БИОФИЗИКА, 2021, том 66, № 4, с. 741-749
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 577.3;57.042
РЕГУЛЯЦИЯ ОКИСЛИТЕЛЬНЫХ ПРОЦЕССОВ В ОРГАНАХ МЫШЕЙ
ПРИ ДЕЙСТВИИ ХИМИЧЕСКИХ И ФИЗИЧЕСКИХ ФАКТОРОВ
В МАЛЫХ ДОЗАХ
© 2021 г. А.Г. Кудяшева*, Н.Г. Загорская*, О.В Раскоша*, Л.Н. Шишкина**
*Институт биологии Коми научного центра УрО РАН, 167982, Сыктывкар, ул. Коммунистическая, 28
**Институт биохимической физики им. Н.М. Эмануэля РАН, 119334, Москва, ул. Косыгина, 4
E-mail: shishkina@sky.chph.ras.ru
Поступила в редакцию 29.11.2019 г.
После доработки 13.04.2021 г.
Принята к публикации 20.04.2021 г.
Изучено влияние раздельного и совместного действия нитрата свинца в широком диапазоне доз,
уранилнитрата и хронического γ-излучения в малой дозе на формирование окислительного стресса
в печени мышей линии CBA (самцы) и морфофункциональное состояние щитовидной железы в
ранние и отдаленные сроки после окончания воздействий. Выявлен сложный и неоднозначный
характер изменений параметров системы регуляции окислительных процессов в печени мышей и
морфофизиологические перестройки в щитовидной железе. Масштаб и направленность этих
изменений зависели от возраста животных, химической природы токсикантов и их концентрации,
времени после окончания воздействий. Анализ литературы и полученные данные позволяют
сделать заключение о невозможности прогнозирования последствий обитания млекопитающих в
неблагоприятных экологических условиях путем экстраполяции результатов из области больших
доз к малым дозам полютантов.
Ключевые слова: регуляция, перекисное окисление липидов, нитрат свинца, уранилнитрат, хроническое
облучение, малые дозы.
DOI: 10.31857/S0006302921040141
зателями физико-химической системы регуля-
В настоящее время не вызывает сомнения, что
ции в тканях опытных животных [3].
воздействие любых повреждающих факторов на
биологические объекты вызывает формирование
Хозяйственная деятельность человека способ-
комплекса биохимических реакций, рассматри-
ствует появлению новых территорий, биота кото-
ваемых как окислительный стресс. Это обуслов-
рых подвергается хроническому воздействию
лено нарушением регуляции окислительных про-
факторов разной природы. Среди загрязнителей
цессов в тканях, стационарный уровень которых
распространены ионы свинца, уранилнитрат,
в норме поддерживается физико-химической си-
низкоинтенсивное ионизирующее излучение.
стемой регуляции, функционирующей как на
Поступление свинца в организм человека в тече-
ние длительного времени вызывает повреждение
мембранном, так и органном уровнях [1, 2]. Ана-
головного мозга и почек, оказывает токсическое
лиз взаимосвязей между различными показателя-
ми данной регуляторной системы позволил пред-
воздействие на развитие плода и процесс сперма-
тогенеза [4-6]. Природа токсичности соединений
ложить новый подход для оценки биологических
урана при поступлении в организм обусловлена
последствий воздействия повреждающих факто-
как радиоактивными свойствами изотопов урана,
ров разной природы и мощности по изменению
так и их химическим влиянием на обмен веществ
масштаба и направленности корреляций между
[7-9]. Установлено, что при длительном обита-
тесно взаимосвязанными в норме разными пока-
нии природных популяций мышевидных грызу-
Сокращения: ПОЛ
- перекисное окисление липидов,
нов на территориях с повышенным радиацион-
ПДК - предельно допустимая концентрация, ФЛ - фос-
ным фоном в Республике Коми или загрязнен-
фолипиды, ФХ - фосфатидилхолин, ФЭ - фосфатидил- ных радионуклидами в зоне аварии на
этаноламин, ФИ - фосфатидилинозит, ФС - фосфати-
Чернобыльской АЭС процессы перекисного
дилсерин, КЛ - кардиолипин, ФК - фосфатидная кисло-
та, ЛФХ
- лизоформы фосфолипидов, СМ
-
окисления липидов (ПОЛ) участвуют в их адапта-
сфингомиелин.
ции к изменившимся условиям окружающей сре-
741
742
КУДЯШЕВА и др.
ды за счет перехода системы регуляции на новый
Мыши групп А и Б пили чистую воду, осталь-
уровень функционирования [10, 11].
ные группы мышей с питьевой водой получали
Эффективность действия стресс-факторов мо-
растворы нитрата свинца (квалификации «ч.д.а.»)
в расчетных концентрациях, обеспечивающих
жет быть связана и с изменением состояния щи-
товидной железы, так как синтезируемые ее фол-
поступление в организм ионов Pb2+ в указанных
ликулярными клетками тиреоидные гормоны
выше концентрациях за весь период эксперимен-
оказывают влияние практически на все виды об-
та. Предельно допустимая концентрация (ПДК)
мена веществ, усиливают окислительные процес-
ионов свинца в питьевой воде для водных объек-
сы в организме, стимулируют теплообразование,
тов бытового пользования составляет 0.03 мг/л,
пролиферацию и дифференцировку клеток [12].
следовательно, общее потребление токсиканта
Изучению последствий сочетанного действия
превышает ПДК в 2-200 раз. При этом поступле-
факторов разной природы на организм млекопи-
ние в организм нитрат-иона за месяц даже при
тающих уделяется значительно меньше внимания
концентрации нитрата свинца 0.3 мг/кг в 75 раз
по сравнению с действием отдельных факторов
ниже его ПДК.
[13]. Способность токсикантов перемещаться на
В экспериментах с уранилнитратом мыши
значительные расстояния от источника загрязне-
(возраст пять месяцев) были разделены на четыре
ния и непредсказуемость их совместного дей-
группы по восемь-десять особей в каждой:
ствия требуют более детального исследования ме-
группа 1 -контроль;
ханизма формирования ранних и отдаленных эф-
группа 2 - облучение;
фектов
при хроническом поступлении
группа 3 - уранилнитрат (UO2(NO3)2 · 6H2O);
токсикантов в организм в течение длительного
времени.
группа 4 - облучение + уранилнитрат;
Целью работы являлось изучение ранних и от-
Удельная активность уранилнитрата составля-
даленных биофизических и биохимических эф-
ла 25 Бк/л (0.00428 г/л), что соответствует содер-
фектов в тканях мышей линии CBA, подвергав-
жанию солей урана в природных водах на терри-
шихся хроническому низкоинтенсивному γ-из-
тории с повышенным уровнем естественной ра-
лучению в малой дозе в сочетании с нитратом
диоактивности в Республике Коми [10]. Каждое
свинца в широком диапазоне доз или уранилнит-
животное за 30 суток получало вместе с водой в
рата.
среднем 0.18 мг уранилнитрата.
γ-Облучение мышей обеспечивали два источни-
МАТЕРИАЛЫ И МЕТОДЫ
ка 226Ra с активностью 0.474 · 106 и 0.451 · 106 кБк,
разнесенные на расстоянии 2.5 м. Геометрия их рас-
Объектами исследования были половозрелые
положения обеспечивала сравнительно однородное
мыши линии СВА (самцы), обычно используе-
облучение всех животных (неравномерность облу-
мые при оценке токсических свойств разных со-
чения не превышала 6%). Данный уровень излуче-
единений. Животные были получены из научной
ния имитировал условия низкоинтенсивного внеш-
коллекции экспериментальных животных ИБ
него γ-излучения на участках с повышенным фоном
естественной радиоактивности в Республике Коми
rf.ru/usu/471933/). В течение одного месяца мы-
[10]. Суммарную поглощенную дозу (1.6 сГр) опре-
шей ежедневно подвергали воздействию хрони-
деляли термолюминесцентными дозиметрами с де-
ческого γ-излучения с мощностью дозы 2 мР/ч,
текторами ДТГ-4 (LiF) с последующей регистраци-
одновременно с этим животные потребляли с пи-
ей на термолюминесцентной установке ДВГ-02ТМ
тьевой водой токсиканты. Были проведены две
(НПП «Доза», Россия).
серии экспериментов.
Забой мышей одновременно опытных и кон-
В первой серии с нитратом свинца мыши (воз-
трольных групп осуществляли в первые сутки
раст два с половиной-три месяца) были разделе-
(ранний период) и спустя 30 суток (отдаленный
ны на семь групп по семь-десять особей в каждой:
период) после окончания воздействия при со-
А - контроль;
блюдении принципов биоэтики и гуманного об-
Б - облучение;
ращения с животными [14] в утренние часы, что-
бы избежать влияния суточных колебаний на ис-
I
- облучение
+ нитрат свинца в дозе
следованные показатели. Необходимо также
0.003 г/кг;
отметить, что эксперименты проводили в весен-
II
- облучение
+ нитрат свинца в дозе
ний сезон (март - май)
0.01 г/кг;
Печень сразу после забоя помещали на лед.
III - облучение + нитрат свинца в дозе
Липиды из гомогената печени выделяли по мето-
0.03 г/кг;
ду Блая и Дайера в модификации Кейтса [15]. Ка-
IV - облучение + нитрат свинца в дозе 0.1 г/кг;
чественный и количественный состав фосфоли-
V - облучение + нитрат свинца в дозе 0.3 г/кг.
пидов (ФЛ) определяли методом тонкослойной
БИОФИЗИКА том 66
№ 4
2021
РЕГУЛЯЦИЯ ОКИСЛИТЕЛЬНЫХ ПРОЦЕССОВ В ОРГАНАХ МЫШЕЙ
743
Состав фосфолипидов печени мышей в опытах с нитратом свинца и уранилнитратом в контрольных группах и
спустя сутки после хронического γ-облучения в дозе 1.6 сГр.
Относительное содержание фракции (%P)
Фракции
Группа 1 (контроль).
Группа 2
Группа А (контроль).
Группа Б (облучение).
фосфолипидов
Возраст 6 месяцев,
(γ-облучение). Опыт
Возраст 3.5-4.0
Опыт с нитратом
n = 12
с уранилнитратом,
месяца, n = 25
свинца, n = 22
n = 20
ЛФХ
3.37 ± 0.24
1.65 ± 0.20***
3.90 ± 0.20
7.09 ± 0.31***
СМ
3.34 ± 0.05
2.15 ± 0.22***
4.43 ± 0.39
3.70 ± 0.19
ФХ
52.2 ± 0.9
57.3 ± 0.7***
47.1 ± 0.7
42.50 ± 0.65***
ФИ+ФС
9.57 ± 0.10
6.20 ± 0.17**
10.37 ± 0.50
10.85 ± 0.62
ФЭ
26.9 ± 1.0
28.70 ± 0.19
28.9 ± 0.6
28.84 ± 0.23
КЛ+ФК
4.58 ± 0.45
3.96 ± 0.42
5.30 ± 0.65
7.03 ± 0.59
Примечание. n - Количество хроматографических дорожек; здесь и далее (на рисунках): различия между кон-
тролем и вариантом эксперимента достоверны при * - р < 0.1, ** - p < 0.05 *** - p < 0.01.
хроматографии с использованием силикагеля ти-
Щитовидную железу сразу после извлечения
па G (Sigma, США) [16]. В качестве подвижной
помещали в 10%-й формалин с последующей
фазы использовали смесь «хлороформ-метанол-
стандартной гистологической обработкой. Па-
ледяная уксусная кислота-вода» в соотношении
рафиновые срезы тиреоидной ткани толщиной
50 : 30 : 8 : 4. Проявление хроматограмм проводи-
4-5 мкм окрашивали реактивом Шиффа с до-
ли в парах йода. Количественный анализ отдель-
краской ядер гематоксилином Карачи. На гисто-
ных фракций ФЛ проводили на спектрофотомет-
логических препаратах с использованием окуляр-
ре Спекол-211 (Carl Zeiss Jena, Германия) при
ной линейки измеряли высоту клеток фоллику-
длине волны 800 нм по образованию фосфорно-
лярного эпителия, а также длинный (L) и
молибденового комплекса в присутствии аскор-
короткий (В) поперечники фолликулов (от 50 до
биновой кислоты. Оценивали количественные
100 замеров). Средний диаметр фолликулов рас-
соотношения отдельных фракций ФЛ и обоб-
считывали по формуле D = (L + B)/2. По отноше-
щенные показатели состава липидов: процентное
нию среднего диаметра фолликула к высоте фол-
содержание ФЛ в составе общих липидов и отно-
ликулярного эпителия рассчитывали индекс
шение содержания основных фракций ФЛ клеток
функции железы [17]. Морфометрические пара-
млекопитающих фосфатидилхолина и фосфа-
метры анализировали при помощи программы
тидилэтаноламина (ФХ/ФЭ), отражающие
AxioVision LE под микроскопом AxioScope A1
структурное состояние мембранной системы ор-
(Carl Zeiss, Германия) при увеличении от 250× до
гана; соотношение сумм более легкоокисляемых
1250×, осуществляя их измерение в соответствии
и более трудноокисляемых фракций ФЛ
с имеющимися в литературе методическими ре-
(∑ЛОФЛ/∑ТОФЛ), характеризующее способ-
комендациями, в том числе разработанными для
ность липидов к окислению [2]. Последнее соот-
эндокринных органов [18, 19].
ношение вычисляли по формуле:
Результаты обрабатывали методами вариаци-
∑ЛОФЛ/∑ТОФЛ = (ФИ + ФС + ФЭ + КЛ +
онной статистики [20], используя Microsoft Office
+ ФК)/(ЛФХ + СМ + ФХ),
Excel 2007, программу Statistica 6.0. и пакет про-
грамм KINS [21]. Для оценки результатов стати-
где ФИ - фосфатидилинозит, ФС - фосфатидил-
стического анализа использовали стандартные
серин, КЛ - кардиолипин, ФК - фосфатидная
кислота, ЛФХ
- лизоформы фосфолипидов,
параметры: R - коэффициент корреляции, b -
СМ - сфингомиелин. Для определения состава
коэффициент линейной регрессии, p - уровень
ФЛ для каждой пробы анализировали по четыре-
достоверности различий (вероятность ошибки) и
шесть параллельных дорожек. Все измерения
критерий Стьюдента. В таблице и на рисунках ре-
проводили для каждого животного индивидуаль-
зультаты представлены в виде среднеарифмети-
но. Общее количество мышей в экспериментах
ческих значений с указанием их среднеквадра-
составило 174 особи.
тичных ошибок (M ± m).
БИОФИЗИКА том 66
№ 4
2021
744
КУДЯШЕВА и др.
Рис. 1. Высота фолликулярного эпителия (% от контроля) в щитовидной железе мышей линии CBA (самцы) в ранние
(а) и отдаленные (б) сроки после воздействия γ-излучения и уранилнитрата: 1 - хроническое γ-облучение в
суммарной дозе 1.6 сГр; 2 - потребление с питьевой водой уранилнитрата в течение 30 суток; 3 - совместное действие
обоих факторов.
РЕЗУЛЬТАТЫ
контрольной величины (рис. 1). При этом в отда-
ленный период во всех вариантах эксперимента
Воздействие на организм любых неблагопри-
наблюдается нормализация данного параметра.
ятных факторов вызывает существенные измене-
Как показано в работах [18, 26], высота фоллику-
ния во всех его системах. Так, в работе [22] были
лярного эпителия является информативным по-
выявлены достоверные изменения состояния
казателем, отражающим функциональное состо-
процессов ПОЛ в головном мозге и крови мышей
яние щитовидной железы. Достоверные различия
при совместном действии низкоинтенсивного
этого показателя от контрольного значения во
излучения в малой дозе и наличии нитрата свин-
всех опытных группах в ранние сроки после
ца в питьевой воде. Так как интенсивность про-
окончания воздействий (рис. 1) свидетельствуют
цессов ПОЛ обусловливает влияние физико-хи-
о существенных изменениях индекса функции
мических свойств липидов на координацию взаи-
щитовидной железы.
мосвязей между различными параметрами
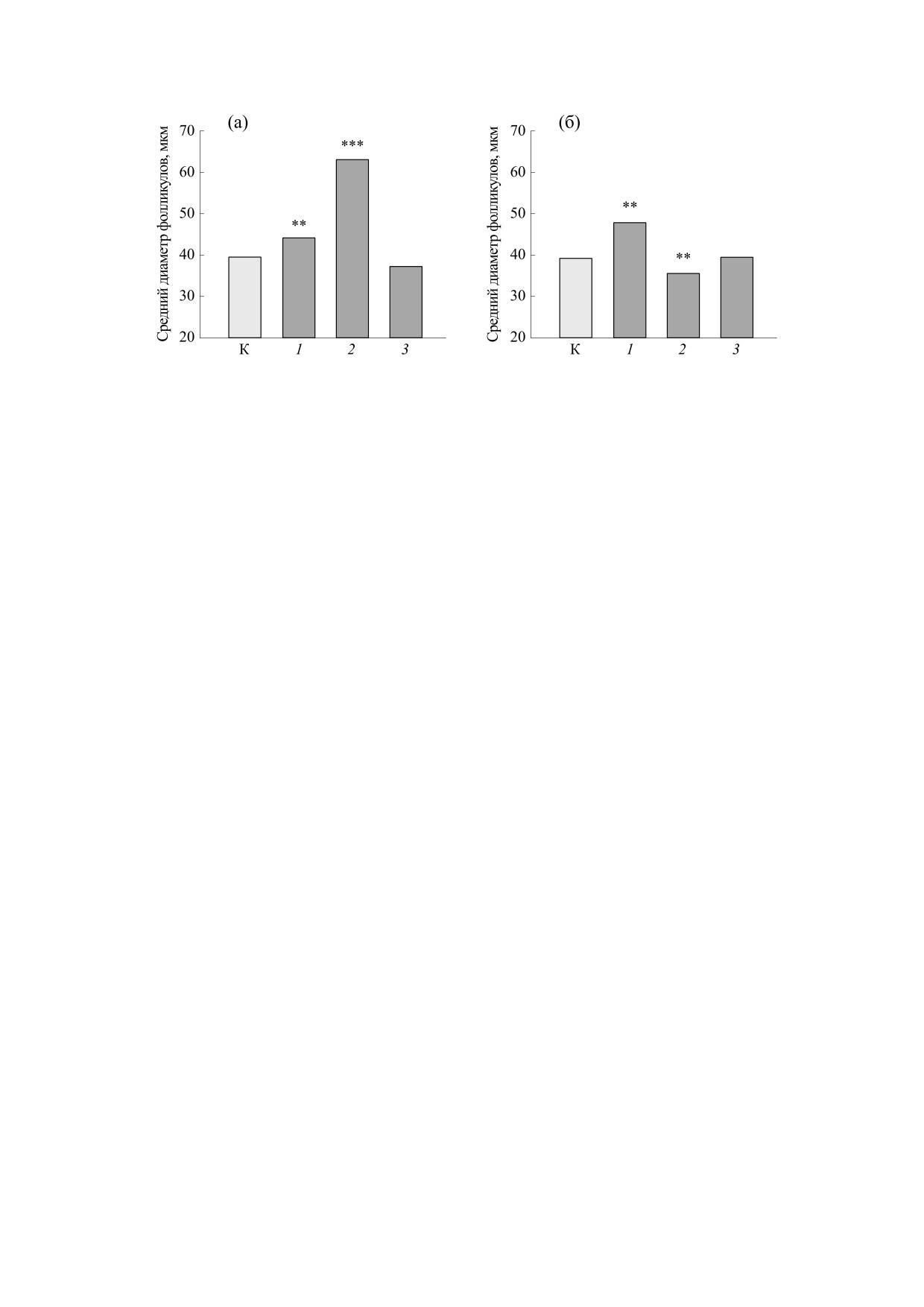
Измерение диаметров фолликулов в тиреоид-
системы регуляции и баланс биохимических
ной паренхиме показало, что у облученных жи-
функций в биологических объектах [23, 24], то
вотных средние значения этого показателя были
необходимо отметить, что липиды головного
выше нормы как в ранние, так и в отдаленные
мозга и эритроцитов крови, в отличие от липидов
сроки после радиационного воздействия (рис. 2).
печени, обладают низким антиоксидантным ста-
При этом в ранние сроки после потребления
тусом [25].
только уранилнитрата средний диаметр фоллику-
лов достоверно (р ≤ 0.001) увеличивается относи-
Поскольку синтезируемые клетками щитовид-
ной железы тиреоидные гормоны усиливают
тельно контрольных значений, а спустя месяц по-
сле прекращения его потребления - уменьшается
окислительные процессы в организме, то обнару-
(рис. 2). В щитовидной железе мышей, которые
женное изменение морфофункциональных пара-
испытывали одновременно воздействие обоих
метров тиреоидной ткани мышей в ответ на дей-
факторов, средний диаметр фолликулов и индекс
ствие хронического γ-излучения и уранилнитрата
также свидетельствует об участии щитовидной
функции были в пределах контрольных значений
независимо от времени, прошедшего после окон-
железы мышей в формировании последствий
чания воздействия факторов.
действия исследуемых факторов. Результаты,
представленные на рис. 1 и 2, позволяют заклю-
Выбор печени как основного органа для изуче-
чить, что выраженность и направленность состо-
ния механизма формирования последствий сов-
яния щитовидной железы зависели от времени,
местного действия повреждающих факторов раз-
прошедшего после воздействия. Так, в ранний
ной природы на регуляцию процессов ПОЛ не
период после окончания облучения животных
случаен. Именно печень является одним из ос-
и/или наличия в питьевой воде уранилнитрата в
новных органов как биосинтеза липидов, так и
тиреоидной паренхиме обнаружено статистиче-
биотрансформации поступающих в организм
ски значимое увеличение высоты фолликулярно-
ксенобиотиков. Известно, что показатели систе-
го эпителия, в то время как при их совместном
мы регуляции ПОЛ обладают выраженной суточ-
действии значения этого показателя были ниже
ной и сезонной вариабельностью [23, 27]. Чтобы
БИОФИЗИКА том 66
№ 4
2021
РЕГУЛЯЦИЯ ОКИСЛИТЕЛЬНЫХ ПРОЦЕССОВ В ОРГАНАХ МЫШЕЙ
745
Рис. 2. Средний диаметр фолликулов щитовидной железы мышей линии СВА (самцы) в ранние (а) и отдаленные (б)
сроки после воздействия γ-облучения и уранилнитрата: К - контроль; 1 - хроническое γ-облучение в суммарной дозе
1.6 сГр; 2 - потребление с питьевой водой уранилнитрата в течение 30 суток; 3 - совместное действие обоих факторов.
минимизировать влияние сезонных колебаний
структурным состоянием мембранной системы
исходного состояния параметров системы регу-
печени в 2.65 раза выше в группе более молодых
ляции ПОЛ, обе серии экспериментов проводили
мышей: b = 0.53 ± 0.11 и 0.20 ± 0.05 соответственно.
в весенний период. Для адекватной оценки меха-
Различия исходных величин параметров си-
низма формирования последствий воздействий
стемы регуляции ПОЛ вызывает и неодинаковый
токсикантов разной природы прежде всего было
характер изменения в соотношении фракций ФЛ
необходимо сопоставить ответ процессов ПОЛ в
в ранние сроки после окончания хронического
печени мышей разного возраста на воздействие
облучения. Так, в группе более молодых мышей
хронического ионизирующего излучения в малой
облучение приводит к уменьшению доли ФХ и
дозе. Это обусловлено высокой чувствительно-
росту в 1.8 раза доли лизоформ ФЛ, в то время как
стью окислительных процессов к низкоинтен-
в группе мышей старшего возраста доля ФХ в со-
сивному облучению млекопитающих в малых до-
ставе ФЛ печени достоверно увеличивается при
зах, которая была выявлена в лабораторных экс-
снижении доли лизоформ ФЛ в два раза. Необхо-
периментах [28] и в исследованиях на природных
димо обратить внимание на практически посто-
популяциях грызунов [10, 11]. Состав ФЛ печени
янное относительное содержание и у мышей раз-
интактных мышей CBA в контрольных группах и
ных возрастных групп, и в ранние сроки после
в ранние сроки после окончания одного облуче-
хронического облучения в малой дозе второй ос-
ния в обоих вариантах опытов представлен в таб-
новной фракции ФЛ печени - ФЭ, который ло-
лице.
кализуется преимущественно на внутренней сто-
роне мембран [29].
Анализ полученных данных свидетельствует о
том, что мембранная система печени более моло-
Сохранение функционирования системы ре-
дых особей характеризуется достоверно менее
гуляции ПОЛ в группах животных, подвергнутых
жесткой структурированностью (ФХ/ФЭ
=
различным воздействиям, оценивают, сопостав-
= 1.629 ± 0.019) по сравнению с аналогичным по-
ляя масштаб и характер обратной корреляцион-
казателем для более старых особей (ФХ/ФЭ =
ной взаимосвязи между соотношением способ-
= 1.940 ± 0.035, p < 0.01). При этом липиды
ности липидов к окислению (∑ЛОФЛ/∑ТОФЛ) и
печени являются более окисляемыми (∑ЛОФЛ/
структурным состоянием мембранной системы
∑ТОФЛ = 0.877 ± 0.015) в опыте с нитратом свин-
органа (ФХ/ФЭ) в группах контрольных и опыт-
ца по сравнению со значением 0.697 ± 0.008 в
ных животных [2]. Данная взаимосвязь выявлена
опыте с уранилнитратом (p < 0.01). Это обуслов-
в обеих контрольных группах мышей с коэффи-
лено более высоким содержанием ФЛ в составе
циентами корреляции R = -0.87 ± 0.10 (n = 6) и
общих липидов в печени более молодых мышей
R = -0.83 ± 0.11 (n = 8) в экспериментах с нитра-
(50.4 ± 1.4%) по сравнению с их долей в составе
том свинца и уранилнитратом соответственно.
общих липидов в возрастной группе мышей
Следовательно, регуляция процессов ПОЛ в
(37.7 ± 1.4%), поскольку именно ФЛ являются ос-
группах мышей линии CBA разного возраста от-
новными субстратами окисления в биологиче-
носительно стабильна. Однако в ранние сроки
ских мембранах. При этом закономерно, что ко-
после окончания радиационного воздействия бо-
эффициент линейной регрессии обратной взаи-
лее существенные нарушения регуляции
мосвязи между окисляемостью липидов и
ПОЛ наблюдаются в экспериментах с нитратом
БИОФИЗИКА том 66
№ 4
2021
746
КУДЯШЕВА и др.
Рис. 3. Отношение основных фракций (ФХ/ФЭ) и соотношение сумм более легко- и трудноокисляемых
фосфолипидов в печени мышей линии CBA (самцы) спустя 30 суток после окончания воздействия γ-излучения и
нитрата свинца: А - возрастной контроль; Б - хроническое γ-облучение в суммарной дозе 1.6 сГр; совместное
действие облучения и нитрата свинца в дозах 0.003 г/кг (I), 0.01 г/кг (II), 0.03 г/кг (III), 0.1 г/кг (IV) и 0.3 г/кг (V).
свинца (аналогичная корреляции отсутствует,
става ФЛ в печени мышей выявлены спустя 30 су-
R = -0.54), в то время как для более возрастной
ток после окончания совместного действия фак-
группы мышей в экспериментах с уранилнитра-
торов в экспериментах со свинцом, что следует из
том обнаружено только уменьшение взаимосвя-
данных, представленных на рис. 3 и 4. Спустя ме-
занности между данными показателями
сяц после прекращения воздействий наличие в
(R = -0.75 ± 0.19) относительно контроля. Тем не
питьевой воде мышей нитрата свинца при одно-
менее ответ процессов ПОЛ в печени мышей раз-
временном действии облучения вызывает досто-
ных возрастных групп на хроническое облучение
верное уменьшение окисляемости липидов пече-
в малой дозе имеет однотипный характер.
ни по сравнению с аналогичным показателем в
Как известно, среди органов интактных лабо-
контроле при всех исследованных дозах токси-
раторных грызунов печень характеризуется са-
канта. Отношение же ФХ/ФЭ возрастает при до-
мой низкой интенсивностью процессов ПОЛ и
зах нитрата свинца 0.003 и 0.1 г/кг, снижается при
наиболее высокой обеспеченностью липидов ан-
0.03 и 0.3 г/кг при сохранении приблизительно на
тиоксидантами [25, 28]. Это приводит к тому, что
уровне контрольных значений при дозе 0.01 г/кг
нарушения взаимосвязей между каталазной ак-
(рис. 3).
тивностью печени и интенсивностью процессов
ПОЛ в ранние сроки и после раздельного дей-
Среди продуктов деградации ФЛ важную роль
ствия облучения, и при его совместном действии
играют лизоформы ФЛ, обладающие детергент-
с нитратом свинца не столь значительны [22].
ными свойствами [29]. В отдаленные сроки после
В отдаленные сроки после хронического облу-
окончания воздействий достоверный рост отно-
чения в малой дозе незначительные изменения в
сительного содержания лизоформ ФЛ наблюда-
относительном содержании некоторых минор-
ется как после хронического облучения, так и при
ных фракций ФЛ выявлены в обоих эксперимен-
совместном действии облучения и нитрата свин-
тах, что обусловливает поддержание обобщенных
ца, кроме мышей группы IV, получивших токси-
показателей состава ФЛ на близких к контроль-
кант в дозе 0.1 г/кг (рис. 4).
ным значениям величинах. В серии эксперимен-
та с уранилнитратом самые существенные изме-
В группах мышей более старшего возраста
нения в составе ФЛ печени спустя месяц после
(эксперименты с уранилнитратом) хроническое
окончания воздействий выявлены только при на-
облучение в малой дозе вызывает достоверный
личии в питьевой воде мышей уранилнитрата
рост отношения ФХ/ФЭ и доли лизоформ в ФЛ
(группа 3). Интересно отметить, что достоверные
печени, а также уменьшение способности липи-
различия в составе ФЛ в этой группе мышей об-
наружены именно для более легкоокисляемых
дов к окислению в отдаленные сроки после воз-
ФЛ: уменьшение доли ФЭ на 5.7% (p < 0.01) и
действия (рис. 5 и 6). Спустя 30 суток после окон-
снижение в 1.9 раза доли суммарной фракции
чания воздействий выявлено уменьшение отно-
ФИ + ФС при росте относительного содержания
шения ФХ/ФЭ после совместное действие
КЛ + ФК в 2.5 раза.
низкоинтенсивного облучения и уранилнитрата
Существенные различия как в соотношении
(рис. 5) и доли лизоформ ФЛ при потреблении
фракций ФЛ, так и обобщенных показателей со-
мышами только уранилнитрата (рис. 6).
БИОФИЗИКА том 66
№ 4
2021
РЕГУЛЯЦИЯ ОКИСЛИТЕЛЬНЫХ ПРОЦЕССОВ В ОРГАНАХ МЫШЕЙ
747
Рис. 5. Отношение основных фракций (ФХ/ФЭ) и со-
Рис. 4. Доля лизоформ в составе фосфолипидов в пе-
отношение сумм более легко- и более трудноокисля-
чени мышей линии CBA (самцы) спустя 30 суток по-
емых фосфолипидов в печени мышей линии CBA
сле окончания воздействия γ-излучения и нитрата
(самцы) спустя 30 суток после окончания воздей-
свинца: А - возрастной контроль; Б - хроническое
ствия γ-излучения и уранилнитрата: К - контроль;
γ-облучение в суммарной дозе 1.6 сГр; совместное
1 - потребление с питьевой водой уранилнитрата в
действие облучения и нитрата свинца в дозах
течение 30 суток; 2 -хроническое γ-облучение в сум-
0.003 г/кг (I), 0.01 г/кг (II), 0.03 г/кг (III), 0.1 г/кг (IV)
марной дозе 1.6 сГр; 3 - совместное действие обоих
и 0.3 г/кг (V).
факторов.
ОБСУЖДЕНИЕ
ные сроки после изученных воздействий измене-
ния показателей состава ФЛ в печени мышей
Анализ представленных выше данных свиде-
более выражены при раздельном действии облу-
тельствует о том, что исходное состояние пара-
чения или уранилнитрата (рис. 5 и 6), в то время
метров системы регуляции ПОЛ оказывает суще-
как при их совместном действии достоверные из-
ственное влияние на механизм формирования
менения обнаружены лишь для отношения ос-
последствий воздействия хронического γ-излуче-
новных фракций ФЛ (рис. 5). Это может быть
ния в малой дозе. Так, более существенные изме-
обусловлено как разной чувствительностью пока-
нения структурного состояния мембранной си-
зателей и их нормализацией в зависимости от
стемы печени и способности липидов к окисле-
природы повреждающего воздействия, так и не-
нию в ранние сроки после облучения выявлены в
одинаковой способностью хронического излуче-
печени более молодых мышей по сравнению с
ния в малых дозах и уранилнитрата влиять на
животными старшей возрастной группы.
процессы гемодинамики и микроциркуляции в
В печени в отдаленные сроки после совмест-
ного действия облучения и нитрата свинца выяв-
лен нелинейный (полимодальный) характер
изменения доли лизоформ ФЛ (рис. 4) и обоб-
щенных показателей состава ФЛ (рис. 3) в зави-
симости от концентрации нитрата свинца: мак-
симальный эффект проявляется при мини-
мальной из исследованных доз токсиканта
(0.003 г/кг). Ранее нелинейный характер анало-
гичных показателей состава ФЛ был обнаружен в
головном мозге и эритроцитах крови мышей в
ранние сроки после совместного действия дан-
ных факторов [22].
Формирование последствий зависит и от при-
роды токсиканта. Так, структурно-функциональ-
ные изменения тиреоидной ткани в ранние сроки
после окончания воздействия более выражены в
Рис. 6. Доля лизоформ в составе фосфолипидов
группах мышей при раздельном действии изучен-
печени мышей линии СВА (самцы) спустя 30 суток
ных факторов (рис. 1 и 2). Совместное действие
после окончания воздействия γ-излучения и
уранилнитрата: К - контроль; 1 - потребление с
облучения и уранилнитрата вызвало достоверное
питьевой водой уранилнитрата в течение 30 суток; 2 -
изменения только одного из изученных показате-
хроническое γ-облучение в суммарной дозе 1.6 сГр;
лей (рис. 1). Необходимо отметить, что в отдален-
3 - совместное действие обоих факторов.
БИОФИЗИКА том 66
№ 4
2021
748
КУДЯШЕВА и др.
тканях, взаимосвязанных с процессами инакти-
КОНФЛИКТ ИНТЕРЕСОВ
вации токсикантов в печени и их экскреции с
Авторы заявляют об отсутствии конфликта
мочой.
интересов.
Существенные изменения количественных
соотношений среди минорных фракций ФЛ при
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
действии хронического облучения и химических
Все применимые международные, националь-
токсикантов могут оказывать влияние и на их
ные и институциональные принципы ухода и ис-
функционирование как сигнальных молекул, по-
пользования животных при выполнении работы
скольку наличие сигнальных функций у минор-
были соблюдены.
ных фракций ФЛ и продуктов их окисления пока-
зано во многих исследованиях [30-34].
СПИСОК ЛИТЕРАТУРЫ
1.
Ye. B. Burlakova, N. P. Pal’mina, and Ye. L. Mal’tseva,
ЗАКЛЮЧЕНИЕ
Membrane Lipid Oxidation (CRC Press, 1991).
2.
Л. Н. Шишкина, Е. В. Кушнирева и М. А. Смотря-
Таким образом, действие хронического низко-
ева, Радиац. биология. Радиоэкология 44 (3), 289
интенсивного излучения в малых дозах и химиче-
(2004).
ских токсикантов разной природы приводит к
3.
Л. Н. Шишкина, М. А. Климович и М. В. Козлов,
возникновению нарушений в органах лаборатор-
Биофизика 59 (2), 380 (2014).
ных животных, что характерно для формирова-
4.
Ф. С. Дзугкоева, Р. Л. Беликова и С. Г. Дзугкоев,
Нефрология и диализ 5 (3), 18 (2003).
ния окислительного стресса, обусловленного
5.
S.E. Martin, W. Griswold, in Environmental Science
нарушениями в физико-химической системе
and Technology Briefs for Citizens (Kansas State Univer-
регуляции ПОЛ. Масштаб и направленность из-
sity, USA, 2009), No 15, pp. 1-6.
менений показателей морфофункционального
6.
A. Agrawal, Adv. Life Sci. 2 (2), 29 (2012).
состояния щитовидной железы и параметров си-
7.
Ю. И. Москалев, Радиобиология инкорпорированных
стемы регуляции ПОЛ в печени мышей линии
радионуклидов (Энергоатомиздат, М., 1989).
CBA зависят от времени после окончания воздей-
8.
M. D. Sztajnkrycer and E. J. Otten, Mil. Med. 169 (3),
ствия, природы токсиканта и его концентрации в
212 (2004).
питьевой воде, возраста мышей. Выявление более
9.
S. V. Gudkov, A. V. Chernikov, and V. I. Bruskov, Rus.
выраженных изменений морфофизиологических
J. Gen. Chem. 86 (6), 1531 (2016).
параметров в щитовидной железе в ранние сроки
10.
A. G. Kudyasheva, L. N. Shishkina, O. G. Shevchenko,
et al., J. Environ. Radioactivity 97, 39 (2007).
после воздействий облучения и уранилнитрата
свидетельствует о высокой чувствительности ее
11.
L. N. Shishkina, A. G. Kudyasheva, and N. G. Zag-
orskaya, The Lessons of Chernobyl: 25 Years Later (Nova
функционального состояния к неблагоприятным
Science Publ., New York, 2012).
экологическим факторам.
12.
Л. К. Жункейра и Ж. Карнейро, Гистология: атлас:
учеб. пособие (ГЭОТАР-Медиа, М., 2009).
Сложный и неоднозначный характер ответных
13.
В. Г. Петин и Б. И. Сынзыныс, Комбинированное
реакций органов млекопитающих на воздействие
воздействие факторов окружающей среды на биоло-
факторов разной природы в малых дозах, нели-
гические системы (ИА-ТЭ, Обнинск, 1998).
нейный характер изменения структурного состо-
14.
О. В. Раскоша и А. И. Кичигин, Вестн. Института
яния мембранной системы тканей и способности
биологии Коми НЦ УрО РАН, № 3, 19 (2016).
их липидов к окислению, зависимость эффекта
15.
М. Кейтс, Техника липидологии (Мир, М., 1975).
от исходного состояния параметров системы ре-
16.
Биологические мембраны: Методы (Мир, М., 1990).
гуляции ПОЛ свидетельствуют о невозможности
17.
B. Л. Быков, Частная гистология человека (крат-
прогнозирования последствий обитания млеко-
кий обзорный курс) (СОТИС, М., 2002).
питающих в неблагоприятных экологических
18.
Г. Г. Автандилов, Медицинская морфометрия (Ме-
условиях путем экстраполяции результатов из об-
дицина, М., 1990).
ласти высоких доз к малым дозам поллютантов.
19.
O. K. Хмельницкий, Цитологическая и гистологи-
ческая диагностика заболеваний щитовидной желе-
зы (СОТИС, М., 2002).
20. Г. Ф. Лакин, Биометрия (Высш. шк., М., 1990).
ФИНАНСИРОВАНИЕ РАБОТЫ
21. Э. Ф. Брин и С. О. Травин, Хим. физика 10 (6), 830
(1991).
Работа выполнялась в рамках государственных
22. L. N. Shishkina, A. G. Kudyasheva, O. G. Shevchenko,
заданий Института Биологии ФИЦ Коми НЦ
et al., Heavy Metals and Other Pollutants in the Environ-
УрО РАН (№ гос. регистрации АААА-А18-
ment: Biological Aspects (Apple Acad. Press, Toronto,
118011190102-7) и Института биохимической фи-
2017).
зики им. Н.М. Эмануэля РАН (№ 44.4, гос. № те-
23. М. В. Козлов, В. В. Урнышева, Е. В. Кушнирева
мы: 0084-2019-0014).
и др., Биофизика 52 (4), 693 (2007).
БИОФИЗИКА том 66
№ 4
2021
РЕГУЛЯЦИЯ ОКИСЛИТЕЛЬНЫХ ПРОЦЕССОВ В ОРГАНАХ МЫШЕЙ
749
24. Н. В. Хрустова, М. В. Козлов и Л. Н. Шишкина,
29. Р. Геннис, Биомембраны: Молекулярная структура
Биофизика 56 (4), 668 (2011).
и функции (Мир, М., 1997).
25. Л. Н. Шишкина и Н. В. Хрустова, Биофизика 51
30. G. Leonarduzzi, M. C. Arkan, H. Başağa, et al., Free
(2), 340 (2006).
Radic. Biol. Med. 28 (9), 1370 (2000).
26. О. В. Раскоша и О. В. Ермакова, Теоретическая и
31. V. E. Kagan, G. G. Boriscnko, Y. Y. Tyirina, et al., Free
прикладная экология, № 2, 55 (2013).
Radic. Biol. Med. 37 (12), 11 (2004).
27. L. N. Shishkina and E. B. Burlakova, in Chemical and
32. X. Wang, S. P. Devaiah, and W. Zhang, Prog. Lipid
Biochemical Kinetics. New Hotizons. Vol II: Biological
Res. 45, 250 (2006).
Kinetics, Ed. by E. B. Burlakova and S. D. Varfolomeev
(VSP, Leden, Boston, 2005), pp. 334-364.
33. Т. И. Торховская, О. М. Ипатова, Т. С. Захарова
и др., Биохимия 72 (2), 149 (2007).
28. М. А. Климович, К. Ф. Сергейчев, Д. М. Карфи-
дов, и др., Технологии живых систем 7 (8), 17
34. J. P. Slotte and B. Ramstedt, Eur. J. Lipid Sci. Technol.
(2010).
109, 977 (2007).
Regulation of the Oxidative Processes in Organs of Mice under Action
of the Chemical and Physical Factors at Low Doses
A.G. Kudyasheva*, N.G. Zagorskaya*, O.V. Raskosha*, and L.N. Shishkina**
*Institute of Biology, Komi Science Centre, Ural Branch of the Russian Academy of Sciences,
Kommunisticheskaya ul. 28, Syktyvkar, Republic of Komi, 167982 Russia
**Emanuel Institute of Biochemical Physics, Russian Academy of Sciences, ul. Kosygina 4, Moscow, 119334 Russia
Influence of the separate and combined action of the lead nitrate at the wild range of doses, uranyl nitrate and
the chronic γ-irradiation at the low dose on the development of the oxidative stress in organ of mice CBA
(males) and morphophysiological state of thyroid gland was studied at the early and remote periods after ac-
tion. Complex and significant different character of changes of parameters of the oxidation process regulation
in liver of mace (males) and the morphological parameters in its thyroid gland is revealed. The scale and di-
rection of these changes depend on the age of animals, the nature of toxicants and their concentrations, and
also period after actions. Analysis of literature and the data obtained allow us to make it a conclusion that the
development of consequences for animals living under unfavorable ecological conditions may not predict us-
ing the extrapolation of data which were obtained at large doses to sphere of low doses.
Keywords: regulation, lipid peroxidation, lead nitrate, uranyl nitrate, chronic irradiation, low doses
БИОФИЗИКА том 66
№ 4
2021