × 15 с, для предохра-
карственных препаратов, в том числе и экстрак-
нения липосом от окисления сосуд с суспензией
тов Алое, поэтому целесообразно определить фи-
помещали в смесь воды со льдом. Для отделения
зико-химические характеристики таких липосом
липосом от примесей, образующихся, в том чис-
[12, 17-19].
ле, и после «озвучивания», дисперсию липосом
Цель настоящей работы - изучение кинетики
центрифугировали на центрифуге Hettich (Герма-
ингибирующего действия экстрактов пяти видов
ния), при 12000 g (13000 об/мин) и 4°С в течение
рода Алое на модели инициированного окисления
20 мин, а супернатант отбирали для дальнейшей
липосом из ФХ и определение их АОА, а также
работы [16, 21]. Для определения концентрацион-
сравнение размеров и ζ-потенциала липосом с
ных зависимостей ингибирующего действия экс-
введенными в них экстрактами Алое.
трактов Алое мы ввели величину содержания су-
хих листьев в известном объеме экстракта, вве-
денного в суспензию липосом, к массе ФХ в
МАТЕРИАЛЫ И МЕТОДЫ
липосомах С = mС.Л/mФХ, где mС.Л - масса сухих
Получение экстрактов Алое. В работе использо-
листьев в введенном объеме экстракта в мг,
вали пять представителей рода Алое: A. аrborescens,
mФХ - масса навески ФХ в мг.
A. vera, A. pillansii, A. congolensis A. marlothii, выра-
щенных в коллекции суккулентных растений в
Инициирование окисления липосом. Для ини-
Институте физиологии растений им. К.A. Тими-
циирования окисления липосом использовали
рязева РАН (Москва) при естественном освеще-
водорастворимый азо-инициатор
2,2′-азо-
нии и 16-часовом фотопериоде. Листья взрослых
бис(амидинопропан)дигидрохлорид (ААРН; Flu-
(годовалых) растений измельчали до фрагментов
ka, Германия) с конечной концентрацией в дис-
БИОФИЗИКА том 66
№ 3
2021
КИНЕТИКА ИНИЦИИРОВАННОГО ОКИСЛЕНИЯ ЛИПОСОМ
497
персии липосом 0.33 мМ. Исходный раствор
±5%. Суммарная ошибка составляла не более
ААРН (200 мМ) готовили на дистиллированной
±10%.
воде при Т ≈ 0°С и хранили в морозильнике. Пе-
Методы статистической обработки. Результаты
ред измерением разбавляли в десять раз буфер-
экспериментов представлены как средняя вели-
ным раствором в сосуде, помещенном в лед, из
чина ± стандартная ошибка средней при помощи
которого отбирали 50 мкл для анализа. Окисле-
программного пакета Microsoft® Office Excel.
ние липосом с концентрацией ФХ 0.1 мг/мл про-
Статистическую обработку результатов измере-
водили в спектрофотометрической кварцевой
ний осуществляли с использованием t-критерия
кювете с конечным объемом рабочей жидкости
Стьюдента [22]. Различия считали статистически
3 мл, термостатированной при физиологической
достоверными при уровне различия p ≤ 0.05. Из-
температуре (37°С). Кинетику увеличения опти-
мерения повторяли не менее трех раз. Погреш-
ческой плотности ΔА (А - А0), т.е. приращения
ность определения ингибирующей активности
образования диеновых конъюгатов (ДК) в про-
экстрактов Алое не превышала 10%.
цессе перекисного окисления липидов, регистри-
ровали во времени при λ = 234 нм на двухлучевом
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
спектрофотометре (Perkin Elmer, США). Для ис-
ключения влияния спектра поглощения ААРН в
Сначала были изучены спектры поглощения
кювету сравнения добавляли такое же количество
этанольных экстрактов всех исследованных ви-
ААРН, как и в рабочую кювету, а вместо диспер-
дов Алое (рис. 1). Они существенно различались
сии липосом - такой же объем буфера. Добавле-
между собой. Наибольшее поглощение компо-
ние 100 мкл 70%-го этанола вместо экстрактов
нентами всех экстрактов наблюдается в диапазо-
при формировании «чистых» липосом не изменя-
не 200-400 нм, что характерно для фенольных со-
ло параметры окисления по сравнению с «бес-
единений [20]. Это относится к таким веществам,
спиртовыми» липосомами в пределах ошибки из-
содержащимся в листьях Алое, как алоин, ало-
мерений.
энин, алоэ-эмодин, алоэзин, алоэрезин, алоэно-
зид и др. [23-26]. Для A. arborescens содержание
Основными показателями, которые использо-
алоэнина составляет 20.7% от суммы фенольных
вали при изучении кинетики образования ДК,
соединений, на долю алоинов приходится 26.2%
были: τ - период индукции, Vm - максимальная
[24]. Для A. arborescens и A. vera, эти фенольные
скорость окисления, tm - время достижения Vm
метаболиты определяют, главным образом, анти-
[16, 27]. Спектры поглощения этанольных экс-
радикальную активность [25, 26]. Спектры погло-
трактов Алое регистрировали в диапазоне 200-
щения трех чистых метаболитов Алое приведены в
700 нм на вышеупомянутом спектрофотометре.
работе [23]. Для алоэнина максимумы полос по-
глощения 218 и 308 нм, для алоэрезина - 216 и
Метод динамического светорассеяния. Для
302 нм, у барбалоина - четыре пика поглощения:
определения среднего размера липосом в суспен-
при 226, 270, 298 и 360 нм [23]. Сопоставление
зии использовали метод динамического свето-
этих и наших данных (рис. 1) позволяет каче-
рассеяния. Измерения проводили на высокочув-
ственно оценить вклад указанных фенольных ме-
ствительном приборе Zetasizer Nano ZS (Malvern
таболитов в спектральные характеристики экс-
Instruments, Великобритания) по оценке корре-
трактов исследуемых видов Алое. Так, основной
ляционной функции флуктуаций интенсивности
максимум поглощения этанольного экстракта
лазерного излучения с λ = 633 нм, рассеянного на
листьев A. аrborescens (спектр 1) отмечен около
липосомах под углом 173° [21]. Концентрация
300 нм, что может свидетельствовать о наличии в
ФХ-липосом с введенным экстрактами Алое со-
нем алоина, алоэнина, алоэрезина, как и отмеча-
ставляла ≈0.5 мг/мл, температура 25°С, индекс
лось в [23, 24]. Для этанольного экстракта листьев
полидисперсности варьировал в пределах 0.23-
A. pillansii (спектр 3) характерны три ярко выра-
0.25. Для каждой пробы осуществляли регистра-
женные полосы поглощения в области 270, 300 и
цию пяти-семи распределений интенсивности по
360 нм, что предполагает наличие в нем не только
размерам частиц, усреднение проводили по раз-
алоина, алоэнина, алоерезина, но и других фе-
мерам частиц с максимальной интенсивностью
нольных метаболитов. Похожий спектр поглоще-
рассеяния с погрешностью не более ±5%. При
ния имеет экстракт A. vera (спектр 2), однако кон-
измерении ζ-потенциала липосом прибор ис-
центрации его составляющих в полтора-два раза
пользовали в режиме регистрации электрофоре-
меньше. Для экстракта A. marlothii (спектр 5)
тической подвижности частиц, по которой рас-
можно отметить две полосы поглощения вблизи
считывали распределение для ζ-потенциала.
300 и 250 нм. Экстракт A. сongolensis (спектр 4)
Усреднение максимальных значений ζ-потенци-
имеет очень высокий пик вблизи 230 нм. Возмож-
ала проводили по трем повторным измерениям.
но, в этом случае доминирует максимум погло-
Калибровку проводили по стандартной диспер-
щения барбалоина, присутствующего в высокой
сии полистирольного латекса с погрешностью
концентрации, на фоне поглощения алоэнина,
БИОФИЗИКА том 66
№ 3
2021
498
САЖИНА и др.
Рис. 1. Спектры поглощения 70%-х этанольных экстрактов пяти видов Алое: 1 - A. аrborescens, 2 - A. vera, 3 - A. pillansii,
4 - A. сongolensis, 5 - A. marlothii. Разведение в 200 раз.
алоерезина и других близких метаболитов. Не ис-
golensis) (см. рис. 3). Накопление ДК в максимуме
ключено также наличие во всех изученных экс-
кривой 5 несколько выше, чем у других (~10%),
трактах антронов, антрохинонов, алоэсапонари-
что, возможно, связано с присутствием в экстрак-
нов, производных эмодина, также имеющих не-
те A. marlothii фитостеролов или других липидных
сколько полос поглощения в этой области
субстратов [1]. Максимальная скорость окисле-
спектра [1].
ния липосом, от которой зависит τ, заметно сни-
Определение суммарного содержания феноль-
жается у кривых 4 и 5 по сравнению с остальными
ных соединений в изученных экстрактах (СФС в
кривыми, что в значительной степени связано с
мг-экв галловой кислоты на 1 г сухого листа) по-
расходованием к этому времени ненасыщенных
казало небольшие и близкие их значения для
липидов ФХ, от концентрации которых главным
A. arborescens и A. vera (рис. 2). В экстрактах
образом и зависит скорость окисления [16, 27, 28].
остальных видов их уровень был в пять-шесть раз
При больших временах окисления на скорость
выше. Исходя из этих данных, можно отметить
может влиять также снижение концентрации
высокую способность к биосинтезу вторичных
ААРН [27]. Учитывая величину молярного коэф-
фенольных метаболитов у A. pillansii, A. marlothii и
A. сongolensis в сравнении с «традиционными» ви-
дами Алое.
Следующим этапом нашего исследования яви-
лось изучение процессов инициированного окис-
ления липосом из ФХ и ингибирования окисле-
ния экстрактами Алое, введенными в липосомы.
Механизм и соответствующие уравнения иници-
ированного ААРН радикально-цепного окисле-
ния жирных кислот (ЖК) в липосомах подробно
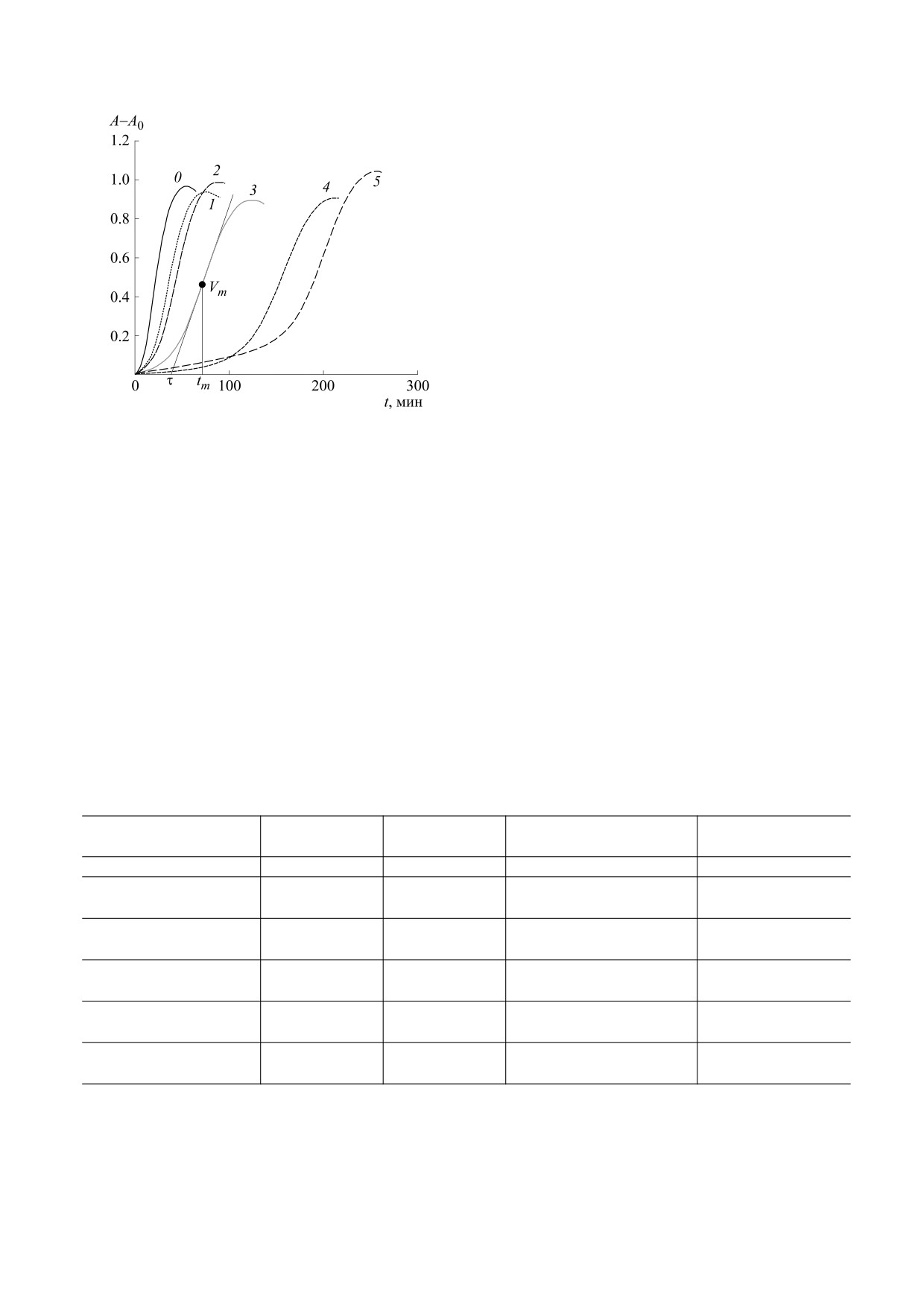
рассмотрены в работах [14, 16, 27, 28]. На рис. 3
представлены сравнительные кинетические кри-
вые образования ДК (в процессе инициирован-
ного окисления липосом без экстракта Алое и с
включенными в них экстрактами пяти видов Алое
с равными значениями С = mС.Л./mФХ, где mС.Л. -
масса сухих листьев в введенном в липосомы объ-
еме экстракта в мг, mФХ - масса ФХ в липосомах
Рис. 2. Содержание фенольных соединений (СФС, в
в мг.
мг-экв галловой кислоты на г сухого листа) в
этанольных экстрактах листьев изученных видов
При одинаковых значениях С значительно бо-
Алое: 1 - A. аrborescens, 2 - A. vera, 3 - A. pillansii, 4 -
лее продолжительные периоды индукции демон-
A. сongolensis,
5
- A. marlothii; *
- p < 0.05 по
стрируют экстракты 5 (A. marlothii) и 4 (A. сon-
отношению к образцу 1.
БИОФИЗИКА том 66
№ 3
2021
КИНЕТИКА ИНИЦИИРОВАННОГО ОКИСЛЕНИЯ ЛИПОСОМ
499
липосом с экстрактом 1 (A. arborescens) примерно
в два раза. Если учесть, что τ = tm - (A - A0)m/Vm
(рис. 3), то оценки влияния снижения Vm на τ по-
казывают, что для больших значений tm (рис. 3,
кривые 4 и 5) эти изменения для τ не очень значи-
тельны (~5-7%) и меньше общей погрешности
определения АОА.
Для сравнения степени ингибирования окисле-
ния липосом экстрактами Алое мы приняли без-
размерный параметр (τ - τ0)/τ0, где τ0 - период
индукции в отсутствие экстрактов для кривой 0
(τ0 = (5.0 ± 0.5) мин), обусловленный кинетикой
процесса накопления ДК и зависящий от концен-
траций ФХ и ААРН [16, 27-29]. Для ААРН ско-
рость инициирования радикалов Ri = ki[AAPH],
где ki = 1.3 ∙ 1013 ∙ е-112600/RT - константа скорости
Рис. 3. Кинетика образования ДК в процессе иници-
ированного окисления ФХ-липосом: 0 - без введе-
этой реакции [29]. При Т = 37°С, ki = 1.36 ∙ 10-6 с-1.
ния в них экстрактов, с введенными экстрактами: 1 -
Согласно законам жидкофазного окисления угле-
A. arborescens,
2
- A. vera,
3
- A. pillansii,
4
-
A. сongolensis,
5
- A. marlothi, [ФХ] = 0.1 мг/мл,
водородов и липидов с квадратичным обрывом це-
[ААРН] = 0.33 мМ, С = 0.14 мгС.Л./мгФХ, Т = 37°С,
пей, при неингибированном окислении cкорость
- опти-
А - оптическая плотность ДК при 234 нм, А0
образования ДК (гидропероксидов НР) d[HP]/dt =
ческая плотность при t = 0 мин, Vm - максимальная
= kр/(2kt)0.5⋅[LH]⋅Ri0.5, где kр и kt - эффективные
скорость окисления, tm - время достижения Vm, τ -
период индукции.
константы скорости продолжения и обрыва
цепей окисления, [LH] - концентрация окисляе-
мых липидов. Для ингибированного окисления
фициента поглощения ДК при λ = 234 нм, равную
d[HP]/dt = kр/2ki[AО]⋅[LHo]⋅Ri, где [AО] - концен-
274000 М-1 см-1 [15, 16], по наклону касательной
трация ингибитора (антиоксиданта). Если [AО]
можно определить максимальную скорость Vm
снижается линейно со временем ([AО] = [AО]о -
формирования ДК для всех кинетических кривых
- Rit/f), то по окончании ингибирования [AО] = 0,
на рис. 3 (см. таблицу). Максимальная скорость
а t = τ, и для разных кинетических кривых на рис. 3
окисления липосом с включенным экстрактом 4
можно сделать сравнительные оценки величин
(A. сongolensis) снижается по сравнению с Vm для
f⋅[AО]о = Riτ, где f - стехиометрический коэффи-
Результаты экспериментов после их статистической обработки
№ образца и вид Алое
[AO]o·f, ± SD,
y = аnх + b
АОА ± SD, нмоль
Vm ± SD,
нМ/мин
мкМ
Trэкв/мгС.Л.
0
117.2 ± 1.2
-
-
-
1. A. аrborescens
91.2 ± 1.5
0.43 ± 0.03
y = 73.88x + 0.91
22.8 ± 2.0
r2 = 0.9899
2. A. vera
80.3 ± 2.7*
0.54 ± 0.03*
y = 96.04x + 0.20
30.2 ± 2.1*
r2 = 0.9977
3. A. pillansii
54.7 ± 1.7*
1.38 ± 0.04*
y = 357.02x + 1.12
112.3 ± 8.6*
r2 = 0.9941
3. A. сongolensis
43.8 ± 2.9*
2.94 ± 0.04*
y = 732.87x - 1.63
230.2 ± 9.1*
r2 = 0.9984
3. A. marlothii
47.4 ± 3.2*
4.08 ± 0.05*
y = 996.65x + 2.11
310.4 ± 11.2*
r2 = 0.9983
Примечание. В таблице представлены: максимальная скорость окисления (Vm) на кривых, показанных на рис. 3; величины
[AO]o·f и уравнения регрессии у = аnх + b (n = 1-5) для зависимостей (τ - τ0)/τ0 от С (рис. 4); значения антиоксидантной
активности. Показатели Vm, АОА и [AO]o·f выражены как среднее ± стандартное отклонение; * - p < 0.05 по отношению к
показателям образца 1.
БИОФИЗИКА том 66
№ 3
2021
500
САЖИНА и др.
циент ингибирования, [AО]о - «эффективная» на-
чальная концентрация антиоксиданта в образце.
При [AAPH] = 0.33 мМ, Ri = = 0.45 нМ/с. Расчет
f⋅[AО]о для исследуемых экстрактов приведен в
таблице. Все величины выражены как средняя ве-
личина ± стандартная ошибка средней.
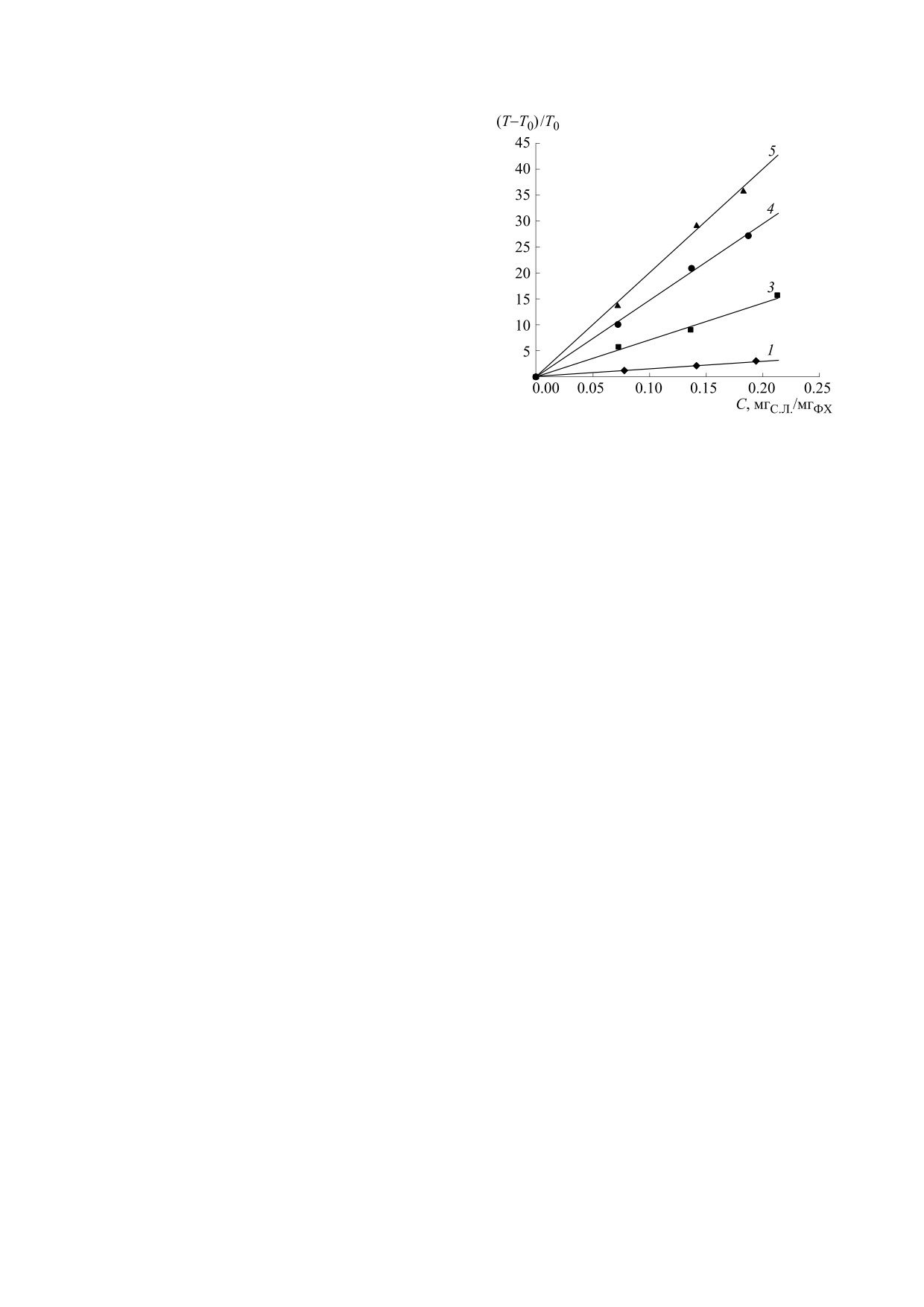
Зависимости параметра (τ - τ0)/τ0 от (С) пред-
ставлены на рис. 4.
В указанных пределах С эти зависимости
линейны (p < 0.01), а наклон их определяет инги-
бирующую (антиокислительную) активность
экстрактов Алое. Из этих зависимостей были по-
лучены соответствующие уравнения регрессии
(у = ах + b) для пяти образцов липосом (см. таб-
лицу).
Для выражения ингибирующей (антиокси-
Рис. 4. Зависимости (τ-τ0)/τ0 от удельного содержа-
дантной) активности экстрактов Алое в эквива-
ния сухих листьев в экстракте, введенном в липосо-
лентах тролокса (Trэкв) была проведена калибров-
мы, к массе ФХ в липосомах (С, мгС.Л./мгФХ) для: 1 -
ка с этанольным раствором тролокса (С = 1 мМ),
A. arborescens, 3 - A. pillansii, 4 - A. сongolensis, 5 -
A. marlothi. Для A. vera эта зависимость практически
вводимом в известных объемах (V) в липосомы.
совпадает с зависимостью 1, поэтому не приведена.
АОА исследуемых образцов определялась по ка-
либровочной прямой зависимости (τ-τ0)/τ0 для
стеме. Такая разница объясняется значительным
тролокса от его удельного содержания в ФХ-ли-
отличием в этих системах активности к захвату
посомах (С∙V, нмоль Tr/mФХ, мг). В таблице пред-
радикалов ДФПГ и ААРН разными фенольными
ставлены результаты определения АОА изучен-
метаболитами. На этих моделях определены так-
ных экстрактов в Trэкв, т.е. в нмоль Trэкв/мгC.Л.
же антирадикальные активности чистых вторич-
Как видно из таблицы, по значениям АОА ли-
ных фенольных метаболитов - алоина, алоэ-эмо-
дируют образцы 5 (A. marlothii) и 4 (A. сongolensis),
дина, алоэрезина, алоэзина А, алоэзона, алоэни-
они имеют антиокислительную активность в 13 и
на A. Для первого метода самыми активными
10 раз выше по сравнению с A. arborescens, однако
оказались алоэзон, алоэзин, алоэ-эмодин (351,
значение СФС у них только в 5-6 раз выше, чем у
343 и 222 мкМ Trэкв), для второго - алоэрезин А и
A. arborescens (рис. 2), а «эффективная» концен-
алоэ-эмодин (144 и 116 мкМ Trэкв), остальные ме-
трация f⋅[AО]о - в 9 и 6 раз. Для образца 3 (A. pil-
таболиты проявили в 2-30 раз меньшую антира-
lansii) эта разница сглаживается: СФС и АОА
дикальную активность [25]. Алоин, считающийся
больше этих величин для A. arborescens в 4.4 и в
главным антиоксидантом в Алое, показал в ДФ-
4.8 раза, причем спектр экстракта A. pillansii за-
ПГ-системе меньшую антирадикальную актив-
метно выше остальных, что свидетельствует о
ность (90 мкМ Trэкв), как и в системе ORAC
главном вкладе в АОА этого экстракта активных
(84 мкМ Trэкв) [25]. Масс-спектроскопический
фенольных соединений. Все это свидетельствует
о сложном антиоксидантном профиле в экстрак-
скрининг A. arborescens [25] и A. vera [25,33] вы-
тах этих растений и значительной разнице в АОА
явил такие классы антиоксидантов, как антроны
составляющих их фенольных компонентов.
(алоин, алоэсапонарин), пироны (алоэнин), хро-
моны (алоэзин, алоэрезин), флавоны, гидрокси-
В разных модельных системах фенольные ме-
кислоты (хлорогеновая, феруловая, кофейная)
таболиты Алое проявляют, как правило, неодина-
и др. [33].
ковую АОА. На модели со стабильным ДФПГ-ра-
дикалом и методом ORAC (oxygen-radical absorb-
Для нашей модельной системы в свободно-ра-
ing capacity) для экстрактов разных частей листьев
дикальных цепных реакциях инициированного
A. arborescens были определены значения антира-
окисления ненасыщенных липидных компонен-
дикальной активности [25, 30-32]. Для экстрак-
тов в бислое липосом кроме пероксидного ради-
тов из целых листьев A. arborescens значения анти-
кала (инициатора ААРН) образуются промежу-
радикальной активности составили 71 мкМ Trэкв
точные радикальные интермедиаты жирных кис-
с ДФПГ-радикалом и 2671 мкМ Trэкв в ORAC-си-
лот фосфолипидов (алкильные, пероксильные и
БИОФИЗИКА том 66
№ 3
2021
КИНЕТИКА ИНИЦИИРОВАННОГО ОКИСЛЕНИЯ ЛИПОСОМ
501
«чистых» липосом зависит, главным образом, от
условий их приготовления и жирнокислотного
состава используемого ФХ и уменьшается с уве-
личением в нем процентного содержания поли-
ненасыщенных ЖК [38]. С введением в липосо-
мы экстрактов Алое их жирорастворимые компо-
ненты встраиваются в различные области
липидного слоя липосом [35, 36, 39]. Они могут
взаимодействовать с группами липидных поляр-
ных головок на границе раздела «липид-вода»
мембран и защищать липидный бислой от агрес-
сивных молекул. Если повреждающая молекула
является окислителем, этот защитный эффект
может способствовать общему антиоксидантно-
му действию экстракта. «Ригидификация» липид-
ной мембраны компонентами экстракта может
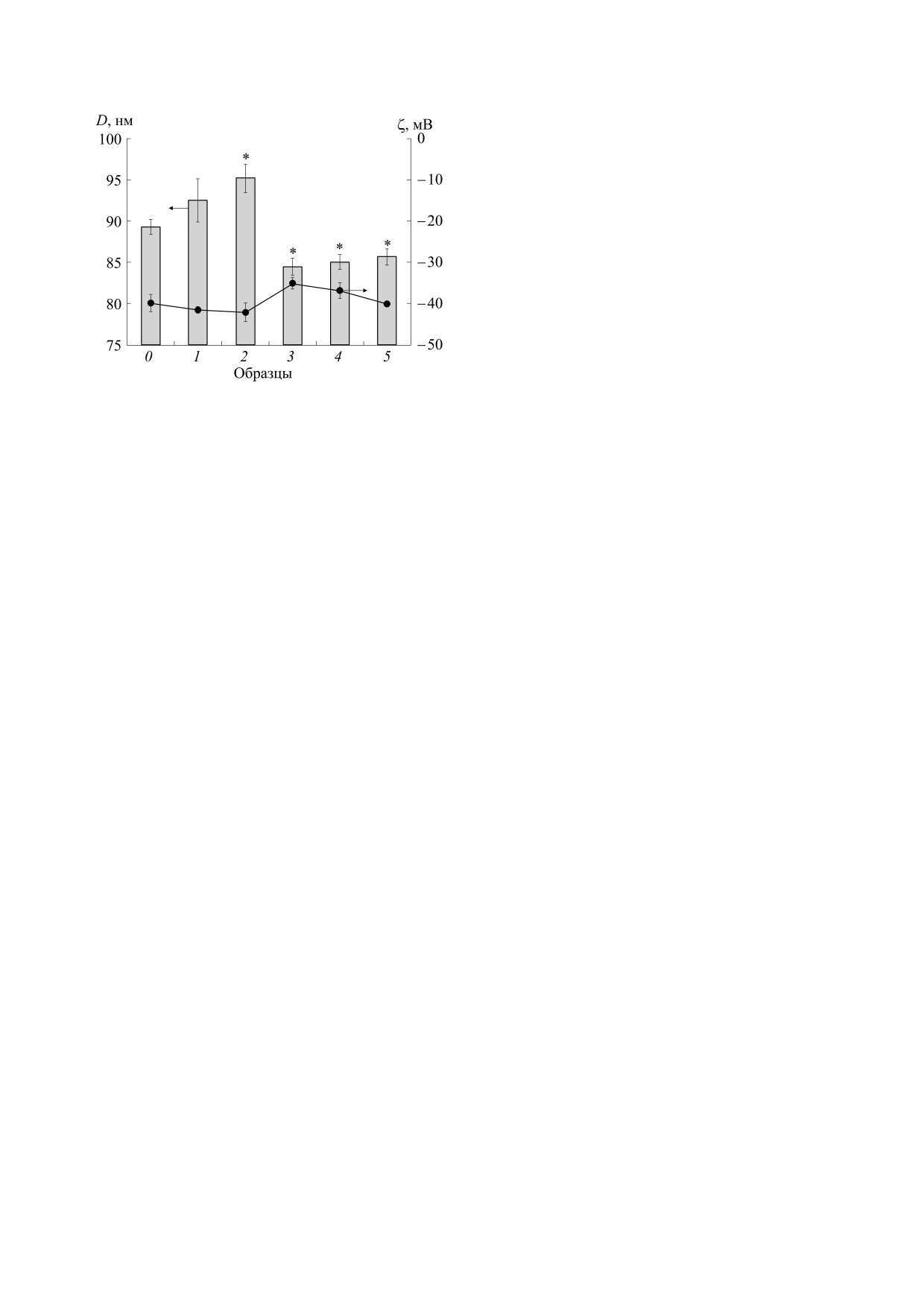
Рис. 5. Средний размер (D, показан столбиками на
диаграмме) и ζ-потенциал липосом: 0 - без экстрак-
препятствовать диффузии в нее кислорода и ра-
тов, 1 - A. аrborescens, 2 - A. vera, 3 - A. pillansii, 4 -
дикалов AAPH, снижая скорость окислительных
,
A. сongolensis, 5 - A. marlothii, С = 0.14 мгС.Л/мгФХ
реакций и повышая ингибирование окисления
[ФХ] = 0.5 мг/мл; * - p < 0.05 по отношению к
липидов. Возможно, с таким изменением струк-
образцу 0 .
туры липидного бислоя и связано уменьшение
среднего размера липосом, наблюдаемое нами
др. радикалы), с которыми взаимодействуют ан-
для экстрактов 3, 4 и 5 (рис. 5). Компоненты экс-
тиоксиданты экстрактов и обрывают цепь окис-
тракта 2, по-видимому, проявляют противопо-
ления [15, 16, 27, 28]. Подобные реакции происхо-
ложный эффект при встраивании в липидный
дят и в мембранах клеток живых организмов [34],
слой, как бы «разрыхляя» его и увеличивая размер
поэтому ингибирование окисления липосом ан-
липосом.
тиоксидантами при Т = 37°С дает более реальную
Уплотнение липидного бислоя активными
«физиологичную» модель для определения АОА
компонентами экстрактов 3 и 4 приводит к пере-
различных соединений. Наблюдаемое в нашей
стройке его структуры и, возможно, переориента-
работе значительное превышение АОА A. marlo-
ции и перегруппировке фосфолипидов в нем, что
thii и A. сongolensis над АОА A. arborescens и A. vera
приводит к уменьшению абсолютной величины
может свидетельствовать о наличии в листьях
отрицательного ζ-потенциала соответствующих
этих видов Алое фенольных метаболитов с высо-
липосом по сравнению с этим значением для «чи-
кой ингибирующей активностью в сложных сво-
стых» липосом [21]. Для экстрактов 1 и 2 наблю-
бодно-радикальных процессах, протекающих в
дается небольшое (≈5%) увеличение отрицатель-
процессе перекисного окисления липидов.
ного ζ-потенциала липосом, однако это только
Введение экстрактов Алое в липосомы приво-
тенденция, достоверного различия нет.
дит к перераспределению компонентов Алое в
разных частях липосомы: в липидном бислое ло-
ВЫВОДЫ
кализуются липофильные соединения, а в цен-
тральной части гидрофильные. Это приводит к
Проведено сравнительное исследование кине-
изменению структуры бислоя липосом и их физи-
тики ингибирующего действия этанольных экс-
ко-химических характеристик [35, 36], в частно-
трактов пяти видов Алое в модельной системе
сти размера и поверхностного заряда (ζ-потенци-
ААРН-индуцированного окисления липосом из
ала) [21, 37]. Эти характеристики для липосом с
ФХ и выявлены новые виды Алое: A. marlothii и
экстрактами Алое, введенными в липосомы,
A. сongolensis, обладающие, соответственно, в 13 и
представлены на рис. 5.
10 раз большей антиоксидантной активностью,
чем традиционные виды Алое: A. arborescens и
Можно отметить, что наиболее активные экс-
А. vera. Значения суммарного содержания фе-
тракты (экстракты 3, 4 и 5) достоверно уменьша-
нольных соединений в исследуемых экстрактах
ют средний размер липосом в сравнении с «чи-
Алое различаются только в 5-6 раз для A. arbo-
стыми» липосомами, в то время как экстракты с
rescens и A. marlothii и A. сongolensis, что может сви-
меньшей АОА (экстракты 1 и 2) увеличивают раз-
детельствовать о наличии в последних активных
мер липосом. Эта тенденция сохраняется и для
фенольных антиоксидантов в высоких концен-
других значений удельной концентрации. Размер
трациях. Проведен анализ спектров поглощения
БИОФИЗИКА том 66
№ 3
2021
502
САЖИНА и др.
изученных экстрактов на основе спектров погло-
13. A. H. Thomas, A. Catala, and M. Vignoni, Biochim.
щения основных индивидуальных фенольных
Biophys. Acta 1858, 139 (2016).
метаболитов Алое, который показал значитель-
14. M. Mosca, A. Cerlie, and L. Ambrosone, Chemistry
ную разницу в составе изученных видов Алое.
and Physics of Lipids 164, 158 (2011).
Установлено влияние введенных в липосомы
15. A. V. Lokhmatikov, N. Voskoboynikova, D. A. Chere-
экстрактов на их размер: наиболее «активные»
panov, et al., Oxidative Medicine and Cellular Longev-
экстракты A. marlothii, A. сongolensis и A. pillansii
ity 1, 1 (2016).
уменьшают средний размер липосом в сравнении
16. N. N Sazhina, A. S. Antipova, M. G. Semenova, and
с «чистыми» липосомами, а экстракты с меньшей
N. P. Palmina, Rus. J. Bioorg. Chem. 45 (1), 34 (2019).
АОА увеличивают, что, вероятно, связано с изме-
17. N. Kuznetsova, A. Kandyba, V. Vostrov, et al., J. Drug
нением липидной структуры липосом компонен-
Delivery Sci. Technol. 19 (1), 51 (2009).
тами экстрактов.
18. M. G. Semenova, Food Hydrocolloids 68, 114 (2017).
Результаты работы позволяют рекомендовать
19. M. G. Semenova, A. S. Antipova, L. E. Belyakova,
A. marlothii, A. сongolensis и A. pillansii для более
et al., Food Hydrocolloids 42, 149 (2014).
глубокого изучения их биологической активно-
сти и создания на их основе различных медицин-
20. M. H. Запрометов, Фенольные соединения и методы
ских препаратов.
их исследования. Биохимические методы в физиоло-
гии растений, (M.: Наука. 1971). 185.
21. H. H. Сажина, И. Г. Плащина, М. Г. Семенова и
КОНФЛИКТ ИНТЕРЕСОВ
Н. П. Пальмина, Коллоид. журн. 82 (1), 89 (2020).
Авторы заявляют об отсутствии конфликта
22. K. Дерффель, Статистика в аналитической химии
интересов.
(Мир, М., 1994).
23. Y. Gutterman and E. Chauser-Volfson, Biochemical
Systematics and Ecology 28 (9), 825 (2000).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ НОРМ
24. Д. Н. Оленников, И. Н. Зилфикаров и Т. A. Ибра-
Настоящая статья не содержит описания ка-
гимов, Химия растит. cырья, № 3, 77 (2010).
ких-либо исследований с использованием людей
25. L. Lucinia, M. Pellizzonia, R. Pellegrinob, et al., Food
или животных в качестве объектов.
Chemistry 170, 501 (2015).
26. Д. Н. Оленников, И. Н. Зилфикаров, Т. A. Ибраги-
СПИСОК ЛИТЕРАТУРЫ
мов и др., Химия растительного сырья, № 3, 83
1. B. Salehi, S. Albayrak, H. Antolak, et al., Int. J. Mol.
(2010).
Sci. 19, 2843. (2018).
27. P. A. C. McPherson, A. Bole, K. A. Cruz, et al., Chem.
2. M. Akaberi, Z. Sobhani, B. Javadi, et al., Biomedicine
Phys. Lipids 165, 682 (2012).
& Pharmacotherapy 84, 759 (2016).
28. I. Pinchuk and D. Lichtenberg, Chem. Phys. Lipids
3. D. I. Sánchez-Machado, J. López-Cervantes, R. Sen-
178, 632014 (2014).
dón, A. Sanches-Silva, Trends Food Sci. Technol. 61,
29. E. T. Denisov and I. B. Afanas’ev, Oxidation and Anti-
94 (2017).
oxidants in Organic Chemistry and Biology (Boca Raton:
4.
4. E. Misawa, M. Tanaka, K. Nomaguchi, et al., Agric.
CRC Press. 2005).
Food. Chem. 60, 2799 (2012).
30. A. Seyoum, K. Asres, and F. K. El-Fiky, Phytochemis-
5. I. Cock, In Novel Natural Products: Therapeutic Effects
try 67, 2058 (2006).
in Pain, Arthritis and Gastro-intestinal Diseases, Ed. by
31. H. Hu, X. Hu, and H. Qiuhui, J. Agricul. Food Chem.
K. Rainsford, M. Powanda, and M. Whitehouse
51, 7788 (2003).
(Springer, Basel, 2015), pp. 179-235.
32. D. Huang, B. Ou, M. Hampsch-Woodil, et al., J. Agri-
6. N. Ghannam, M. Kingston, I. A. Al-Meshaal, et al.,
cul. Food Chem. 50, 4437 (2002).
Horm. Res. Pediatr. 24, 288 (1986).
33. S. Lee, S. G. Do, S. Y. Kim, et al., J. Agricul. Food
7. R. Lawrence, P. Tripathi, and E. Jeyakumar, Braz. J.
Chem. 60, 11222 (2012).
Microbiol. 40, 906 (2009).
34. Н. М. Эмануэль, Е. Т. Денисов и З. К Майзус, Цеп-
8. Q. Pan, H. Pan, H. Lou, et al., Cancer Cell Int. 13, 69
ные реакции окисления углеводородов в жидкой фазе,
(2013).
(Наука, М., 1965).
9. S.-Y. Lin, W.-W. La, C.-C. Ho, et al., Anticancer Res.
35. N. P. Palmina, E. L. Maltseva, V. I. Binjukov, et al.,
29, 327 (2009).
Biophysics 63 (1), 53 (2018).
10. M.E. Clement, G.Tringali, D. Triggiani, B. Giardina,
36. A. R. Neves, C. Nunes, H. Amenitsch, and S. Reis,
Nat. Prod. Commun. 10 (11), 1993 (2015).
Soft Matter 12, 2118 (2016).
11. H. H. Сажина, П. В. Лапшин и Н. В. Загоскина,
37. S. Gal1, L. Pinchuk, and D. Lichtenberg, Chem. Phys.
Химия растит. сырья, № 2, 169 (2015).
Lipids 126, 95 (2003).
12. N. P. Palmina, E. L. Maltseva, V. I. Binjukov, et al.,
38. E. A. Tehrany, C. J. F. Kahn, C. Baravian, et al., Col-
Biophysics 63 (1), 53 (2018).
loids Surfaces B: Biointerfaces 95, 75 (2012).
БИОФИЗИКА том 66
№ 3
2021
КИНЕТИКА ИНИЦИИРОВАННОГО ОКИСЛЕНИЯ ЛИПОСОМ
503
39. P. Strugała, S. Cyboran-Mikołajczyk, A. Dudra, et al.,
J. Membrane Biol. 249, 393 (2016).
Kinetics of Initiated Oxidation of Phosphatidylcholine Liposomes with Introduced Aloe
Extracts and Determination of Their Antioxidant Activity
N.N. Sazhina*, P.V. Lapshin**, and N.V. Zagoskina**
*Emanuel Institute of Biochemical Physics, Russian Academy of Sciences, ul. Kosygina 4, Moscow, 119334 Russia
**Timiryazev Institute of Plant Physiology, Russian Academy of Sciences, Botanicheskaya ul. 35, Moscow, 127276 Russia
Numerous species of the genus Aloe have attracted the attention of researchers because of different biological
benefits and their capacity to act as antioxidants. This paper presents a comparative study of the kinetics of
the inhibitory effect of ethanol extracts of five species of Aloe which was made using a model system of the
oxidation of phosphatidylcholine liposomes. It was found that plant extracts of A. marlothii and A. congolensis,
have higher antioxidant activity (by a factor of 13 and 10, respectively) than the best known Aloe species:
A. arborescens and A. vera. The total phenolic content values of A. marlothii and A. congolensis are higher than
those of A. arborescens and A. vera but in a lesser degree (by a factor of 5-6) than antioxidant activity was.
This may indicate the presence of very active phenolic antioxidants in A. Marlothii and A. congolensis. Anal-
ysis of the influence of extracts introduced into liposomes on the liposome size showed that the most active
extracts of A. marlothii, A. congolensis and A. pillansii reduce the average liposome size compared to pure li-
posomes, and extracts with weaker antioxidant activity increase it, what is probably due to changes in the lipid
structure of liposomes by the components of extracts. Based on the results obtained, A. marlothii, A. congo-
lensis, and A. pillansii are suitable for studying other types of their biological activity that might contribute to
new drug development.
Keywords: Аloe, phenolic metabolytes, kinetics, oxidation, liposomes, phosphatidylcholine
БИОФИЗИКА том 66
№ 3
2021