БИОФИЗИКА, 2021, том 66, № 4, с. 704-710
БИОФИЗИКА КЛЕТКИ
УДК 577.352.4
РЕГУЛЯЦИЯ ЦИКЛА СИНАПТИЧЕСКИХ ВЕЗИКУЛ И ОБРАЗОВАНИЯ
АКТИВНЫХ ФОРМ КИСЛОРОДА В СИНАПТОСОМАХ МОЗГА КРЫС
ПРИ ДЕЙСТВИИ ЛАКТАТА
© 2021 г. Т.Г. Дубовская*, С.В. Гриневич**, С.В. Федорович*, **
*Институт биофизики и клеточной инженерии НАН Беларуси, 220072, Минск, ул. Академическая, 27, Беларусь
**Биологический факультет Белорусского государственного университета,
220030, Минск, ул. Курчатова, 10, Беларусь
E-mail: sergeifedorovich@yahoo.co.uk; fedorovich@bsu.by
Поступила в редакцию 07.12.2020 г.
После доработки 03.03.2021 г.
Принята к публикации 10.03.2021 г.
Различные метаболиты способны модулировать функционирование пресинаптических окончаний
нейронов. Мы изучили влияние лактата и агониста его рецептора HCAR1 3,5-дигидрокси-
бензойной кислоты на цикл синаптических везикул и образование активных форм кислорода в си-
наптосомах мозга крыс. Лактат в концентрации 4 мМ, в отличие от пирувата, фактически не влияет
на цикл синаптических везикул. С помощью флуоресцентного зонда акридинового оранжевого
продемонстрировано, что 3,5-дигидроксибензоевая кислота в концентрации 500 мкМ активирует
эндоцитоз. Эта стимуляция позволяет скомпенсировать его потенциальное ингибирование. Лактат,
но не 3,5-дигидроксибензоевая кислота вызывает увеличение флуоресценции 2',7'-дихлорофлуо-
ресцина диацетата, свидетельствующее об образовании активных форм кислорода. С помощью
флуоресцентного зонда митосокс мы также показали образование супероксид-аниона в митохон-
дриях при действии лактата. Таким образом, лактат способен влиять на работу пресинаптических
окончаний нейронов за счет стимуляции образования активных форм кислорода и активации эндо-
цитоза, опосредованной рецептором лактата HCAR1.
Ключевые слова: синаптосомы, лактат, синаптические везикулы, эндоцитоз, активные формы кисло-
рода, митохондрии.
DOI: 10.31857/S0006302921040098
энергии для поддержания синаптической транс-
Глюкоза является основным, но не единствен-
миссии [7, 8].
ным энергетическим субстратом для нейронов.
Показано, что нейрональные митохондрии также
В то же время физиологическая роль молоч-
способны усваивать кетоновые тела, пируват,
ной кислоты не исчерпывается участием в мета-
лактат [1-4]. Все эти вещества химически отно-
болизме. К настоящему времени накоплено до-
сятся к монокарбоксилатам. Более того, согласно
вольно много доказательств, что лактат является
концепции астроцит-нейронального лактатного
также сигнальной молекулой [6]. Для нее иденти-
челнока лактат как минимум в некоторых случаях
фицирован рецептор гидроксикарбоксиловой
является основным энергетическим субстратом
кислоты 1 (HCAR1), связанный с G-белком [6],
[5, 6]. Согласно этой гипотезе, глюкоза преиму-
который экспрессируется в том числе и на плаз-
щественно перерабатывается в лактат в астроци-
матической мембране нейронов [6, 9].
тах, молочная кислота в свою очередь транс-
Нервный импульс переходит от нейрона к
портируется в нейроны. Синаптические мито-
нейрону с помощью особых веществ - нейроме-
хондрии по своим свойствам достаточно
диаторов. Нейромедиаторы хранятся в синапти-
сильно отличаются от митохондрий из тела ней-
ческих везикулах. При увеличении цитозольной
ронов, но в то же время они также могут исполь-
концентрации кальция в пресинаптическом
зовать монокарбоксилаты в качестве источника
окончании происходит экзоцитоз, слияние мем-
бран синаптических везикул и плазматической
Сокращения: HCAR1 - рецептор гидроксикарбоксиловой
мембраны. Для того чтобы подготовиться к экзо-
кислоты 1, ДГБК - 3,5-дигидроксибензойная кислота,
цитозу, синаптическая везикула должна пройти
АФК - активные формы кислорода, DCFDA - 2',7'-ди-
хлорофлуоресцина диацетат.
эндоцитоз, докинг, прайминг [10]. Стадия докин-
704
РЕГУЛЯЦИЯ ЦИКЛА СИНАПТИЧЕСКИХ ВЕЗИКУЛ
705
га включает в себя образование физического кон-
лота (HEPES) (Merck, Германия), DCFDA, 3,3'-
такта между мембранами синаптической везику-
дипропилтиадикарбоцианин (DiSC3(5),
3,5-ди-
лы и плазматической мембраны. Но для дальней-
гидроксибензойная кислота, пируват натрия, лак-
шего слияния еще должен пройти этап АТФ-
тат натрия, кетоглутарат натрия, хлорид дифени-
зависимого прайминга, вероятно, заключающий-
лйодиния (Sigma, США),
5,5',6,6'-тетрахлоро-
ся в разборке белковых комплексов [10]. Многие
1,1'3,3'-тетраэтилбензимидазоло-карбоцианин йо-
стадии цикла синаптических везикул являются
дид (JC-1), акридиновый оранжевый (Serva, Гер-
энергозависимыми процессами [10, 11].
мания), митосокс (ThermoFisher Scientific, США)
Все остальные реактивы - квалификации не ниже
Ранее было показано, что негликолитические
«ос.ч.».
энергетические субстраты способны быть источ-
ником энергии для поддержания цикла синапти-
Выделение синаптосом. Синаптосомы изоли-
ческих везикул [3, 12]. В то же время замена
ровали из полушарий мозга крыс по методу Хайо-
глюкозы на пируват или кетоновые тела в инкуба-
ша [23]. Полученный осадок суспендировали в
ционной среде ингибирует эндоцитоз в синапто-
среде А следующего ионного состава (в мМ):
сомах мозга крыс [3].
NaCl - 132, KCl - 5, MgCl2 - 1.3, NaH2PO4 - 1.2,
Для пирувата рецепторы не известны, для ке-
глюкоза - 10, HEPES - 10, рН 7.4. Суспензию си-
тоновых тел рецепторы находятся в микроглиаль-
наптосом (10-20 мг/мл белка) преинкубировали
ных клетках и их, скорее всего, в отличие от ре-
10 мин при 37°С, после чего помещали на лед и
цепторов для лактата, нет в нейронах [13, 14]. По-
использовали в эксперименте в течение 3 ч. В
этому в данной работе мы изучили действие
экспериментах с использованием различных
лактата и агониста его рецептора 3,5-дигидрокси-
энергетических субстратов и изучением мем-
бензойной кислоты (ДГБК) на цикл синаптиче-
бранного потенциала или цикла синаптических
ских везикул. Также мы проверили, что лактат,
везикул глюкозу замещали 4 мМ пирувата или
как и пируват, способен поддерживать потенциал
4 мМ лактата. Также в экспериментах по изуче-
плазматической мембраны синаптосом [1, 3].
нию образования АФК или митохондриального
потенциала эти вещества дополнительно вносили
Недавно было показано, что сигнальная функ-
к уже имеющейся в инкубационной среде глюко-
ция лактата может опосредоваться увеличением
зе (10 мМ). Их растворяли в инкубационной
образования активных форм кислорода (АФК)
среде А и вносили в объеме 20 мкл. При регистра-
[15]. Показано, что пируват ведет к образованию
ции флуоресценции в инкубационную среду до-
пероксида водорода во внутрисинаптосомальных
бавляли 2 мМ CaCl2.
митохондриях [16]. Поэтому мы изучили образо-
вание активных форм кислорода (АФК) в синап-
Определение потенциала плазматической мем-
тосомах при действии лактата.
браны. Потенциал плазматической мембраны си-
наптосом определяли с помощью флуоресцент-
В качестве объекта исследования были ис-
ного зонда DiSC3(5) по Waseem and Fedorovich
пользованы синаптосомы, изолированные пре-
[19] c некоторыми модификациями [24]. Флуо-
синаптические окончания нейронов. Они сохра-
ресценцию измеряли на спектрофлуориметре
няют основные свойства интактных нервных тер-
Cary Eclipse (Varian, США) в термостатируемой
миналей, способны к экзоцитозу и эндоцитозу [3,
кювете (37°С) при λвозб = 640 нм, λрег = 688 нм и
17]. Синаптосомы также содержат митохондрии,
которые способны метаболизировать как глюко-
постоянном перемешивании. После выделения
зу, так и негликолитические энергетические суб-
синаптосомы преинкубировали в среде А с соот-
страты [3, 17].
ветствующими энергетическими субстратами
30 мин при 37°С. К 1.8 мл инкубационной среды
Экзоцитоз и эндоцитоз изучали с помощью
А добавляли 200 мкл суспензии синаптосом (до
флуоресцентного зонда акридинового оранжево-
конечной концентрации белка 1 мг/мл), после
го [18], потенциал плазматической мембраны - с
чего в кювету вносили 1 мкМ DiSC3(5). Через од-
помощью зонда DiSC3(5) [19], митохондриаль-
ну минуту добавляли 150 мМ KCl. В контрольном
ный потенциал - с помощью флуоресцентного
эксперименте добавляли 150 мМ NaCl. В даль-
зонда JC-1 [20], образование АФК - с помощью
нейшем из опытной кривой вычитали контроль-
флуоресцентного зонда 2',7'-дихлорофлуоресци-
ную.
на диацетата (DCFDA) [21], образование су-
Регистрация экзоцитоза и эндоцитоза. Экзоци-
пероксидного анион-радикала в митохондриях -
тоз и эндоцитоз определяли с помощью флуорес-
с помощью флуоресцентного зонда митосокса
[22].
центного зонда акридиновый оранжевый по Zoc-
carato et al. [18] c некоторыми модификациями
[24]. Флуоресценцию измеряли на спектрофлуо-
МАТЕРИАЛЫ И МЕТОДЫ
риметре Cary Eclipse (Varian, США) в термостати-
руемой кювете
(37°С) при λвозб = 494 нм,
В работе были использованы реактивы: 4-(2-
гидроксиэтил)пиперазин-1-этансульфоновая кис-
λрег = 529 нм и постоянном перемешивании. По-
БИОФИЗИКА том 66
№ 4
2021
706
ДУБОВСКАЯ и др.
сле выделения синаптосомы преинкубировали в
ной среды. В дальнейшем из опытной кривой вы-
среде с лактатом или агонистом лактатного ре-
читали контрольную.
цептора 30 мин при 37°С. К 1,8 мл среды А с 2 мМ
Определение мембранного потенциала митохон-
СaCl2 добавляли 200 мкл синаптосом (до конеч-
дрий. Мембранный потенциал митохондрий (ΔΨ)
ной концентрации белка 1 мг/мл), после чего в
определялся с помощью флуоресцентного зонда
кювету вносили 5 мкМ акридинового оранжево-
JC-1 по Chinopoulos et al. [20] с небольшими мо-
го. Через 10 мин, необходимых для уравновеши-
дификациями [26]. Флуоресценцию измеряли на
вания зонда между синаптосомами и инкубаци-
спектрофлуориметре Cary Eclipse в термостатиру-
онной средой, в кювету добавляли 60 мМ КСl или
емой кювете (37°С) при λвозб = 490 нм, λрег =
в качестве контроля делали аналогичную добавку
= 595 нм и постоянном перемешивании. После
NaCl. В дальнейшем из опытной кривой вычита-
выделения синаптосомы (концентрация белка
ли контрольную.
5 мг/мл) инкубировали в среде A 15 мин при 37°С
Определение образования АФК. Образование
в присутствии 10 мкг/мл зонда. Избыток экстрак-
АФК определяли с помощью флуоресцентного
леточного зонда удаляли центрифугированием, а
зонда DCFDA по LeBel and Bondy [21] c некото-
осадок суспендировали в инкубационной среде.
рыми модификациями [25]. После дополнитель-
Далее к 1.8 мл среды А в кювете добавляли 200 мкл
ной однократной отмывки в десятикратном объе-
синаптосом (до конечной концентрации белка
ме стандартной инкубационной среды А осадок
1 мг/мл). Через 50 с добавляли энергетические
синаптосом суспендировали в этой же среде. К
субстраты. В контрольном эксперименте добав-
суспензии (10-15 мг/мл белка) добавляли зонд
ляли аналогичное количество инкубационной
DCFDA до конечной концентрации 25 мкМ и
среды. В дальнейшем из опытной кривой вычита-
20% (вес/объем) Pluronic F-127/DMSO (конечная
ли контрольную.
концентрация детергента не превышала 0.05%).
Белок определяли с помощью метода Лоури
Инкубировали 60 мин при 30°С. Загруженные си-
[27] c использованием бычьего сывороточного
наптосомы однократно отмывали в десятикрат-
альбумина как стандарта. Результаты представля-
ном объеме среды А. Осадок суспендировали в
ли в виде среднего арифметического индивиду-
2.0-2.5 мл среды А.
альных измерений и стандартной ошибки
среднего арифметического. Для анализа данных
Флуоресценцию измеряли на спектрофлуори-
использовали однофакторный дисперсионный
метре Cary Eclipse (Varian, США) при постоянном
анализ (ANOVA) с посттестом Бонферрони. Разли-
помешивании в термостатируемой кювете (37°С)
чия считали статистически значимыми при
при λвозб/рег = 501/525 нм.
р < 0.05.
К 1.8 мл среды А добавляли 200 мкл суспензии
синаптосом до конечной концентрации белка
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
1 мг/мл. Через 50 с в кювету вносили соответству-
ющий энергетический субстрат. В контрольном
На первом этапе мы показали, что лактат мо-
эксперименте в кювету добавляли аналогичное
жет использоваться синаптосомами для поддер-
количество растворителя. В дальнейшем из опыт-
жания потенциала плазматической мембраны
ной кривой вычитали контрольную кривую.
(рис. 1). В то же время мы обнаружили неболь-
Определение образования супероксид-аниона во
шую, но достоверную деполяризацию в экспери-
внутрисинаптосомальных митохондриях. Образо-
ментах, где синаптосомы использовали только
вание супероксид-аниона во внутрисинаптосо-
лактат в качестве источника энергии (рис. 1).
мальных митохондриях определялось с помощью
Сходный эффект мы в свое время наблюдали и
флуоресцентного зонда митосокса [22]. Флуорес-
при использовании пирувата
[3]. Потенциал
ценцию измеряли на спектрофлуориметре Cary
плазматической мембраны не изменялся в экспе-
Eclipse в термостатируемой кювете (37°С) при
риментах, в которых лактат добавляли к глюкозо-
λвозб = 510 нм, λрег = 580 нм и постоянном переме-
содержащей инкубационной среде. Это говорит о
том, что рецептор лактата не вносит вклад в депо-
шивании. После выделения синаптосомы инку-
ляризацию пресинаптического окончания. Не-
бировали в среде A 10 мин при 37°С в присутствии
давно было показано, что лактат может регулиро-
5 мкМ зонда. Избыток экстраклеточного зонда
вать возбудимость нейронов как минимум в неко-
удаляли центрифугированием, а осадок суспен-
торых областях головного мозга [6, 28]. В то же
дировали в соответствующей инкубационной
время наши данные показывают, что подобная
среде. Далее к 1.8 мл среды А в кювете добавляли
регуляция, опосредованная рецепторами молоч-
200 мкл синаптосом, предварительно загружен-
ной кислоты, не распространяется на пресинап-
ных зондом (до конечной концентрации белка
тические окончания (рис. 1).
1 мг/мл). Через одну минуту добавляли энергети-
ческие субстраты. В контрольном эксперименте
Далее мы изучили действие лактата на цикл
добавляли аналогичное количество инкубацион-
синаптических везикул в синаптосомах. Ранее мы
БИОФИЗИКА том 66
№ 4
2021
РЕГУЛЯЦИЯ ЦИКЛА СИНАПТИЧЕСКИХ ВЕЗИКУЛ
707
показали, что замена глюкозы на кетоновые тела
приводит к некоторой активации экзоцитоза и
ингибированию эндоцитоза, а замена глюкозы на
пируват - к ингибированию эндоцитоза [6]. Для
изучения цикла синаптических везикул мы ис-
пользовали флуоресцентный зонд акридиновый
оранжевый. Увеличение флуоресценции свиде-
тельствует об экзоцитозе, последующее сниже-
ние - о компенсаторном эндоцитозе [18, 24].
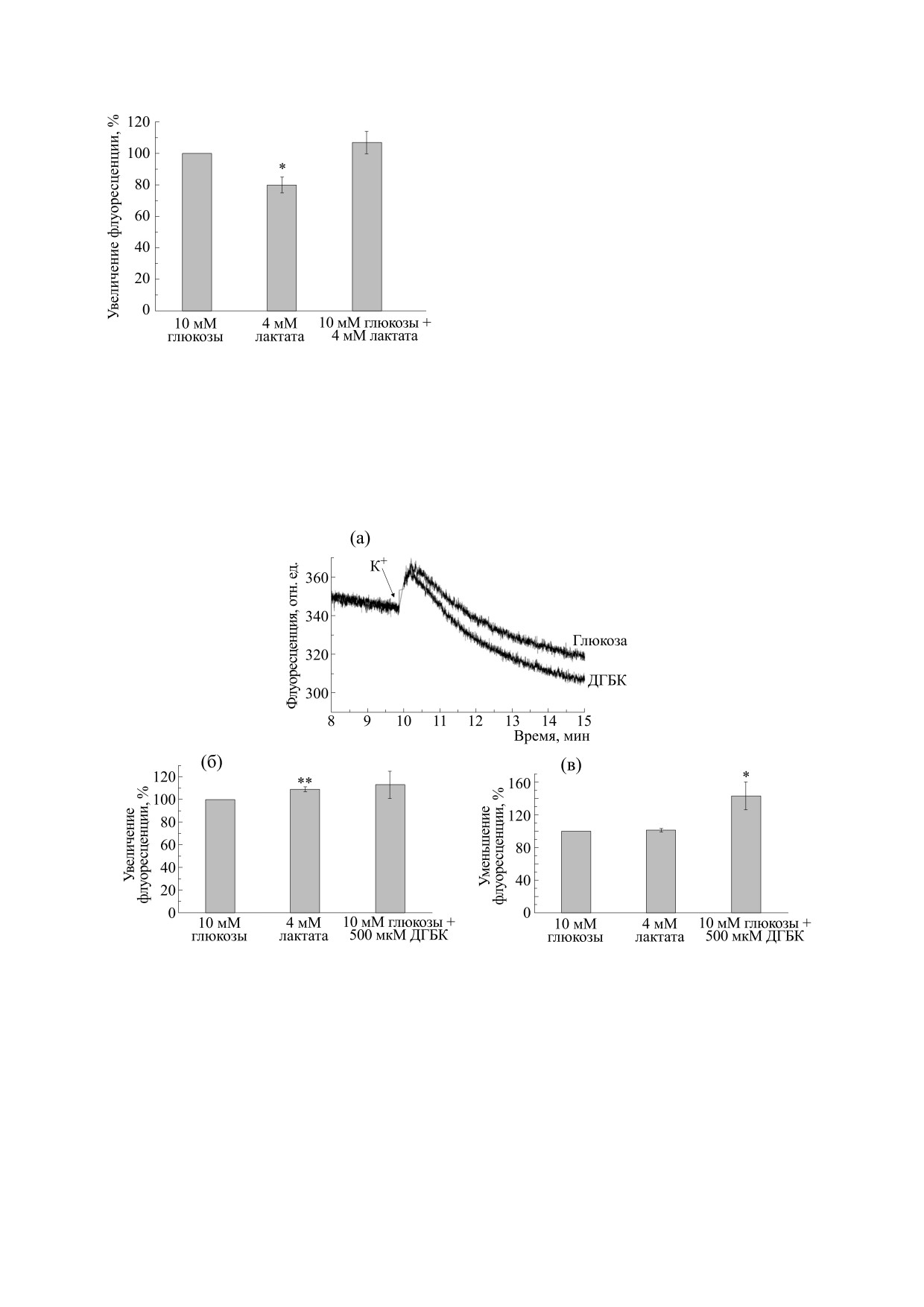
Рис. 2 показывает, что замена глюкозы на лактат
также приводит к небольшому, но достоверному
увеличению экзоцитоза. В то же время полностью
отсутствовало
ингибирование
эндоцитоза
(рис. 2), характерное для других негликолитиче-
ских энергетических субстратов [6]. Мы исполь-
зовали лактат в концентрации 4 мМ, так как ра-
Рис. 1. Влияние лактата на потенциал плазматиче-
ской мембраны синаптосом. За 100% взято увеличе-
нее в наших экспериментах была использована
ние флуоресценции за 4 мин после добавки 150 мМ
такая же концентрация пирувата [6]. Агонист
КСl в контрольной глюкозосодержащей среде.
HCAR1 ДГБК не влияет на экзоцитоз, но активи-
Представлены средние значения шести эксперимен-
рует эндоцитоз (рис. 2). Таким образом, наши ре-
тов ± Sх.
зультаты позволяют предположить, что актива-
Рис. 2. Влияние лактата на цикл синаптических везикул в синаптосомах. (а) - Ответ флуоресцентного зонда акриди-
нового оранжевого на калиевую деполяризацию в контрольной глюкозосодержащей инкубационной среде и в инку-
бационной среде, также содержащей 500 мкМ ДГБК. В указанный стрелкой момент времени в кювету вносили 60 мМ
КCl. Представленные кривые отражают результаты четырех независимых измерений. (б) - Влияние лактата на экзо-
цитоз в синаптосомах мозга крыс: Инкубационная среда содержала 10 мМ глюкозы, либо 4 мМ лактата, либо 10 мМ
глюкозы + 500 мкМ ДГБК. За 100% взято увеличение флуоресценции после добавки КСl в контрольной глюкозосо-
держащей среде. **- P ≤ 0.01 по отношению к контролю (ответ в глюкозосодержащей среде). Представлены средние
значения четырех экспериментов ± Sх. (в) - Влияние лактата на эндоцитоз в синаптосомах мозга крыс. Инкубацион-
ная среда содержала 10 мМ глюкозы, либо 4 мМ лактата, либо 10 мМ глюкозы + 500 мкМ ДГБК. За 100% взято умень-
шение флуоресценции за 5 мин после добавки КСl в контрольной глюкозосодержащей среде. *- P ≤ 0.05
по отношению к контролю (ответ в глюкозосодержащей среде). Представлены средние значения четырех экспери-
ментов ± Sх.
БИОФИЗИКА том 66
№ 4
2021
708
ДУБОВСКАЯ и др.
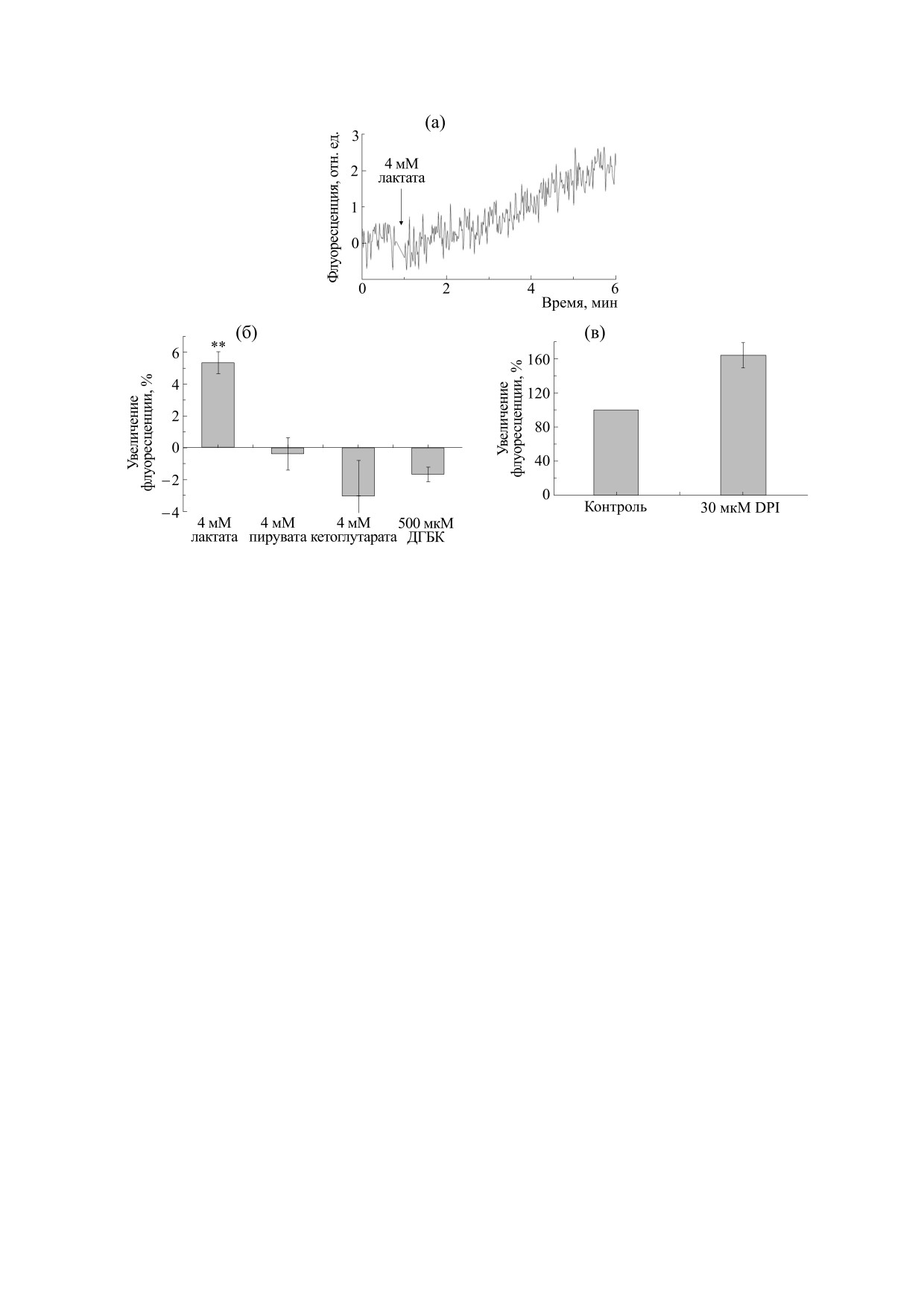
Рис. 3. Влияние монокарбоксилатов на образование АФК в синаптосомах мозга крыс. (а) - Увеличение
флуоресценции DCFDA при действии лактата. В указанный стрелкой момент времени в кювету вносили 4 мМ
лактата. Представленная кривая отражает результаты одиннадцати независимых измерений. (б) - Влияние
монокарбоксилатов на флуоресценцию DCFDA в синаптосомах. В глюкозосодержащую инкубационную среду
добавляли 4 мМ либо лактата, либо пирувата, либо кетоглутарата или 500 мкМ ДГБК. За 100% взято увеличение
флуоресценции за 5 мин после добавления 1 мМ H2O2 в контрольной глюкозосодержащей среде. ** - P ≤ 0.01 по
отношению к 0. Представлены средние значения шести экспериментов ± Sх для лактата, четырех экспериментов для
пирувата, трех экспериментов для кетоглутарата и тринадцати экспериментов - для ДГБК. (в) - Влияние хлорида
дифенилйодиния на образование АФК, индуцированное лактатом: синаптосомы были обработаны 30 мкМ хлорида
дифенилйодиния, за 100% взято увеличение флуоресценции в контроле после добавления 4 мМ лактата.
Представлены средние значения двенадцати экспериментов ± Sх .
ция эндоцитоза при помощи HCAR1 позволяет
ние пероксида водорода в синаптосомах за счет
замаскировать его ингибирование, вызванное
активации кетоглутаратдегидрогеназы и пируват-
лактатом как негликолитическим энергетиче-
дегидрогеназы [16]. В то же время рис. 3б показы-
ским субстратом. Так как лактат в физиологиче-
вает, что эти вещества, в отличие от лактата, не
ских условиях гораздо более важен для питания
ведут к увеличению флуоресценции DCFDA.
нейронов, чем пируват или кетоновые тела, мы
Флуоресцентный зонд DCFDA реагирует прежде
предполагаем, что обнаруженный нами механизм
всего на гидроксильный радикал [29]. Гидрок-
компенсации обеспечивает стабильность синап-
сильный радикал в наибольшей степени спосо-
тической трансмиссии. В то же время стоит отме-
бен повреждать нейроны по сравнению с другими
тить, что лактат, в отличие от пирувата, является
АФК, но он является вторичным по сравнению с
формой, которая хорошо проходит через клеточ-
пероксидом водорода или суперкосиданион ра-
ную мембрану, именно поэтому для транспорта
дикалом [30]. Поэтому наши результаты позволя-
из астроцитов в нейрон используется лактат, а не
ют предположить, что лактат обладает более
пируват. Однако в нейроне лактат переходит в
сильным прооксидантным действием, чем пиру-
пируват, который в конечном итоге и является
ват и кетоглутарат. Видимо, в случае пирувата и
энергетическим субстратом для митохондрий.
кетоглутарата количество образуемого пероксида
водорода, являющегося первичной АФК, недо-
Рис. 3а показывает, что добавление 4 мМ лак-
статочно для синтеза гидроксильного радикала.
тата к суспензии синаптосом приводит к увеличе-
нию флуоресценции DCFDA, свидетельствую-
В синаптосомах различные факторы могут вы-
щее о накоплении АФК. Ранее было показано,
зывать образование АФК за счет образования
что кетоглутарат и пируват вызывают образова-
супероксидного анион-радикала в электрон-
БИОФИЗИКА том 66
№ 4
2021
РЕГУЛЯЦИЯ ЦИКЛА СИНАПТИЧЕСКИХ ВЕЗИКУЛ
709
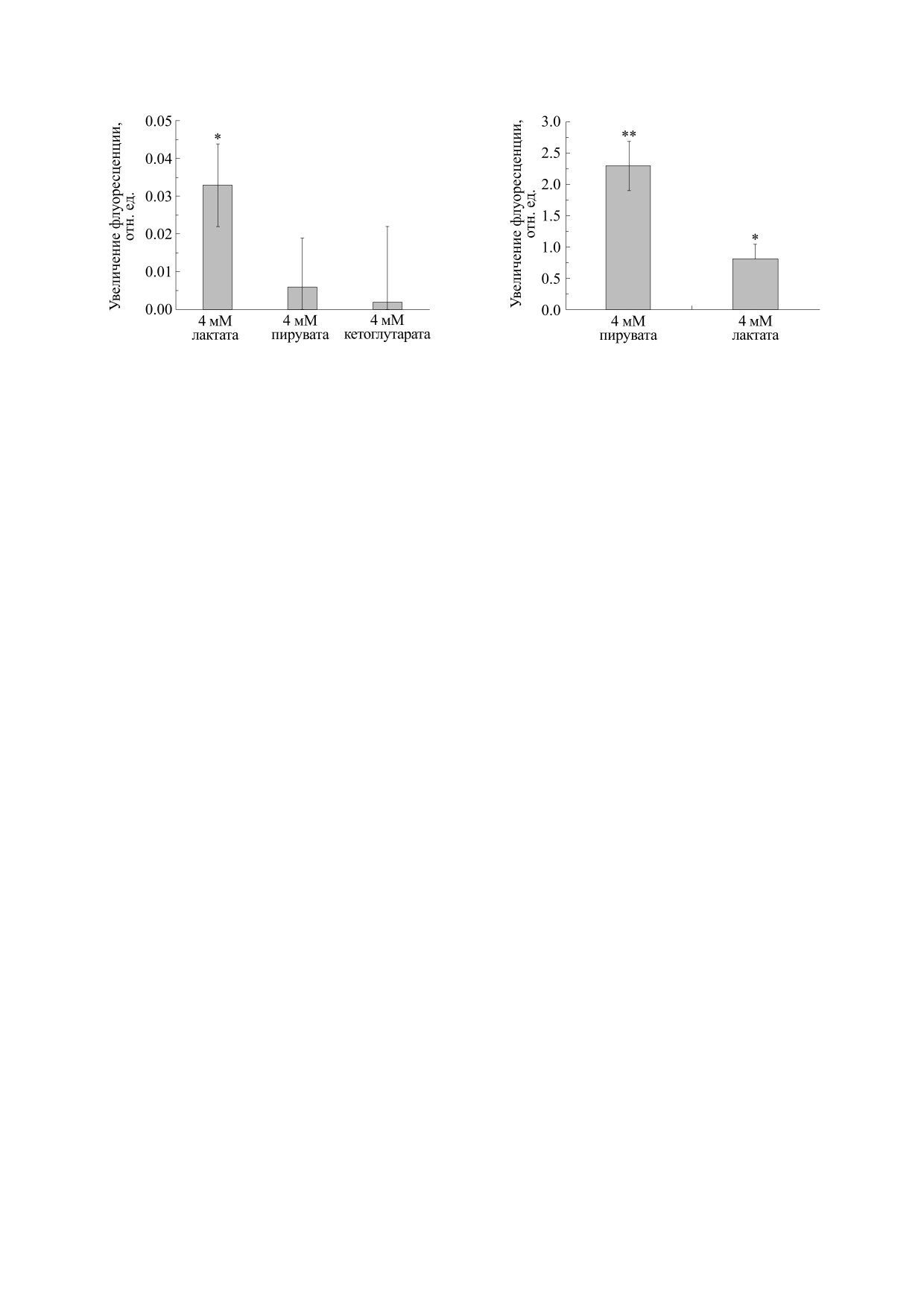
Рис. 4. Влияние монокарбоксилатов на образование
Рис. 5. Влияние монокарбоксилатов на митохондри-
супероксид-аниона в митохондриях. В глюкозосо-
альный потенциал. В глюкозосодержащую инкуба-
держащую инкубационную среду добавляли 4 мМ
ционную среду добавляли 4 мМ лактата или пирувата.
лактата, пирувата или кетоглутарата. *- P ≤ 0.05 по
* - P ≤ 0.05 по отношению к контролю; ** - P ≤ 0.01
отношению к контролю. Представлены средние зна-
по отношению к контролю. Представлены средние
чения семи экспериментов ± Sх.
значения четырех экспериментов ± Sх.
транспортной цепи митохондрий или NADPH
шивается какой-то другой фактор. Этим факто-
оксидазой [25, 26, 31]. Мы изучили действие спе-
ром, видимо, не является рецептор лактата
цифического ингибитора NADPH-оксидазы хло-
HCAR1, так как его специфический агонист
рида дифенилйодиния [25, 26, 31] на увеличение
ДГБК не вызвал образования АФК в синаптосо-
образования АФК при действии лактата. Рис. 3в
мах (рис. 3а). В то же время было предположено,
показывает, что хлорид дифенилйодиния увели-
что существуют и другие типы рецепторов лактата
чивает, а не ингибирует влияние молочной кис-
в нейронах [6]. Возможно, именно этот до сих пор
лоты на флуоресценцию DCFDA. Далее мы ис-
неидентифицированный рецептор и является
пользовали флуоресцентный зонд митосокс, ко-
стимулом для образования супероксид-аниона в
торый специфичен по отношению к супероксид-
электронтранспортной цепи внутрисинаптосо-
аниону, образующемуся в митохондриях [22].
мальных митохондрий.
Рис. 4 показывает, что лактат, но не пируват или
Наши результаты показывают, что лактат вы-
кетоглутарат, вызывает увеличение флуоресцен-
зывает образование супероксид-аниона в элек-
ции митосокса, свидетельствующее об образова-
тронтранспортной цепи во внутрисинаптосо-
нии супероксид-аниона в электронтранспортной
мальных митохондриях. Этот эффект не связан с
цепи митохондрий.
активацией HCAR1 (рис. 3). Молочная кислота, в
Ранее было показано, что пируват вызывает
отличие от других негликолитических энергети-
гиперполяризацию внутрисинаптосомальных
ческих субстратов, не ингибирует эндоцитоз. Это
митохондрий [1]. Гиперполяризация митохон-
связано с активацией эндоцитоза под действием
дрий стимулирует образование АФК в митохон-
рецептора лактата HCAR1 (рис. 2). Таким обра-
дриях [32, 33]. Поэтому мы изучили влияние лак-
зом, лактат способен регулировать функциони-
тата на потенциал внутрисинаптосомальных ми-
рование пресинаптических окончаний нейронов
тохондрий. Рис. 5 показывает, что и пируват, и
за счет стимуляции образования АФК в элек-
лактат вызывают увеличение флуоресценции
тронтранспортной цепи митохондрий и актива-
зонда JC-1 при λрег = 595 нм. Это свидетельствует
ции эндоцитоза, опосредованной HCAR1.
о гиперполяризации [20]. Мы также наблюдали
уменьшение флуоресценции при λрег = 595 нм
ФИНАНСИРОВАНИЕ РАБОТЫ
(данные не показаны), что подтверждает гипер-
поляризацию [20]. В то же время эффект пирувата
Работа выполнена при финансовой поддержке
был более выражен, чем эффект лактата (рис. 5),
Белорусского Республиканского фонда фунда-
что делает связь между потенциалом митохон-
ментальных исследований (грант № Б19-001).
дрий и образованием АФК не столь однозначной.
Более сильная гиперполяризация в случае пиру-
вата скорее всего связана с тем, что, в отличие от
КОНФЛИКТ ИНТЕРЕСОВ
лактата, это вещество сразу используется в цикле
Кребса. Вероятно, в физиологический процесс
Автор заявляет об отсутствии конфликта инте-
регуляции образования супероксид-аниона вме-
ресов.
БИОФИЗИКА том 66
№ 4
2021
710
ДУБОВСКАЯ и др.
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
15. A. Tauffenberger, H. Fiumelli, S. Almustafa, et al., Cell
Death Dis. 10, 653 (2019).
Все применимые международные, националь-
16. L. Tretter, B. Liktor, and V. Adam-Vizi, Neurochem.
ные и институциональные принципы ухода и ис-
Res. 30, 1331 (2005).
пользования животных при выполнении работы
17. B. G. Wilhelm, S. Mandad, S. Truckenbrodt, et al.,
были соблюдены.
Science 344, 1023 (2014).
18. F. Zoccarato, L. Cavallini, and A. Alexandre, J. Neuro-
chem. 72, 625 (1999).
СПИСОК ЛИТЕРАТУРЫ
19. T. V. Waseem and S. V. Fedorovich, Neurochem. Res.
1. R. A. Kauppinen and D. G. Nicholls, FEBS Lett. 199,
35, 1188 (2010).
222 (1986).
20. C. Chinopoulos, L. Tretter, and V. Adam-Vizi, J. Neu-
2. M. C. McKenna, J. T. Tildon, J. H. Stevenson, et al.,
rochem. 73, 220 (1999).
Dev. Neurosci. 15, 320 (1993).
21. C. P. LeBel and S. C. Bondy, Neurochem. Int. 17, 435
3. S. V. Hrynevich, T. V. Waseem, A. Hebert, et al.,
(1990).
Neurochem. Int. 93, 73 (2016).
22. A. Y. Abramov, A. Scorziello, and M. R. Duchen, J.
4. Т. Г. Дубовская, С. В. Гриневич и С. В. Федорович,
Neurosci. 27, 1129 (2007).
Биофизика 64, 500 (2019).
23. F. Hajos, Brain Res. 93, 485 (1975).
5. L. Pellerin and P. J. Magistretti, J. Cereb. Blood Flow
24. S. V. Hrynevich, T. G. Pekun, T. V. Waseem, et al.,
Metab. 32, 1152 (2012).
Neurochem. Res. 40, 1188 (2015).
6. P. J. Magistretti and I. Allaman, Nature Rev. Neurosci.
25. A. V. Alekseenko, V. V. Lemeshchenko, T. G. Pekun,
19, 235 (2018).
et al., Neurosci. Lett. 513, 238 (2012).
7. S. V. Fedorovich, T. V. Waseem, and L. V. Puchkova,
26. T. G. Pekun, V. V. Lemeshchenko, T. I. Lyskova, et al.,
Rev. Neurosci. 28, 363 (2017).
J. Mol. Neurosci. 49, 211 (2013).
8. M. J. Devine and J. T. Kittler, Nature Rev. Neurosci.
27. O. Lowry, H. Rosenbrough, H. Farr, et al., J. Biol.
19, 63 (2018).
Chem. 193, 265 (1951).
9. H. C. Abrantes, M. Briquet, C. Schmuziger, et al., J.
28. N. Sada, S. Lee, T. Katsu, et al., Science 347, 1362
Neurosci. 39, 4422 (2019).
(2015).
10. T. C. Sudhof, Neuron 80, 675 (2013).
29. T.-I. Setsukinai, Y. Urano, K. Kakinuma, et al., J. Biol.
11. S. V. Fedorovich and T. V. Waseem, Rev. Neurosci. 29,
Chem. 278, 3170 (2003).
825 (2018).
30. B. Halliwell, J. Neurochem. 97, 1634 (2006).
12. A. S. Tarasenko, M. V. Linetska, L. G. Storchak, et al.,
31. A. Tarasenko, O. Krupko, and N. Himmelreich, Neu-
J. Neurochem. 99, 787 (2006).
rochem. Int. 61, 1044 (2012).
13. C. C. Blad, C. Tang, and S. Offermans, Nat. Rev. Drug
32. S. S. Korshunov, V. P. Skulachev, and A.A. Starkov,
Discov. 11, 603 (2012).
FEBS Lett 416, 15 (1997).
14. S. P. Fu, J. F. Wang, W. J. Xue, et al., J. Neuroinflam-
33. D. B. Zorov, M. Juhaszova, and S. J. Sollott, Physiol.
mation 12, 9 (2015)
Rev. 94, 909 (2014).
Regulation of the Synaptic Vesicle Cycle and Reactive Oxygen Species Generation
in Rat Brain Synaptosomes in the Presence of Lactate
T.G. Dubouskaya*, S.V. Hrynevich**, and S.V. Fedorovich*, **
*Institute of Biophysics and Cell Engineering, National Academy of Sciences of Belarus,
ul. Akademicheskaya 27, Minsk, 220072 Belarus
**Faculty of Biology, Belarusian State University, ul. Kurchatova 10, Minsk, 220030 Belarus
Different metabolites are able to modulate function of presynaptic endings of neurons. We investigated the
influence of lactate and 3,5-dihydroxybenzoic acid, an agonist for the lactate receptor HCAR1, on the syn-
aptic vesicle cycle and the generation of reactive oxygen species in rat brain synaptosomes. Supplementation
of lactate (4 mM) versus pyruvate had no significant effect on the synaptic vesicle cycle. With the help of a
fluorescent dye acridine orange we demonstrated that 3,5-dihydroxybenzoic acid in the concentration of
500 μM is amenable for eliciting endocytosis thereby preventing its potential inhibition during this stimulus.
In the presence of lactate but not 3,5-dihydroxybenzoic acid there was an increase in 2′,7′-dichlorofluores-
cein diacetate fluorescence, indicating reactive oxygen species generation. Also, using a fluorescent dye Mi-
toSOX, we demonstrated the formation of superoxide anion in mitochondria after the addition of lactate.
Therefore, lactate can influence the function of presynaptic endings of neurons through stimulation of reac-
tive oxygen species generation and activation of endocytosis mediated by lactate receptor HCAR1.
Keywords: synaptosomes, lactate, synaptic vesicles, endocytosis, ROS, mitochondria
БИОФИЗИКА том 66
№ 4
2021