БИОФИЗИКА, 2021, том 66, № 3, с. 611-624
ДИСКУССИИ
УДК 577.35
ГИПОТЕЗА ОБ ЭНТРОПИЙНОМ ИНВАРИАНТЕ
ДЛЯ БИОЛОГИЧЕСКИХ ОРГАНИЗМОВ
© 2021 г. В.Т. Волов
Самарский государственный университет путей сообщения, 443110, Самара, ул. Свободы, 2в
E-mail: vtvolov@mail.ru
Поступила в редакцию 20.12.2019 г.
После доработки 18.01.2021 г.
Принята к публикации 22.01.2021 г.
Представлены обзор исследований, посвященных проблеме собственного (физиологического) вре-
мени биологических организмов и интегральных характеристик их жизнедеятельности, и результа-
ты оригинальных исследований автора. Сформулирована гипотеза об инвариантности вновь вве-
денной интегральной характеристики жизни организма - функционала действия энтропии. Со-
гласно данной гипотезе, значение функционала действия энтропии является одинаковым для всех
организмов конкретного вида. Базируясь на предложенной гипотезе, выявлена связь между соб-
ственным и хронологическим временами жизни организма. Полученная формула этой связи вклю-
чает фундаментальную функцию теории структуры И. Пригожина - второй дифференциал энтро-
пии. Показано, что собственное время жизни организма имеет фрактальную структуру. Предложе-
на формула математической меры - детерминированной энтропии метаболизма организма,
позволившая осуществить проверку выдвинутой гипотезы для некоторых видов организмов. Об-
суждаются возможные перспективы тестирования предложенной гипотезы для организмов.
Ключевые слова: константа Рубнера, энтропийный инвариант, собственное (физиологическое) время
организма, функционал действия энтропии, второй дифференциал энтропии.
DOI: 10.31857/S0006302921020236
В фундаментальных работах И. Пригожина [1,
Необходимо отметить, что проблема собствен-
2, 3] разработана теория структуры или неравно-
ного (индивидуального) времени развития орга-
весной термодинамики, ставшая основой про-
низмов имеет довольно длинную историю в био-
рывного направления в науке конца ХХ-начала
логической науке, начиная с классических работ
ХХI века - синергетики. Преимуществом термо-
М. Рубнера [5] и его последователей [6-10], а так-
динамического подхода И. Пригожина является
же работ, представленных в исследованиях конца
отсутствие необходимости эмпирического опре-
XX - начала XXI веков [11-18], где, в частности,
деления параметров порядка системы [4], по-
предпринимались попытки термодинамического
скольку в данном подходе используется универ-
подхода решения данной проблемы. Осознание
сальный аппарат термодинамики (II начало тер-
того, что живой организм существует в собствен-
модинамики, теорема о минимуме производства
ном времени, скорость течения которого зависит
энтропии, понятие «локального равновесия»).
от различных процессов, происходящих в орга-
Главной причиной, побудившей И. Пригожи-
низме, пришло достаточно давно. Одна из первых
на заняться разработкой «теории структуры», бы-
попыток формализации понятия собственного
ло, с его точки зрения, противоречие, заключав-
времени организма была предпринята Г. Бакма-
шееся в отсутствии в термодинамике такой важ-
ном [7]. В качестве определяющего процесса Бак-
нейшей величины, как время, с одной стороны, и
ман выделил процесс роста организма. Собствен-
противоречие со II началом термодинамики фак-
ное время организма по Бакману могло прини-
та существования организмов - с другой сторо-
мать как положительные, так и отрицательные
ны. Однако, как известно, время жизни организ-
значения, а в момент достижения половой зрело-
мов варьируется в достаточно широком диапазо-
сти становилось равным нулю. Функция роста
не, т.е. продолжительность жизни организма
массы организма также была принята за основу
конкретного вида может существенно отличаться
при определении собственного времени в работах
от максимального значения в популяции. Данное
[18, 19].
обстоятельство побудило автора статьи преодо-
В работах [14-21] дано определение единицы
леть данное противоречие.
собственного времени организма, которая опре-
611
612
ВОЛОВ
деляется как время удвоения массы и растет с уве-
tβ
личением общей массы организма.
Physiological time =
q(t)dt
,
(2)
∫
Необходимо подчеркнуть, что собственное
t
α
время, определенное в работе [18] как τ(t) =
где tα и tβ - соответственно начало и конец рас-
w(t)/W’(t), имеет место только для организмов не
имеющих границ роста.
сматриваемого периода. Физиологическому вре-
мени (2) придается размерность ЕМ-1 (выражает-
В исследовании [20] в качестве меры собствен-
ся в ккал/г сухой массы, понимаемой как соот-
ного времени был принят период эмбрионально-
ветствующее
количество
единиц
го развития - интервал времени между появлени-
физиологического времени).
ем одноименных фаз митоза двух последователь-
ных делений дробления.
Определение
Базируясь на предложенной Рубнером гипоте-
собственного (физиологического) времени, ос-
зе о приблизительно равном количестве энергии,
нованное на процессе метаболизма, предложил
используемом единицей массы взрослого живот-
Дж. Райс [21]. Он в качестве основы собственного
ного за период жизни от момента прекращения
(физиологического) времени взял введенное М.
роста до смерти для некоторых видов млекопита-
Боддингтоном [22] понятие «показатель абсолют-
ющих, Э. Бауэр [6] распространил это равенство
ного метаболизма» (absolute metabolic scope) -
потребления энергии на единицу массы живот-
константу, равную произведению максимальной
ного на весь период жизни и на животных других
уровней организации. Данную количественную
длительности жизни (пропорциональной W1/4) и
меру Бауэр [6] назвал константой Рубнера, кото-
скорости базального метаболизма, деленному на
рая определялась как произведение основного
единицу массы (W-1/4, где W - масса взрослого
обмена за сутки на продолжительность жизни в
организма (достигшего границы роста). Данная
сутках, деленное на массу организма. Тем не ме-
константа представляет собой минимальное ко-
нее позже Бауэр подтвердил неправомочность
личество энергии, потребленное единицей
расширения предложенного понятия и предпо-
массы организма за время жизни, и не зависит от
ложил, что константа Рубнера является критери-
значения W. Данная константа имеет размер-
ем принадлежности животного к определенному
ность ЕМ-1Т-1. Пренебрежение временем разви-
уровню эволюционного развития и определяется
тия организма от момента зарождения до момен-
приблизительно одинаковой свободной энергией
та достижения массы W приводит к тому, что по-
зародышевых клеток внутри него. С ростом слож-
казатель абсолютного метаболизма оказывается
ности организации животных в процессе эволю-
равным константе Рубнера и может служить, со-
ции данная константа увеличивается.
гласно работе [21], инвариантом максимального
Анализ научной литературы по данному во-
возраста.
просу приводит к выводу, что в большинстве слу-
Недостатком введенного Боддингтоном «по-
чаев этому понятию придается не тот смысл, что
казателя абсолютного метаболизма» было то, что
оно имело первоначально. В биологической ли-
показатель не учитывал изменения массы и ско-
тературе конца ХХ - начале ХХI века часто кон-
рости удельного метаболизма организма в онто-
стантой Рубнера называют суммарный удельный
генезе, в частности, в раннем возрасте скорость
метаболизм за весь период жизни животного [11-
метаболизма выше, чем в остальное время жизни.
19]. Например, в исследованиях [18, 19], базиру-
Учитывая данный факт, Райс ввел понятие
ясь на вышеотмеченных работах [6, 21], констан-
«удельный метаболизм за время жизни» и опреде-
той Рубнера называют следующую величину:
лил его следующим образом:
t
max
t
max
q t)
= ∫ qt)dt
,
(3)
t
max
0
q(t)dt
Specific limetime metabolism =
∫
,
(1)
где
q
−
усредненный удельный метаболизм за
t0
]. Принимается, что ве-
весь период жизни [0,tmax
где q(t) = Q(t)/w(t) - удельная скорость потребле-
личина (3) может служить характеристикой вида,
ния энергии (кислорода), Q(t) - потребление
т.е. имеет определенное значение для каждого ви-
энергии организмом в единицу физического вре-
да. Отсюда следует, что имеет место существен-
мени, w(t) - значение массы организма в текущий
ное отличие от определения, данного Рубнером:
момент времени, tmax и t0 - продолжительность
а) константой Рубнера называют не суммарное
жизни и начальное время организма соответ-
количество энергии, использованное единицей
ственно.
активной массы организма за всю жизнь от мо-
Вводя гипотезу о пропорциональности скоро-
мента прекращения роста до смерти, а суммар-
сти физиологического времени и удельного мета-
ный метаболизм единицы массы организма за
болизма, Боддингтон дал его определение [21]:
всю жизнь (Еmax); б) считается доказанным по-
БИОФИЗИКА том 66
№ 3
2021
ГИПОТЕЗА ОБ ЭНТРОПИЙНОМ ИНВАРИАНТЕ
613
i
i
i
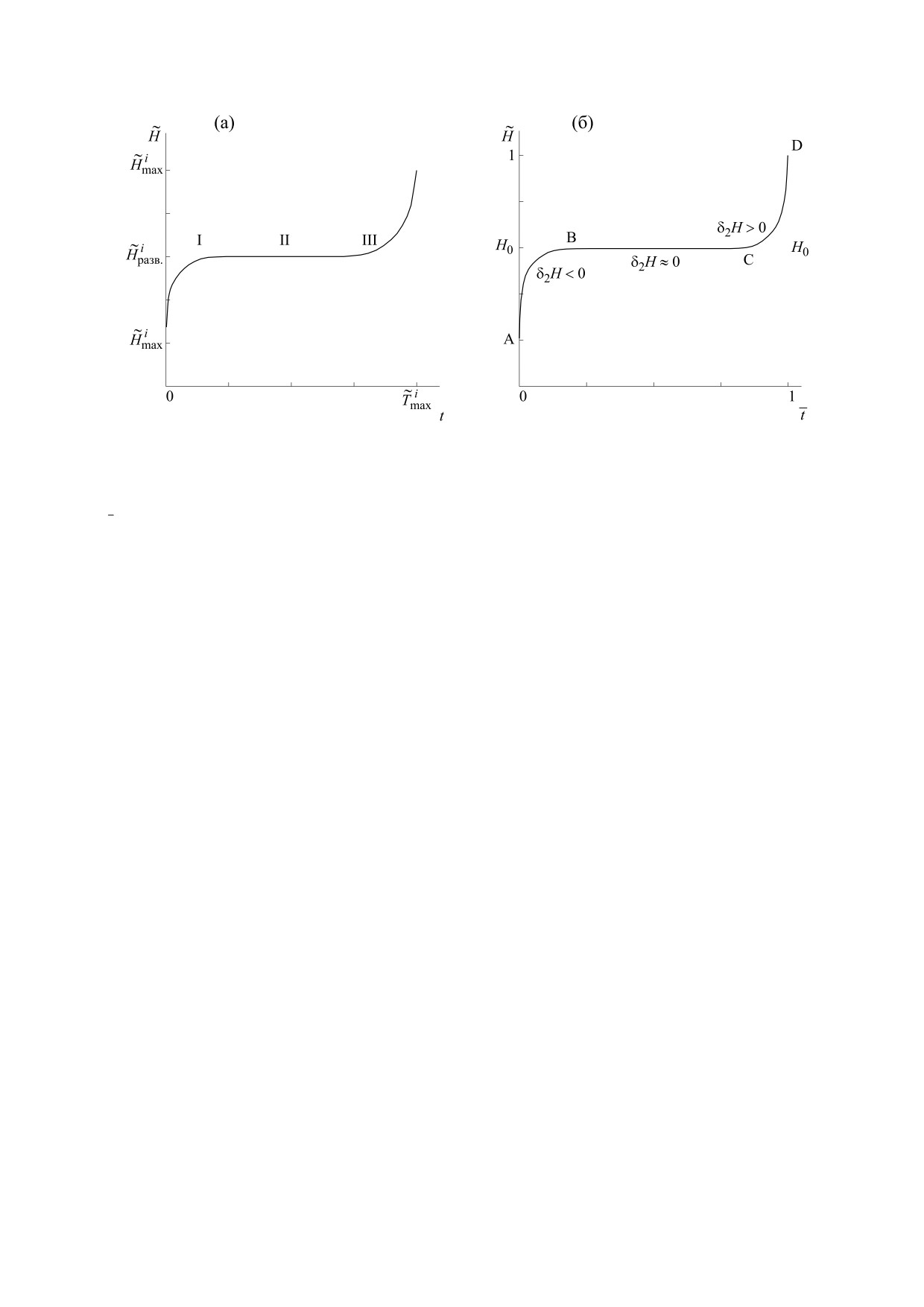
Рис. 1. Иллюстрация эталонного тренда энтропии в течении жизни организма: (а) -
-
N N,N ,
максимальное и минимальное значения энтропии, а также значение энтропии развитого организма i-го вида
i
соответств енно,
- фактическое время жизни i-го вида организма; (б) - H0 - энтропия «золотого сечения»,
fact
T
нормированное время организма.
t
-
стоянство величины Еmax для вида. Между тем
щихся систем вдали от равновесия необходима
tmax
информация из дополнительных эмпирических
данных по квадратичной форме - производству
Рубнер показал, что величину
qt)dt
можно
∫
избыточной энтропии. Данное противоречие
tA
преодолевается в цитируемой выше литературе с
считать постоянной только при достаточно боль-
шом разбросе конкретных значений.
помощью фрактально-кластерной теории [23-
25], разработанной для самоорганизующихся си-
Резюмируя вышеизложенное, можно заклю-
стем класса «организм». В статье используются
чить, что, как отмечено в работе [18], постулиро-
эквивалентные понятия «собственное» и «физио-
вание постоянства «константы Рубнера» для кон-
логическое» время развития организма.
кретного вида организмов, в том числе пойкило-
термных животных, например для моллюсков и
рыб, не имеющих границы роста [17], представля-
ФУНКЦИОНАЛ ДЕЙСТВИЯ ЭНТРОПИИ
ется необоснованным.
И ЕГО ИНВАРИАНТНОСТЬ
В представленной статье вводится новая инте-
Смысл определяемого нами функционала дей-
гральная характеристики жизни организма - так
ствия энтропии заключается в следующем. Допу-
называемый функционал действия энтропии и
стим, что имеется тренд энтропии развития орга-
выдвигается гипотеза его инвариантности для ор-
низма, представленной на рис. 1. В контексте на-
ганизмов конкретного вида. На основе данной
стоящего исследования, энтропия представляет
гипотезы определяется собственное (физиологи-
ческое) время жизни организма и устанавливает-
собой математическую меру, отвечающую прави-
лам построения теории математической энтро-
ся связь между собственным и хронологическим
пии [8]. Представленная математическая мера
временем. Базируясь на предложенной гипотезе
и фундаментальном аппарате неравновесной тер-
определяет интегральную характеристику состоя-
модинамики
[1-4], а также инструментарии
ния всего организма. Этой измеряемой характе-
фрактально-кластерной теории [23-25], закла-
ристикой может быть, например, метаболизм,
дываются основы термодинамической теории
рост организма (для организмов, не имеющих
эволюции организма. Следует отметить, что ап-
границ роста), водно-белковый, кислотно-ще-
парат линейной термодинамики неравновесных
лочной баланс, липидный обмен, иммунный ста-
процессов И. Пригожина предназначен для ис-
тус, гормональный фон и т.д., а также нормиро-
следования самоорганизующихся систем, нахо-
ванная мера всех вышеперечисленных характери-
дящихся вблизи положения равновесия [1-3], в
стик организма. Из данной иллюстрации следует,
то время как для исследования самоорганизую-
что имеется три периода: первый - устойчивый
БИОФИЗИКА том 66
№ 3
2021
614
ВОЛОВ
2
2
~
рост
(
δ
H >
0
)
; второй - гомеостаз (δ
H ≈
0) и
В дальнейшем мы будем обозначать через
H
i
третий - неустойчивое развитие (старение) «ор-
энтропию, относящуюся к определенному i-му
2
~
ганизма» (δ
H <
0). Эталонный тренд эволюции
виду организмов. Величину
H
i
, удовлетворяю-
энтропии (рис. 1) согласуется, во-первых, со вто-
щую свойствам
1-5, удобно нормировать:
рым началом термодинамики для необратимых
H
i
H
=
,
неравновесных процессов
(dH / dt ≥ 0), во-вто-
i
max
H
i
рых, с фундаментальной теоремой неравновес-
ной термодинамики о минимуме производства
~
max
где
H
- максимальное значение энтропии ор-
энтропии [1-3] и, в-третьих, с опытом жизнедея-
i
тельности самоорганизующихся систем различ-
ганизма данного типа, и считать, что
0≤
H
≤
1
i
ной природы - биологическим, социально-эко-
Введем новую физическую величину - функ-
номическим и др. [23-25]. Производство энтро-
ционал действия энтропии - для самоорганизую-
пии (P = dH/dt), согласно эталонному тренду, на
щейся системы, определяемую следующим обра-
этапе развития (I) уменьшается до нуля (II этап),
зом:
а затем начинает экспоненциально возрастать
(III этап). Согласно положениям неравновесной
t
2
термодинамики, I этап соответствует уменьше-
action
F
= ∫H
dt,
(4)
H
i
i
нию диссипативных процессов в системе, второй
t1
этап - балансу регенеративных и диссипативных
процессов, а III этап
- интенсивному
где t1 и t2 - начальное и конечное время ее разви-
увеличению диссипаций в системе, которые в ко-
~
тия;
H
- размерное текущее значение энтропии
нечном результате приводят к ее полному разру-
i
шению.
определенного организма.
Например, для человека энтропийный тренд
Предлагаемая гипотеза об энтропийном инва-
(рис. 1) можно интерпретировать следующим об-
рианте для организмов заключается в постулиро-
разом: первый этап связан с энергией роста ре-
вании инвариантности функционала действия
бенка, второй - энергетический баланс между ре-
action
энтропии
F
:
H
генеративными и диссипативными процессами
i
i
организма, а третий - например, с энергией роста
T
fact
T
max
ideal
патологических образований, приводящих в ко-
H t)dt =
i
H t)dt = const ≡ γ
i
,
(5)
∫
i
∫
нечном счете к его смерти.
0
0
Согласно общему определению энтропии [26,
~
где
t
- тренд энтропии конкретного орга-
27], энтропия «организма» есть функция
H
его
i
состояния, которая удовлетворяет следующим
низма из выбранного
i -го вида как функции вре-
свойствам:
fact
мени t,
T
i
- фактическое время жизни организ-
N
ideal
ма,
N t)
- идеальный (эталонный) тренд эн-
i
1) Аддитивность
H
H
, т.е. энтропия си-
l
≥∑
max
l=1
тропии,
T
- идеальное (максимальное)
i
стемы не может быть меньше суммы энтропий
фактическое время жизни организма. Соотноше-
подсистем, из которых она состоит (здесь знак
ние (5) представляет константу для конкретного
«>» относится к случаю взаимозависимости под-
-го вида организма.
i
систем).
~
Согласно свойству 2) энтропии имеем
2) Приращение энтропии
dH
есть сумма ее
inner
приращения за счет взаимодействия с внешними
dH
= dH
+ dH
ext.
(6)
i
i
i
~
системами
dH
ext и ее приращения за счет внут-
inner
Здесь внутреннее изменение энтропии
dH
~
i
ренних изменений в системы
dH
inner:
обусловлено естественным старением системы, а
ext
inner
ext
ее внешнее изменение
dH
связанно с взаимо-
dH = dH
+ dH
i
действием с внешней средой (другими система-
3) Внутреннее приращение энтропии всегда
ext
ми);
dH
i
может иметь как положительное, так и
~
неотрицательно, т.е.
dH
inner ≥ 0.
отрицательное значение. Ниже мы будем иссле-
~
~
~
довать только случай слабой зависимости
4) Ограниченность
∃Н
>
0 :
H ≤
Н
max
max
ext
H
i
inner =
fH
i
). Идеальный или эталонный тренд
ext
5) Неотрицательность
H
0
энтропии (рис. 1) реализуется при
dH
→ min.
i
БИОФИЗИКА том 66
№ 3
2021
ГИПОТЕЗА ОБ ЭНТРОПИЙНОМ ИНВАРИАНТЕ
615
ideal
fact
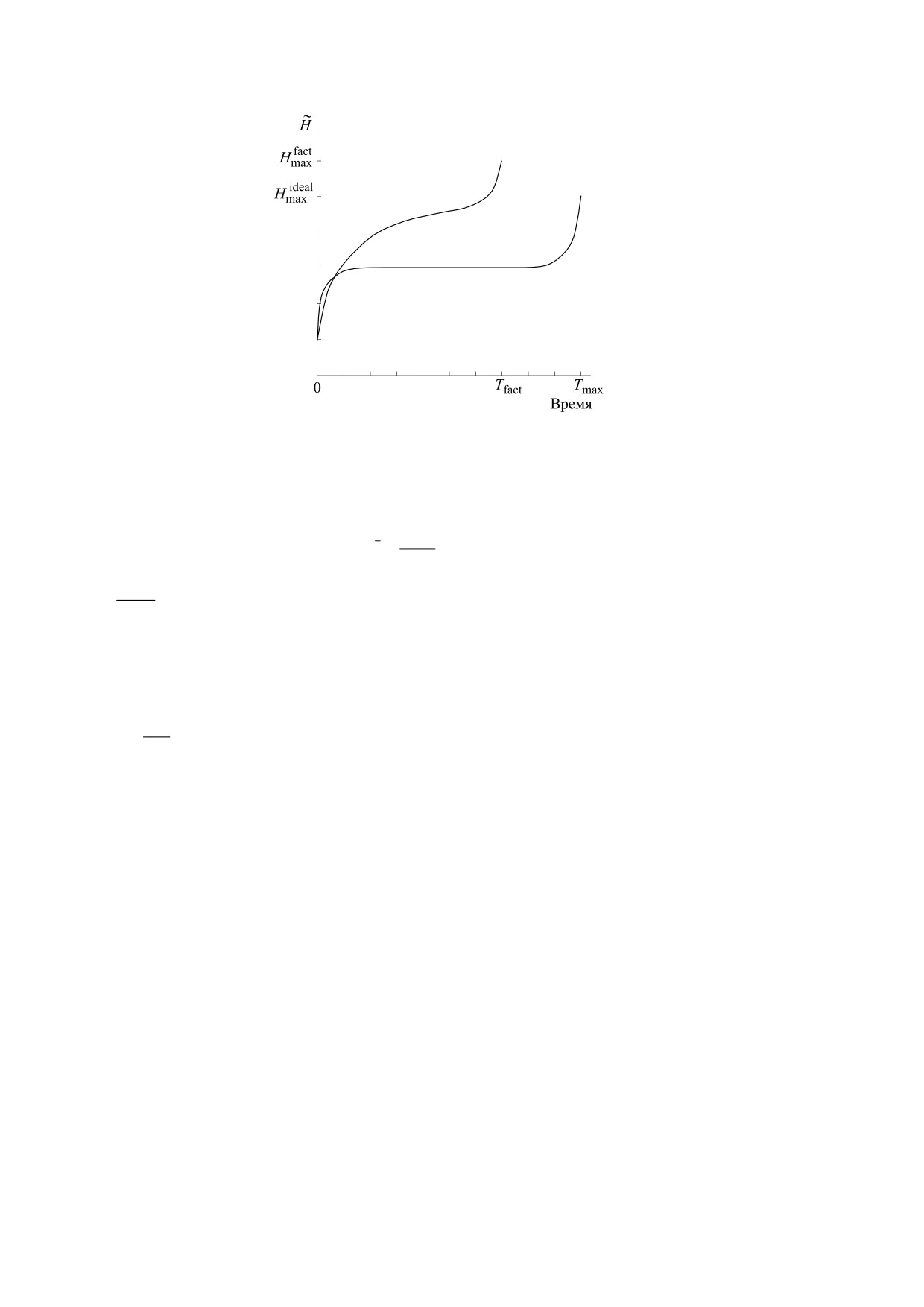
Рис. 2. Иллюстрация гипотезы о постоянстве функционала действия энтропии для организма.
H
max
,
H
max
-
максимальное значение идеального и фактического трендов соответственно; Tmax, Tfact - максимальное и
фактическое время жизни организма конкретного вида соответственно.
отрезка пополам (0.5/0.5), и «золотое сечение»
t
Введя безразмерные обозначения
t
=
и
(0.38/0.62). Таким образом, у живых существ про-
max
T
i
блема «золотого сечения» четко коррелирует с
ideal
обобщенной проблемой устойчивости (гомеоста-
H
i
β =i
, уравнение (5) можно переписать в
за) организмов. Не случайно в этой связи была
ideal
H
max
разработана математическая теория «золотого се-
виде:
чения» [27].
ξ
i
1
В соответствии с теоремой о среднем для мо-
max
ideal
ideal
H
H
dt
=
H
H
dt
,
(7)
нотонных функций, принципом симметрии
i
∫
i
imax
∫
i
0
0
(ΔАВH
≈
ΔCH D)0и принципом «золотого сече-
0
fact
ния» [28] интеграл (7) будет равен:
i
T
где
ξ
i
=
. В дальнейшем при рассмотрении
max
1
T
ideal
i
H
dt ≡H
≈ 0.618.
(8)
конкретного организма мы будем опускать ин-
∫
0
0
декс i.
При расчете интеграла (7) по эталонному трен-
2. СВЯЗЬ ИНДИВИДУАЛЬНОГО ВРЕМЕНИ
ду энтропии (рис. 1б) был использован принцип
ОРГАНИЗМА С ХРОНОЛОГИЧЕСКИМ
симметрии и принцип «золотого сечения» [27].
ВРЕМЕНЕМ
Проблеме взаимосвязи
«золотого сечения» и
строения и функционирования организмов, на-
Для организма, находящегося в реальных
чиная с Древней Греции и в эпоху Возрождения,
условиях, фактическое время жизни T будет от-
посвящено значительное количество публикаций
личаться от максимального времени жизни орга-
и исследований. Люди находили «золотое сече-
низма Tmax, соответствующего идеальному (эта-
ние» в различных пропорциях живых организмов
лонному) тренду (рис. 2). Если перенести данное
[24, 27, 28]. Эта пропорция проявилась не только
утверждение на человека, то можно представить
в пропорциях организма, но и в функциональных
следующую ситуацию: в горах Азербайджана, где,
характеристиках организмов (нормативы артери-
как известно, живут долгожители, родились близ-
ального давления (80/120 мм рт. ст.), соотноше-
нецы. Один из них остался жить в горах, в эколо-
ние осевой и вращательной составляющих крово-
гически чистом месте, дышал ионизированным
тока в сосудах и так далее). Существует мнение,
горным воздухом, питался натуральными про-
отраженное в различных научных публикациях,
дуктами, выращенными в горах, не имел вредных
что наличие «золотого сечения» во множестве на-
привычек и так далее и, как результат, дожил до
блюдаемых в природе фактов отнюдь не случай-
своих 120 лет. Другой близнец спустился с гор в
но, а закономерно. Как известно, существуют не-
город, где экологическая обстановка была далека
сколько видов баланса: дихотомия, т. е. деление
от международных стандартов, питался продук-
БИОФИЗИКА том 66
№ 3
2021
616
ВОЛОВ
тами с разнообразными добавками, имел весь
тропийного функционала (действия энтропии)
спектр вредных привычек и постоянные стрессы
была сформулирована для всего времени жизни
и, как результат, дожил до 69 лет. Налицо реали-
организма. Требование инвариантности действия
зация, так называемого, принципа «шагреневой
энтропии для любого временного интервала при-
кожи» В данном случае «аналогом» «шагреневой
водит к появлению новой временной шкалы -
кожи» является интегративная мера состояния
шкалы индивидуального (или собственного) вре-
action
мени θ. А именно, мы потребуем, чтобы для лю-
здоровья - это энтропийный инвариант
F
-
Н
бого временного отрезка [t1, t2] существовал такой
энтропия действия. Увеличение энтропии, адап-
отрезок [θ1, θ2] на оси индивидуального времени,
тированной организмом, приводит к сокраще-
что
нию длительности жизни.
Однако в силу свойства ограниченности
θ
2
t
2
условной энтропии (4) выполнение гипотезы об
H
(θ')d
θ'
=β
H
dt
∫
∫
0
энтропийном инварианте на всем протяжении
θ
1
t
1
жизни организма может реализовываться только
Это означает, что имеется следующее биектив-
на определенном интервале шкалы хронологиче-
ное отображение между переменными хроноло-
ского времени:
гического t и индивидуального времени θ, кото-
H0⋅Tmax < Tfact < Tmax.
рое выражается следующей формулой:
θ
При временах жизни организма Tfact, меньших
значения H0⋅Tmax, для выполнения гипотезы эн-
H
(
')
θ θ'
=β
H
0
Δt
(11)
∫
0
тропийного инварианта необходимо ввести но-
вую шкалу времени (индивидуальное время орга-
и которое в дифференциальной форме прини-
низма) описываемую переменной θ, которая бу-
мает вид:
дет отличаться от шкалы хронологического
H
(θ
)
времени (переменная t). Индивидуальное время
dθ
=
dt
(12)
всей жизни организма будем обозначать как Θ.
βH
0
Оно определяется требованием выполнения сле-
Разобьем временной отрезок [0,Δt] на конеч-
дующей гипотезы:
ные временные шаги величиной Δtj, где j - номер
Θ
T
max
шага. Такому разбиению будет соответствовать
ideal
H
(τ)d
τ=
H t)dt
(9)
разбиение интервала [0, θ] на отрезки величиной
∫
∫
0
0
Δθj. Используя теорему о среднем и разлагая с
Из условия ограниченности энтропии (4) и
точностью до членов второго порядка в ряд Тей-
выполнения энтропийного инварианта следует,
лора значение энтропии H(θ′) на отрезке [0, θ] от-
что индивидуальное время организма имеет
носительно ее равновесного значения Hj на j-м
фрактальную структуру. Под термином «фрак-
временном отрезке после интегрирования в урав-
тальная» структура понимается дробная степен-
нении (11), получаем:
ная зависимость между физическими величина-
Δθj = WjΔtj,
(13)
ми в исследуемом процессе или явлении. Из со-
отношения (10) при предположении равенства
β
H
0
/H
j
максимальной продолжительности жизни орга-
гдеW
j
(H)
≡
, Hj - равновесное зна-
2
низма в собственном (Θ) и хронологическом (фи-
δ
H
j
δ
H
j
1+
+
зическом) времени Tmax следует D
= Θ, где
H
j
H
j
fact
D ≥ 1. Величина D может быть интерпретирована
чение энтропии на j-м отрезке, δHj и δ2Hj - чле-
как формальный аналог фрактальной размерно-
ны, пропорциональные значениям первого и
сти собственного времени жизни организма и
второго дифференциала тренда энтропии. Дина-
определена из уравнения
мика изменения численного значения второго
D
дифференциала тренда энтропии определяет
T
fact
T
max
устойчивость функционирования организма и в
ideal
H τ)dτ=β ∫ H
t)dt = βH
T
,
(10)
∫
0
max
соответствии с предложенной гипотезой энтро-
0
0
пийного инварианта показывает отличие его соб-
ideal
ственного (физиологического) времени от хроно-
H
max
где
β=
логического.
H
max
Соотношение (13) позволяет определить за-
Возникает вопрос: как связано текущее инди-
медление или ускорение индивидуального време-
видуальное время в жизни организма и хроноло-
ни организма по сравнению с хронологическим
гическое время? Гипотеза об инвариантности эн-
временем как функцию энтропийного фактора.
БИОФИЗИКА том 66
№ 3
2021
ГИПОТЕЗА ОБ ЭНТРОПИЙНОМ ИНВАРИАНТЕ
617
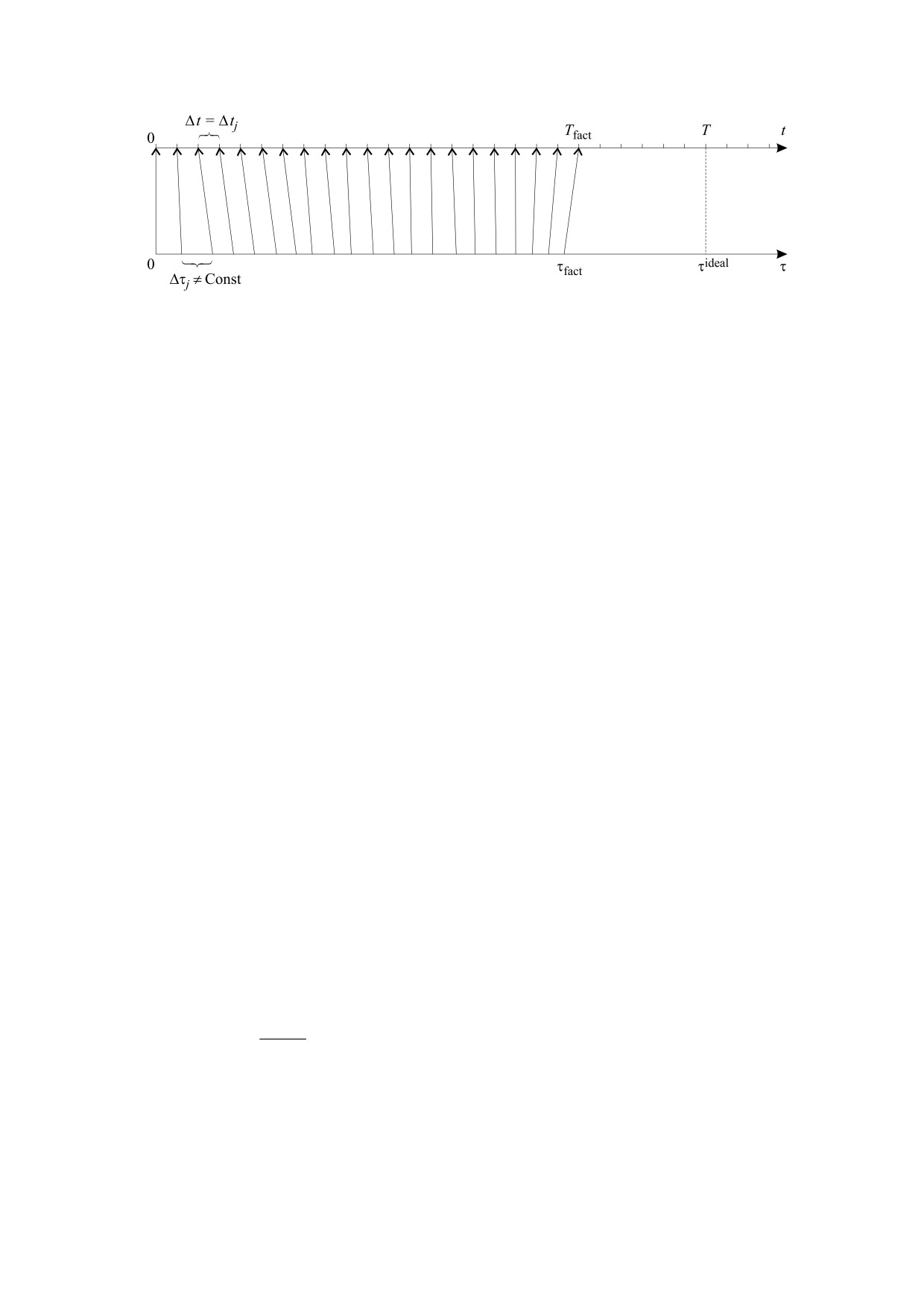
Рис. 3. Иллюстрация связи индивидуального времени с хронологическим временем: T и Tfact - максимальная и
фактическая продолжительность жизни организма, τfact и τideal - фактическое и максимальное (идеальное) значение
собственного времени организма.
Как следует из формулы (13), текущее значение
жет принимать значение как больше, так и мень-
индивидуального времени организма зависит от
ше единицы.
фундаментальной величины теории И. Пригожи-
на - второго дифференциала энтропии δ2H, а
НЕПРЕРЫВНО-ДИСКРЕТНОЕ
также от первого дифференциала энтропии δH. В
ИНДИВИДУАЛЬНОЕ ПРОСТРАНСТВО-
случае отрицательного значения δ2H и при усло-
ВРЕМЯ ОРГАНИЗМА
вии, что |δ2H| > |δH|, собственное время организма
Изучению свойств непрерывно-дискретного
Δθi больше хронологического Δti, в противном
пространства-времени посвящено значительное
случае собственное время меньше хронологиче-
число исследований философов, начиная с ан-
ского. На рис. 3 представлена геометрическая ил-
тичных времен [28, 29] и до современного изуче-
люстрация соотношения (13). Изоморфное соот-
ния данной проблемы [30]. В работе В.И. Вернад-
ветствие между индивидуальным и хронологиче-
ского [31] время рассматривается как единство
ским временами определяется кривизной тренда
длительности (дления) и бренности (разрушения
энтропии, которая определяется из опыта или
и длительности). Существенное число исследова-
расчета, например, используя аналитический ап-
ний в психофизиологии посвящено проблеме ин-
парат фрактально-кластерной теории [19].
дивидуального времени [32-37]. Психика челове-
Уравнение (13), формализующее гипотезу ин-
ка, дублирующая работу центральной нервной
вариантности функционала действия энтропии
системы для более совершенной адаптации орга-
для полной продолжительности организма (гло-
низма и личности в окружающей действительно-
бальная инвариантность), можно переписать с
сти (изначально только к биологической и мате-
учетом выше приведенных допущений для отрез-
риальной, а затем и к социальной), является наи-
ка времени (локальная инвариантность) следую-
более сложной, а поэтому и наиболее
щим образом:
информативной системой на предмет исследова-
ния собственного времени. Проблема психофи-
D
j
Δt
= Δθ
,
(14)
зиологического параллелизма была поднята Р.
j
j
Декартом еще в ХVIII веке. С тех пор многочис-
где величина Dj может быть интерпретирована
ленные и разноплановые работы по выявлению
как аналог фрактальной размерности шкалы соб-
механизмов и характера взаимосвязи психиче-
ственного времени организма. Эта величина мо-
ского и физиологического открыли путь к созда-
жет быть определена как формальное решение
нию психофизиологии, нейропсихологии ряда и
уравнения
наук о мозге. Протекание психических функций,
D
i
как и их развитие, неизбежно привело к вопросу
Δt
=W
⋅ Δt
,
(15)
j
j
j
индивидуального времени. В тех редких трудах, в
т.е.
которых этот вопрос был поставлен открыто, бы-
ли получены мировые открытия в области психо-
logW
j
D
=
1+
(16)
логии. Так, например, на основе идей Л.С. Вы-
j
logΔt
готского были открыты законы гетерохронии в
j
развитии центральной нервной системы, а также
Из соотношения (16) следует, что размерность
общие законы психического развития в норме и
Dj текущего индивидуального времени на j-м от-
при дизонтогенезе, легшие в основу дефектоло-
резке, в отличие от усредненной размерности ин-
гии, специальной психологии, коррекционной
дивидуального времени жизни организма D, мо-
педагогики и др. [32]. Кроме того, сама постанов-
БИОФИЗИКА том 66
№ 3
2021
618
ВОЛОВ
ка проблемы темпа и времени развития высших
Примем, что минимальный масштаб времени
психических функций как «психологических си-
в шкале хронологического и индивидуального
стем», их несовпадения во времени, предопреде-
времени один и тот же:
лило появление науки нейропсихологии, открыв-
шей законы функционирования головного мозга
Δtmin = Δθmin.
при формировании и реализации высших психи-
ческих функций, и понимание основ происхож-
Тогда
дения психики как системы (культурно-истори-
ческая теория). Однако прецедент в постановке
Δθj = Nj⋅Δθmin,
вопроса о собственном (индивидуальном) психи-
ческом времени был совершен еще З. Фрейдом
где Nj - натуральное число, и мы имеем:
[33], вне связи с физиологией. Идея влечения к
смерти в психодинамике больного неврозом и
Nj = Wj⋅Mj.
(18)
здорового индивида получила адекватное разви-
тие в теории Г.М. Назлояна [34], введшего поня-
Из анализа различных процессов в организмах
тие «психической смерти» и «психической пау-
следует, что имеет место иерархия их длительно-
зы», ставшей основой уникальной психотехноло-
сти. Так, например, у человека время импульсов в
гии в лечении шизофрении [34]. Восприятие и
нейронах мозга соответствует Δt = 10-5 с, α-цик-
переживание времени как когнитивный феномен
лы мозга имеют длительность Δt = 0.1 с, частота
был исследован в работах Б.И. Цуканова, вы-
сердечных сокращений составляет примерно 1
явившего инвариант временного самоощущения,
Гц, т.е. один удар в секунду. Однако в контексте
который оказался врожденной константой био-
настоящего исследования важен минимальный
логических часов индивида [35]. В обыденной
промежуток времени, в течение которого проис-
жизни феномен ускорения и замедления индиви-
ходит значимые измеряемые изменения состоя-
дуального времени известен по данным интро-
ния организма. Возможно, инвариант временно-
спекции (самоотчет на основе субъективных дан-
го самоощущения, выявленного Б.И. Цукановым
ных). Так, по ощущению в детстве (этап I на
[39], который оказался врожденной константой
рис. 1) собственное время течет медленнее, а в
биологических часов индивида, может быть свя-
зрелом возрасте, с годами, все быстрее и быстрее
зан с минимальным масштабом времени Δtmin.
(этап III на рис.1).
Однако данное предположение требует отдельно-
Наиболее близкой работой в этом ряду к на-
го изучения.
стоящему исследованию является монография
Очевидно, что в контексте предлагаемой гипо-
В.И. Жаркова [30]. Сформулированные в выше-
тезы энтропийного инварианта целесообразно
приведенных работах свойства дискретности
ввести понятие условного «кванта» действия эн-
пространства и индивидуального времени и кон-
тропии или «кванта» старения организма, пред-
тинуальности единого пространства-времени со-
ставляющего собой порцию энтропии, получае-
ответствуют предложенному энтропийному ин-
мой (генерируемой) организмом за минимально
варианту. Дискретность энтропии сложной си-
возможный промежуток времени своего развития
стемы (организма) может быть объяснена
Δtmin:
особенностью термодинамического метода опи-
сания явлений: любой неравновесный процесс
εi = Δtmin⋅Hi.
(19)
заменяется совокупностью равновесных процес-
сов. В контексте настоящей работы непрерыв-
ность пространства-времени сложной системы
Примем, что при выборе масштаба деления
проявляется в инвариантности функционала дей-
шкалы времени внутри отрезка длительности Δti
ствия энтропии (4).
кривизна тренда энтропии будет иметь постоян-
ное значение (δ2H = const). Уравнение (15) с уче-
Следует отметить, что минимальный масштаб
том «квантования» действия энтропии примет
времени, на котором можно исследовать самоор-
следующий вид:
ганизующуюся систему, будет различен для раз-
личных видов организмов, но для любой эволю-
logW
(H)
ции системы каждый ее временный этап будет
j
D
=
1+
(20)
j
кратен ее минимальному временному масштабу
log
(
M
⋅Δt
)
j
min
Δtmin, т.е.
В уравнении (20) осуществлена перенорми-
Δti = Mi⋅Δtmin,
(17)
ровка времени на максимальное время жизни ор-
ганизма и оно дает связь между размерностью те-
где Mi - натуральное число, зависящее от номера
кущего индивидуального времени, энтропией и
этапа i.
минимальным масштабом времени.
БИОФИЗИКА том 66
№ 3
2021
ГИПОТЕЗА ОБ ЭНТРОПИЙНОМ ИНВАРИАНТЕ
619
Таблица 1. Показатели суммарного за потенциальное время жизни удельного метаболизма щуки из разных
водоемов, а также параметры аппроксимации роста на основе уравнения Берталанфи
Водоем
Winf, г.
k
Tfact , годы
W, г
Emax, ккал/г
Fi(Hi)
Источник
Бобков и др.,
Оз. Баунт
3212
0.621
9
1946
91
0.619
1987 [40]
Руденко,
Оз. Чернявское
4917
0.570
10
3040
93
0.627
2000 [43]
Руденко,
Оз. Кудо
8801
0.394
14
5261
114
0.622
2000 [43]
Оз. Жемчужное
7525
0.368
15
4496
123
0.618
Оз. Сомнино
17411
0.257
19
10633
132
0.6077
Оз. Ужо
13259
0.267
21
8048
150
0.621
Бобков,
Оз. Кокотель
22823
0.240
24
13870
157
0.631
Соколов,198
8 [41]
Руденко,
Оз. Кривое
52519
0.175
32
31890
176
0.632
2000 [43]
Оз.Красненькое
54064
0.121
47
33232
248
0.624
Щука
5762
0.29
19
3465
0.607
(Красненькое)
Среднее)
142.7 ± 45.9
0.622 ± 0.01
ОБСУЖДЕНИЕ ВОЗМОЖНОСТИ ПРОВЕРКИ
ние всей жизни. В связи с данным фактом функ-
ГИПОТЕЗЫ ЭНТРОПИЙНОГО
ция роста может быть аппроксимирована уравне-
ИНВАРИАНТА ОРГАНИЗМА
нием Берталанфи [18]. В табл. 1 и 2 представлен
Проверка выдвинутой гипотезы о глобальной
суммарный за потенциальное время жизни удель-
и локальной инвариантности энтропийного
ный метаболизм (Emax) щуки и окуня из разных
функционала F(H)-действия энтропии может, в
водоемов, а также параметры аппроксимации ро-
частности, апробирована на опытных данных по
ста рыб на основе уравнения Берталанфи
метаболизму организмов [36-40]. Как известно,
рост беспозвоночных и рыб продолжается в тече-
(k,Winf,Tfact,Emax):
W(t) = [(Winf)1/3 - ((Winf)1/3 - (w0)1/3)exp(-kt/3)]3,
(21)
где Winf - асимптота уравнения (21), k - параметр
Предлагаемая математическая мера - детер-
минированная энтропия метаболизма - будет
уравнения,
w
- значение активной массы орга-
0
иметь следующий вид:
низма в момент времени t = 0. Скорость потреб-
ления кислорода единицей массы рыбы опреде-
Hi(qi(t) = [-αilog(qi(t)/qimax)]μi,
(23)
ляется по следующей аллометрической формуле
где αi = -1/(log(qimin/qimax), μi = fi(Tifact/Timax)H0/fi,
[18]:
- минимальное и максимальное зна-
qimin и qimax
q(t) = 0.31(w(t))-0.19.
(22)
чение удельного потребления кислорода (мета-
болизма) организмом i-го вида соответственно,
БИОФИЗИКА том 66
№ 3
2021
620
ВОЛОВ
Таблица 2. Показатели суммарного за потенциальное время жизни удельного метаболизма окуня из разных
водоемов, а также параметры аппроксимации роста на основе уравнения Берталанфи
Водоем
Winf, годы
k
Tfact , годы
W, г
Emax, ккал/г
Fi(Hi)
Источник
Озера
Оз. Сосновое
797
0.438
12
468
131
0.57
Хакассии…,
[42]
Руденко,
Оз. Жемчужное
1039
0.240
23
629
232
0.566
2000 [43]
Оз. Рубанково
810
0.248
23
507
239
0.567
Озера
Оз. Бугаево
6850
0.234
24
4132
179
0.601
Хакасси,
1976 [42]
195.3 ± 43.8
0.575 ± 0.025
Среднее
Tifact и Timax - фактическая и максимальная про-
q(t) = q0(1 - ω1+βtβ), (25)
(25)
должительность жизни i-го вида организма соот-
ветственно, fi - константа i-го вида организма.
где ω(M) = 0.41M-0.25, β = 2.6 ± 0.18.
Из данной формулы следует, что предложен-
Необходимо отметить, что различие алломет-
ная нормированная мера - детерминированная
рических соотношений (22) и (25) принципиаль-
энтропия метаболизма i-го организма Hi - соот-
ных различий не имеет, так как оно обусловлено
ветствует всем правилам построения математиче-
разными аппроксимациями экспериментальных
ской энтропии [27]. Она позитивна (Hi > 0), изме-
данных, используемыми авторами представлен-
ных работ, и для них всегда характерна отрица-
няется в пределах от нуля до единицы и аддитив-
тельная степенная зависимость метаболизма от
на. Энтропийный функционал Fi(Hi) согласно
массы организма.
выражению (7) определится следующим образом:
Как следует из табл. 3, для других видов рыб
Tfact
из разных водоемов значение энтропийного ин-
F(H
i
)
= ∫ H q(t))
i
i
dt
/T
fact
(24)
варианта F(Hi) также принимает значение, близ-
0
кое к энтропии «золотого сечения» (F(Hi) =
= 0.6125 ± 0.08). В табл. 3 параметры p и r - досто-
Константа fi для i-го вида организма в формуле
верность и коэффициент корреляции опытных
детерминированной энтропии метаболизма орга-
данных. Таким образом, для разных видов рыб
низма (23) определяется по калибровке функцио-
получено подтверждение инвариантности энтро-
нала (24) на значение энтропии «золотого сече-
пийного функционала метаболизма при фикси-
ния» H0 для организма в популяции с максималь-
рованном значении f показателя энтропии (24).
ной продолжительностью жизни Tmax.
Среднее значение энтропийного инварианта
Как следует из табл. 1 и 2, энтропийный инва-
F(Hi) при фиксированном значении f (f = δ =
риант F(Hi) для рыб (щуки, окуни) из разных во-
= 0.46609 - первая постоянная Фегенбаума) для
доемов имеет практически постоянное значение.
рассмотренных видов рыб составляет <F(Hi)> =
Так, например, для щук и окуней с различной
= 0.603 ± 0.028, т.е. отклонение от среднего значе-
продолжительностью жизни значение энтропий-
ния не превышает 5%. В то же время отклонения
ного инварианта F(Hi) составляет соответственно
от среднего значения потребления полной энер-
0.622 ± 0.01 и 0,575 ± 0.025. В работе [44] предло-
гии (кислорода) на один грамм веса организма
жены соотношения роста и метаболизма для рыб,
Еmax за весь период жизни превышают
30%
отличающиеся от аллометрических соотношений
(табл. 1 и 2). Необходимо отметить, что в формуле
в работе (22):
детерминированной энтропии метаболизма (23)
БИОФИЗИКА том 66
№ 3
2021
ГИПОТЕЗА ОБ ЭНТРОПИЙНОМ ИНВАРИАНТЕ
621
Таблица 3. Результаты расчетов величины энтропийного инварианта F(Hi) для некоторых популяций рыб из
разных водоемов (
w
,
M
,
m
[г],
[годы])
0
∞
max
t
max
Вид, водоем
w0
p
r
k
Winf
Tfact
W
F(Hi)
Густера
(Волгоград.
0.005
0.91 ± 0.04
1.09 ± 0.15
0.28
1867
20
1131
0.606
водохр.)
Ерш (Рубанково)
0.001
0.67 ± 0.06
1.08 ± 0.16
1.22
34
5
22
0.58
Лещ (Жижицкое)
0.001
0.95 ± 0.03
1.00 ± 0.20
0.17
6111
34
3772
0.65
Лещ (Локново)
0.001
0.92 ± 0.02
1.23 ± 0.10
0.27
3087
21
1855
0.66
Плотва
0.011
0.95 ± 0.05
0.75 ± 0.18
0.16
2908
35
1788
0.604
(Жемчужное 1)
Плотва
0.011
0.94 ± 0.08
0.78 ± 0.24
0.20
1876
29
1161
0.6012
(Жемчужное 2)
Плотва (Сомино)
0.011
0.89 ± 0.16
0.96 ± 0.52
0.34
754
17
470
0.602
Плотва
0.011
0.90 ± 0.07
0.98 ± 0.26
0.31
1013
18
612
0.6
(Чернявское)
Среднее
0.613 ± 0.033
показатель f для рыб разных видов равен первой
гипотезы энтропийного инварианта не превыша-
универсальной константе Фегенбаума (δ = 4.669),
ет 2-3%.
связанной с процессом удвоения логистического
Косвенным подтверждением гипотезы об эн-
отображения, что, возможно, имеет глубинную
тропийном инварианте для организмов также яв-
связь с единицей внутреннего времени, введен-
ляются исследования, проведенные в работах
ной в работе [18], которая определяется как время
[41-43]. Нормированная энтропия двигательной
удвоения массы организма для пойкилотермных
активности молодых здоровых людей соответ-
животных.
ствует энтропии «золотого сечения», что можно
сопоставить II этапу эталонного тренда энтропии
Как отмечено выше, для расчета составляю-
(рис. 1а).
щих энтропийного фактора W(H), в частности,
Как отмечено выше, феномен ускорения и за-
можно использовать аппарат неравновесной тер-
медления индивидуального времени известен по
модинамики [1], фрактально-кластерной теории
данным интроспекции: в детстве (этап I на рис. 1)
[23-25] и аппарат построения условных энтро-
собственное время течет медленнее, а в зрелом
пий [26, 27].
возрасте, с годами, все быстрее и быстрее (этап III
Проведенный в работе [25] анализ на основе
на рис. 1). Это также является косвенным под-
фрактально-кластерной теории
[25] развития
тверждением предложенной гипотезы.
39 видов организмов, список которых начинается
с древнейших хламидомонад, гидр и до Homo
ЗАКЛЮЧЕНИЕ
sapiens, также позволяет количественно подтвер-
дить постулируемую гипотезу - в среднем откло-
Представлен обзор исследований, посвящен-
нение по рассматриваемым видам от выполнения ных проблеме собственного (физиологического)
БИОФИЗИКА том 66
№ 3
2021
622
ВОЛОВ
времени биологических организмов и интеграль-
стоянная Фегенбаума) для рассмотренных ви-
ных характеристик их жизнедеятельности, и из-
дов рыб составляет <F(Hi)> = 0.603 ± 0.028, т.е.
ложены результаты оригинальных исследований
отклонение от среднего значения не превышает
автора. Проверка в многочисленных исследова-
5%. Интересным является факт, что в формуле
ниях [15-21, 39] гипотезы Рубнера [5], постулиру-
(23) детерминированной энтропии метаболизма
ющей для некоторых видов млекопитающих при-
организма показатель f для рыб разных видов ра-
близительно равное количество энергии, исполь-
вен первой универсальной константе Фегенбау-
зуемое единицей массы взрослого животного за
ма, связанной с процессом удвоения логистиче-
период жизни от момента прекращения роста до
ского отображения, что, возможно, имеет глу-
смерти, показало, что расширение постоянства
бинную связь с единицей собственного
«константы Рубнера» для видов, в том числе пой-
(физиологического) времени, введенной в рабо-
килотермных животных, например для моллюс-
те [18], которая определяется как время удвое-
ков и рыб, не имеющих границы роста, представ-
ния массы организма для пойкилотермных жи-
ляется необоснованным.
вотных.
Ввиду данного факта была предложена гипоте-
Необходимо позиционировать единицу соб-
за энтропийного инварианта для организмов.
ственного времени организма, введенную в пред-
Суть данной гипотезы заключается в постулиро-
ставленной статье, и единицу собственного (фи-
вании инвариантности вновь введенной физиче-
зиологического) времени, введенную в работе
ской величины - функционала действия энтро-
[18]. Единица собственного (физиологического)
пии для полной продолжительности жизни орга-
времени, введенная авторами работы [18], пред-
низма (глобальная инвариантность) и для любого
ставляет собой значимый временной период, в
отрезка жизни организма, в течение которого
течение которого происходит удвоение массы ор-
имеет место измеряемое изменение его состоя-
ганизма, не имеющего границу роста.
ния (локальная инвариантность). Одним из след-
В контексте настоящего исследования едини-
ствий данной гипотезы является полученная
ца собственного времени - это минимальный
связь индивидуального времени организма с хро-
промежуток времени, в течение которого проис-
нологическим временем. Полученная формула
ходит измеряемое изменение состояния организ-
включает фундаментальную физическую величи-
ма. Этой минимальной единицей собственного
ну теории структуры И. Пригожина - второй
времени организма (для человека) может быть
дифференциал энтропии.
инвариант временного самоощущения, выявлен-
Введено понятие «кванта» действия энтропии
ного Б.И. Цукановым [3], который оказался
или «кванта» старения организма, представляю-
врожденной константой биологических часов
щего собой порцию энтропии получаемой (гене-
индивида и может быть связан с минимальным
рируемой) организмом за минимально возмож-
масштабом времени Δtmin. Также кандидатом на
ный промежуток времени Δtmin развития системы
минимальную единицу собственного времени
i-го типа.
организма может быть цитохрон (время деления
Другим следствием данной гипотезы являет-
клетки). Однако данные предположения требуют
ся утверждение о фрактальной структуре инди-
отдельного изучения.
видуального времени организма. Важно отме-
Для дальнейшей проверки предложенной ги-
тить, что имеет место глубинная аналогия между
потезы энтропийного инварианта должны быть
Вторым началом термодинамики (ΔS ≥ 0), где
проведены широкомасштабные исследования с
знак равенства изменения термодинамической
использованием опытных данных по метаболиз-
энтропии ΔS относится к обратимым (идеаль-
му и росту организмов для следующих организ-
ным процессам)) и отклонением размерности
мов: 1) млекопитающих; 2) птиц; 3) пойкило-
индивидуального времени жизни организма от
термных (моллюски, рыбы); 4) насекомых; 5) ам-
его идеального значения ΔD = D - Dideal ≥ 0 (где
фибий и 6) организованных клеточных структур.
знак равенства относится к организмам, имею-
Несомненно, актуальным исследованием мо-
щим идеальный (эталонный) тренд энтропии).
жет быть:
Проверка выдвинутой гипотезы о глобальной и
локальной инвариантности функционала дей-
1) определение зависимости продолжительно-
ствия энтропии F(H) была апробирована на
сти жизни организмов (высших млекопитающих)
опытных данных по метаболизму рыб разных
от аккумулированной организмом энтропии,
видов [36-38]. Для разных видов рыб получено
обусловленной внешними условиями и внутрен-
подтверждение инвариантности функционала
ними причинами;
действия энтропии при фиксированном значе-
2) в сравнительном анализе результатов жиз-
нии показателя энтропии f. Среднее значение
недеятельности тех организмов, продолжитель-
энтропийного инварианта F(Hi) при фиксиро-
ность жизни которых находится во взаимно одно-
ванном значении f (f = δ = 0.46609 - первая по-
значном соответствии с размерами и массой тела
БИОФИЗИКА том 66
№ 3
2021
ГИПОТЕЗА ОБ ЭНТРОПИЙНОМ ИНВАРИАНТЕ
623
(моллюски, рыбы и др.) по валидным методам
17.
А. А. Зотин и И. Г. Владимирова, Изв. РАН. Сер.
оценки [6-19] и предлагаемой гипотезе.
биол. № 3, 331 (2001).
18.
А. Ф. Алимов и Т. И. Казанцева, Изв. РАН. Сер.
биол., № 3, 347 (2007).
БЛАГОДАРНОСТИ
19.
А. Ф. Алимов и Т. И. Казанцева, Докл. РАН 396
Автор выражает благодарность доц. к.ф.-м.н.
(4), 561 (2004).
А.П.Зубареву, и к.псих.н. В.В. Волову за обсужде-
ния и ценные замечания по статье.
20.
Т. А. Детлаф, в кн. Конструкция времени в есте-
ствознании. Ч. 1. Междисциплинарное исследование
(Изд-во МГУ, М., 1996), сс. 59-64.
КОНФЛИКТ ИНТЕРЕСОВ
21.
J. O. Reiss, Amer. Natur. 134 (2), 170 (1989).
Автор заявляет об отсутствии конфликта инте-
22.
M. J. Boddington, J. Theor. Biol. 75 (4), 443 (1978).
ресов.
23.
V. T. Volov, arXiv preprint, arXiv: 1309.1415 (2013).
24.
В. П. Бурдаков, Эффективность жизни (Энергоиз-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
дат, М., 1997).
Настоящая работа не содержит описания ис-
25.
В. Т. Волов, Экономика. Флуктуации и термодина-
следований с использованием людей и животных
мика (Изд-во СНЦ РАН, Самара, 2001).
в качестве объектов.
26.
Н. Ингленд и Дж. Мартин, Математическая тео-
рия энтропии (Мир, М., 1988).
СПИСОК ЛИТЕРАТУРЫ
27.
A. Stakhov, Chaos, Solitons & Fractals 26 (2), 263
1. П. Гленсдорф и И. Пригожин, Термодинамическая
(2005).
теория структуры устойчивости и флуктуации
28.
C. Аристотель, О частях животных (Биомедгиз,
(Мир, М., 1973).
ВПМ, 1937).
2. И. Пригожин и И. Стенгерс, Прядок из хаоса: Но-
вый диалог человека с природой (Прогресс, М.,
29.
М. А. Дынник, Диалектика Гераклита Эфесского
1986).
(РАНИОН, М., 1929).
3. И. Пригожин, Введение в термодинамику необра-
30.
В. И. Жарков, Непрерывно-дискретное простран-
тимых процессов (ИИЛ, М., 1960).
ство и время микрообъектов (Наука, Новосибирск,
4. Г. Хакен, Информация и самоорганизация. Макро-
1971).
скопический подход к сложным системам (Мир, М.,
31.
В. И. Вернадский, Изв. АН СССР. Отд. мат. и
1991).
естеств. наук 4, 511 (1932).
5. M. Rubner, Das problem der Lebensdauer und seine
32.
Л. С. Выготский, Психология развития (Эксмо, М.,
Beziehungen zu Wachstum und Ernährung (München;
2004).
Berlin, 1908).
33.
З. Фрейд, По ту сторону принципа удовольствия
6. B. E. Lebensdauer, Biol. Zentralblatt 51 (1/2), 74
(Фолио, М., 2013).
(1931).
34.
7. G. Backman, Wachstum und organiche Zeit (J.A. Barth,
gi/rojdenie-i-smert.html.
Leipzig, 1943).
35.
Б. И. Цуканов, Время в психике человека (Астро-
8. L. P. du Noüy, Biological time (Methuen, L., 1936).
принт, Одесса, 1999).
9. S. Brody, Growth 1 (1), 60 (1973).
36.
А. И. Бобков, Л. Ф. Калягин и Н. Ф. Калягина, в
10. S. Brody, Bioenergetics and growth (Reinhold Publ.
Сб. науч. тр. ГосНИОРХ (1987), вып. 272, сс. 52-66.
Corp., N.Y., 1945).
37.
А. И. Бобков и А. В. Соколов, в Сб. науч. тр. Гос-
11. А. И. Зотин, Термодинамический подход к пробле-
НИОРХ (1988), вып. 279, сс. 118-130.
мам развития, роста и старения (Наука, М., 1974).
38.
Озера Хакасии и их рыбохозяйственная оценка, под
12. А. И. Зотин, в сб. Биология старения (Наука, Л.,
ред. Б.П. Сигиневича (Кн. изд-во, Красноярск,
1982), сс. 116-129.
1976).
13. А. И. Зотин, Термодинамическая основа реакции ор-
39.
Г.П. Руденко, Продукционные особенности ихтио-
ганизмов на внешние и внутренние факторы (Наука,
ценозов малых и средних озер Северо-запада и их
М., 1988).
классификация (ГосНИОРХ, СПб., 2000).
14. А. И. Зотин и А. А. Зотин, Онтогенез 27 (2), 147
40.
R. E. Ricklefs, J. Avian Biol. 31, 2 (2000).
(1996).
41.
В. Т. Волов, Известия Самарского научного цен-
15. А. И. Зотин и Р. С. Зотина, Феноменологическая
тра РАН 5 (2), 204 (2018).
теория развития, роста и старения организма (На-
42.
V. T. Volov and V. V. Volov, arXiv: 1510.02679 [q-
ука, М., 1993).
bio.NC].
16. А. И. Зотин и Т. А. Алексеева, Физиол. журн. 30
43.
В. Т. Волов и В. В. Волов, Нац. психологич. журн.
(1), 59 (1984).
4 (24), 98 (2016).
БИОФИЗИКА том 66
№ 3
2021
624
ВОЛОВ
The Hypothesis about Entropy Invariant for Biological Organisms
V.T. Volov
Samara State University of Railway Transport, ul. Svobody 2v, Samara, 443110 Russia
The article presents an overview of studies devoted to the problem of intrinsic factors (physiological history)
and extrinsic factors that shape the life history of biological organisms including new original author’s results.
A hypothesis about the invariance of a newly introduced extrinsic factor as the functional of the entropy ac-
tion has been formulated. According to this hypothesis, the value of the entropy action functional is the same
for all organisms of a particular species. Based on the proposed hypothesis, the interaction has been found
between the intrinsic (chronological) and environmental (extrinsic) factors that control the growth of biolog-
ical organism. The resulting formula for this interaction includes the fundamental function of I. Prigogine's
theory of structure; it is the second differential of entropy. It is shown that the intrinsic factor of an organism
has a fractal structure. A formula for a mathematical measure - the deterministic entropy of the organism 's
metabolism - has been proposed, and allowed us to test this hypothesis for some groups of organisms. The
perspectives of the proposed hypothesis testing for organisms are discussed.
Keywords: Rubner’s constant, entropy invariant, intrinsic (physiological) factor of an organism, functional of en-
tropy action, second differential of entropy
БИОФИЗИКА том 66
№ 3
2021